Analysis of Gene Expression Signatures for Osteogenic 3D Perfusion-Bioreactor Cell Cultures Based on a Multifactorial DoE Approach


Abstract
:1. Introduction
2. Experimental Section
2.1. Ti6Al4V Scaffolds
2.2. hPDC Culture
2.3. qPCR
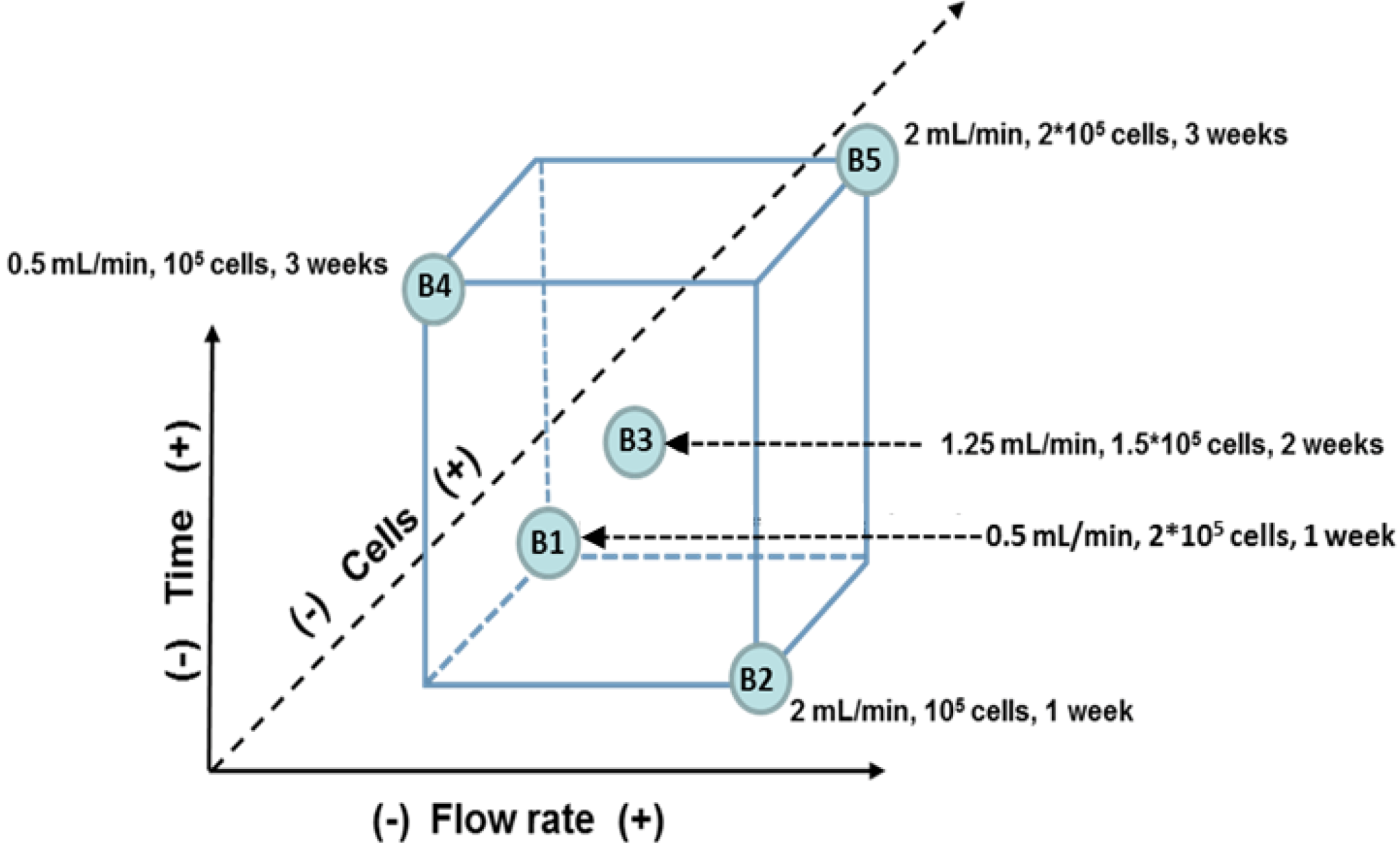
2.4. Multi-Factorial Analysis

2.5. DNA Content Analysis
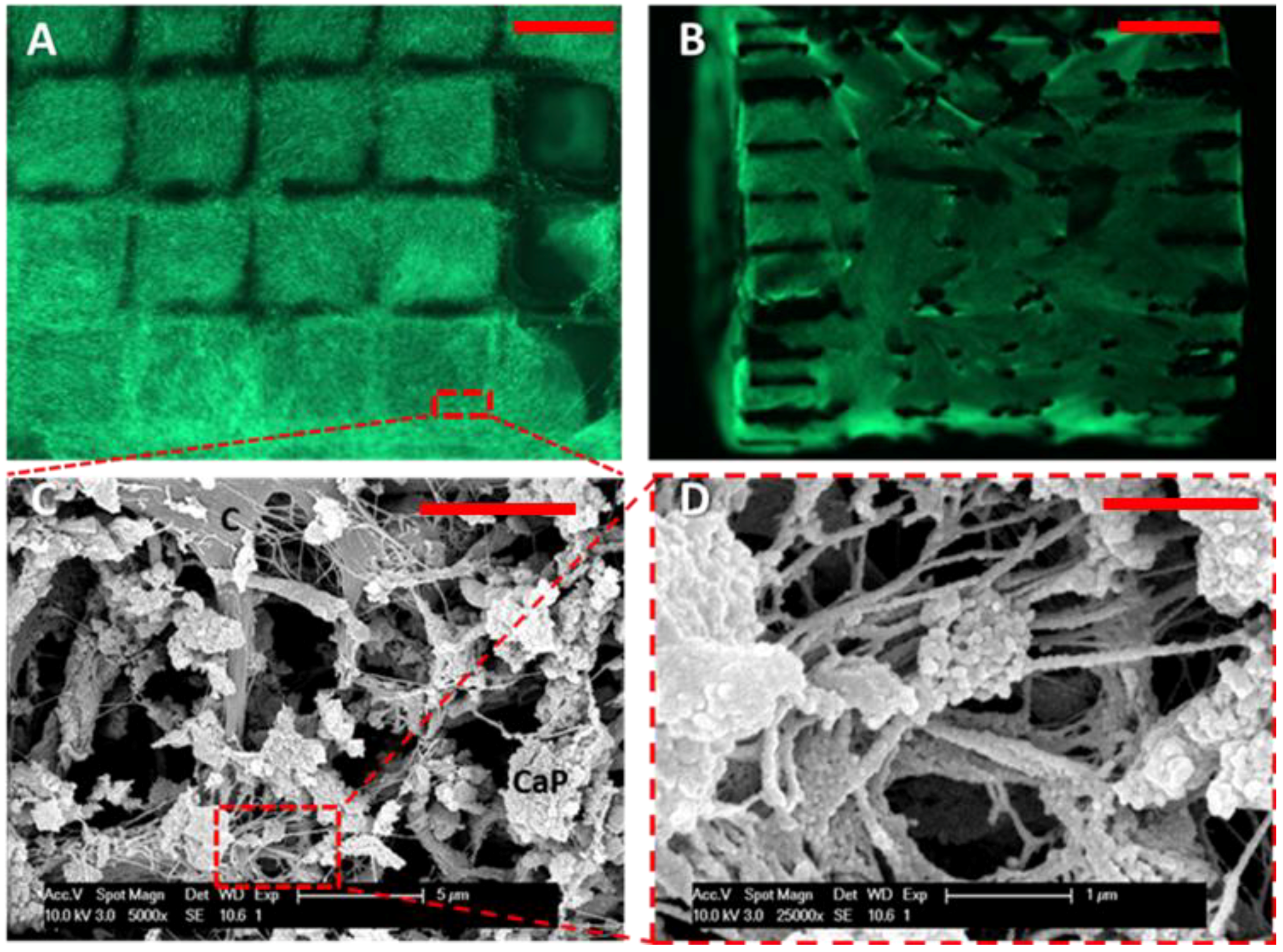
2.6. Live/Dead Assay
2.7. Scanning Electron Microscopy Analysis
2.8. Picrosirius Red Collagen Assay
2.9. Alizarin Red Mineralization Assay
3. Results and Discussion
3.1. Cell Proliferation



| Name | Experimental conditions | DNA content (ng/mL) (average, n = 3) |
| B1 | 0.5 mL/min, 2 × 105 cells, 7 days | 10,000 |
| B2 | 2 mL/min, 105 cells, 7 days | 7,600 |
| B3 | 1.25 mL/min, 1.5 × 105 cells, 14 days | 14,000 |
| B4 | 0.5 mL/min, 105 cells, 21 days | 34,000 |
| B5 | 2 mL/min, 2 × 105 cells, 21 days | 28,200 |
| (B) | ||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
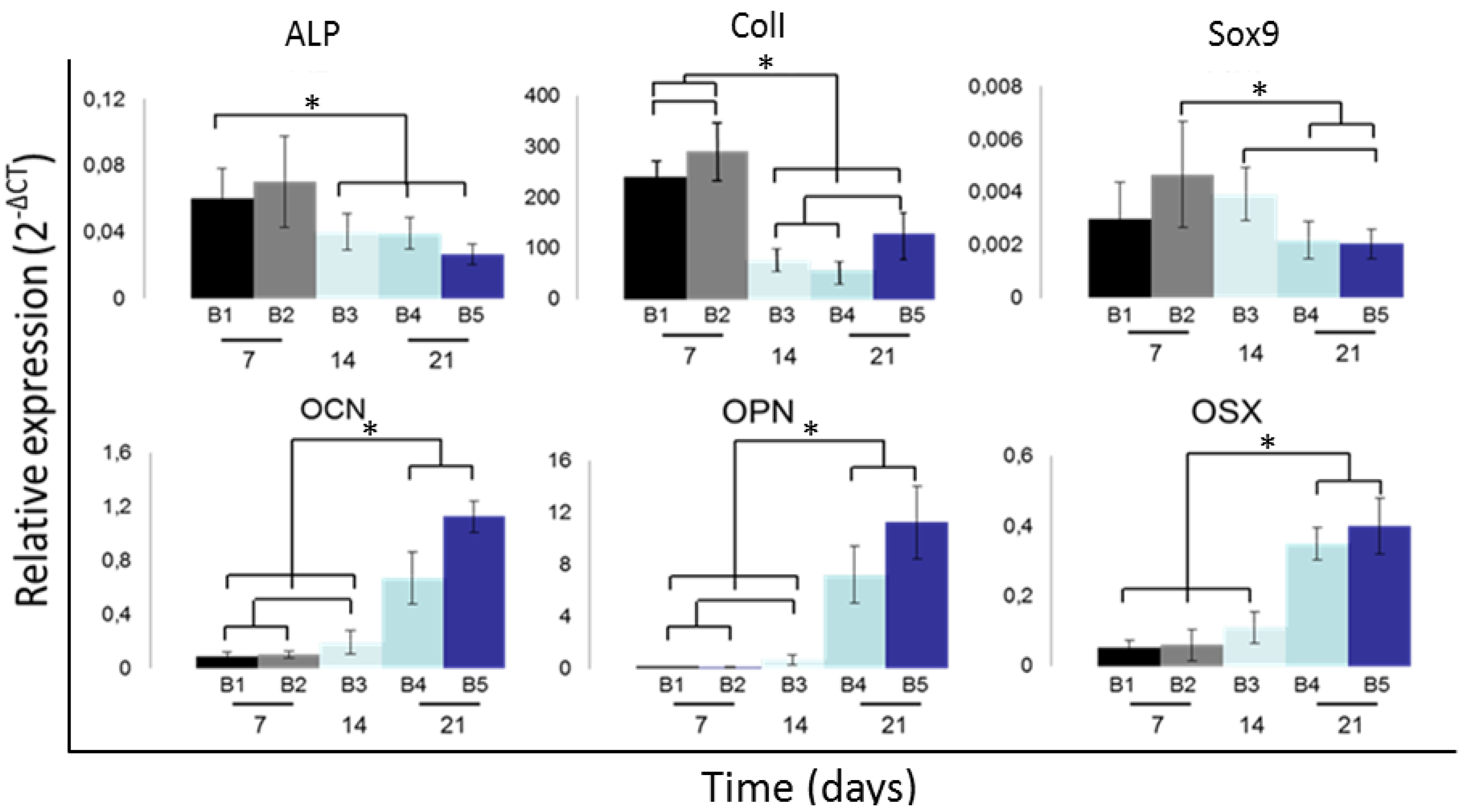
3.2. Osteogenic Cell Differentiation: An Overview


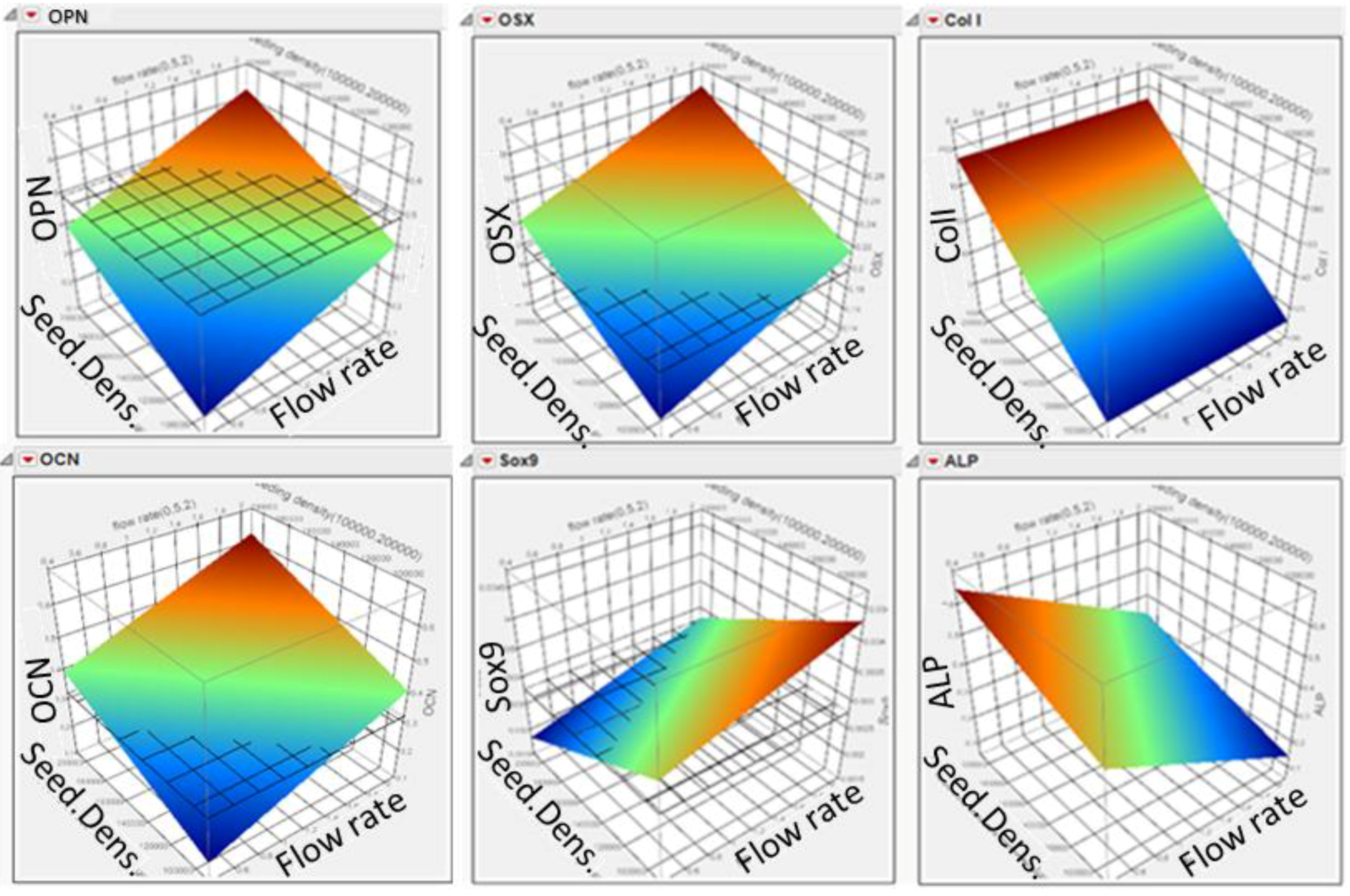
3.3. DoE Gene Expression Analysis: Flow Rate and Seeding Density Effect

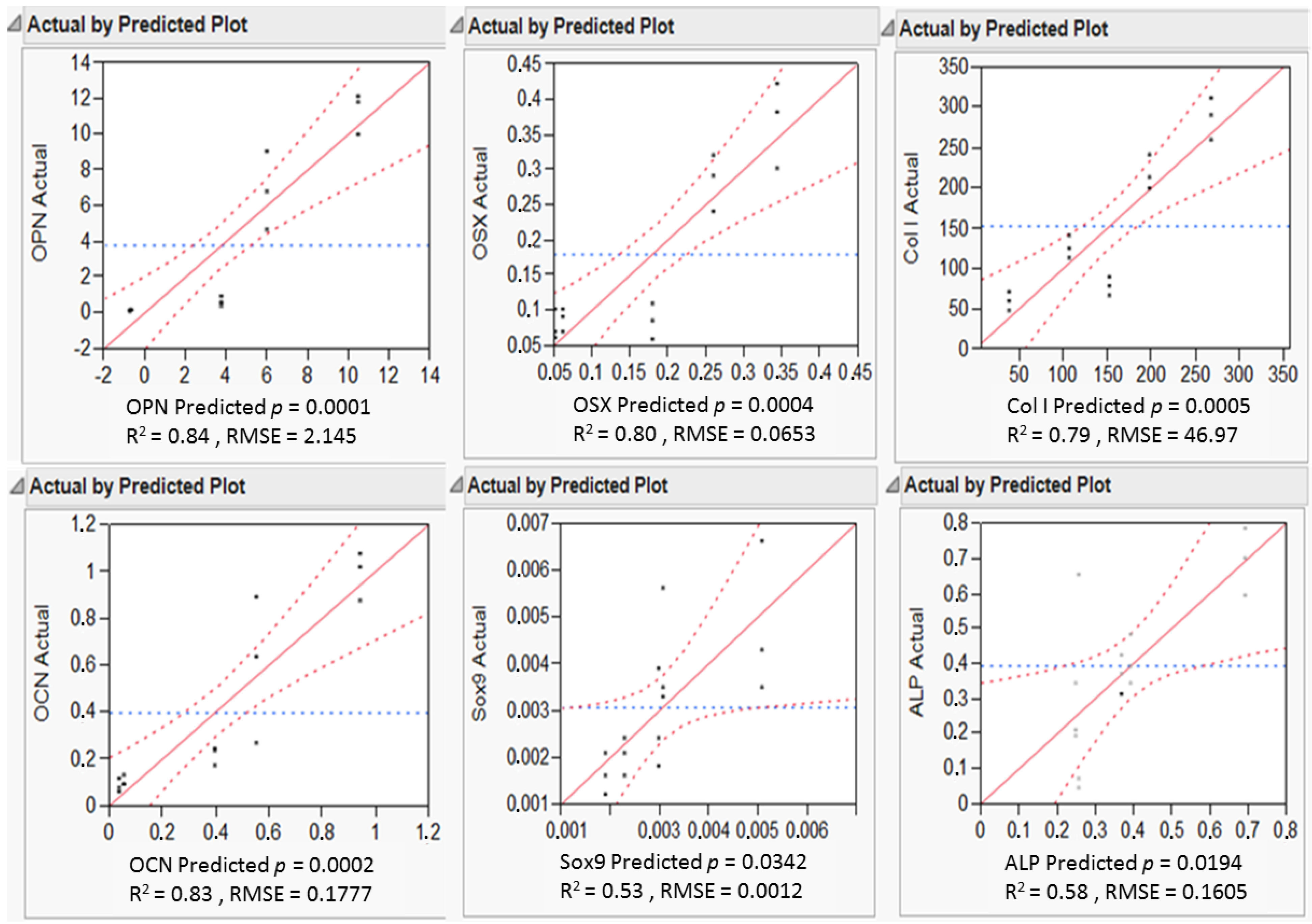
3.4. Highlighting Gene Expression Signature Windows via Response Surface Methodology


4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Laurencin, C.T.; Ambrosio, A.M.A.; Borden, M.D.; Cooper, J.A. Tissue engineering: Orthopedic applications. Annu. Rev. Biomed. Eng. 1999, 1, 19–46. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.M. Biomaterials for bone tissue engineering. Mater. Today 2008, 11, 18–25. [Google Scholar] [CrossRef]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H.K. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, G.N.; Sikavitsas, V.I.; Mikos, A.G. Design of a flow perfusion bioreactor system for bone tissue-engineering applications. Tissue Eng. 2003, 9, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Grayson, W.L.; Bhumiratana, S.; Cannizzaro, C.; Vunjak-Novakovic, G. Bioreactor cultivation of functional bone grafts. Methods Mol. Biol. 2011, 698, 231–241. [Google Scholar] [PubMed]
- Datta, N.; Pham, Q.P.; Sharma, U.; Sikavitsas, V.I.; Jansen, J.A.; Mikos, A.G. In vitro generated extracellular matrix and fluid shear stress synergistically enhance 3D osteoblastic differentiation. Proc. Natl. Acad. Sci. USA 2006, 103, 2488–2493. [Google Scholar] [CrossRef] [PubMed]
- Yeatts, A.B.; Fisher, J.P. Bone tissue engineering bioreactors: Dynamic culture and the influence of shear stress. Bone 2011, 48, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Placzek, M.R.; Chung, I.M.; Macedo, H.M.; Ismail, S.; Blanco, T.M.; Lim, M.; Cha, J.M.; Fauzi, I.; Kang, Y.; Yeo, D.C.L.; Ma, C.Y.J.; et al. Stem cell bioprocessing: Fundamentals and principles. J. R. Soc. Interface 2009, 6, 209–232. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, E.; Thomas, R.J.; Williams, D.J. Current understanding and challenges in bioprocessing of stem cell-based therapies for regenerative medicine. Br. Med. Bull. 2011, 100, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Lenas, P.; Moos, M.; Luyten, F.P. Developmental engineering: A new paradigm for the design and manufacturing of cell-based products. Part I: From three-dimensional cell growth to biomimetics of in vivo development. Tissue Eng. Part B 2009, 15, 381–394. [Google Scholar]
- Thomas, R.J.; Hourd, P.C.; Williams, D.J. Application of process quality engineering techniques to improve the understanding of the in vitro processing of stem cells for therapeutic use. J. Biotechnol. 2008, 136, 148–155. [Google Scholar] [CrossRef]
- Ratcliffe, E.; Hourd, P.; Guijarro-Leach, J.; Rayment, E.; Williams, D.J.; Thomas, R.J. Application of response surface methodology to maximize the productivity of scalable automated human embryonic stem cell manufacture. Regen. Med. 2013, 8, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Panoskaltsis, N.; Ye, H.; Mantalaris, A. Optimization of in vitro erythropoiesis from CD34+ cord blood cells using design of experiments (DOE). Biochem. Eng. J. 2011, 55, 154–161. [Google Scholar] [CrossRef]
- Papantoniou, I.I.; Chai, Y.C.; Luyten, F.P.; Schrooten, J.I. Process quality engineering for bioreactor-driven manufacturing of tissue-engineered constructs for bone regeneration. Tissue Eng. Part C 2013. [Google Scholar] [CrossRef]
- Chen, Y.; Bloemen, V.; Impens, S.; Moesen, M.; Luyten, F.P.; Schrooten, J. Characterization and optimization of cell seeding in scaffolds by factorial design: Quality by design approach for skeletal tissue engineering. Tissue Eng. Part C 2011, 17, 1211–1221. [Google Scholar] [CrossRef]
- Hunt, M.M.; Meng, G.L.; Rancourt, D.E.; Gates, I.D.; Kallos, M.S. Factorial experimental design for the culture of human embryonic stem cells as aggregates in stirred suspension bioreactors reveals the potential for interaction effects between bioprocess parameters. Tissue Eng. Part C 2014, 20, 76–89. [Google Scholar] [CrossRef]
- Cartmell, S.H.; Porter, B.D.; Garcia, A.J.; Guldberg, R.E. Effects of medium perfusion rate on cell-seeded three-dimensional bone constructs in vitro. Tissue Eng. 2003, 9, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Sikavitsas, V.I.; Bancroft, G.N.; Lemoine, J.J.; Liebschner, M.A.K.; Dauner, M.; Mikos, A.G. Flow perfusion enhances the calcified matrix deposition of marrow stromal cells in biodegradable nonwoven fiber mesh scaffolds. Ann. Biomed. Eng. 2005, 33, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bjerre, L.; Bunger, C.E.; Kassem, M.; Mygind, T. Flow perfusion culture of human mesenchymal stem cells on silicate-substituted tricalcium phosphate scaffolds. Biomaterials 2008, 29, 2616–2627. [Google Scholar] [CrossRef] [PubMed]
- McCoy, R.J.; O’Brien, F.J. Influence of shear stress in perfusion bioreactor cultures for the development of three-dimensional bone tissue constructs: A review. Tissue Eng. Part B 2010, 16, 587–601. [Google Scholar] [CrossRef]
- Kim, J.; Ma, T. Perfusion regulation of hMSC microenvironment and osteogenic differentiation in 3D scaffold. Biotechnol. Bioeng. 2012, 109, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.C.; Roberts, S.J.; van Bael, S.; Chen, Y.; Luyten, F.P.; Schrooten, J. Multi-level factorial analysis of Ca2+/Pi supplementation as bio-instructive media for in vitro biomimetic engineering of three-dimensional osteogenic hybrids. Tissue Eng. Part C 2012, 18, 90–103. [Google Scholar] [CrossRef]
- Bitar, M.; Brown, R.A.; Salih, V.; Kidane, A.G.; Knowles, J.C.; Nazhat, S.N. Effect of cell density on osteoblastic differentiation and matrix degradation of biomimetic dense collagen scaffolds. Biomacromolecules 2008, 9, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Eyckmans, J.; Lin, G.L.; Chen, C.S. Adhesive and mechanical regulation of mesenchymal stem cell differentiation in human bone marrow and periosteum-derived progenitor cells. Biol. Open 2012, 1, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Dean, D.; Mikos, A.G.; Fisher, J.P. Effect of initial cell seeding density on early osteogenic signal expression of rat bone marrow stromal cells cultured on cross-linked poly(propylene fumarate) disks. Biomacromolecules 2009, 10, 1810–1817. [Google Scholar] [CrossRef] [PubMed]
- Salter, E.; Goh, B.; Hung, B.; Hutton, D.; Ghone, N.; Grayson, W.L. Bone tissue engineering bioreactors: A role in the clinic? Tissue Eng. Part B 2012, 18, 62–75. [Google Scholar]
- Lim, M.; Shunjie, C.; Panoskaltsis, N.; Mantalaris, A. Systematic experimental design for bioprocess characterization: Elucidating transient effects of multi-cytokine contributions on erythroid differentiation. Biotechnol. Bioproc. E 2012, 17, 218–226. [Google Scholar] [CrossRef]
- Pyka, G.; Burakowski, A.; Kerckhofs, G.; Moesen, M.; van Bael, S.; Schrooten, J.; Wevers, M. Surface modification of Ti6Al4V open porous structures produced by additive manufacturing. Adv. Eng. Mater. 2012, 14, 363–370. [Google Scholar] [CrossRef]
- Eyckmans, J.; Luyten, F.P. Species specificity of ectopic bone formation using periosteum-derived mesenchymal progenitor cells. Tissue Eng. 2006, 12, 2203–2213. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.C.; Roberts, S.J.; Schrooten, J.; Luyten, F.P. Probing the osteoinductive effect of calcium phosphate by using an in vitro biomimetic model. Tissue Eng. Part A 2011, 17, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.C.; Truscello, S.; Bael, S.V.; Luyten, F.P.; Vleugels, J.; Schrooten, J. Perfusion electrodeposition of calcium phosphate on additive manufactured titanium scaffolds for bone engineering. Acta Biomater. 2011, 7, 2310–2319. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sonnaert, M.; Roberts, S.J.; Luyten, F.P.; Schrooten, J. Validation of a PicoGreen-based DNA quantification integrated in an RNA extraction method for two-dimensional and three-dimensional cell cultures. Tissue Eng. Part C 2012, 18, 444–452. [Google Scholar] [CrossRef]
- Van Bael, S.; Chai, Y.C.; Truscello, S.; Moesen, M.; Kerckhofs, G.; van Oosterwyck, H.; Kruth, J.P.; Schrooten, J. The effect of pore geometry on the in vitro biological behavior of human periosteum-derived cells seeded on selective laser-melted Ti6Al4V bone scaffolds. Acta Biomater. 2012, 8, 2824–2834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Pathi, P.; Grayson, W.; Xing, Q.; Locke, B.R.; Ma, T. Effects of oxygen transport on 3-d human mesenchymal stem cell metabolic activity in perfusion and static cultures: Experiments and mathematical model. Biotechnol. Prog. 2005, 21, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Kommareddy, K.P.; Lange, C.; Rumpler, M.; Dunlop, J.W.C.; Manjubala, I.; Cui, J.; Kratz, K.; Lendlein, A.; Fratzl, P. Two stages in three-dimensional in vitro growth of tissue generated by osteoblastlike cells. Biointerphases 2010, 5, 45–52. [Google Scholar] [CrossRef]
- Pyka, G.; Kerckhofs, G.; Papantoniou, I.; Speirs, M.; Schrooten, J.; Wevers, M. Surface roughness and morphology customization of additive manufactured open porous Ti6Al4V structures. Materials 2013, 6, 4737–4757. [Google Scholar] [CrossRef]
- Kreke, M.R.; Goldstein, A.S. Hydrodynamic shear stimulates osteocalcin expression but not proliferation of bone marrow stromal cells. Tissue Eng. 2004, 10, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Grayson, W.L.; Marolt, D.; Bhumiratana, S.; Frohlich, M.; Guo, X.E.; Vunjak-Novakovic, G. Optimizing the medium perfusion rate in bone tissue engineering bioreactors. Biotechnol. Bioeng. 2011, 108, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- McCoy, R.J.; Jungreuthmayer, C.; O’Brien, F.J. Influence of flow rate and scaffold pore size on cell behavior during mechanical stimulation in a flow perfusion bioreactor. Biotechnol. Bioeng. 2012, 109, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.J.; Chen, Y.; Moesen, M.; Schrooten, J.; Luyten, F.P. Enhancement of osteogenic gene expression for the differentiation of human periosteal derived cells. Stem. Cell Res. 2011, 7, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Stiehler, M.; Bunger, C.; Baatrup, A.; Lind, M.; Kassem, M.; Mygind, T. Effect of dynamic 3-D culture on proliferation, distribution, and osteogenic differentiation of human mesenchymal stem cells. J. Biomed. Mater. Res. Part A 2009, 89A, 96–107. [Google Scholar]
- Barradas, A.M.; Fernandes, H.A.; Groen, N.; Chai, Y.C.; Schrooten, J.; van de Peppel, J.; van Leeuwen, J.P.; van Blitterswijk, C.A.; de Boer, J. A calcium-induced signaling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials 2012, 33, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
- Vehof, J.W.M.; de Ruijter, A.E.; Spauwen, P.H.M.; Jansen, J.A. Influence of rhBMP-2 on rat bone marrow stromal cells cultured on titanium fiber mesh. Tissue Eng. 2001, 7, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Z.; Weir, M.D.; Xu, H.H.K. Effect of cell seeding density on proliferation and osteodifferentiation of umbilical cord stem cells on calcium phosphate cement-fiber scaffold. Tissue Eng. Part A 2011, 17, 2603–2613. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Reilly, G.C.; Zhen, X.C.; Yellowley, C.E.; Chen, Q.; Donahue, H.J.; Jacobs, C.R. Osteopontin gene regulation by oscillatory fluid flow via intracellular calcium mobilization and activation of mitogen-activated protein kinase in MC3T3-E1 osteoblasts. J. Biol. Chem. 2001, 276, 13365–13371. [Google Scholar] [CrossRef] [PubMed]
- Kreke, M.R.; Huckle, W.R.; Goldstein, A.S. Fluid flow stimulates expression of osteopontin and bone sialoprotein by bone marrow stromal cells in a temporally dependent manner. Bone 2005, 36, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Scaglione, S.; Wendt, D.; Miggino, S.; Papadimitropoulos, A.; Fato, M.; Quarto, R.; Martin, I. Effects of fluid flow and calcium phosphate coating on human bone marrow stromal cells cultured in a defined 2D model system. J. Biomed. Mater. Res. Part A 2008, 86A, 411–419. [Google Scholar] [CrossRef]
- Meinel, L.; Karageorgiou, V.; Hofmann, S.; Fajardo, R.; Snyder, B.; Li, C.; Zichner, L.; Langer, R.; Vunjak-Novakovic, G.; Kaplan, D.L. Engineering bone-like tissue in vitro using human bone marrow stem cells and silk scaffolds. J. Biomed. Mater. Res. Part A 2004, 71, 25–34. [Google Scholar] [CrossRef]
- Yu, X.J.; Botchwey, E.A.; Levine, E.M.; Pollack, S.R.; Laurencin, C.T. Bioreactor-based bone tissue engineering: The influence of dynamic flow on osteoblast phenotypic expression and matrix mineralization. Proc. Natl. Acad. Sci. USA 2004, 101, 11203–11208. [Google Scholar] [CrossRef]
- Bancroft, G.N.; Sikavitsast, V.I.; van den Dolder, J.; Sheffield, T.L.; Ambrose, C.G.; Jansen, J.A.; Mikos, A.G. Fluid flow increases mineralized matrix deposition in 3D perfusion culture of marrow stromal osteloblasts in a dose-dependent manner. Proc. Natl. Acad. Sci. USA 2002, 99, 12600–12605. [Google Scholar] [CrossRef] [PubMed]
- Yourek, G.; McCormick, S.M.; Mao, J.J.; Reilly, G.C. Shear stress induces osteogenic differentiation of human mesenchymal stem cells. Regen. Med. 2010, 5, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Sikavitsas, V.I.; Bancroft, G.N.; Holtorf, H.L.; Jansen, J.A.; Mikos, A.G. Mineralized matrix deposition by marrow stromal osteoblasts in 3D perfusion culture increases with increasing fluid shear forces. Proc. Natl. Acad. Sci. USA 2003, 100, 14683–14688. [Google Scholar] [CrossRef] [PubMed]
- Grellier, M.; Bareille, R.; Bourget, C.; Amedee, J. Responsiveness of human bone marrow stromal cells to shear stress. J. Tissue Eng. Regen. Med. 2009, 3, 302–309. [Google Scholar] [CrossRef] [PubMed]
- McCoy, R.J.; O'Brien, F.J. Visualizing feasible operating ranges within tissue engineering systems using a “windows of operation” approach: A perfusion-scaffold bioreactor case study. Biotechnol. Bioeng. 2012, 109, 3161–3171. [Google Scholar] [CrossRef]
- Barbosa, H.S.C.; Fernandes, T.G.; Dias, T.P.; Diogo, M.M.; Cabral, J.M.S. New insights into the mechanisms of embryonic stem cell self-renewal under hypoxia: A multifactorial analysis approach. PLoS One 2012, 7. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Papantoniou, I.; Sonnaert, M.; Lambrechts, T.; Aerts, J.-M.; Geris, L.; Luyten, F.P.; Schrooten, J. Analysis of Gene Expression Signatures for Osteogenic 3D Perfusion-Bioreactor Cell Cultures Based on a Multifactorial DoE Approach. Processes 2014, 2, 639-657. https://doi.org/10.3390/pr2030639
Papantoniou I, Sonnaert M, Lambrechts T, Aerts J-M, Geris L, Luyten FP, Schrooten J. Analysis of Gene Expression Signatures for Osteogenic 3D Perfusion-Bioreactor Cell Cultures Based on a Multifactorial DoE Approach. Processes. 2014; 2(3):639-657. https://doi.org/10.3390/pr2030639
Chicago/Turabian StylePapantoniou, Ioannis, Maarten Sonnaert, Toon Lambrechts, Jean-Marie Aerts, Lies Geris, Frank P. Luyten, and Jan Schrooten. 2014. "Analysis of Gene Expression Signatures for Osteogenic 3D Perfusion-Bioreactor Cell Cultures Based on a Multifactorial DoE Approach" Processes 2, no. 3: 639-657. https://doi.org/10.3390/pr2030639