Cost-Effective Analysis of Control Strategies to Reduce the Prevalence of Cutaneous Leishmaniasis, Based on a Mathematical Model

Abstract
:1. Introduction
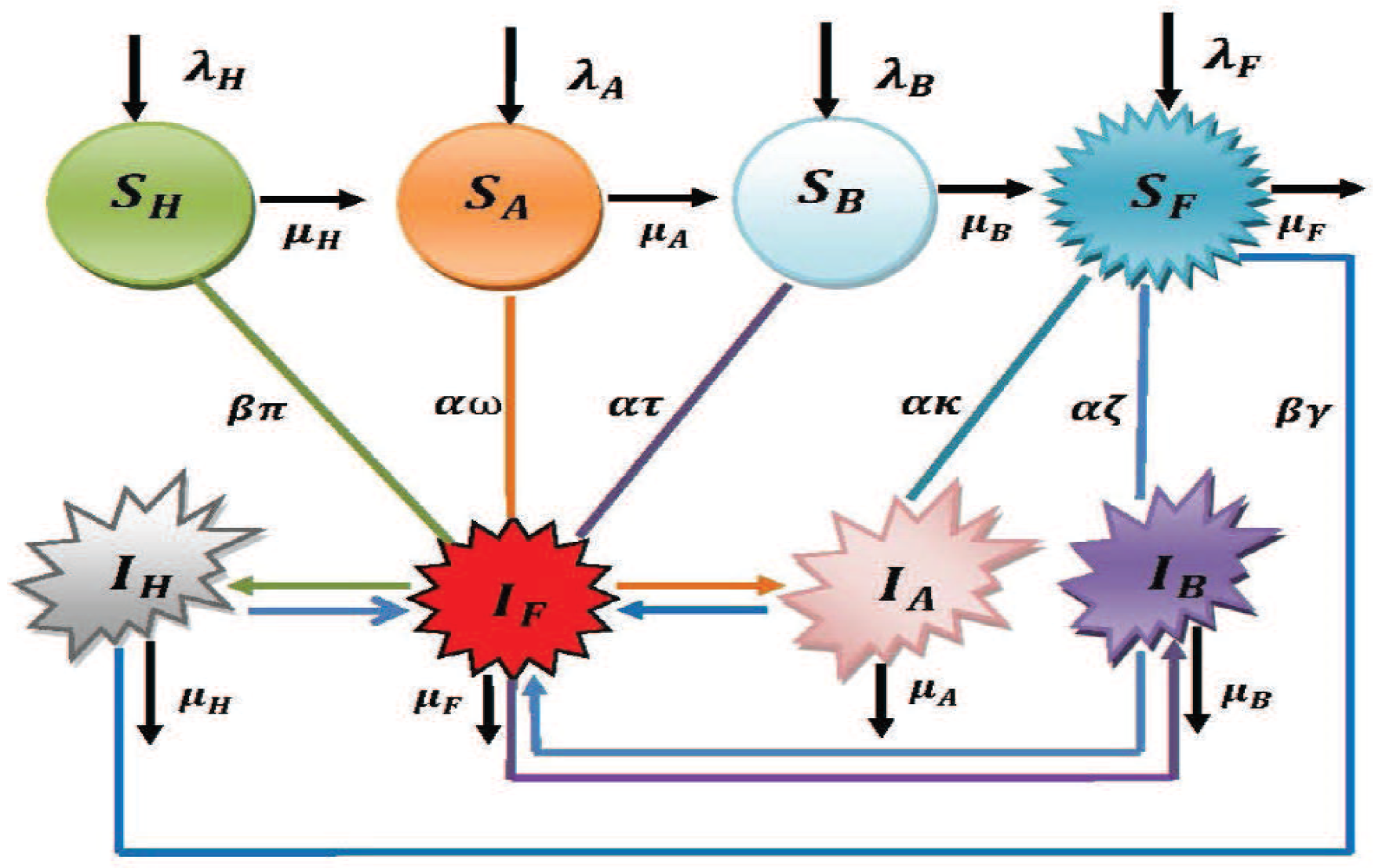
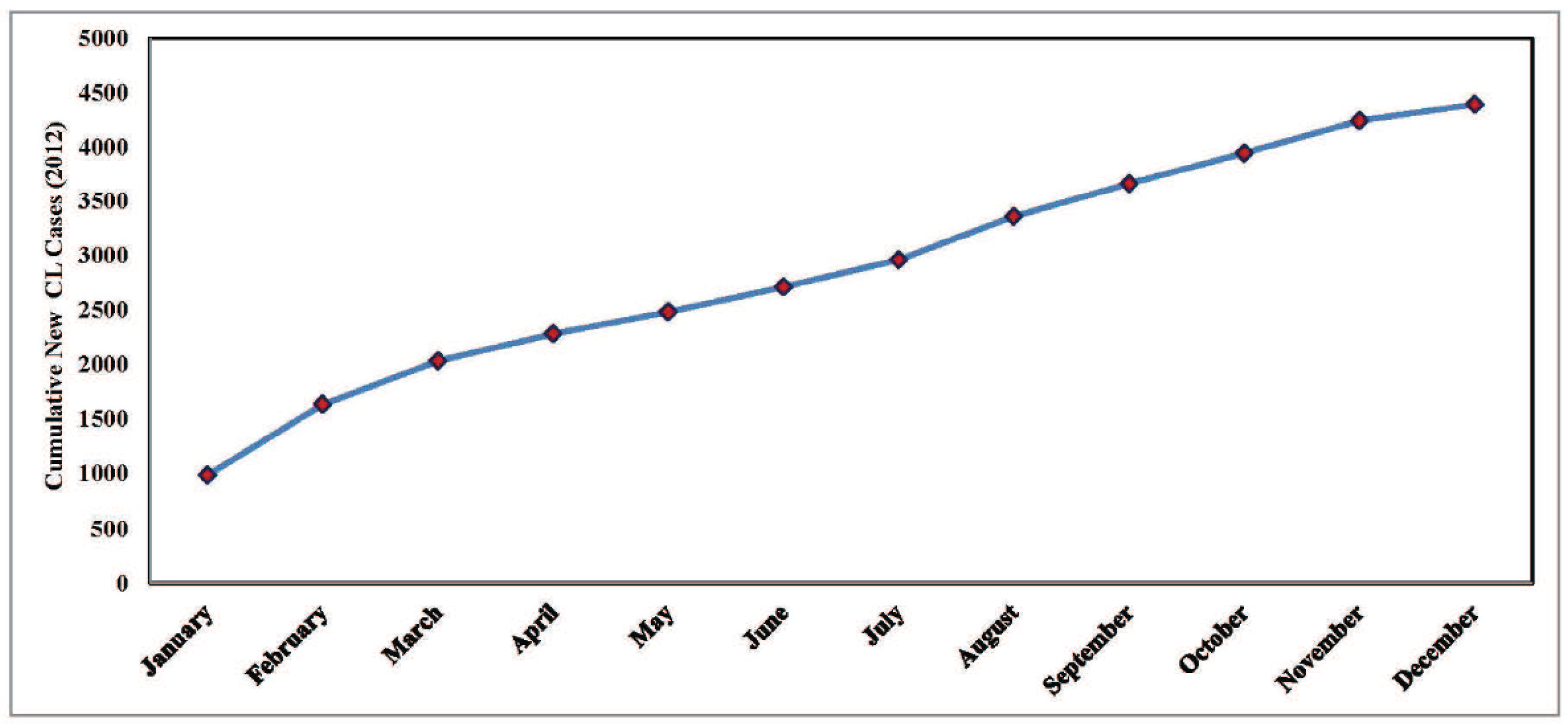
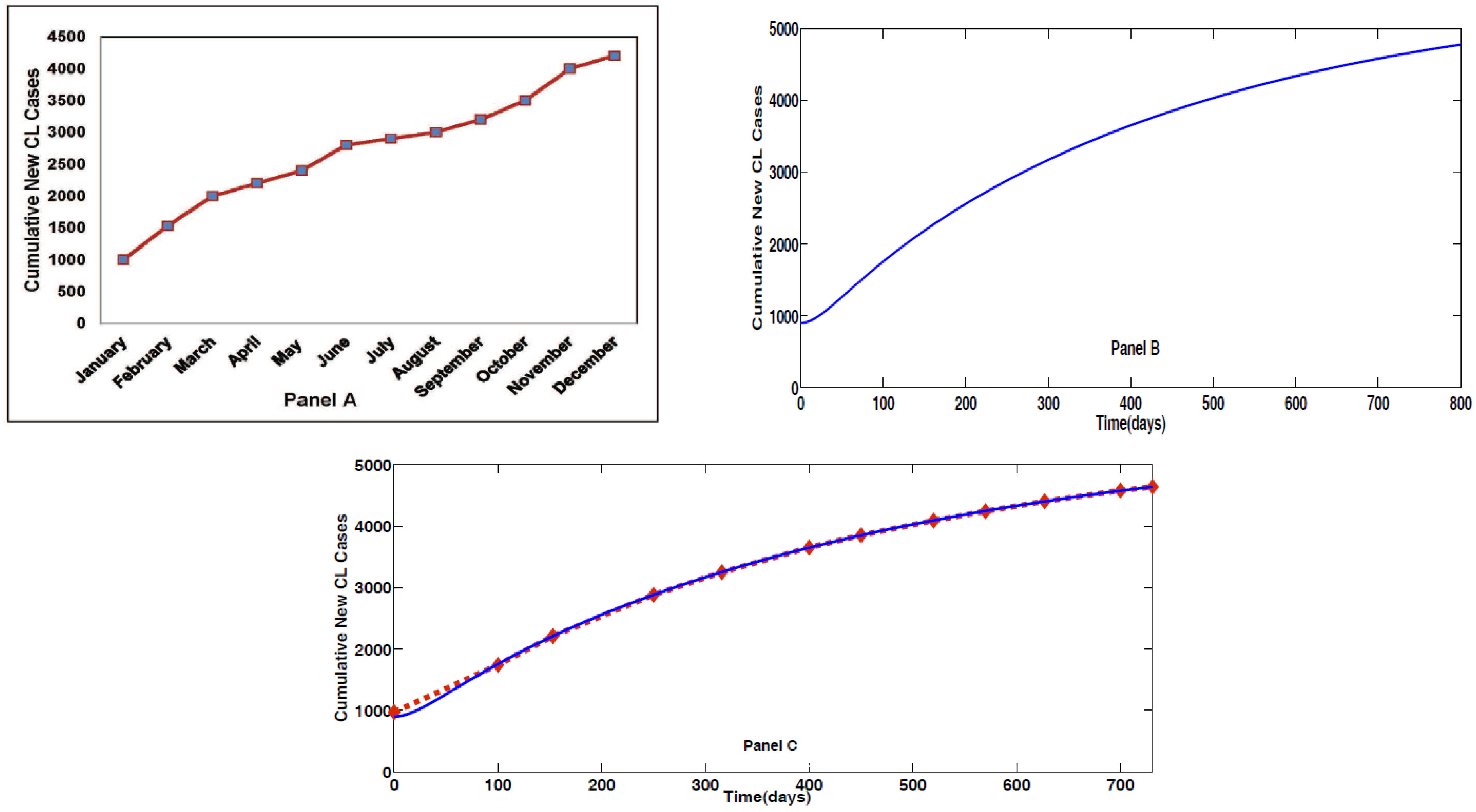
2. Model Formulation through Schematic Diagram and Its Validation
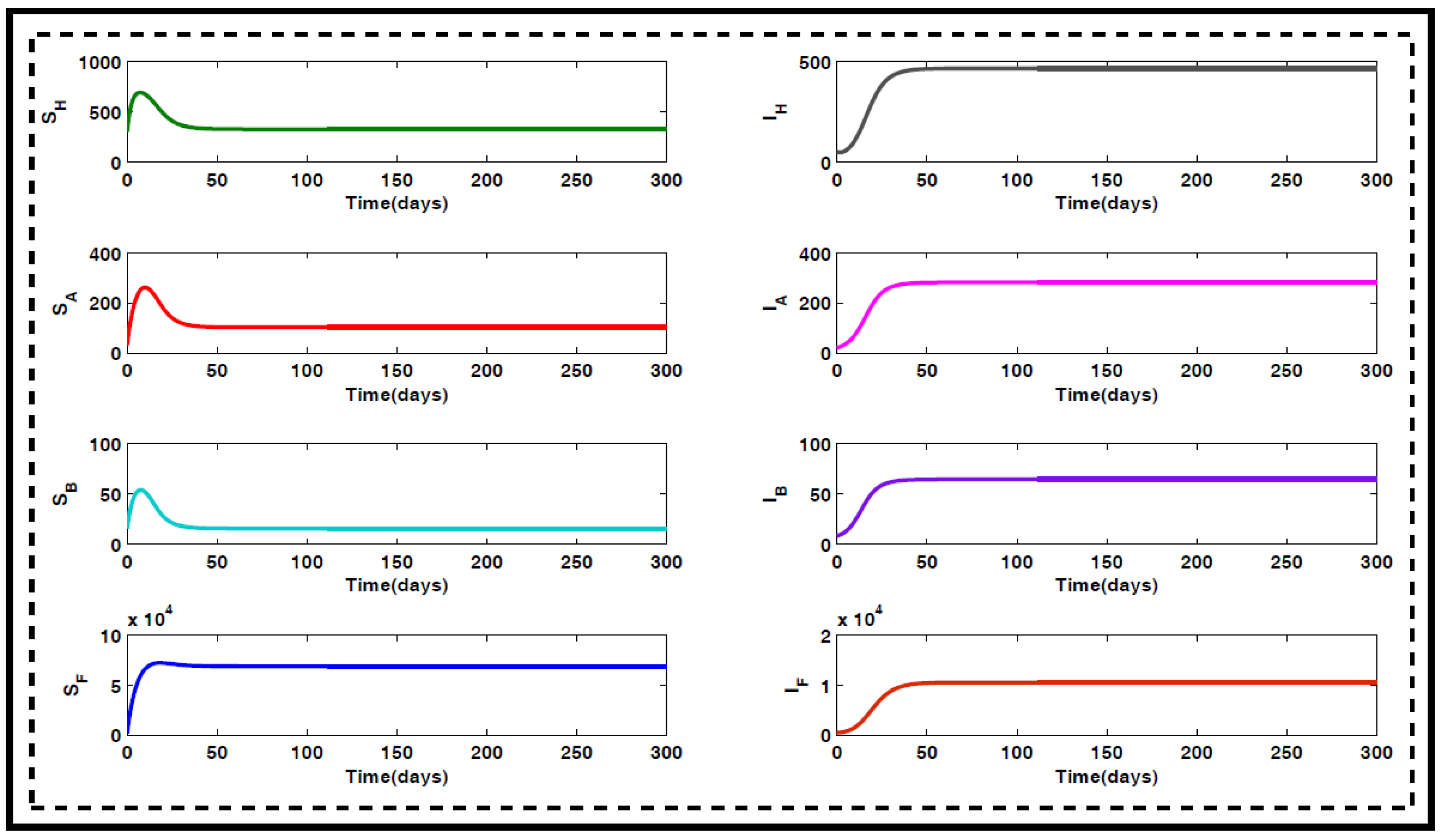
2.1. Properties of the Model
2.2. Existence Condition
- and
2.3. Analytical Study of the Formulated Model
3. Control Theoretic Approach for the Proposed Model
- (i)
- The control variable acts as a prevention of human infection using drugs and the use of insecticide-treated bed nets to reduce infection.
- (ii)
- The control variable represents the use of medicines for the prevention of an infected reservoir population of Type A.
- (iii)
- The control variable represents the use of effective medicines for the prevention of an infected reservoir population of Type B.
- (iv)
- The control variable corresponds to measures like spraying insecticide on residences and other places where sand flies can breed and live in order to kill them at all stages.
Existence of the Optimal Control
- (i)
- , where and and
- (ii)
- , where and ,
- (i)
- , where and and
- (ii)
- , where and ,
- (i)
- , where and and
- (ii)
- , where and ,
- (i)
- , where and and
- (ii)
- , where and .
- Case 1:
- , subject to the condition :
- Case 2:
- , subject to the condition and :
- Case 3:
- , subject to the condition and :
- Case 1:
- , subject to the condition :
- Case 2:
- , subject to the condition and :
- Case 3:
- , subject to the condition and :
- Case 1:
- , subject to the condition :
- Case 2:
- , subject to the condition and :
- Case 3:
- , subject to the condition and :
- Case 1:
- , subject to the condition :
- Case 2:
- , subject to the condition and :
- Case 3:
- , subject to the condition and :
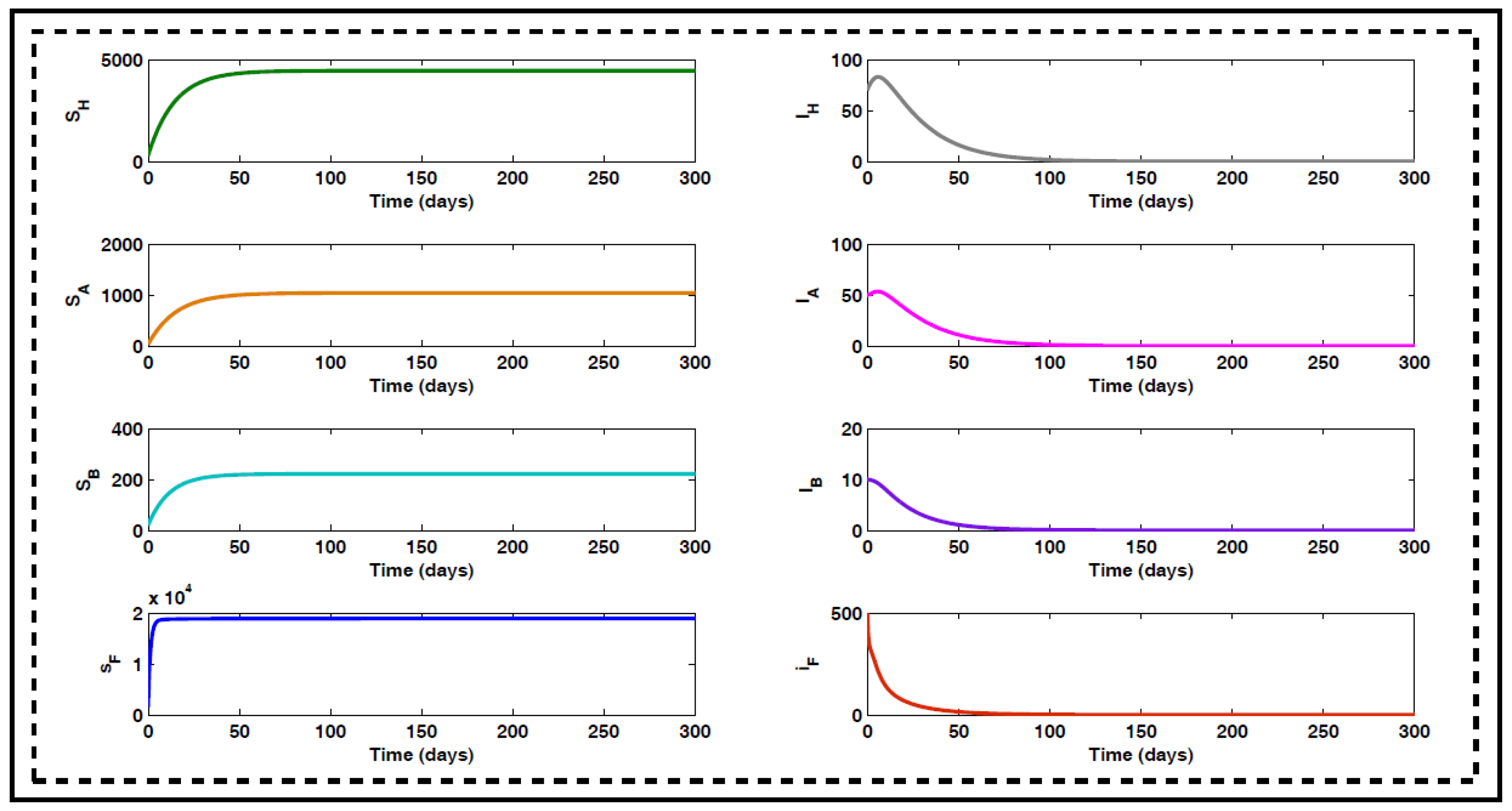
4. Numerical Simulation
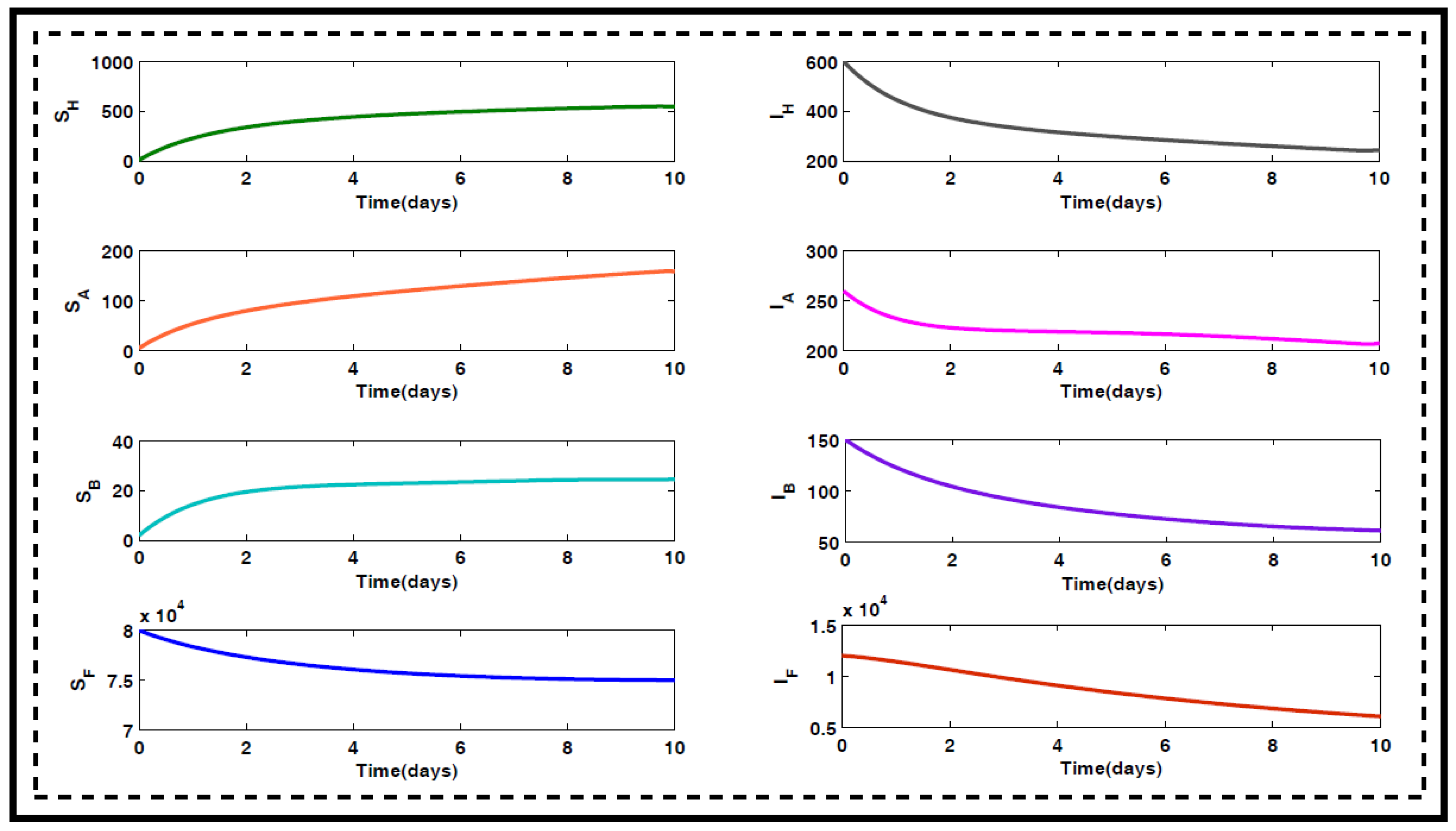
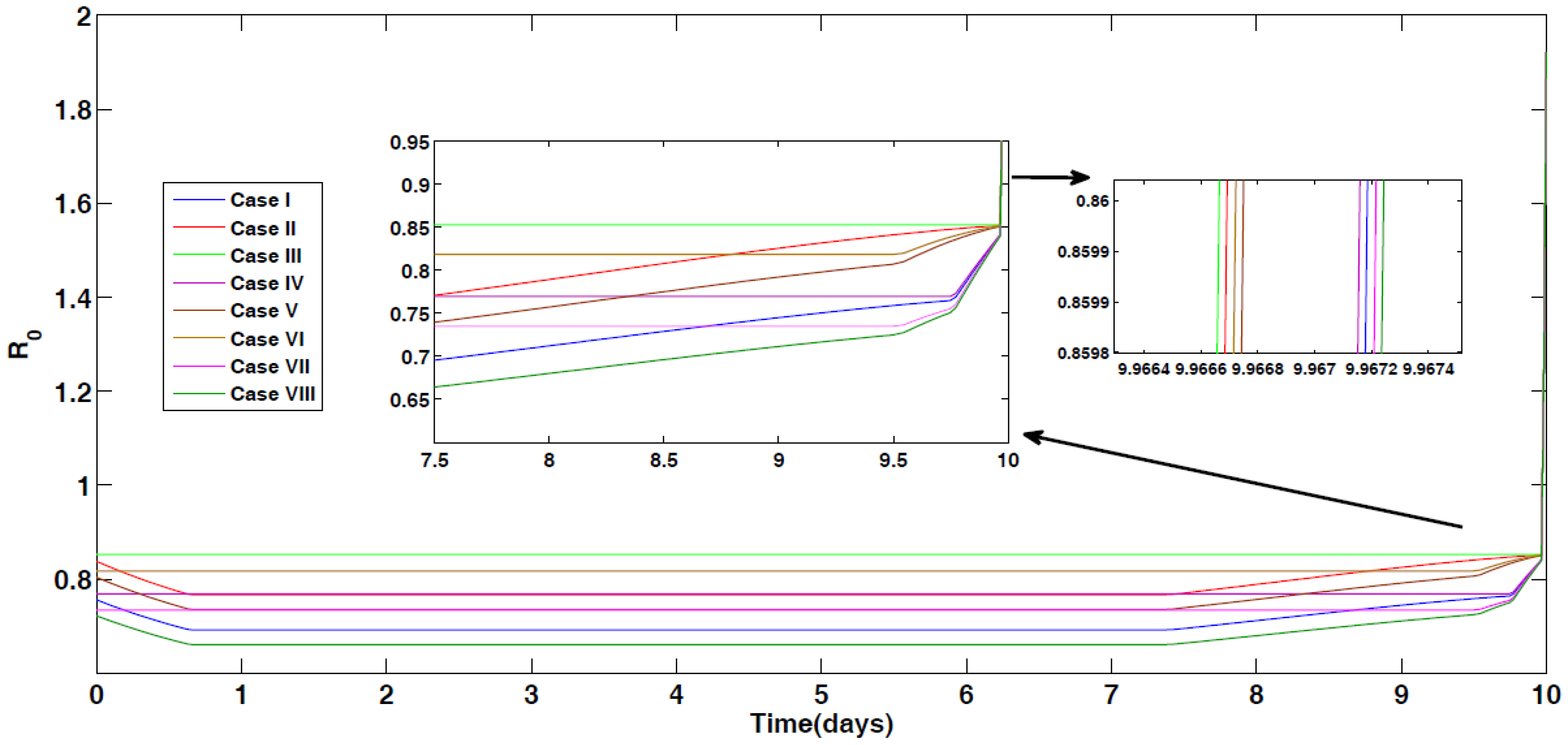
4.1. Optimal Control for Different Cases
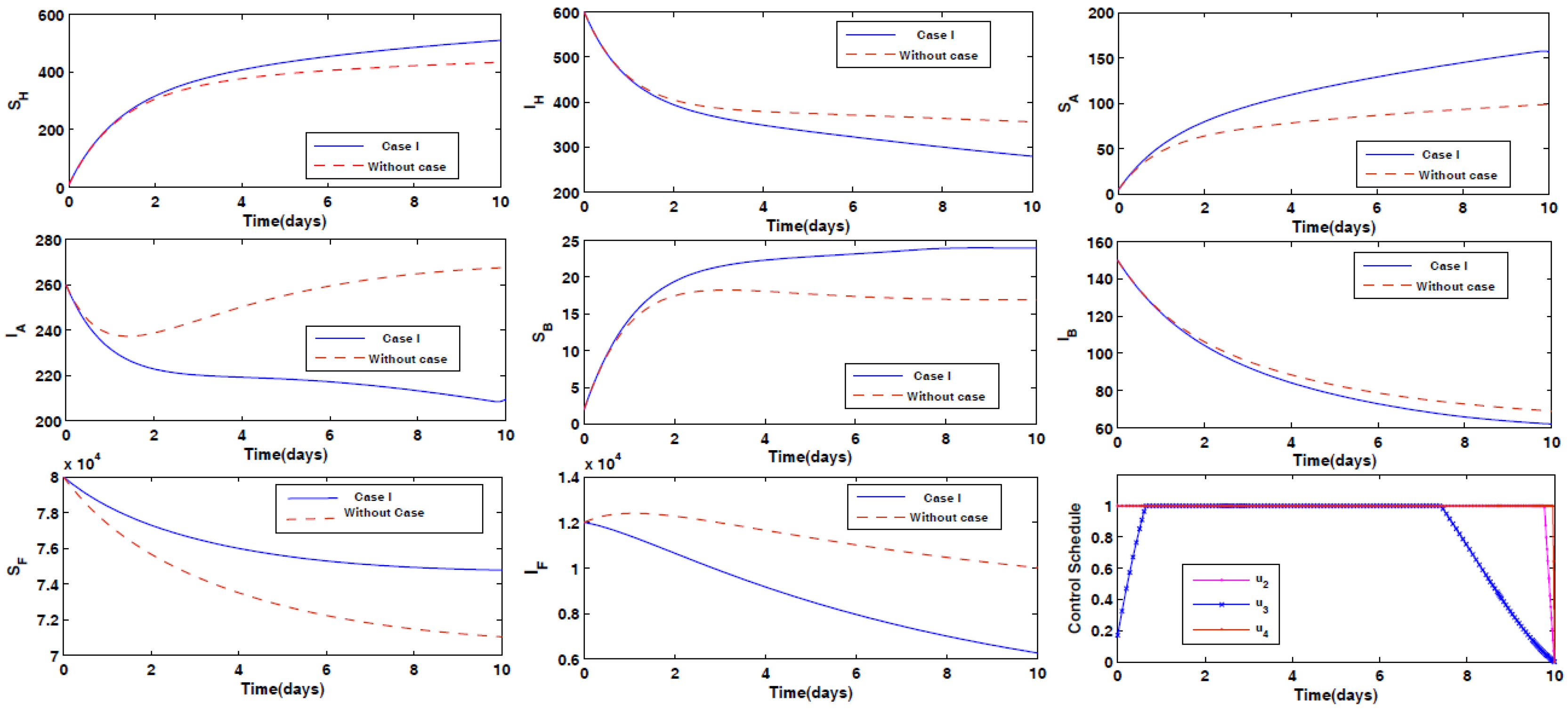
- Case I: Prevention of the infection of animals Type A and Type B by the disease, along with spraying insecticides on the sand fly vectors.
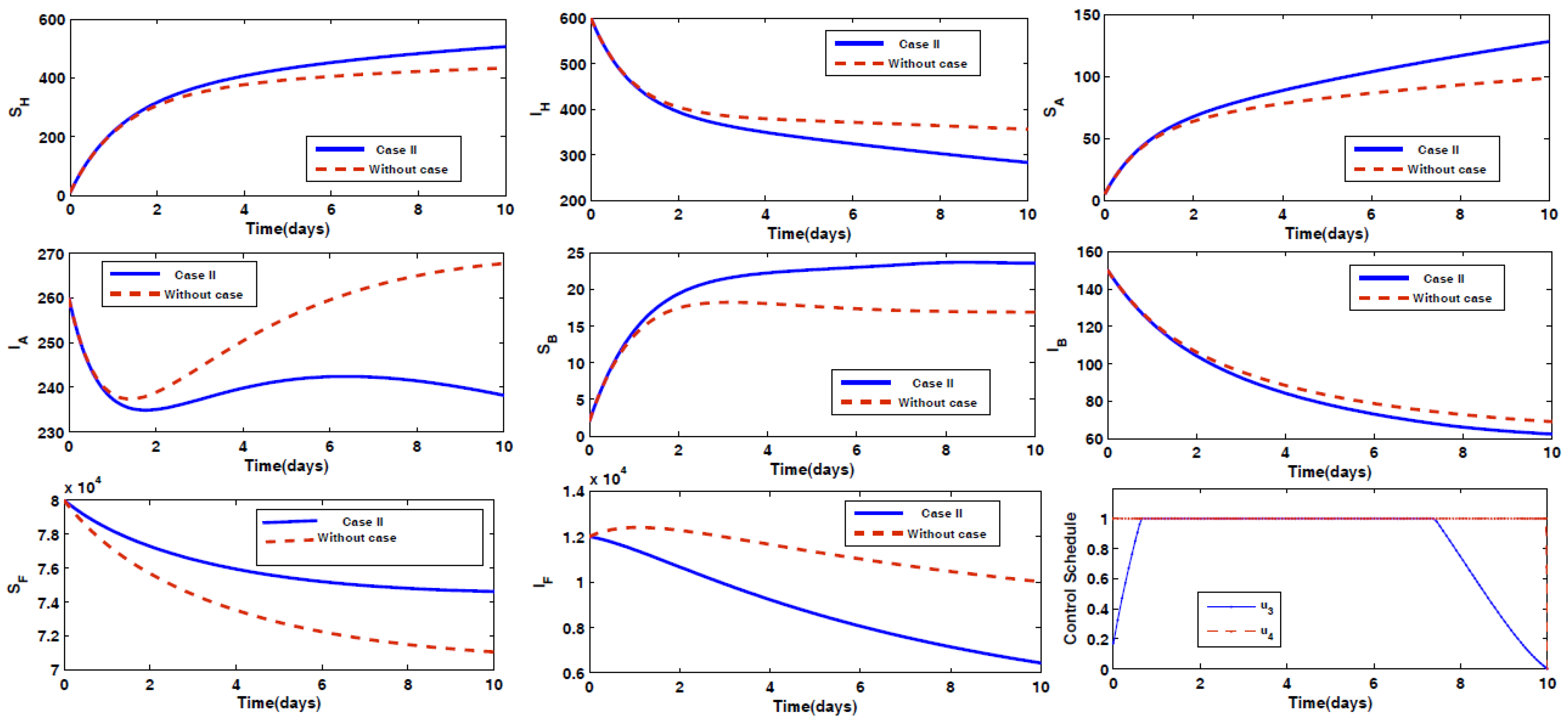
- Case II: Prevention of animal Type B being infected by the disease, along with spraying insecticides on the sand fly vectors.
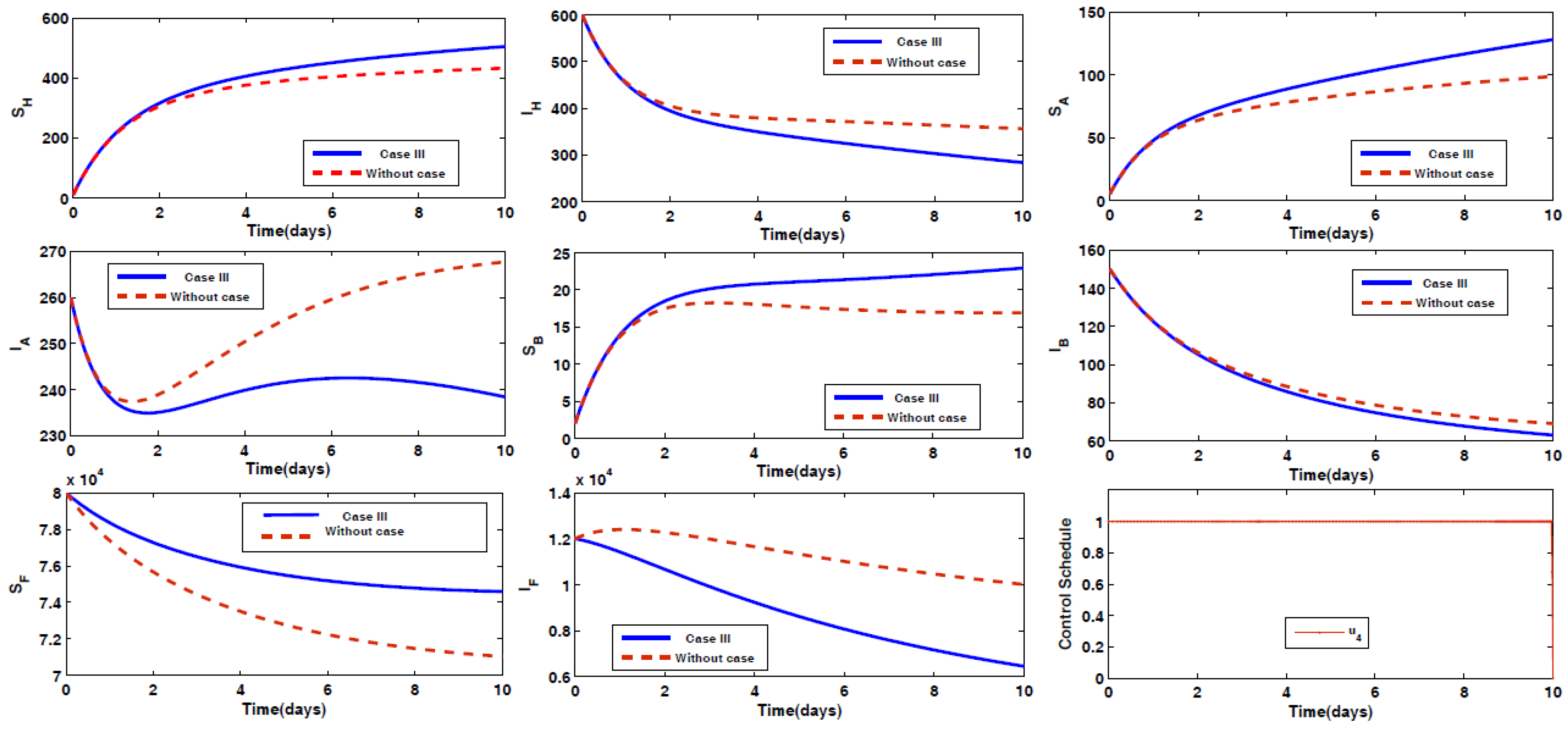
- Case III: Spraying insecticides on the sand fly vectors.
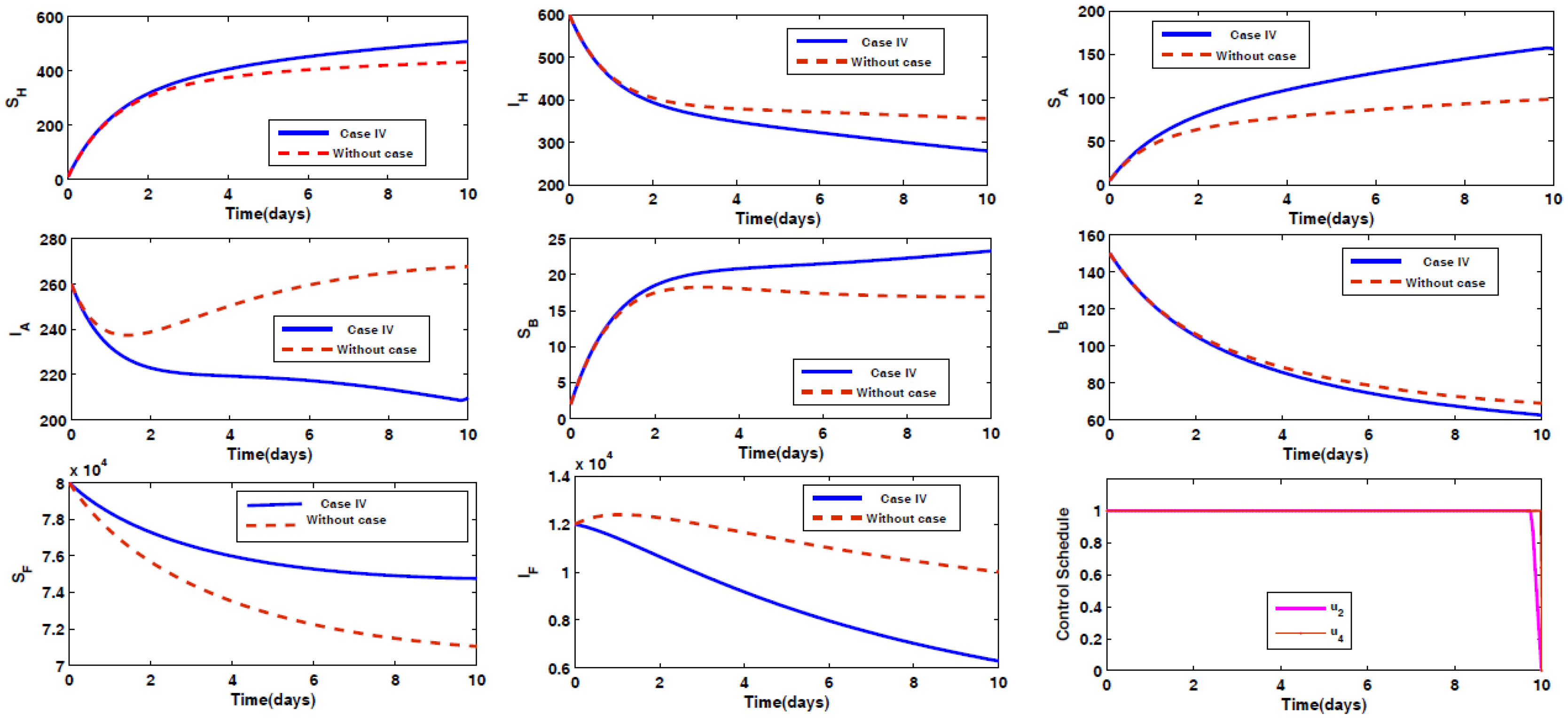
- Case IV: Prevention of animal Type A being infected by the disease, along with spraying insecticides on the sand fly vectors.
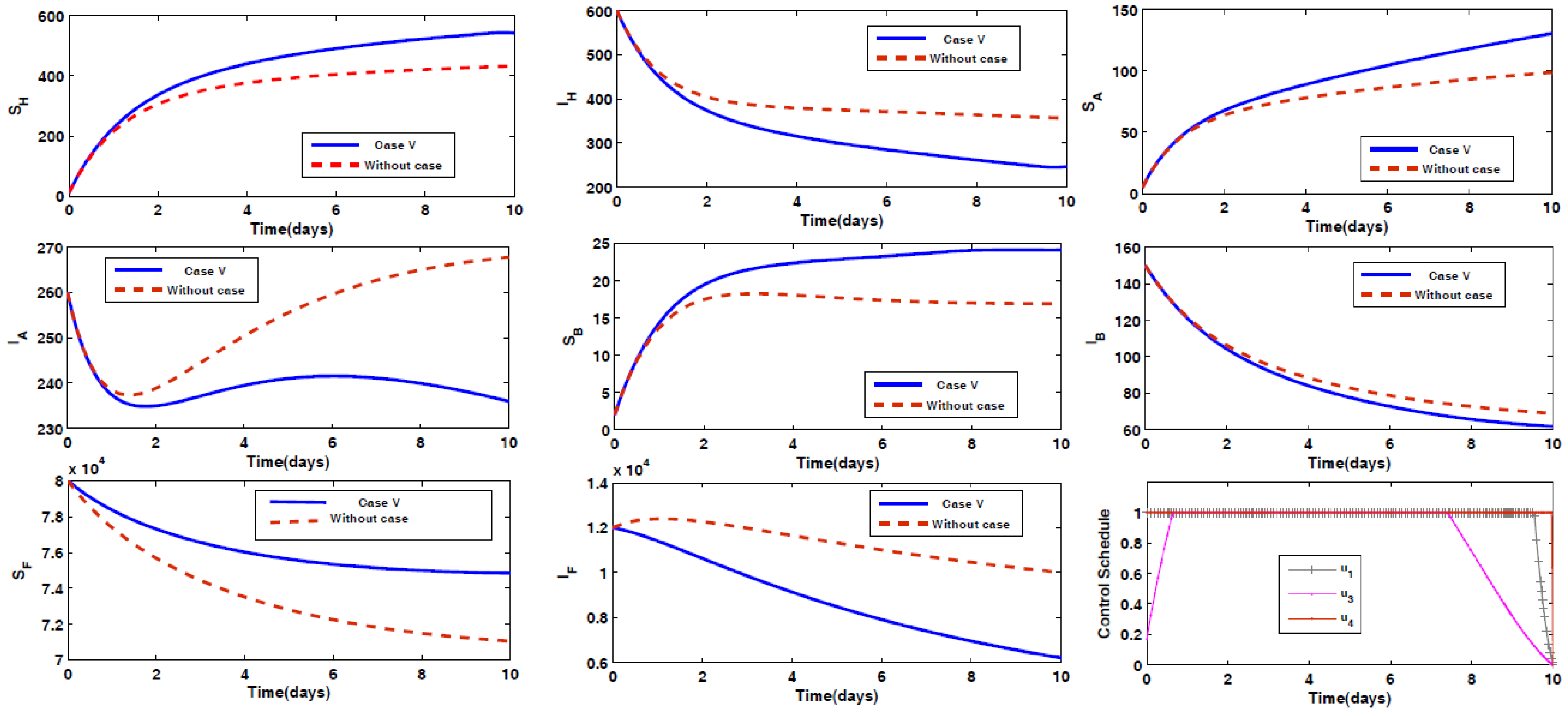
- Case V: Prevention of animal Type B being infected by the disease, along with spraying insecticides on the sand fly vectors.
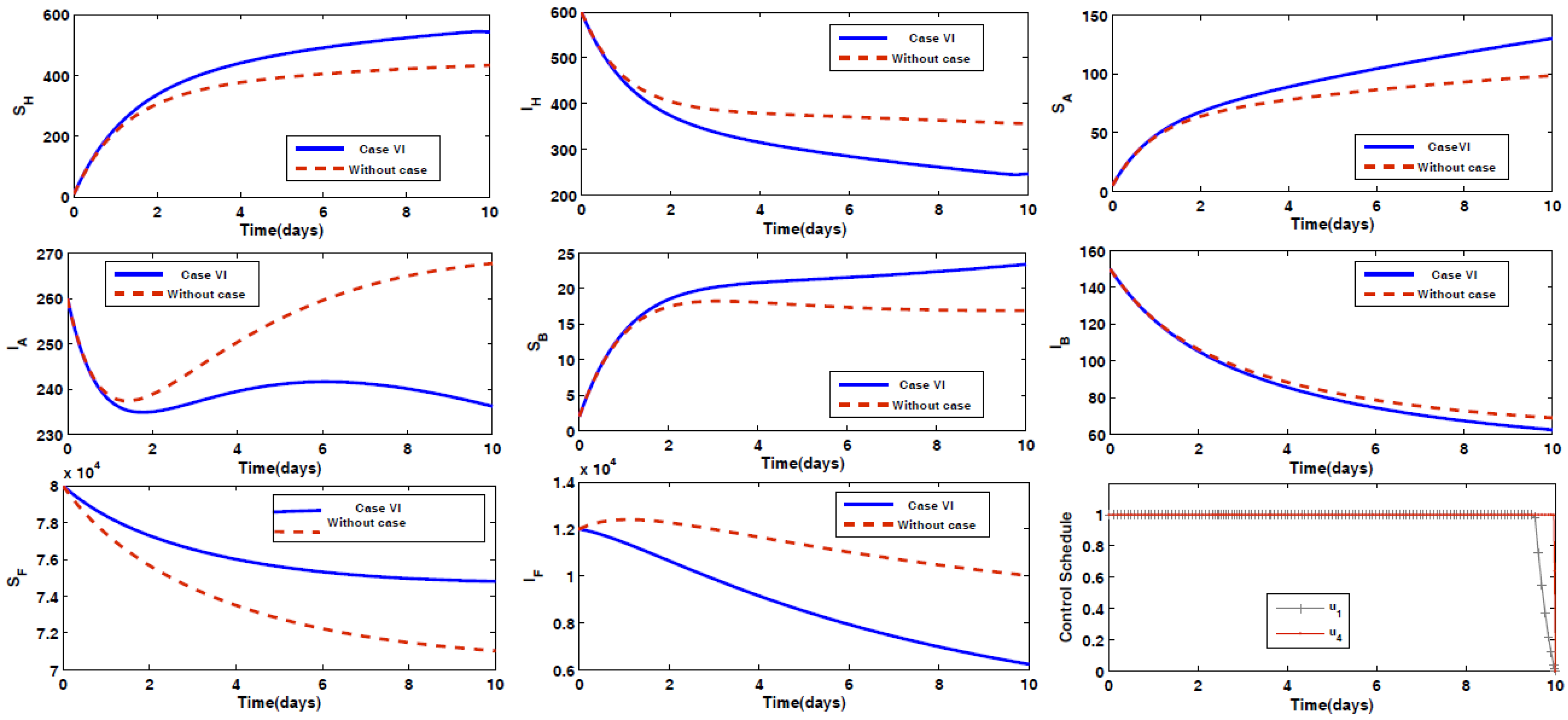
- Case VI: Prevention of humans being infected by the disease, along with spraying insecticides on the sand fly vectors.
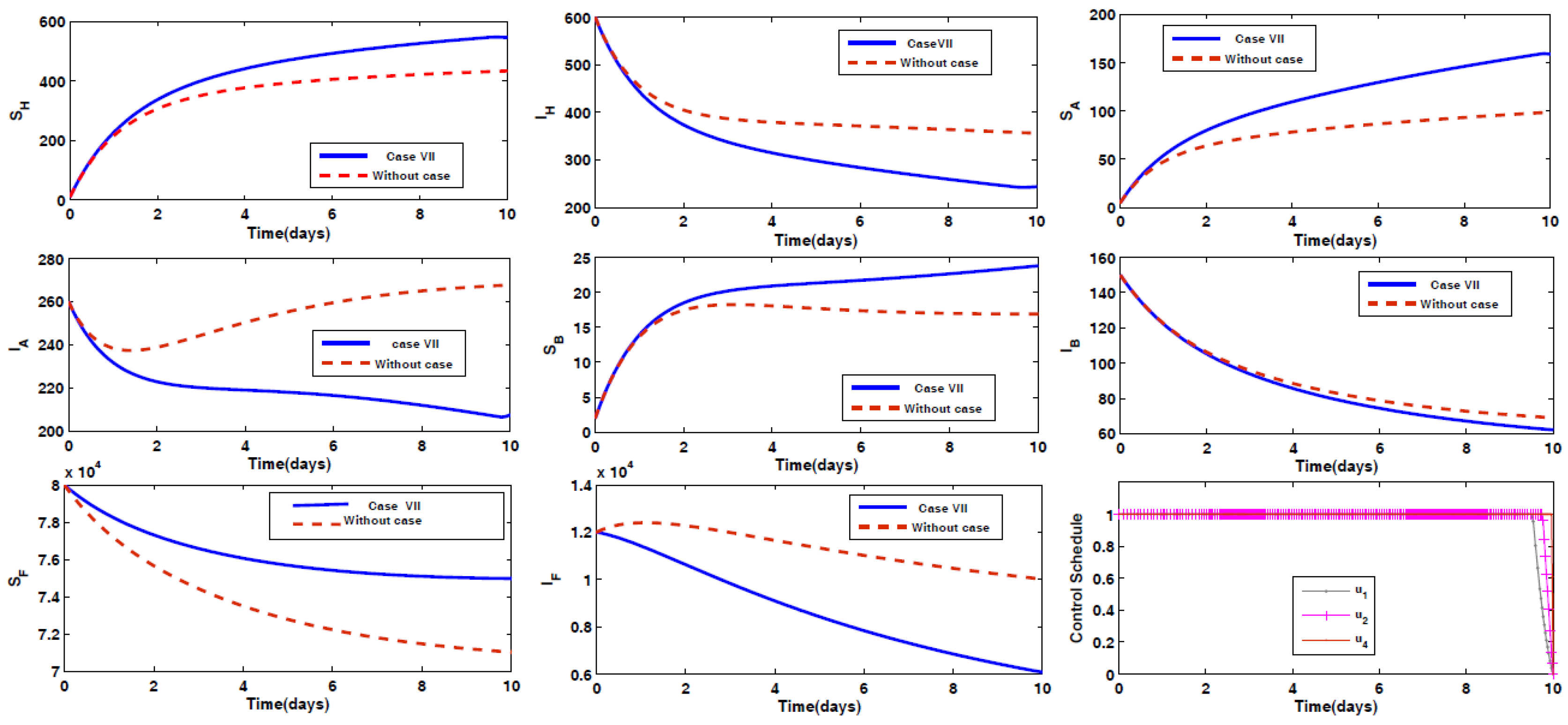
- Case VII: Prevention of humans and animal Type A being infected by the disease, along with spraying insecticides on the sand fly vectors.
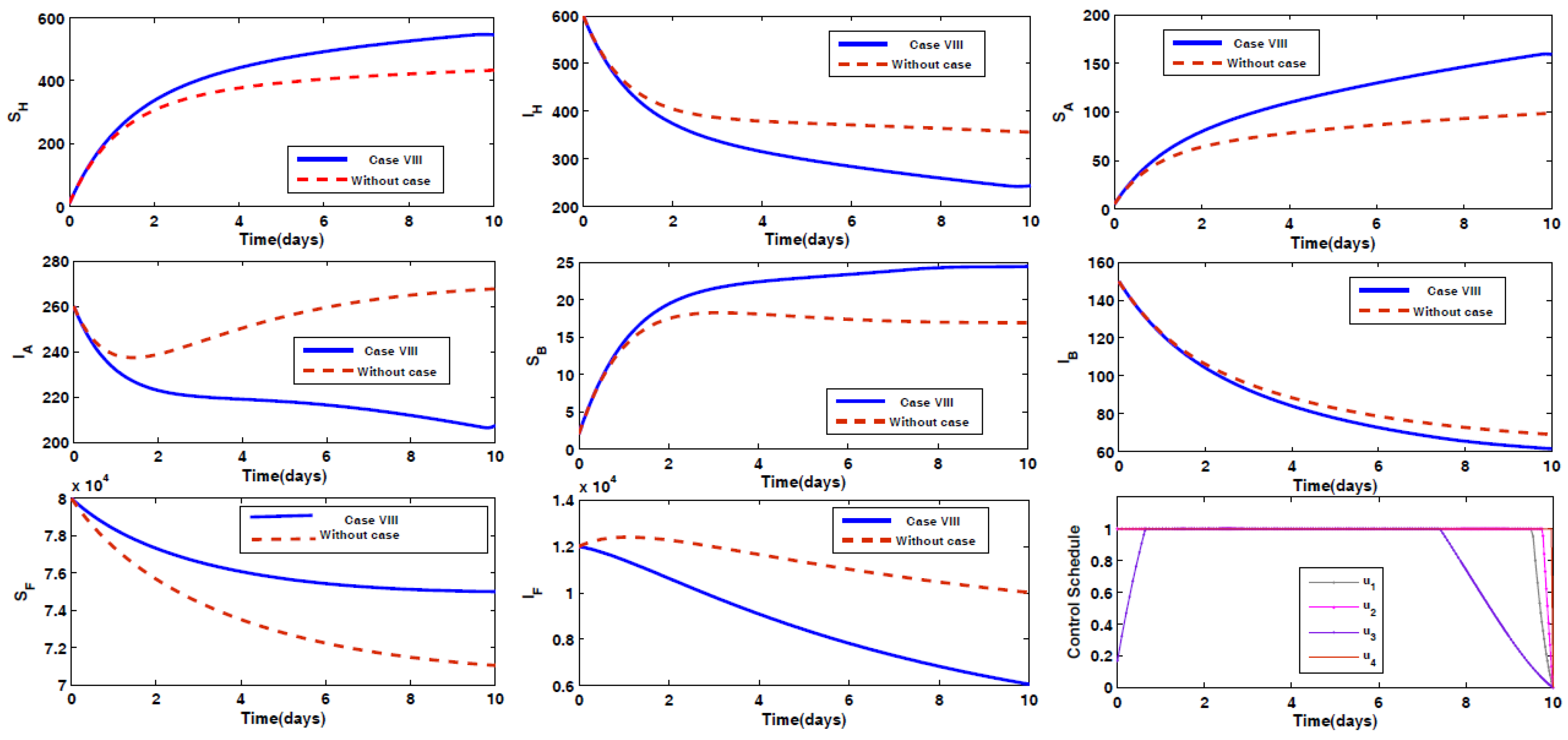
- Case VIII: Prevention of humans and animals Type A and Type B being infected by the disease, along with spraying insecticides on the sand fly vectors.
4.2. Impact of Optimal Control on the Different Cases Proposed
4.3. Rescued Population and Vector Reduction for Different Cases
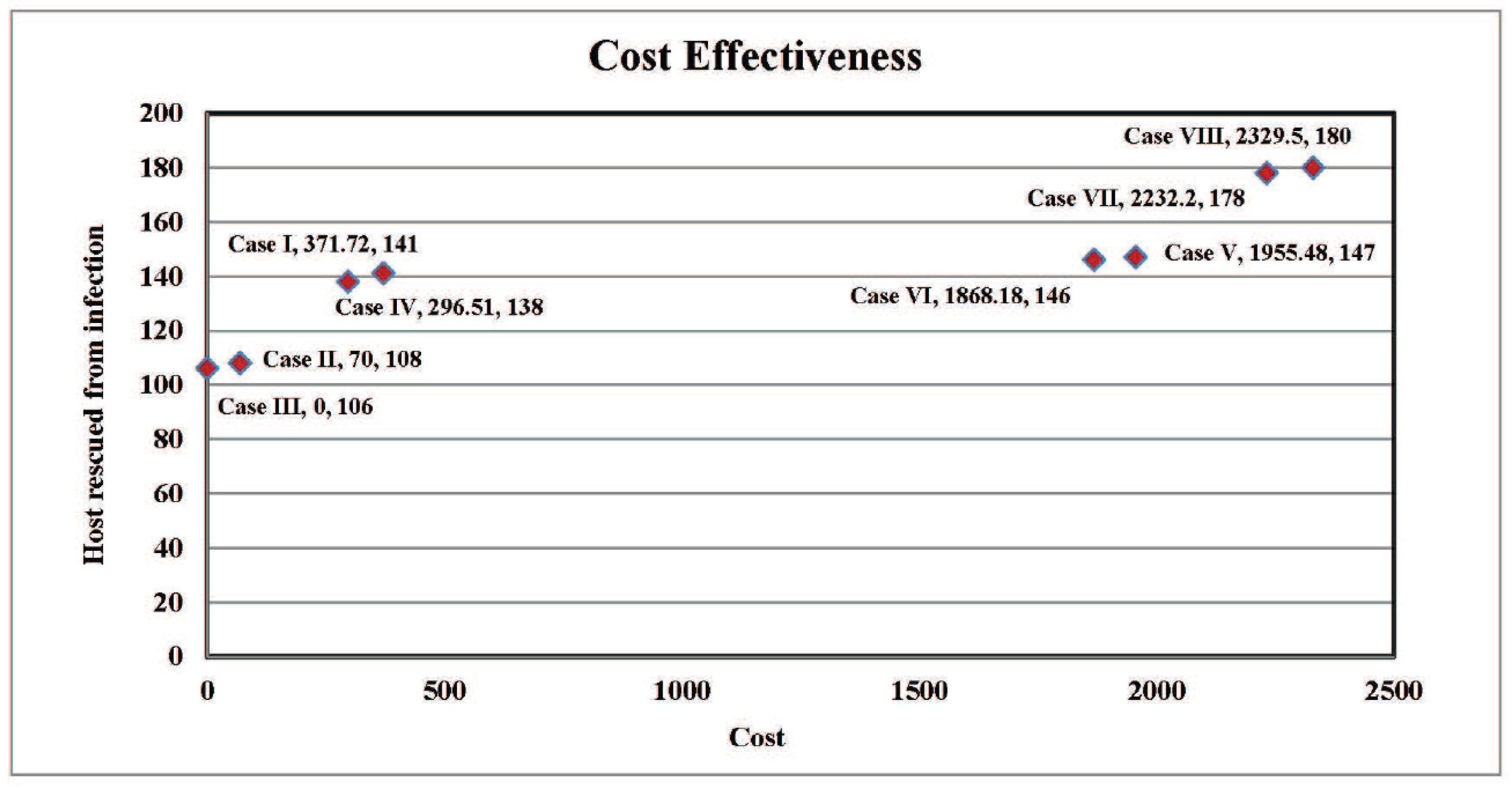
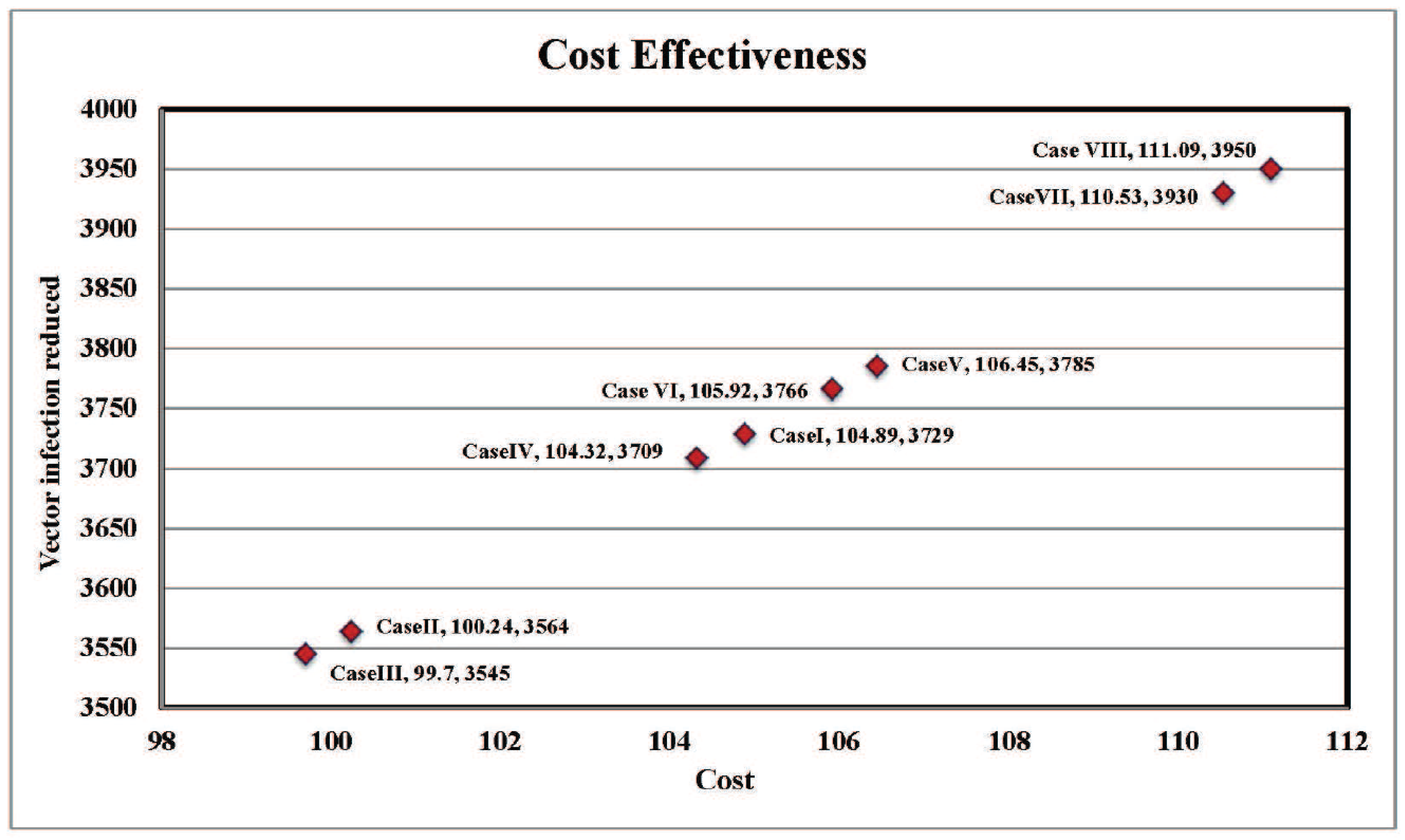
4.4. Cost-Effectiveness of the Different Cases
4.5. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A
- Case I: Optimal Use of Drug for Prevention From Disease for Animals Type A and Type B with Spray of Insecticides on Vector

- Case II: Optimal Use of Drug for Prevention of Disease for Animal Type B with Spray of Insecticides on Vector

- Case III: Optimal Use of Spray of Insecticides on the Sand Fly Vector

- Case IV: Optimal Use of Drug for Prevention from Disease for Animal Type A with Spray of Insecticides on Vector

- Case V: Optimal Use of Drug for Prevention from Disease for Human and Animal Type B with Spray of Insecticides on Vector

- Case VI: Optimal Use of Drug for Prevention from Disease for Human and Spraying of Insecticides on Vector

- Case VII: Optimal Use of Drug for Prevention from Disease for Human and Animal A with Spray of Insecticides on Vector

- Case VIII: Optimal Use of Drug for Prevention from Disease for Human, Animals Type A and Type B with Spraying of Insecticides on Vector

References
- Sharma, U.; Singh, S. Immunobiology of leishmaniasis. Indian J. Exp. Biol. 2009, 47, 412–423. [Google Scholar] [PubMed]
- Park, K. Preventive and Social Medicine; Banarsidas Bhanot Publishers: Jabalpur, India, 2005. [Google Scholar]
- Chaves, L.F.; Hernandez, M.J. Mathematical modelling of American Cutaneous Leishmaniasis: Incidental hosts and threshold conditions for infection persistence. Acta Trop. 2004, 92, 245–252. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Leishmaniasis. Fact Sheet Updated April 2017. Available online: http://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 18 July 2018).
- ELmojtaba, I.M.; Mugisha, J.Y.T.; Hashim, M.H.A. Mathematical analysis of the dynamics of visceral leishmaniasis in the Sudan. Appl. Math. Comput. 2010, 217, 2567–2578. [Google Scholar] [CrossRef]
- Kassiri, H.; Sharifinia, N.; Jalilian, M.; Shemshad, K. Epidemiological aspects of cutaneous leishmaniasis in Ilam province, west of Iran (2000–2007). Asian Pac. J. Trop. Dis. 2012, 2, S382–S386. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef]
- Kasper, D.L.; Braunwald, E.; Fauci, A.S.; Hauser, S.L.; Longo, D.L.; Jameson, J.L.; Loscalzo, J. Harrison’s Principles of Internal Medicine, 17th ed.; McGraw-Hill: New York, NY, USA, 2008; Volume 1, Chapter 1–216. [Google Scholar]
- Bathena, K. A Mathematical Model of Cutaneous Leishmaniasis. Master’s Thesis, Rochester Institute of Technology, Rochester, NY, USA, 2009. [Google Scholar]
- Miller, E.; Huppert, A. The effects of Host Diversity on Vector-Borne Disease: The Conditions under Which Diversity Will Amplify or Dilute the Disease Risk. PLoS ONE 2013, 8, e80279. [Google Scholar] [CrossRef] [PubMed]
- Jaouadi, K.; Haouas, N.; Chaara, D.; Gorcii, M.; Chargui, N.; Augot, D.; Pratlong, F.; Dedet, J.P.; Ettlijani, S.; Mezhoud, H.; et al. First detection of Leishmania killicki (Kinetoplastida, Trypanosomatidae) in Ctenodactylus gundi (Rodentia, Ctenodactylidae), a possible reservoir of human cutaneous leishmaniasis in Tunisia. Parasites Vectors 2011, 4, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.N.; Roy, P.K.; Mondal, J. Mathematical Model for Suppression of Sand Flies through IRS with DDT in Visceral Leishmaniasis. Am. J. Math. Sci. 2013, 2, 105–112. [Google Scholar]
- Wu, H.J.J.; Massad, E. Mathematical modelling for Zoonotic Visceral Leishmaniasis dynamics: A new analysis considering updated parameters and notified human Brazilian data. Infect. Dis. Model. 2017, 2, 143–160. [Google Scholar]
- Bacaer, N.; Guernaoui, S. The epidemic threshold of vector-borne diseases with seasonality the case of cutaneous leishmaniasis in Chichaoua, Morocco. J. Math. Biol. 2006, 53, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Biswas, D.; Kesh, D.K.; Datta, A.; Chatterjee, A.N.; Roy, P.K. A Mathematical Approach to Control Cutaneous Leishmaniasis Through Insecticide Spraying. Sop Trans. Appl. Math. 2014, 1, 44–54. [Google Scholar] [CrossRef]
- Biswas, D.; Roy, P.K.; Li, X.Z.; Basir, F.A.; Pal, J. Role of macrophage in the disease dynamics of cutaneous leishmaniasis: A delay induced mathematical study. Commun. Math. Biol. Neurosci. 2016, 2016, 1–31. [Google Scholar]
- Roy, P.K.; Li, X.Z.; Biswas, D.; Datta, A. Impulsive Application to Design Effective Therapies Against Cutaneous Leishmaniasis Under Mathematical Perceptive. Commun. Math. Biol. Neurosci. 2017, 2017, 1–17. [Google Scholar]
- Biswas, D.; Datta, A.; Roy, P.K. Combating Leishmaniasis through Awareness Campaigning: A Mathematical Study on Media Efficiency. Int. J. Math. Eng. Manag. Sci. 2016, 1, 139–149. [Google Scholar]
- Zamir, M.; Zaman, G.; Alshomrani, A.S. Sensitivity Analysis and Optimal Control of Anthroponotic Cutaneous Leishmania. PLoS ONE 2016, 11, e0160513. [Google Scholar] [CrossRef] [PubMed]
- Begon, M.; Bennett, M.; Bowers, R.G.; French, N.P.; Hazel, S.M.; Turner, J. A clarication of transmission terms in host-microparasite models: numbers, densities and areas. Epidemiol. Infect. 2002, 129, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, A.; Ruiz-Postigo, A.J.; Pita, J.; Lado, M.; Ben-Ismail, R.; Argaw, D.; Alvar, J. Visceral leishmaniasis outbreak in South Sudan 2009–2012: Epidemiological assessment and impact of a multisectoral response. PLoS Negl. Trop Dis. 2014, 8, e2720. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Singh, V.; Sarkar, R.R. Understanding Visceral Leishmaniasis Disease Transmission and its Control—A Study Based on Mathematical Modeling. Mathematics 2015, 3, 913–944. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Subramanian, A.; ELMojtaba, I.M.; Chattopadhyay, J.; Sarkar, R.R. Optimal combinations of control strategies and cost-effective analysis for visceral leishmaniasis disease transmission. PLoS ONE 2017, 12, e0172465. [Google Scholar] [CrossRef] [PubMed]
- Brett-Major, D.M.; Claborn, D.M. sandfly Fever: What Have We Learned in One Hundred Years? Mil. Med. 2009, 174, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.E.; Bates, P.A. Leishmania Manipulation of sandfly Feeding Behavior Results in Enhanced Transmission. PLoS Pathog. 2007, 3, e91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, L.F.; Coutinho, F.A.B.; Burattini, M.N.; Massad, E. Threshold conditions for infection persistence in complex host-vectors interactions. Comptes Rendus Biol. 2002, 325, 1073–1084. [Google Scholar] [CrossRef]
- Zaman, G.; Kang, Y.H.; Jung, I.H. Stability analysis and optimal vaccination of an SIR epidemic model. BioSystems 2008, 93, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Birkhoff, G.; Rota, G.C. Ordinary Differential Equations, 4th ed.; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Lukes, D.L. Differential Equations: Classical to Controlled in Mathematics in Science and Engineering; Academic Press: New York, NY, USA, 1982; Volume 162. [Google Scholar]
- Kirschner, D.; Lenhart, S.; Serbin, S. Optimal control of the chemotherapy of HIV. J. Math. Biol. 1997, 35, 775–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimozako, H.J.; Wu, J.; Massad, E. The Preventive Control of Zoonotic Visceral Leishmaniasis: Efficacy and Economic Evaluation. Comput. Math. Methods Med. 2017, 2017, 4797051. [Google Scholar] [CrossRef] [PubMed]
- Kaabi, B.; Ahmed, S.B. Assessing the effect of zooprophylaxis on zoonotic cutaneous leishmaniasis transmission: A system dynamics approach. Biosystems 2013, 114, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Okosuna, K.O.; Rachid, O.; Marcus, N. Optimal control strategies and cost-effectiveness analysis of a malaria model. BioSystems 2013, 111, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Sardar, T.; Mukhopadhyay, S.; Bhowmick, A.R.; Chattopadhyay, J. An optimal cost effectiveness study on Zimbabwe cholera seasonal data from 2008–2011. PLoS ONE 2013, 8, e81231. [Google Scholar] [CrossRef] [PubMed]
- Stauch, A.; Sarkar, R.R.; Picado, A.; Ostyn, B.; Sundar, S.; Rijal, S.; Boelaert, M.; Dujardin, J.; Duerr, H. Visceral Leishmaniasis in the Indian Subcontinent: Modelling Epidemiology and Control. PLoS Negl. Trop. Dis. 2011, 5, e1405. [Google Scholar] [CrossRef] [PubMed]
- Costs of Medicines in Current Use for the Treatment of Leishmaniasis (Annex6), Drug Prices (January 2010). Available online: http://www.who.int/leishmaniasis/research/978_92_4_12_949_6_Annex6.pdf (accessed on 18 July 2018).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Range | Default Value | Reference |
|---|---|---|---|---|
| Recruitment rate of human population | 300–318 | 317 | [5] | |
| Recruitment rate of sand fly population | 14,950–15,000 | 14,950 | [5] | |
| Recruitment rate of animal population of Type A | 70–150 | 73 | [5] | |
| Recruitment rate of animal population of Type B | 3–40 | 20 | [32] | |
| Death rate of human population | 0.000007–0.0001 | 0.00004 | ||
| Death rate of sand fly population | 0.188–0.795 | 0.189 | [5] | |
| Death rate of animal population of Type A | 0.06–0.21 | 0.19 | [5] | |
| Death rate of animal population of Type B | 0.089–0.255 | 0.25 | [32] | |
| Biting rate of sand fly on human | 0.15–0.29 | 0.24 | [5,9] | |
| Biting rate of sand fly on animals of Type A and Type B | 0.15–0.25 | 0.16 | [5] | |
| Transmission probability of CL in sand fly | 0.12–0.24 | 0.18 | [5,9] | |
| Transmission probability of CL on animal of Type A | 0.11–0.172 | 0.12 | [32] | |
| Transmission probability of CL on animal of Type B | 0.02–0.071428 | 0.05 | [5,9] | |
| Transmission probability of CL in sand fly from infected human | 0.11–0.25 | 0.14 | [5,9] | |
| Transmission probability of CL in sand fly from infected animal A | 0.07–0.21 | 13 | [5,9] | |
| Transmission probability of CL in sand fly from infected animal B | 0.04–0.21 | 0.12 | [32] |
| Cases | Human | Animal A | Animal B | Vector Reduction |
|---|---|---|---|---|
| Case I | 12.67 | 22.31 | 4.67 | 31.10 |
| Case II | 12 | 11.15 | 4.67 | 29.7 |
| Case III | 11.83 | 11.15 | 4.00 | 29.54 |
| Case IV | 12.50 | 12.92 | 4.00 | 30.90 |
| Case V | 18.16 | 5.17 | 4.67 | 31.54 |
| Case VI | 18 | 11.92 | 4.67 | 31.38 |
| Case VII | 18.5 | 23.07 | 4.67 | 32.75 |
| Case VIII | 18.60 | 23.07 | 5.33 | 32.92 |
| Cases | Host Reduction | Cost (in $) | Vector Reduced | Cost (in $) |
|---|---|---|---|---|
| Case I | 141 | 371.716 | 3729 | 104.878 |
| Case II | 108 | 70 | 3564 | 100.238 |
| Case III | 106 | 0 | 3545 | 99.70 |
| Case IV | 138 | 296.514 | 3709 | 104.316 |
| Case V | 147 | 1955.482 | 3785 | 106.453 |
| Case VI | 146 | 1868.184 | 3766 | 105.919 |
| Case VII | 178 | 2232.198 | 3930 | 110.531 |
| Case VIII | 180 | 2329.496 | 3950 | 111.094 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, D.; Dolai, S.; Chowdhury, J.; Roy, P.K.; Grigorieva, E.V. Cost-Effective Analysis of Control Strategies to Reduce the Prevalence of Cutaneous Leishmaniasis, Based on a Mathematical Model. Math. Comput. Appl. 2018, 23, 38. https://doi.org/10.3390/mca23030038
Biswas D, Dolai S, Chowdhury J, Roy PK, Grigorieva EV. Cost-Effective Analysis of Control Strategies to Reduce the Prevalence of Cutaneous Leishmaniasis, Based on a Mathematical Model. Mathematical and Computational Applications. 2018; 23(3):38. https://doi.org/10.3390/mca23030038
Chicago/Turabian StyleBiswas, Dibyendu, Suman Dolai, Jahangir Chowdhury, Priti K. Roy, and Ellina V. Grigorieva. 2018. "Cost-Effective Analysis of Control Strategies to Reduce the Prevalence of Cutaneous Leishmaniasis, Based on a Mathematical Model" Mathematical and Computational Applications 23, no. 3: 38. https://doi.org/10.3390/mca23030038