
Postbiotics: Functional Food Materials and Therapeutic Agents for Cancer, Diabetes, and Inflammatory Diseases

1
Department of Biotechnology, Rathinam College of Arts and Science, Eachanari, Coimbatore 641021, Tamil Nadu, India
2
Department of Stem Cell and Regenerative Biotechnology, Konkuk University, Seoul 05029, Republic of Korea
*
Authors to whom correspondence should be addressed.
Foods 2024, 13(1), 89; https://doi.org/10.3390/foods13010089
Submission received: 15 November 2023
/
Revised: 21 December 2023
/
Accepted: 22 December 2023
/
Published: 26 December 2023
(This article belongs to the Section Food Microbiology)
Abstract
:Postbiotics are (i) “soluble factors secreted by live bacteria, or released after bacterial lysis, such as enzymes, peptides, teichoic acids, peptidoglycan-derived muropeptides, polysaccharides, cell-surface proteins and organic acids”; (ii) “non-viable metabolites produced by microorganisms that exert biological effects on the hosts”; and (iii) “compounds produced by microorganisms, released from food components or microbial constituents, including non-viable cells that, when administered in adequate amounts, promote health and wellbeing”. A probiotic- and prebiotic-rich diet ensures an adequate supply of these vital nutrients. During the anaerobic fermentation of organic nutrients, such as prebiotics, postbiotics act as a benevolent bioactive molecule matrix. Postbiotics can be used as functional components in the food industry by offering a number of advantages, such as being added to foods that are harmful to probiotic survival. Postbiotic supplements have grown in popularity in the food, cosmetic, and healthcare industries because of their numerous health advantages. Their classification depends on various factors, including the type of microorganism, structural composition, and physiological functions. This review offers a succinct introduction to postbiotics while discussing their salient features and classification, production, purification, characterization, biological functions, and applications in the food industry. Furthermore, their therapeutic mechanisms as antibacterial, antiviral, antioxidant, anticancer, anti-diabetic, and anti-inflammatory agents are elucidated.
1. Introduction
Food stands as a fundamental human requirement crucial for sustaining all aspects of life. Nutrients such as lipids, carbohydrates, proteins, fibers, phytochemicals, antioxidants, vitamins, probiotics, and prebiotics provide different health benefits [1,2,3,4]. Whole grains and dietary fiber serve as rich sources of vitamins, minerals, and slowly digestible energy. Intense innovation in the functional food industry has led to the production of a vast spectrum of health-promoting bioactive components, including prebiotics, phytochemicals, natural antioxidants, and bioactive peptides [5]. Emerging food technologies are actively used to process food products, resulting in increased probiotic functionality, prebiotic stability, and higher concentrations of bioactive compounds [6].
Probiotics, as defined by the Food and Agriculture Organization of the United Nations–World Health Organization (FAO-WHO), are “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [7]. The majority of probiotic products contain a defined and constrained list of microbial taxa, primarily lactic acid bacteria (LAB) like Lactobacillus spp.and Bifidobacterium spp., recognized as generally safe (GRAS) [7]. While yogurt is a common carrier of probiotics, other fermented (cheese and yogurt) and non-fermented (cereal, chocolate bars, fruit juices, and smoothies) meals may also incorporate probiotics. Probiotics impact the gut microbiota by suppressing infections, inhibiting their adherence to tissues, and impeding their establishment in the gut [8,9]. Furthermore, probiotics contribute to immune system development, the production of essential nutrients, and the reinforcement of intestinal barrier integrity by activating genes linked to tight junction signaling [9].
According to a consensus statement by the International Scientific Association of Probiotics and Prebiotics, “Prebiotics is a substrate that is specifically utilized by host microorganisms to provide a health advantage”. Prebiotics, found in dietary fibers, phenolics, phytochemicals, conjugated linoleic acids, polyunsaturated fatty acids, human milk oligosaccharides (HMOs), and various oligosaccharides, can alter the composition of the microbiota by promoting the growth of specific species, thereby benefiting the host’s health [10,11]. A synbiotic, commonly defined as a synergistic mixture of probiotics and prebiotics, beneficially affects the host by improving the survival and colonization of live beneficial microorganisms in the gastrointestinal tract [12,13].
Postbiotics are “soluble factors secreted by live bacteria, or released after bacterial lysis, such as enzymes, peptides, teichoic acids, peptidoglycan-derived muropeptides, polysaccharides, cell-surface proteins and organic acids”; (ii) “non-viable metabolites produced by microorganisms that exert biological effects on the hosts”; and (iii) “compounds produced by microorganisms, released from food components or microbial constituents, including non-viable cells that, when administered in adequate amounts, promote health and wellbeing”. These molecules mediate beneficial biological activities directly or indirectly when administered to consumers [14,15,16,17,18]. Described as “preparations of inanimate microorganisms and/or their components that confer a health benefit to the host”, postbiotics are produced by bacterial and fungal species like Lactobacillus, Bifidobacterium, Streptococcus, Eubacterium, Faecalibacterium, and Saccharomyces, which are naturally found in fermented foods such as yogurt, sauerkraut, pickled vegetables, and kombucha [19,20,21]. Several commercial postbiotics, primarily used to treat gastrointestinal or immune-related disorders, are available as supplements or in food matrices [22]. Comprising bacterial lysates with cell surface proteins, bacterial enzymes, peptides, metabolites, and lower organic acids like lactic acid, postbiotics derive their efficacy from microbial metabolites, proteins, lipids, carbohydrates, vitamins, organic acids, cell wall components, and other complex compounds produced in the fermented matrix [20,23]. Food processing techniques, including heat, sonication, irradiation, and high pressure, can impact postbiotic composition, with potential effects on the microorganisms involved in fermentation [24,25,26,27,28,29,30,31,32,33,34]. Various analytical techniques such as matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry and HPLC are suggested for postbiotic identification [35,36,37]. Studies have assessed the bioactivity and health effects of postbiotics, including intracellular metabolites and cell wall components, both in vitro and in vivo [38]. Dairy-based items, like kefir, kombucha, yogurt, and pickled vegetables, naturally contain postbiotics [39].
Producer strains, primarily Lactobacillus and Bifidobacterium varieties, and other species like Streptococcus, Akkermansia muciniphila, Eubacterium hallii, Faecalibacterium, and Saccharomyces boulardii, can be used to extract postbiotics in situ. Milk products containing exopolysaccharides (EPSs) made by lactic acid bacteria (LAB), particularly Lacticaseibacillus rhamnosus, enhance the physicochemical and sensory properties of food-based products [20,40]. Streptococcus and Faecalibacterium species are also potential sources of postbiotics [14,23]. Postbiotic supplements, primarily produced by Lactobacillus and Bifidobacterium spp., have been shown to lower blood pressure, indicating their potential in treating hypertension [38].
As highlighted by Klemashevich [41], the intestinal microbiota influences various gastrointestinal tract processes, including inflammation, pathogen defense, and immune system development. A study on Lactobacillus helveticus MB2-1 demonstrated the potent capacity of the crude culture extract and pure EPS to scavenge three different types of free radicals and chelate ferrous ions [42]. This review offers a concise history and explanation of postbiotics, covering their classification, manufacturing, purification, and characterization, along with insights into their biological activities and potential applications in the food industry.Furthermore, therapeutic mechanisms of postbiotics as antibacterial, antiviral, antioxidant, anticancer, anti-diabetic, and anti-inflammatory agents are discussed.
2. History and Concept of Postbiotics
More than 2000 years ago, Hippocrates asserted that “all disease begins in the gut”. Today, there is no doubt that the intestinal microbiome plays a critical role in the pathogenesis of various systemic autoimmune and inflammatory disorders [43]. The emergence of studies on intestinal flora was initiated by the Russian microbiologist Elie Metchnikoff, who began working with Louis Pasteur at the Pasteur Institute in Paris in the early 1900s. Louis Pasteur (1822–1895) made significant advancements in pasteurization, microbial fermentation, and vaccination, and Louis Pasteur was the inventor of these procedures. During his lifetime, he developed treatments for several of the deadliest diseases worldwide, including rabies, anthrax, tuberculosis, cholera, and smallpox. It has been suggested that discovering treatments for cancer, heart disease, diabetes, and Alzheimer’s disease would be the modern-day equivalent of his accomplishments. In addition to Pasteur’s remarkable achievements, he contributed to the germ theory of disease by popularizing the idea that germs cause disease. Consequently, antibiotic medications were developed, leading to a century of bacterial fear. Over the past several decades, the overuse of antibiotics has damaged the immune system, destroyed the microbiome, and given rise to infectious diseases with lethal “superbug” antibiotic resistance.
In the early 1900s, Metchnikoff developed a keen interest in understanding how and why people age while working at the Pasteur Institute. During the early 1900s, Bulgaria boasted an extraordinarily long average lifetime of 87 years, with four out of every 1000 people over 100 years old, according to figures collected during Metchnikoff’s study of 36 nations. Metchnikoff hypothesized that “bad” bacteria in the intestinal tract, producing toxins, were responsible for the aging process. He attributed this to what he termed “intestinal auto-intoxication”, leading to age-related bodily deterioration and breakdown [44]. Metchnikoff ultimately developed a profound intuitive understanding that earned him the title of the “Founding Father of Probiotics”. According to Metchnikoff, the daily consumption of fermented milk products, such as yogurt and kefir, accounted for the long and healthy lives of Bulgarians. He recognized that lactic acid produced by bacteria in milk during fermentation created an acidic environment in the gastrointestinal tract (GI tract). Metchnikoff postulated that this environment prevented the formation of “bad” microorganisms, resulting in less “intestinal auto-intoxication”, improving health, and extending lifespan (Figure 1).
The current state of microbiome science has significantly benefited from the development of postbiotics. The collection of microorganisms, including bacteria, fungi, and viruses, residing inside and on our bodies is known as the “human microbiome”. The first successful sequencing of the human genome was undertaken by the Human Genome Project, with a budget of USD3 billion over 13 years (1990–2003). This paved the way for cures for many chronic degenerative diseases. However, the Human Genome Project also led to technology allowing researchers to sequence genomes rapidly and affordably. The Human Microbiome Project, resulting in the publication of over 350 articles, is considered the “birth” of the modern age of microbiome science.
Our ancestors recognized the vital role fermented foods and bacteria play in health, especially gut health [45]. The health of a host is significantly influenced by the gastrointestinal microbiota (GM) due to the protective roles of microorganisms [46]. Microbial dysbiosis, or an imbalance in microbial populations, has been linked to various human disorders, such as obesity, diabetes, colon cancer, inflammatory bowel diseases, neurological issues, psychiatric difficulties, and allergies [47,48,49,50,51]. The GM seems to contribute to the onset and course of disease in these circumstances [52,53,54]. Probiotics and prebiotics play a crucial role in repairing intestinal microbial habitats and thus benefiting health [55,56]. Numerous studies have shown that strategies for altering GM composition or activity, such as supplementation with these substances, are highly effective. Most studies on GM modulators have focused on probiotics and prebiotics. Living bacteria, known as probiotics, support host health by improving intestinal barrier function, infection defense, and immune response modulation [9,11]. Among bacteria, species from the genera Lactobacillus, Bifidobacterium, Streptococcus, and Lactococcus, as well as yeasts from the genus Saccharomyces, are most frequently used [57,58,59]. Postbiotics are live microorganisms that, when administered to a host in sufficient quantities, impart health benefits [60]. Prebiotics are substrates specifically utilized by host microbes and confer health benefits to the host [11]. A symbiotic is defined as a combination of live microorganisms and substrates that are utilized by host microorganisms only when advantageous to host health [61]. Until recently, the term mostly referred to indigestible fibers, but its modern meaning now encompasses bioactive substances from various sources, such as polyunsaturated fatty acids and polyphenols.
Technological advances have enabled the examination, categorization, and characterization of different probiotic-related concepts. The International Scientific Association for Probiotics and Prebiotics (ISAPP), among others, has established an expert consensus and aids in the development of novel concepts. In 2019, ISAPP assembled a panel of specialists to evaluate the definition and use of postbiotics [62,63]. These specialists, with expertise in nutrition, microbial physiology, gastroenterology, pediatrics, food science, and microbiology, defined postbiotics as “the preparation of inanimate microorganisms and/or their components that confer a health benefit on the host”. Effective postbiotics must inactivate microbial cells or cell components, with or without metabolites, to contribute to claimed health benefits [62]. Postbiotics, referred to as “biogenics” by Mitsuoka in 1998, have been accessible in Japan for over a century. It is essential to highlight the most recent changes in the ISAPP consensus statement regarding the definition and scope of postbiotics, in addition to new biotic terminology [18]. The ISAPP defined postbiotics as (i) “soluble factors secreted by live bacteria, or released after bacterial lysis, such as enzymes, peptides, teichoic acids, peptidoglycan-derived muropeptides, polysaccharides, cell-surface proteins and organic acids”; (ii) “non-viable metabolites produced by microorganisms that exert biological effects on the hosts”; and (iii) “compounds produced by microorganisms, released from food components or microbial constituents, including non-viable cells that, when administered in adequate amounts, promote health and wellbeing”.
The concept of postbiotics is relatively recent compared with that of prebiotics and probiotics. Postbiotic supplements, despite not being widely available, are superior to probiotics. This is due to their purity, ease of preparation, long shelf life, large-scale production feasibility, precise actions, and the ability to trigger more focused responses through specific ligand–receptor interactions [64].
Experimental studies suggest that microbial components, distinct from their live counterparts (probiotics), are desirable subjects for investigation. Postbiotics, a group that has recently garnered much attention, exhibit therapeutic effects not restricted to the intestine alone. These effects should be assessed or proven in humans, animals, and other target organs. The panel concluded that postbiotics are intentionally inactivated bacteria containing metabolites or cellular components with positive health effects. Additionally, the target population, including people, pets, and cattle, must derive local and systemic health advantages. The injection of postbiotics onto the host surface appears to be safe and favorable. Postbiotics were defined to exclude pure microbial metabolites and vaccines; however, probiotics are not always a source of antibiotics.
Postbiotics have demonstrated positive immunomodulatory, antibacterial, and anticancer effects, including lowering blood pressure, cholesterol levels, proliferative qualities, inflammation, oxidative stress, and body weight. Healthy individuals tolerate postbiotics well. However, some individuals refrain from consuming probiotic-rich meals to elevate their postbiotic levels, such as those who have recently undergone surgery, those with structural heart abnormalities, those with digestive tract illnesses, pregnant women, and young children. Due to their weakened immune systems, these groups may be more susceptible to negative reactions. Several foods, including high-fiber meals such as oats, flaxseed, and garlic, as well as buttermilk, cottage cheese, fermented pickles, and yogurt, may aid in the gut’s postbiotics. Numerous studies have reported that postbiotic performance may be influenced by both internal and external factors. Interactions between active postbiotic metabolites and internal elements, such as existing microbiota, enzymes, and different dietary components, can inhibit the ability of metabolites to function [65]. Proteolytic enzymes, including pepsin, trypsin, and chymotrypsin, have been associated with postbiotic dysfunction [66,67,68]. Temperature is another external factor influencing the antibacterial activity of postbiotics. According to Mirnejad [69], heat treatment for 30 min at 121 °C reduces the antibacterial efficacy of postbiotics. Therefore, maintaining consistent temperature and pH is essential for the formation of postbiotics.
3. Characteristic Features of Postbiotics
According to ISAPP, a postbiotic is defined as (i) “soluble factors secreted by live bacteria, or released after bacterial lysis, such as enzymes, peptides, teichoic acids, peptidoglycan-derived muropeptides, polysaccharides, cell-surface proteins and organic acids”; (ii) “non-viable metabolites produced by microorganisms that exert biological effects on the hosts”; and (iii) “compounds produced by microorganisms, released from food components or microbial constituents, including non-viable cells that, when administered in adequate amounts, promote health and wellbeing” [11]. Postbiotics are compounds formed after microbes have been rendered inert, dead, or inactivated. In contrast, postbiotics are probiotic-derived products made from food-grade bacteria that, when taken in sufficient doses, have positive effects on health [70]. They consist of cellular components and secreted substances, metabolic waste released by living microorganisms, or are gathered and extracted during cell lysis [16]. The bacteria that constitute a postbiotic can be whole, inert, or broken down into their structural components, such as cell walls. Many postbiotic preparations contain chemicals produced by microbes, such as metabolites, proteins, and peptides. These substances may provide a postbiotic’s overall health benefits but are not required. A postbiotic must originate from a specific microorganism or a combination of microorganisms whose genomic sequences are known and can be created using a well-defined technological process of biomass generation and inactivation that is repeatable [71]. Although postbiotics of fungal origin are also being studied, the majority are formed from bacteria, most frequently Lactobacilli and Bifidobacterium members. Currently, some commercial postbiotics are available as supplements or included in food matrices, most of which are used to treat gastrointestinal or immune system-related disorders [18,20]. Lipoteichoic acid extracted from L. plantarum significantly attenuates Shigella flexneri PGN (flexPGN)-induced pro-inflammatory signals in human monocytic THP-1 cells [72]. Oral administration of Faecalibacterium prausnitzii supernatant decreased the severity of 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced colitis in mice and corrected the dysbiosis associated with TNBS colitis, highlighting its potent anti-inflammatory effects [33]. Saccharomyces cerevisiae is a well-known probiotic-producing yeast used for the bioproduction of metabolites and serves as an anti-inflammatory component [73]. Human polymorphonuclear (PMN) cells treated with cell wall fragments and metabolites obtained from Bacillus coagulans inhibited oxidative stress-induced reactive oxygen species (ROS) formation [73].
Postbiotics have several noteworthy characteristics, such as (i) being relatively safe, (ii) being well-tolerated and related to a decreased risk of adverse effects in vulnerable persons [14,25,74,75], and (iii) being quite safe. According to Zókiewicz et al. [76], they do not carry the risk of spreading antibiotic-resistant genes to commensal or pathogenic bacteria, and (iv) their efficacy is independent of cell viability, resulting in increased stability and shelf life. (v) They demonstrate straightforward industrial (large-scale) production [64]; (vi) they exhibit intriguing technological characteristics, such as the rheological properties of exopolysaccharides (EPSs) used as stabilizers in the food industry [64,77], or the bio-preservative effects of LAB bacteriocins; (vii) they have a broad range of health-promoting effects; and (viii) they include antifungal and antibacterial agents [78] (Figure 2). Metabolomic techniques are widely used to evaluate the metabolic products of bacteria in feces and serum [79]. Many commercial pharmaceutical products, including cytoflora (components of Lactobacillus and Bifidobacterium sp.) and lacteol (inactivated Lactobacillus sps.), Nyaditum resea (inactivated mycobacteria), and others have already been approved for use as dietary supplements, immunomodulators, and instruments to lower the risk of infections [80]. Food ingredients such as immuno-biotics and post-immunobiotics can influence the immune response in two ways: augmentation or inhibition [81].
4. Production, Purification, and Characterization of Postbiotics
Billions of bacteria, containing more than three million genes, reside in the human gastrointestinal tract (GI), with the large intestine housing the most metabolically active and varied microbial community [82]. According to Tanaka and Nakayama [83,84], the human microbiota, which is specific to each individual, begins to appear in the uterus when the fetus consumes amniotic fluid and continues even after delivery. It is essential for human growth and contains more than 1000 different microbial species [27]. An individual’s gut microbiome contains both beneficial and harmful bacteria and reflects familial inheritance. The balance of both bacterial communities is essential; an imbalance disrupts this regular microflora (dysbiosis), which not only affects the GI tract but also has a severe effect on the operation of other organs. According to Carding et al. [85], this increases the risk of a wide range of infections and chronic disorders, including obesity, autism, psychological abnormalities, gastroenteritis, colon dysfunction, and irritable bowel syndrome. Several studies have demonstrated that this balance can be restored using postbiotics, which are less risky than prebiotics and probiotics [86]. Numerous studies have suggested that the beneficial effects of probiotics on human health are not always related to bacterial viability. A significant portion of the health advantages of pre-, pro-, and synbiotics appears to be mediated by different metabolic products, cellular and subcellular structural elements, and intact or ruptured dead microorganisms. Teichoic acid, short-chain fatty acids, vitamins, enzymes, exopolysaccharides, peptides, amino acids, and fermentation byproducts are among the postbiotics, which are structural and metabolic microbial products [16]. Probiotics produce these postbiotic components when they feed on prebiotics during lengthy storage or processing such as pasteurization, baking, or metabolic processes.
Numerous yeasts and bacteria have been utilized as probiotics; however, their postbiotic usage has not been explored. Postbiotic studies have recently attracted immense interest because postbiotics offer a secure substitute for live probiotics. Many functional foods, including the probiotic yeast Saccharomyces cerevisiae var. Boular and functional food products such as grains and fruit juices, have increased shelf stability, sensory characteristics, safety, and health benefits [87]. Modern technologies are used to extract, characterize, and examine the bioactivities of various postbiotic components in preparation for potential therapeutic applications in medicine [88]. Generally, postbiotics are stable and do not require cold chains for industrial use, whereas viable probiotic products must be stored and transported in cold storage facilities. There is no postbiotic interaction with the food matrix, no possibility of developing antibiotic-resistance genes, and no unacceptable taste or odor modifications. Therefore, postbiotics are a secure replacement for patients with immunological weaknesses following transplantation or in babies. Through the communication axes between the gut and target organs, such as the gut–brain, gut–lung, and gut–liver axes, postbiotics affect other organs, both locally and systemically [64].
The bacterial strain, culture medium, and how the bacteria are treated after they have multiplied are important factors that affect the type and quantity of postbiotic products. Food postbiotics are soluble components, such as commodities or metabolic byproducts, generated in a medium during bacterial growth and do not undergo any post-propagation processing [76]. Bacteria may occasionally be subjected to lysis after multiplication using cell fragmentation techniques, including thermal, enzymatic, chemical, sonication, hyperbaric, solvent extraction, or a combination of these [22,89]. These mechanisms add various extra intracellular metabolites and components made from the cell walls to the postbiotic mixture, enriching it and providing the ensuing postbiotics with new functions. The term “postbiotics” refers to a wide range of substances, such as extracellular vesicles (EVs), bacteriocins, enzymes, proteins, peptides, organic acids, vitamins, and other discharged compounds like EPS, cell wall components, polymers, teichoic acids, peptidoglycans, peptidoglycan-derived muropeptides, pili-type forms, cell surface fractions, cell-free extracts and lysates, culture supernatants, or biosurfactants and also the definition of postbiotics as described above [14,18,25,64,74,75,76,77]
Additional extraction and purification techniques such as centrifugation, dialysis, lyophilization, and column purification have been used on the resulting solutions to separate bacterial cells from postbiotic metabolites in both treated and untreated postbiotic mixtures [22,24]. The microorganisms used as starting points and the inactivation methods or techniques used for their production are generally used to characterize postbiotics because each process affects the quality and quantity of the final postbiotics produced and produces different postbiotics with different effects [27]. When prebiotics are consumed, stored, or processed for an extended period, such as during pasteurization or baking, or when metabolized, probiotics produce these postbiotic components. According to Aggarwal et al. [27], these postbiotics can be created using lab-utilizing techniques, including radiation (UV/ionizing), high pressure, high temperature, sonication, and formalin inactivation (Figure 3). Given the complexity of biological compounds with varying degrees of polymerization and glycosidic bonds, qualitative and quantitative analyses of postbiotics typically require complicated equipment and numerous concentration/purification steps using well-known techniques, such as chromatography, spectroscopy, NMR, Fourier transform infrared absorption spectroscopy, and spectrophotometry [19].
In food, complex microbial cultures in the intestine, or as a result of cell lysis, food-grade bacteria can emit postbiotics. Following the removal of the supernatant, the effects may be immediately examined or specific chemicals may be isolated for further study [16,20,22]. According to the analytical goals and type of characterization required, a suitable technique is commonly selected [19]. Various techniques have been developed and used to evaluate postbiotics qualitatively and quantitatively. Depending on the analytical objectives and qualitative and/or quantitative properties of microbial metabolite complexes, many analytical approaches are currently being used to identify postbiotic metabolites [22]. For the purpose of determining the qualitative and/or quantitative analysis and composition of postbiotics, a number of analytical techniques, including gas chromatography (GC), high-performance liquid chromatography (HPLC), thin-layer chromatography (TLC), and spectroscopic methods, have been reviewed in detail [68,90,91,92]. Owing to its high efficiency, resolution, sensitivity, accuracy, and low solvent usage, ultra-performance liquid chromatography offers superior postbiotic separation and identification capabilities [93,94,95]. Thin-layer chromatography has been used to determine whether postbiotics contain various compounds [96,97]. Metabolites in LAB postbiotics were quantified using colorimetric methods [98]. Lin et al.used nuclear magnetic resonance spectroscopy to understand the interactions between postbiotic biological metabolites [99,100]. Headspace solid-phase microextraction GC-MS was used to characterize sixty-two compounds in the volatile profiles of postbiotics from Lactobacillus casei. Additionally, the short-chain fatty acid content of the postbiotics from four different bacterial strains was studied using GC. HPLC is one of the most frequently used analytical techniques for postbiotic analyses, both quantitatively and qualitatively [101]. In a study by Li et al. [102], postbiotics from Lactobacillus plantarum were analyzed using the Fourier transform infrared spectroscopy method (FTIR) (Figure 3).
5. Classification of Postbiotics
The classification of postbiotics depends on various factors, including the type of microorganism, structural composition, and physiological functions. Various postbiotic compounds produced by extracellular and intracellular probiotic bacteria have also been identified. For example, muropeptides are derived from peptidoglycans, exopolysaccharides (EPSs), teichoic acids, surface-protruding molecules such as fimbriae, pili, or flagella that make up cell wall components, secreted proteins/peptides, bacteriocins such as acidophilin, reuterin, and bifidin, cell-free supernatants, organic acids, neurotransmitters, and biosurfactants [64,103]. Owing to their unique physical, chemical, and functional properties, postbiotics are classified into different types, including inactivated and dead probiotics, peptidoglycans, teichoic acids, exopolysaccharides, cell-free supernatants, short-chain fatty acids, bacteriocins, enzymes, and vitamins.
5.1. Inactivated and Dead Probiotics
Although other techniques, such as gamma or UV radiation, tyndallization, sonication, and chemical treatment, are used for the preparation of postbiotics, heat is the most frequently used method for the production of inactivated or dead probiotics. This inactivation process causes differences in the cellular makeup and biological functions [25]. According to studies conducted on experimental models, the biological characteristics of their viable counterparts, such as the ability to scavenge oxygen radicals, reduce inflammatory indicators, and modify host physiology, are retained by nonviable cells [104]. Recent studies on eight different strains of Lactobacillus reuteri suggest that both live and heat-killed cells of these bacteria adhered to caco 2 cell cultures and prevented enteropathogens such as E. coli, Salmonella typhi, Listeria monocytogenes, and Enterococcus faecalis from adhering to them [105].
5.2. Cell-Free Supernatants/Suspensions
Cell-free supernatants (CFSs) are a broad category of biomolecular and active metabolites with low or high molecular weights, such as organic acids, diacetylene, carbon dioxide, and bacteriocin-like substances, which are typically secreted by lactic acid bacteria and yeasts and may help maintain homeostasis in the body [106,107]. The composition of a medium can influence CFS composition. Cell-free supernatants (CFSs) are fluids that include nutrients from the growth medium that are not absorbed by microbes as well as any metabolites left over from microbial development. CFS, which is produced when microbes are fermented, has antibacterial, antibiofilm, anti-inflammatory, antioxidant, and anticancer activities and is used to treat diarrhea [108]. Generally, CFS is obtained from safe bacteria, and the bioactive material can be used as an alternative to common antimicrobials. The metabolites can be isolated from microbial cells by centrifugation and are highly abundant in anti-inflammatory, anticancer, antioxidant, phenolic, and flavonoid chemicals. These metabolites potentially increase the expression of anti-inflammatory cytokines like IL-10 and suppress pro-inflammatory cytokines like TNF α and IL-1β. Owing to the presence of organic acids, proteinaceous compounds, and fatty acids, the CFS generated by LAB may have an antimicrobial effect. Lactic and acetic acids, together with other substances, are principally responsible for the antibacterial action of LAB [109]. The anti-proliferative effects of a cell-free culture filtrate from Lactobacillus fermentum were also reported by Lee [110], who examined the anticancer capabilities of this substance. They used 3D spheroid cultures of colorectal cancer (CRC) cells as a model for their research. According to another study, cell-free Lactobacillus reuterine supernatant, which is likely to contain carbohydrates and fatty acid metabolites, has the potential to be used for the prevention and treatment of dental caries and periodontal diseases.
Hamad reported the antibacterial ability of culture suspensions produced from four probiotic strains, including L. rhamnosus, L. fermentum, L. delbrueckii subsp. lactis, and Pediococcus acidilactici, against Clostridium perfringens [111]. The growth of Staphylococcus aureus, E. coli, Aspergillus niger, and Aspergillus flavus is significantly suppressed by lactic acid, hydrogen peroxide, protein, and diacetyl generated from Lactobacillus and Pediococcus species culture filtrate [112]. The mechanism of inhibition appears to involve the creation of pores in cell membranes and cell lysis caused by lactic acid bacteria-producing bacteriocins, followed by the actions of diacetyl and bacteriocins. Lantibiotics (class I) are among the pore-forming peptides that are produced by lactic acid bacteria. These peptides typically form unstable pores and exhibit a wide range of activity. The majority of bacteriocins have interactions with anionic lipids, which are widely found in Gram-positive bacteria membranes. “Docking molecules” have the potential to improve the conductivity and stability of lantibiotic pores; “wedge-like” pores may be formed by antibiotics; and “carpet” or “barrel stave” pores may be formed by class II bacteriocins, which may increase membrane permeability [113].Hydrogen peroxide, fatty acids, secreted proteins, and organic acids have been detected in the culture suspension of the dental health probiotic Weissella cibaria strain CMU. Organic acids, secreted proteins, and hydrogen peroxide have all been shown to exert antibacterial activities against periodontal pathogens by disrupting cell membranes, lowering the pH of the cytosol, producing hydroxyl radicals, and interfering with cellular metabolic functions [94]. As with several biomolecules, CFS seems to have superior biological effects on host health compared with pure biomolecules [114]. Pyrrolo [1,2-a] pyrazine-1,4-dione has been observed in the CSF of several examined species of lactobacilli using GC-MS analysis. Strain-specific substances such as butyric acid, benzoic acid, biosurfactants (laurostearic acid), different peptides, fatty acids, ethanol, phenol, cyclopentanes, esters, and aldehydes are also present in strain-specific ways. Many of these substances exhibited antioxidant, biofilm removal, and antagonistic activities against L. monocytogenes, indicating their potential application as food additives, particularly L. salivarius [15]. The CFS antibacterial activity of Enterococcus faecalis was found to be thermostable and peaked at a neutral pH of 7.0, supporting its use in food preservation [115]. CFS is produced in various cultures, and bacterial strains exhibit differential functions. CFS derived from L. acidophillus and L. casei has antioxidant and anti-inflammatory effects [86]. Lactobacillus and Bifidobacterium also exert antibacterial activities by inhibiting E. coli strains [116]. It has been postulated that the antioxidant capacity of diverse intracellular fractions formed from Lactobacillus strains mediates an increase in cellular glutathione concentration, which is a significant non-enzymatic antioxidant essential for maintaining the intracellular redox state. However, these non-enzymatic postbiotic antioxidant properties may also have scavenging effects on ROS and reactive nitrogen species [26,117,118].
5.3. Cell Wall Fragments
The cell wall contains various components, including teichoic and lipoteichoic acids. Among the immunogenic components of bacterial cell walls, teichoic acids, lipoteichoic acids, and other compounds can elicit an immune response [119]. The cell wall of Gram-positive bacteria is mostly composed of lipoteichoic and teichoic acids, which account for approximately 60% of the cell wall mass [120]. Different lipoteichoic acid structures among the four strains of Lactobacillus plantarum lead to various immunological reactions in immune cells, as evidenced by the lipoteichoic acid recovered from the K8, K88, K5-5, and K55-5 strains of L. plantarum [121]. Teichoic acids are essential for the pathophysiology and development of antibiotic resistance [122]. According to Lebeer et al. [123], teichoic and lipoteichoic acids exhibit various bioactivities, including anticancer, immunomodulatory, and antioxidant activities.
5.4. Exopolysaccharides
According to Caggianiello et al. [124], lactobacilli and other bacteria produce a variety of homo- and heteropolysaccharides, including kefricin, glucans, and uronic acid. These are collectively referred to as exopolysaccharides (EPSs) and can be released extracellularly, cling to the surface of the microbial cell as a slime layer, or remain firmly attached as a capsule. These macromolecules have the power to defend against phages, phagocytes, and toxic substances; however, they also affect the immune system, physiological processes, lipid metabolism, and pathogen colonization in hosts. According to a study by Dinic et al. [125], EPS from Lactobacillus paraplantarum BGCG11 decreased proinflammatory (IL-I, TNF, and iNOS) and concurrently elevated anti-inflammatory (IL-6 and IL-10) cytokines, thereby reducing inflammation in rats. According to Liu et al. and Wang et al. [74,126], EPSs generated from probiotic Lactobacillus fermentum and Paenibacillus polymyxa cultures show antioxidant activity and may thus have therapeutic effects in diseases such as diabetes, atherosclerosis, and rheumatoid arthritis. Additionally, EPSs extracted from pathogenic E. coli and S. aureus prevented the development of biofilms and suppressed tumor growth and inflammation [74]. The bioremediation, pharmaceutical, food, and textile industries have significant applications of EPSs derived from various bacteria [127]. Examples of food additives include xanthan, alginate, gellen, levans, and pullulan [128]. Centrifugation is the first stage in a multiphase method to extract EPSs, which also includes acid protein removal, cold ethanol precipitation, filtration to remove small molecules, dialysis, and lyophilization [129]. EPSs are essential for cell adhesion and defense, and the structural diversity of EPSs produced by lactic acid bacteria (LAB) enables polymers to have a range of bioactivities, including immunomodulatory, antitumor, antimutagenicity, antioxidant, anti-inflammatory, antihypertensive, antibacterial, antiviral, cholesterol-lowering, and anti-gastrointestinal activities [130]. Khalil et al. [131] reported that EPS generated from Lactobacillus strains showed antibacterial and antioxidant activities and improved lipid metabolism by inhibiting cholesterol absorption. By increasing the activities of antioxidant enzymes, such as catalase, glutathione peroxidase, and superoxide dismutase, and lowering the levels of lipid peroxidation in serum and mouse livers, EPSs generated from Lactococcus lactis subsp. lactis displayed antioxidant activity [132]. Currently, the food industry uses EPSs as emulsifiers, stabilizers, and water-binding agents.
5.5. Enzymes
Enzymes are proteins that catalyze biological reactions. Based on their activities or functions, enzymes can be classified into six primary groups: oxidoreductases, transferases, hydrolases, lyases, isomerases, and ligases [133,134]. A small number of bacterial strains, primarily Bacillus subtilis and Bacillus licheniformis, as well as a few fungal strains, notably, Aspergillus niger and Aspergillus oryzae, are the primary sources of enzymes that are used in a variety of physiological, metabolic, and regulatory processes. A significant amount of glutathione peroxidase was detected in two strains of Lactobacillus fermentum, which was later discovered to possess strong in vitro antioxidant capabilities. Under difficult conditions such as temperature, pH, organic solvents, oxidizing agents, and detergents, Bacillus spp.can produce proteolytic enzymes in large yields that are remarkably stable. Catalase from a genetically modified strain of L. lactis can protect mice from chemically induced colon cancer [135,136,137].
5.6. Short Chain Fatty Acids (SCFAs)
Short-chain fatty acids (SCFAs) are an important class of compounds produced by gut bacteria such as Bacteroides and Firmicutes, which ferment plant polysaccharides [138]. Inulin and fructooligosaccharides, two prebiotics, are fermented to produce SCFAs, primarily acetate, propionate, and butyrate, which are found in the colon and feces at an estimated 60:20:20 molar ratio and aid in the regeneration of the intestinal epithelium [139,140]. In addition, they suppress the production of pro-inflammatory cytokines, preventing the activation of nuclear factor-kappa B (NF-κB). A reduction in atherogenesis in a mouse model was demonstrated using an in vivo butyrate model [141]. Acetate and lactate are produced by bifidobacteria when too many carbon atoms are available for development. Inhibiting the growth of Klebsiella oxytoca, for instance, Lactobacillus acidophilus, Lactobacillus fermentum, Lactobacillus paracasei ATCC 335, and Lactobacillus brevis produced SCFAs by lysing the cell wall [142]. SCFs exert several beneficial effects on health. In addition to improving colonic function and lowering pH, they promote the proliferation of epithelial cells and blood flow in the colon [143]. Bird et al. [144] found that SCFAs significantly lowered the prevalence of colorectal diseases. When colonic bacteria ferment undigested carbohydrates, they produce mostly acetate, propionate, and butyrate in ratios that normally vary from 3:1:1 to 10:2:1. Acetate aids cholesterol regulation and is used as a growth factor by other bacteria. Propionate and butyrate play a role in gluconeogenesis, providing colonocytes and epithelial cells with their main source of energy and promoting apoptosis of colon cancer cells [145].
5.7. Bacteriocins
Lactic acid bacteria (LAB), as well as other eubacteria and archaebacteria, produce tiny ribosomally synthesized peptides or proteins known as bacteriocins that can either kill or impede the growth of other bacteria. According to Soltani et al. [146], the therapeutic utility of bacteriocins as next-generation antimicrobials for reducing the threat posed by drug-resistant pathogenic organisms is highlighted by their restricted broad-spectrum inhibitory effect against bacterial growth. Examples include nisin, subtilosin, lactococcin G&Q, enterocin, lactocyclicin, bovicin, plantaricin, and lacticin, among others [147]. Bacteriocins have demonstrated potential for use in food preservation. Nisin was the first bacteriocin to receive regulatory approval for commercial use as a food preservative from organizations such as the European Food Safety Authority (EFSA), Food and FDA, and Health Canada. Currently, it is used as a food additive in more than 80 countries. Bacteriocins prevent pathogen growth in the GI tract by creating pores in cell membranes, preventing the proper construction of cell walls, and inhibiting enzyme and protein functions. Multibacteriocinogenic strains of L. paracasei and L. taiwanensis show antibacterial activity against E. coli, Salmonella gallinarum, and enteropathogenic E. coli [148,149]. Because of their various qualities, bacteriocins have been widely used in various applications, including medicine, cancer therapy, food, cosmetics, and veterinary medicine.
5.8. Vitamins
Vitamins are thermosensitive chemical substances that are necessary for the body to perform a number of physiological processes, including DNA replication, repair, and methylation, and vitamins must be supplied exogenously.Vitamins play a crucial role in many physiological processes such as bone health, brain function, and blood clotting, and riboflavin acts as a hydrogen carrier in redox reactions. Vitamin K also plays a role as a cofactor of gamma carboxylase activity in blood clotting, and various other critical vitamins, such as vitamin K, and various B-group vitamins, such as folate, riboflavin, cobalamin, pyridoxine, thymine, niacin, and nicotinic acid, are produced by lactic acid bacteria and Bifidobacterium sp. [150]. Numerous fermented foods, including fermented milk, yogurt, and cheese, are major sources of these vitamins, which help the digestive system. In addition to being essential for producing energy, controlling genes, and changing intestinal immunity, B-group vitamins, including B12, B2, B6, B9, and vitamin K, may all be synthesized by the gut microbiome on their own. For example, vitamins B2, B6, and B9 exerted anti-tumorigenic effects against pro-monocytic lymphoma cells [151].Cobalamin, generally known as vitamin B12 (B12), is a water-soluble vitamin essential for maintaining hematopoiesis and neuronal health. It is also an essential nutrient in animal products. Probiotics such as L. sanfranciscensis, L. reuteri, L. rossiae, and L. fermentum, which have been shown to synthesize vitamin B12 and could be useful substitutes for industrial production, have recently been found to contain genes encoding enzymes necessary for cobalamin (B12) synthesis [152,153,154]. In contrast to MK-6, MK-8, and MK-9, which are produced by Bacteroides fragilis, MK-10, MK-11, and MK-12 are produced by Eubacterium lentum, Lactococcus lactis ssp. lactis, and Lactococcus lactiscremoris [155]. Cortés-Martin et al. [156] found that the gut microbiota also produces dietary polyphenols. Aromatic amino acids are generated and metabolized in the gut to function as bioactive molecules in circulatory, renal, and brain systems [157].
5.9. Neurotransmitters
Neurotransmitters, such as serotonin, dopamine, norepinephrine, catecholamines, and acetylcholines, are produced by gut bacteria such as Bifidobacterium, Lactobacillus plantarum, Lactobacillus brevis, and Bacillus subtilis. These neurotransmitters play a major role in brain function via the gut–brain axis through the modulation of enteric nerve signaling. Tryptophan is an amino acid that is transformed into serotonin, which is responsible for mood improvement. Gamma-aminobutyric acid inhibits neurotransmission, and when it does not work, anxiety and depression result. Acetylcholine and catecholamines are essential for CNS activities, such as emotion, memory, learning, and motor control [157,158,159]. According to Patterson et al. [159], microbiome management can cure mental conditions linked to depression, and these compounds appear to have antidepressant potential.
5.10. Extracellular Vesicles
EVs are spherical, lipid bilayer, membrane-bound particles that release commensal bacteria, such as E. coli and Akkermansia muciniphila, into the environment. They are involved in the horizontal transfer of genetic material across bacterial species and contain a variety of substances, including proteins, DNA, RNA, glycolipids, polysaccharides, enzymes, and toxins. According to studies by Ahmadi Badi et al. and Chelakkot et al. [159,160], these substances are thought to regulate the permeability of the gut barrier and signaling pathways, maintain intestinal homeostasis, improve lipid profiles, and facilitate communication between the gut and brain. Survival, competitiveness, pathogenesis, and immunomodulation are some mechanisms regulated by bacterial EVs. They can also swiftly cross the mucosal barrier and interact with the host, thereby lowering the risk of sepsis. Previous studies have shown an association between obesity and reduced barrier integrity. Increased intestinal barrier permeability causes metabolic endotoxemia, which is the primary contributing factor to obesity-related metabolic diseases [161,162,163]. EVs derived from Akkermansia muciniphila reduced fat accumulation, body weight gain, and pathological abnormalities in high-fat diet (HFD)-fed mice; the tested EVs had the most significant effects on adipocyte size, epididymal white adipose tissue (eWAT) weight, lipid balance, and expression of inflammatory cytokines in the adipose tissue and glucose tolerance in diabetic mice. EVs derived from Propionibacterium freudenreichii can mitigate inflammation by modulating the NF-B pathway [161,164,165,166,167].Recently, Gurunathan et al. reported that Pseudomonas aeruginosa-derived outer membrane vesicles exhibited antibacterial and antibiofilm effects against Streptococcus mutans. Extracellular nanovesicles produced by Bacillus licheniformis showed anticancer effects against breast and lung cancer cells [167].
6. Applications of Postbiotics in the Food Industry
Functional foods, such as probiotics, prebiotics, and postbiotics, have recently received considerable attention from researchers, manufacturers, and consumers. The development of innovative functional foods and preventive medicine formulations for improving host health, as well as accurately characterizing their mechanisms of action, is currently the focus of a sizable section of postbiotic research [168]. A variety of food products with bioactive ingredients, such as probiotics, dairy, and non-dairy products, are already available in the market to meet the nutritional needs of consumers with various dietary preferences, such as those who are allergic to milk proteins, lactose intolerant, and vegetarians [168,169]. Source, components, types, and functional aspects of postbiotics is shown in Table 1.
Postbiotics are stable over a wide range of temperatures and pH levels, making it easy to add meals and components before thermal processing without affecting their effectiveness. Producers may gain from this, both technically and financially. They can be used in delivery systems such as pharmaceutical goods and/or functional meals because the right amount of postbiotics can be managed under manufacturing and storage conditions when survival is not the main determining factor [168]. Bacterial lysates containing cell surface proteins, enzymes, peptides, metabolites, neuropeptides, and lower organic acids such as lactic acid are examples of postbiotics. Fermentation is the most common postbiotic source in the food industry. Postbiotics are naturally present in many dairy products, such as kefir, kombucha, yogurt, and pickled vegetables. Generally, Lactobacillus sps., Bifidobacterium, Saccharomyces, Bacillus, Streptococcus, or Faecalibacterium genera are highly effective postbiotic-producing microorganisms in the form of cytoplasmic extracts and cell wall components [33,73,206,207].
According to several studies [28,30,34,72,208], several Lactobacillus species, including L. rhamnosus, L. bulgaricus, L. acidophilus, L. reuteri, L. casei, and L. fermentum, play significant roles in the food industry. On the other hand, the most prevalent probiotic producers of postbiotics are Bifidobacterium sps., including B. bifidum, B. longum, B. breve, and B. longum. According to an in vivo study, mice injected with B. longum showed a strong antibody response; however, the cell wall and cytoplasmic fractions had little effect on the immune system [209]. Additionally, L. plantarum species are thought to be potential food bio-preservatives and benefit animal gut health because they produce metabolites with high levels of mixed organic acids and bacteriocins. Nisin, a bacteriocin produced by Lactococcus lactis subsp. lactis, is used as a preservative in many foods including dairy products, infant formula, and canned soups [210]. EPS produced by LAB, such as L. rhamnosus, which are important in dairy products, may enhance the physicochemical and sensory qualities of food-based products [20]. Owing to the postbiotic composition of the supernatant from Lactobacillus plantarum YML007, the shelf life of soybeans was extended by up to two months [211]. Postbiotic enzymes, including purified phytases from Bifidobacterium pseudocatenulatum and Bifidobacterium longum spp. infantis, increase the amount of myoinositol triphosphate while lowering the amount of phytate in cereal combinations [212].
Cereals lose some vitamin B content when heated or ground. Vitamins B1, B2, B3, B9, B11, and B12 can be produced by additional bacteria as a result of grain fermentation and LAB pretreatment, which can compensate for these vitamin losses. Cereals that underwent LAB fermentation had considerably higher levels of total lysine, protein fractions, sugars, and soluble dietary fiber and a higher bioavailability of calcium, iron, and zinc. Wheat may also produce antioxidant peptides, γ-aminobutyric acid, and angiotensin I-converting enzyme-inhibitory peptides via LAB fermentation [213,214,215].
Postbiotics are chemicals obtained from specific bacteria that prevent microbiological food degradation and increase the shelf life of food. Postbiotics are highly significant in the food industry because they exhibit anti-microbial activity against both pathogenic and spoilage microorganisms through a variety of mechanisms, such as creating cavities in CM, changing proteins in cell walls, and lowering the pH of the bacterial cytoplasm [216]. The nutritional value and organoleptic changes in non-vegetarian food can be preserved by directly applying a postbiotic coating (for example, to fish fillets and slices of meat) or by spraying it (for example, on ground fish and meat), depending on the type of meat and postbiotic. Postbiotics comprising Pediococcus acidilactici, Latilactobacillus sakei, and Staphylococcus xylosus flavonoids and phenolics have been shown to reduce Salmonella typhimurium in chicken drumsticks [217]. By consuming products made from the fermentation of Saccharomyces cerevisiae, it may be possible to limit the amount of Salmonella enterica contamination in poultry products by consuming products made from S. cerevisiae fermentation [218]. Postbiotic-containing preservatives were found to be as effective as frequently used commercial preservatives in preserving vacuum-packaged cooked sausages as natural preservation technologies [219]. Postbiotics can be used as an alternative biocontrol for the safe production of dairy products, fruits, and vegetables. For instance, several bacteriocins and LABs play roles in controlling cheese-blowing errors. Postbiotics can be used as sanitizers in the food industry [15].
7. Overall Therapeutic Effects of Postbiotics
Postbiotics typically contain additional therapeutic and health-promoting components. Metabolites sustain healthy bacteria and reduce the chances of harmful living microorganisms [220]. Numerous bacteria in the gastrointestinal system catabolize indigestible carbohydrates to produce large amounts of butyrate and volatile SCFAs [221]. Enzymes that protect cells against oxidative damage, cancer, or heart disease, recognized as immunostimulants by teichoic acids, peptide antimicrobials that directly combat invasive bacteria in the colon, and the prebiotic/indigestible carbohydrate inulin have been shown to reduce constipation and increase the volume of feces. Postbiotics reduce lipid levels, inhibit fatty acid production, and prevent inflammatory diseases [220,222]. Digestible oligosaccharides enhance calcium absorption in humans, particularly during puberty and menstruation. The amounts of calcium, potassium, and magnesium ions in the intestinal lumen increase when cellular transformation is controlled and prevented in the intestinal lumen [20]. Postbiotics rich in butyrate and oligosaccharides are important for cancer prevention. Because of the presence of bacteriocins and organic acid-based postbiotics, Lactobacillus acidophilus LA5, Lactobacillus salivarius, and Lactobacillus casei 431 displayed antibiofilm effects against L. monocytogenes. Therefore, postbiotics can be used in the food industry to prevent and reduce bacterial biofilm development [16,223].Reactive oxygen species (ROS) can change the nature of lipids and proteins and eventually lead to cellular dysfunction, which can lead to permanent malfunctions such as diabetes and its consequences, microvascular disease, and cardiovascular effects. Living organisms can use enzymatic or non-enzymatic defenses, such as natural antioxidants (vitamins C and E), to quench ROS [224,225]. Superoxide dismutase (SOD), an antioxidant enzyme that aids in reducing free radical accumulation in the intestinal and colon lumen of rats, was found to be active in the cell-free extracts of strains belonging to Lactococcus and S. thermophilus, with Lactococcus exhibiting more activity than S. thermophilus [26].The Lactococcus lactis strain produces catalase (CAT), which is able to increase CAT activity in mice administered 1,2-dimethylhydrazine (DMH), which improves antioxidant capacity by decreasing H2O2 levels and preventing or reducing the severity of intestinal diseases caused by ROS [137]. Several lactic acid bacteria, including Bifidobacterium adolescentis, B. longum, B. infantis, and B. breve, can break down hydrogen peroxide. L. plantarum has been shown to have a significant antioxidant role through an increase in GPx concentration in serum and ruminal fluid of post-weaning lambs.Among the 25 distinct lactobacilli, the human strain L. plantarum 30 B has the highest catalase activity, whereas the human strain L. acidophilus 900 has the highest superoxide anion dismutation activity [67,226,227,228]. Additionally, in an animal model of inflammatory bowel disorder (IBD), L. acidophilus strain 900 suppressed the inflammatory process more effectively than L. plantarum strain 30 B, indicating that H2O2 is less harmful than superoxide anion radicals and ROS [227]. Collectively, the different Lactobacillus strains exhibit anti-inflammatory potential by expressing various antioxidant enzymes. Lactobacillus fermentum E-3 and E-18 were isolated from the intestinal microflora of a healthy child and displayed antioxidant activity by expressing high levels of GPx and MnSOD in the intestine, which are crucial for preventing lipid peroxidation and removing hydrogen peroxide [228].
Any drug or bioactive substance that suppresses inflammation may potentially have anticancer properties because inflammation and carcinogenesis are closely associated. Probiotics from fermented milk by Propionibacterium freudenreichii induce apoptosis in HGT-1 human gastric cancer cells [229]. The supernatant of L. rhamnosus GG lowered the expression of MMP-9, which aided in the breakdown of the intercellular matrix, promoted cancer cell penetration, and increased ZO-1 expression [230]. Lactobacillus plantarum-derived postbiotics exhibited selective cytotoxicity and increased apoptosis against MCF-7 and RG14 PM on HT29, RG11, and RI11 cells [231]. The severity of 2,4,6-trinitrobenzenesulphonic acid (TNBS)-induced colitis in mice was reduced with the oral administration of Faecalibacterium prausnitzii supernatant, and dysbiosis related to TNBS colitis was similarly improved [33]. An anti-inflammatory immunogen derived from yeast culture was shown by Jensen et al. [73] to activate human natural killer cells and B lymphocytes as well as to change the expression of chemokine receptors. According to Jensen et al. [73], cell wall fragments and metabolites from Bacillus coagulans cause human polymorphonuclear (PMN) cells to spontaneously inhibit and produce reactive oxygen species (ROS) in response to oxidative stress.Interleukin-12 (IL-12) production is inhibited and interferon-gamma (IFN-) and tumor necrosis factor-alpha (TNF-) production is stimulated by a peptidoglycan generated by Lactobacillus spp. However, peptidoglycans can also promote the synthesis of pro-inflammatory cytokines in macrophages, such as TNF- or IL-12, as well as the expression of IL-12p35 mRNA [232].
Levan, a polysaccharide derived from Bacillus licheniformis, was found to be efficient in preventing hyperglycemia and oxidative stress induced by diabetes in adult rats, suggesting that adding levan to the diet may help prevent diabetes-related molecular abnormalities such as blood glucose levels, improvement in peripheral sensitivity to residual insulin, and activation of Langerhans islets [233]. In experimentally induced diabetes mellitus (DM), folic acid administration lowers glycemia and improves the activity of specific enzymes, including superoxide dismutase (SOD) and catalase (CAT). Folic acid functions as an antioxidant by lowering the production of superoxide radicals catalyzed by nitric oxide synthase (NOS) [234].EPSs from Bacillus subtilis sp. suppress (BSEPS) control hyperglycemia and dyslipidemia in diabetic rats by increasing insulin levels and decreasing blood glucose and troponin blood concentrations [235].The fact that EPSs from Lactobacillus plantarum H31-2 decreased the amount of glucose in the supernatant of insulin-resistant HepG2 cells suggests that EPS H31-2 may help these cells to take up glucose through the AMPK/PI3K/Akt pathway. The expression of the glycometabolism-related genes glucose transporter 4 (GLUT-4), protein kinase B (Akt-2), and AMP-activated kinase (AMPK) was also elevated by EPS H31-2. According to these findings, Lactobacillus plantarum EPS H31-2 may effectively inhibit pancreatic amylase activity, which lowers blood glucose levels in type 2 diabetes (T2DM) patients, indicating that it may be utilized to both prevent and treat diabetes [236].In a high-fat diet and streptozotocin-induced type 2 diabetes in mice, a mixture of multiple Lactobacillus species reduced fasting blood glucose (FBG), hemoglobin A1c (HbA1C), and leptin levels and improved glucagon-like peptide-1 (GLP-1) levels [237]. In the absence of detectable changes in the composition of the microbiota or metabolome, L. plantarum bacteriocin plantaricin effectively reduced body weight and food intake in mice fed a high-fat diet (HFD) [238]. By activating NOD2, peptidoglycan-containing muramyl dipeptide (MDP) was demonstrated to be an insulin-sensitizing postbiotic that can reduce adipose tissue inflammation and glucose intolerance in obese mice without influencing body weight or altering microbiota composition [239]. NOD2 protects against inflammatory colitis by reducing insulin resistance and inflammation induced by other bacterial products. NF-ҡB and receptor-interacting serine/threonine-protein kinase 2 (RIPK2) are both involved in NOD2 activation [240,241,242]. A preclinical model of Caenorhabditis elegans suggested that Bifidobacterium animalis subsp. lactis BPL1-derived lipoteichoic acids (LTAs) function as novel lipid modulators with fat-reducing capabilities through the insulin-like signaling pathway (IGF-1) and prevent metabolic syndrome and diabetes-related disorders [243].
According to Taverniti and Guglielmetti [25], the postbiotic produced by lactic acid bacteria can quickly interact with immune cells and the epithelium to activate innate immunity. For instance, Lacticaseibacillus rhamnosus HN001 improves leukocyte phagocytic activity, which boosts immunity, and postbiotics produced by L. gasseri TMC0356 have immunomodulatory activity [244], and these bacteria carry out their immunomodulatory activities by increasing Th1-associated cytokine levels and decreasing Th2-related cytokines [245]. One study found that the LPS-induced TLR-4 pathway improved the ability of peptidoglycans from several Lactobacillus species to suppress the production of inflammatory cytokines in macrophage-like cell types [246]. Conversely, a combination of heat-inactivated probiotic strains, including Lactobacillus sps., protected intestinal cells from infection by E. coli in in vitro models of the intestinal mucosa (HT29-MTX cells), ensuring the restoration of tight junction function and membrane integrity, preventing an increase in paracellular permeability and penetration of pathogens into the intestinal epithelium, and modulating cytokine gene expression. Another study found that the probiotic strain Streptococcus thermophilus CRL1190 and its EPSs decreased Helicobacter pylori adhesion and lowered the inflammatory response in a human gastric adenocarcinoma epithelial cell line (AGS cells) [247]. Heat-killed S. boulardii can maintain the gut barrier by preserving intestinal permeability at physiological levels, lowering bacterial translocation, and preventing mucosal lesions; thus, heat-killed S. boulardii treatment can maintain the gut barrier [248]. Similarly, another study found that the metabolic byproducts of an infant formula fermented with Lactobacillus paracasei CBA L74 can protect the host from pathogens and enteric pathogens by inhibiting immune cell inflammation, and these byproducts also have protective effects against colitis [249]. When administered orally to newborn rats infected with E. coli K1, a new secretory protein, HM0539, produced by Lacticaseibacillus rhamnosus GG, was used to prevent and treat diseases related to intestinal barrier dysfunction. HM0539 promoted the development of neonatal intestinal defense and was sufficient to prevent E. coli K1 pathogenesis. The researchers also discovered that HM0539 protected against liver damage, colitis induced by dextran sulfate sodium (DSS), and bacterial translocation induced by LPS/D-galactosamine [250].Postbiotic Lactobacillus casei Zhang (LcZ) increases the production of proinflammatory cytokines and the transcription of TLR2, TLR3, TLR4, and TLR9, boosting the macrophage-mediated innate immune response [251]. LcZ was heat-inactivated and resuspended at a concentration of 106 CFU/mL in PBS. The inactive preparation boosted the secretion of cellular immune markers more than the live preparation, according to a study using the live and inactive forms of Bacillus amyloliquefaciens FPTB16 and Bacillus subtilis FPTB13 [252]. In addition, a mouse study found that combining heat-inactivated LAB with two heat treatments—30 min at 100 °C and 15 min at 121 °C—increased macrophage immunomodulatory activity [253]. Compared with heat-inactivated postbiotics from Enterococcus gallinarum L-1, ultraviolet-inactivated postbiotics were more effective in boosting phagocyte function. The results showed that the postbiotic Lactobacillus gasseri TMC0356 increased IL-12 production in macrophages more compared toprobiotics. This suggests that heat treatment enhances the capacity of the strain to activate IL-12 production in macrophages and that the postbiotic form has a greater immunomodulatory effect than the probiotic form. Postbiotics produced by Lactobacillus acidophilus A2, Lactobacillus gasseri A5,and Lactobacillus salivarius A6 alter the Th1-mediated immune response by promoting the proliferation of IL-10 and IL-12 p70, IFN-production in splenocytes, and IL-12 p70 secretion in dendritic cells, respectively [24,252,253]. SCFAs potentially inhibit the production and function of regulatory T cells by inhibiting histone deacetylase (HDAC) induction and are also important factors in building the link between the microbiome and the immune system [254]. Lactobacillus acidophilus and Lactobacillus casei have been shown to increase interleukin-10 (IL-10) production while simultaneously decreasing pro-inflammatory tumor necrosis factor (TNF-) cytokine secretion [86]. It has been demonstrated that soluble components derived from L. reuteri strain CRL1098 significantly reduce the release of pro-inflammatory mediators such as nitric oxide (NO), cyclooxygenase 2 (COX-2), heat shock proteins 70 (Hsp70), TNF-, and IL-6 [255]. Similar anti-inflammatory effects were demonstrated by S. boulardii, which were attributed to excrete low-molecular-weight-soluble components that play a role in inhibiting NF-ҡB activation and NF-B-mediated IL-8 gene production in monocytes and intestinal epithelial cells [256].
8. Antiviral, Antibacterial, Antioxidants, Anticancer, and Anti-Inflammatory Mechanisms of Postbiotics
8.1. Antiviral
Viral diseases pose significant global public health hazards, underscoring the importance of antiviral medications and vaccines in preventing infections. Postbiotic antiviral effects predominantly manifest through the restraint of viral attachment to host cells, thereby delaying the onset of infection (Figure 4). Postbiotics inhibit viral binding to host cell receptors, preventing viral entry and fortifying the host immune system [257,258].
The immune system plays a pivotal role in actively safeguarding cells, eliminating viruses, and initiating a pro-inflammatory response to establish Th1-type immunity. This involves the production of inflammatory chemokines, cytokines, and interleukins, including TNF, interferons, IL-23, IL-18, and IL12. Additionally, T-lymphocytes, NK cells, and monocytes/macrophages are activated to produce cytokines [259].
Probiotics and their metabolites protect against viral infections by enhancing both innate and adaptive immunity. This results in a reduction in sickness duration, virus shedding, and the frequency of episodes. Furthermore, postbiotics normalize intestinal permeability and augment the production of virus-specific antibodies [260].
Postbiotics exhibit antiviral efficacy by impeding retroviral reverse transcriptases and preventing viral uptake by host cells. The specific probiotic strain used and the type of virus significantly influence the antiviral effects of postbiotics. For instance, Anwar et al. [261] demonstrated that plant-derived probiotics, known as plantaricins, mitigate SARS-CoV-2 infection by modulating the immune system. Plantaricin compounds inhibit COVID-19 pathogenicity by binding to the spike glycoprotein (S) [262]. Metabolites from L. rhamnosus deter viruses from attaching to cell lines [263].
The organic acids of probiotic bacteria, particularly those produced by attaching to the glycoprotein (S) of viruses, exhibit an antiviral mechanism, preventing viruses from attaching to the angiotensin-converting enzyme (ACE2) [264]. Lauric acid and meristic acid are reported to significantly inhibit the growth and development of viruses [265]. Furthermore, organic acids produced by probiotics bind to the spike glycoprotein (S), preventing it from interacting with the angiotensin-converting enzyme 2 (ACE2). Microbial-derived peptides hinder viral proliferation by inhibiting endosomal acidification [266].
8.2. Antibacterial
The primary bioactive components of postbiotics consist of organic acids, including lactic acid, acetic acid, tartaric acid, malic acid, and citric acid. These acids inhibit bacterial growth by reducing pH levels and altering membrane integrity. Bacteriocins contribute to inhibiting bacterial viability by targeting bacterial cytoplasmic membranes and generating spores [168,267,268,269,270].
Short-chain fatty acids, such as lauric and meristic acids, effectively inhibit bacterial formation and proliferation. These acids disrupt the electron transport chain, alter the structure and activity of enzymes, and induce morphological and functional changes in delicate components like proteins. As a result, short-chain fatty acids cause cell lysis and enhance membrane permeability in bacteria [271]. Recently, Gurunathan et al. reported that Pseudomonas aeruginosa-derived outer membrane vesicles exhibited antibacterial and antibiofilm effects against Streptococcus mutans [167]. Antibacterial peptides produced by bacteria exhibit robust antibacterial activity by rupturing microbial membranes and accumulating metabolic nuclei. The mechanisms underlying the antibacterial action of peptides involve creating physical holes that facilitate the leakage of cellular content, causing damage to delicate microbial intracellular components, initiating lethal processes such as inducing hydrolases that negatively impact cell walls, and acidifying the bacterial cell membrane (Figure 4) [168,272,273,274].
8.3. Anticancer
Postbiotics exhibit anticancer properties by inhibiting cell growth and proliferation and enhancing apoptotic effects. Polysaccharides, among the various components of postbiotics, including SCFAs, metabolites, microbial cell fractions, functional proteins, EPSs, and cell lysates, demonstrate the most prominent anticancer activity [13,275]. The molecular mechanisms underlying postbiotic anticancer effects involve the modulation of immune responses, suppression of mutagenesis and carcinogens, activation of pro-apoptotic pathways, decreased bacterial translocation, and increased apoptosis and autophagy for the prevention and treatment of cancer (Figure 4).
Numerous in vitro and in vivo experiments have elucidated the anticancer properties of postbiotics. For instance, SCFAs from propionibacteria inhibit human colorectal cancer cell lines HT-29 and Caco-2 by inducing the loss of mitochondrial transmembrane potential, generating reactive oxygen species, activating caspase-3, modulating Bcl-2 regulation, and inducing nuclear chromatin condensation [276]. The administration of the cytoplasmic fraction of lactic acid bacteria (LAB) enhances specific antitumor activity by modulating cellular immunity [277]. Lactobacillus salivarius REN probiotics inhibit oral carcinogenesis in a dose-dependent manner, providing defense against oxidative damage, reducing COX-2 and PCNA expression, and protecting against oxidative damage [278]. A FACS analysis of human colorectal cancer RKO cells revealed that SCFAs generated by P. freudenreichii increase the sub-G1 phase and decrease the S and G2/M phases [279].
In human colorectal cancer cells (T4056 and HT-29), cell-free L. spp. pentasaccharides induce apoptosis and prevent the progression of the S-phase cell cycle. In human colorectal SW480 cancer cells, cytoplasmic extracts and cell walls of L. lactis spp. demonstrate an antiproliferative effect linked to a lower expression of cyclin D1 [280]. The probiotic-derived p8 protein exhibits antiproliferative activity by inhibiting the p53-p21-Cyclin B1/Cdk1 signal pathway, leading to growth arrest at the G2 phase of the cell cycle in human colorectal DLD-1 cells [281]. Exopolysaccharides derived from L. acidophilus 606 inhibit the proliferation of HT-29 colon cancer cells by directly affecting cell morphology and activating autophagic cell death through the induction of Bcl-2, Bak, Beclin-1, and GRP78 [282].
Postbiotics eliminate tumor cells by modulating immune responses and activate anti-inflammatory and pro-apoptotic cytokines such as TRAIL, interleukin (IL)-10, and TGF-β [283]. SCFAs protect the mucosal layer by lowering the levels of immunomodulators such as prostaglandins in human breast cancer MCF7 cells [284]. Heat-killed L. pentosus b240 promotes the production of immunoglobulin A (IgA), IL-6, IL-10, interferon (IFN)-γ, and tumor necrosis factor, but not IL-4, IL-5, B-cell activating factors, IFN-α, IFN-β, and transforming growth factor-β1 [285]. L. plantarum strain YYC-3 strongly inhibits human colorectal cancer HT-29 and Caco2 cell lines by modulating the immune system, downregulating the expression of inflammatory cytokines interleukin IL-6, IL-17, and IL-22, and reducing the infiltration of inflammatory cells [286]. Extracellular nanovesicles produced by Bacillus licheniformis showed anticancer effects against breast and lung cancer cells [193].These studies collectively indicate that postbiotics possess anticancer properties.
8.4. Anti-Diabetic
Diabetes mellitus is a chronic, irreversible condition characterized by dysregulated insulin response or synthesis in the body. Type 1 diabetes mellitus (T1DM) and type 2 diabetes mellitus (T2DM) represent distinct forms of diabetes. T2DM, accounting for approximately 90% of diabetes cases, arises from insulin-resistant cells requiring elevated insulin levels for effective sugar metabolism. The gut microbiota and obesity share a close connection, and probiotic strains like Lactobacillus pentosus GSSK2 and Lactobacillus plantarum GS26A can contribute to obesity reduction [287]. Probiotics significantly enhance cytokine production and adiponectin levels compared with lyophilized single- or multi-strain formulations [288].
With an elevation in Akkermansia and Prevotella abundance, probiotics such as Lactobacillus plantarum L-14-derived exopolysaccharides activate the SIRT1-IRS1-Akt and GLUT2 pathways, improving lipid and glucose metabolism in insulin-resistant mice [289]. The TLR2-AMPK signaling pathway is activated by postbiotics, exemplified by exopolysaccharides from Lactobacillus plantarum L-14, inhibiting immature cell development into mature adipocytes and regulating body weight and lipid profiles in mice [290].The Lactobacillus brevis long-chain polyphosphate may alleviate intestinal inflammation and enhance intestinal barrier function by triggering the extracellular-regulated protein kinase (ERK) signaling pathway [291]. Muramyl dipeptide mitigates obesity-induced insulin resistance by targeting nucleotide-binding oligomerization domain-containing protein 2 (NOD2) and interferon regulatory factor 4 (IRF4) [239]. The overexpression of NLRC3 enhances colonic epithelial barrier integrity by increasing TNF receptor-associated factor 6 (TRAF6)-mediated ZO-1 and occludin expression. Butyrate improves the intestinal barrier in type 2 diabetic mice by upregulating GPR43 expression and stimulating NLCR3 in a TRAF6-dependent manner [292]. In NAFLD mice, butyrate restores intestinal barrier function by increasing ZO-1 expression and mitigates metabolic disorders and intestinal epithelial impairment in type 2 diabetic mice by promoting insulin secretion without compensatory hyperplasia in pancreatic β cells [293]. Postbiotics increaseantidiabetic activity by increasing insulin production, WAT Browning, glucose homeostasis, and insulin resistance (Figure 4).
8.5. Anti-Inflammatory
Inflammation serves as the initial response of an organism to infection, and several inflammatory disorders, including inflammatory bowel disease (IBD), arthritis, gastritis, asthma, atherosclerosis, and chronic inflammation, are associated with adverse side effects. Dysregulation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and/or mitogen-activated kinase (MAPK) pathways, primarily involved in cell cycle regulation, is a common characteristic feature of these diseases [282,294,295]. A substantial body of evidence suggests the effectiveness of probiotics in alleviating the signs and symptoms of IBD and other inflammatory gastrointestinal diseases. The main molecular targets of the substances present in these probiotics are presumed to be the NF-κB and/or MAPK pathways. Bioactive compounds derived from probiotics emerge as a promising therapeutic strategy for mitigating excessive and/or prolonged inflammatory reactions [296,297,298].
MAPK signaling pathways, including c-Jun N-terminal kinases (JNKs), extracellular signal-regulated kinases (ERKs), and p38 MAP kinases, significantly influence cellular responses such as apoptosis, cytokine production, the regulation of proliferation, and inflammatory responses [299]. Postbiotics induce a robust response in inflammatory pathways by modulating the transcription of pro-inflammatory genes and the expression levels of pro-inflammatory proteins (Figure 4). However, the impact of postbiotics on the inflammatory response is contingent on factors like origin, bacterial species, strains, cell-free supernatant (CFS), and heat-killed bacteria (HKB). Both anti-inflammatory and immunostimulatory effects have been demonstrated, suggesting a heterogeneous structure and composition of different postbiotic media [86,300,301,302].
Inhibiting NF-κB nuclear translocation, IκB phosphorylation, and proteasomal degradation are three mechanisms through which postbiotics from CFS and HKB exert anti-inflammatory effects [303,304,305]. CFS from L. rhamnosus CNCM I-4036 enhances the expression of the IκB component NFKBIA, reinforcing the sequestration of NF-κB in the cytoplasm. Given that a similar set of pattern recognition receptors (PRRs) activates both the NF-κB and MAPK pathways, downstream processes may be impacted simultaneously, attenuating the inflammatory response by reducing NF-κB activity and MAPK activation [9]. Similar results were observed with isolated postbiotic factors, including 8.7 kDa proteins from L. plantarum 10hk2 CFS, DNA obtained from L. rhamnosus LGG, HKB from Weissella cibaria JW15, and CFS from L. salivarius MG4265 [2,121,306].
The injection of L. reuteri extract into mice accelerated wound healing, dependent on the activation of the PI3K/AKT/-catenin/TGF-1 pathway. Additionally, the CSF of L. fermentum, by inhibiting the PI3K/Akt/mTOR pathway, reduces the induction of H2O2-induced senescence in mouse adipocytes [307,308]. In monocytic cells, lipoteichoic acid (LTA) from L. plantarum exhibits anti-inflammatory effects and reduces the generation of TNF-α [72,309]. A crucial factor influencing the production of anti-inflammatory actions is the modification of postbiotic structure. For instance, in L. rhamnosus GG, the removal of D-alanine and substitution with glucosyl substitutions increased anti-inflammatory activity in murine colitis models [310,311]. In human intestinal cells, the elongation factor Tu (eFTu) and chaperone protein GroeL from L. johnsonii La1 stimulate IL-8 secretion in a CD14-dependent manner. CD14, acting as a co-receptor, is believed to amplify signals by inducing TLR4 endocytosis and activating TRIF-dependent pathways, resulting in the production of type I IFNs [312,313]. Probiotic-derived oligonucleotides such as gDNA and CpG motifs dramatically reduce LPS-induced IL-6 mRNA levels in RAW264.7 macrophages. L. plantarum gDNA pretreatment decreases the pro-inflammatory effect of LPS in THP-1 monocytic cells [170,314,315].
9. Conclusions and Future Perspectives
Since 2013, international research projects on postbiotics have seen a considerable increase. This review discusses a number of postbiotic topics, including historical viewpoints, significant developmental milestones, and noteworthy characteristics.The production, purification, and characterization of postbiotics, as well as their classification and the role of specific compounds, such as proteins, fatty acids, peptides, bacteriocins, enzymes, organic acids, extracellular vesicles, exopolysaccharides, and antioxidants, in the food industry are discussed. Given their unique properties, postbiotics exhibit promise as tools to control pathogenic microorganisms and function as antioxidants, anticancer, anti-diabetic, and anti-inflammatory agents. The therapeutic effects and mechanisms of postbiotics as antiviral, antibacterial, antioxidants, anticancer, and anti-inflammatory agents are presented. Although postbiotics exhibit pronounced effects on the food and health industries, further research is essential to establish standardized ways to evaluate the quantitative effects in a more reasonable, scientific, and accurate manner. Studies should also delve into the effects of postbiotics on health and their specific processes, necessitating in-depth investigation into the biological response of metabolites and host–postbiotic interactions using different omics approaches. This will bolster the wholesome and long-term growth of postbiotic preparations. Despite the scarcity of studies on postbiotic applications in the food industry and medicine, larger, higher-quality, and more rigorous trial data are required to demonstrate their benefits. Several countries have incorporated postbiotics into their national legislation, making goods available to the public despite the absence of defined global regulatory standards. This study contributes valuable standards for the evaluation of postbiotics, including their definition, mechanism, efficacy, and safety.
While postbiotic therapy’s safety and potential hazards remain incompletely studied and understood, further multicenter research is essential to establish outcomes and safety profiles for different postbiotics. Numerous metabolic, clinical, mechanistic, and biomedical studies are needed to identify new types of postbiotics, determine safe doses, and elucidate compound chemical structures. Understanding the mechanisms of immunomodulation by food is crucial for advancing and creating novel health solutions in the future.
Author Contributions
S.G.: conceptualization, writing—original draft preparation, and writing—review and editing; P.T.: figures and references; J.-H.K.: validation, resources, and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We would have never been able to complete this review without the people who have contributed to prebiotics, probiotics, and postbiotics and owe our gratitude to them. We have cited as many references as permitted and apologize to the authors of publications we have not cited because of the above limitation. This study was supported by BiOrgan Solution Ltd., Seoul, Republic of Korea.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bhat, F.M.; Riar, C.S. Effect of chemical composition, granule structure and crystalline form of pigmented rice starches on their functional characteristics. Food Chem. 2019, 297, 124984. [Google Scholar] [CrossRef] [PubMed]
- Ghadimi, D.; Helwig, U.; Schrezenmeir, J.; Heller, K.J.; De Vrese, M. Epigenetic imprinting by commensal probiotics inhibits the IL-23/IL-17 axis in an in vitro model of the intestinal mucosal immune system. J. Leukoc. Biol. 2012, 92, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Gul, K.; Singh, A.K.; Jabeen, R. Nutraceuticals and Functional Foods: The Foods for the Future World. Crit. Rev. Food. Sci. Nutr. 2016, 56, 2617–2627. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.R.; Mortimer, D.; Rose, M.; Smith, F.; Steel, Z.; Panton, S. Recently listed Stockholm convention POPs: Analytical methodology, occurrence in food and dietary exposure. Sci. Total Environ. 2019, 678, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Balthazar, C.F.; Guimarães, J.F.; Coutinho, N.M.; Pimentel, T.C.; Ranadheera, C.S.; Santillo, A.; Albenzio, M.; Cruz, A.G.; Sant’Ana, A.S. The future of functional food: Emerging technologies application on prebiotics, probiotics and postbiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2560–2586. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nature Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef]
- Collado, M.C.; Isolauri, E.; Salminen, S.; Sanz, Y. The impact of probiotic on gut health. Curr. Drug Metab. 2009, 10, 68–78. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Vyas, U.; Ranganathan, N. Probiotics, prebiotics, and synbiotics: Gut and beyond. Gastroenterol. Res. Pract. 2012, 2012, 872716. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Gurry, T. Synbiotic approaches to human health and well-being. Microb. Biotechnol. 2017, 10, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef] [PubMed]
- Altveş, S.; Yildiz, H.K.; Vural, H.C. Interaction of the microbiota with the human body in health and diseases. Biosci. Microbiota Food Health 2020, 39, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-González, P.F.; Aguilar-Toalá, J.E.; García, H.S.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Protective Effect of the Intracellular Content from Potential Probiotic Bacteria against Oxidative Damage Induced by Acrylamide in Human Erythrocytes. Probiotics Antimicrob Proteins 2020, 12, 1459–1470. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Barros, C.P.; Pires, R.P.S.; Guimarães, J.T.; Abud, Y.K.D.; Almada, C.N.; Pimentel, T.C.; Sant’Anna, C.; De-Melo, L.D.B.; Duarte, M.C.K.H.; Silva, M.C.; et al. Ohmic heating as a method of obtaining paraprobiotics: Impacts on cell structure and viability by flow cytometry. Food Res. Int. 2021, 140, 110061. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Amores-Arrocha, A.; Roldán, A.; Jiménez-Cantizano, A.; Caro, I.; Palacios, V. Evaluation of the use of multiflora bee pollen on the volatile compounds and sensorial profile of Palomino fino and Riesling white young wines. Food Res. Int. 2018, 105, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and paraprobiotics: From concepts to applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Kuipers, E.J.; Peppelenbosch, M.P. Functional genomic analyses of the gut microbiota for CRC screening. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 741–745. [Google Scholar] [CrossRef] [PubMed]
- de Almada, C.N.; Almada, C.N.; Martinez, R.C.R.; Sant’Ana, A.S. Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci. Technol. 2016, 58, 96–114. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Sabharwal, V.; Kaushik, P.; Joshi, A.; Aayushi, A.; Suri, M. Postbiotics: From emerging concept to application. Front. Sustain. Food Syst. 2022, 6, 887642. [Google Scholar] [CrossRef]
- Choi, S.S.; Kim, Y.; Han, K.S.; You, S.; Oh, S.; Kim, S.H. Effects of Lactobacillus strains on cancer cell proliferation and oxidative stress in vitro. Lett. Appl. Microbiol. 2006, 42, 452–458. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, H.; Cho, I.Y.; Paik, H.D.; Park, E. Dietary supplementation of probiotic Bacillus polyfermenticus, Bispan strain, modulates natural killer cell and T cell subset populations and immunoglobulin G levels in human subjects. J. Med. Food. 2006, 9, 321–327. [Google Scholar] [CrossRef]
- Kaji, R.; Kiyoshima-Shibata, J.; Nagaoka, M.; Nanno, M.; Shida, K. Bacterial teichoic acids reverse predominant IL-12 production induced by certain Lactobacillus strains into predominant IL-10 production via TLR2-dependent ERK activation in macrophages. J. Immunol. 2010, 184, 3505–3513. [Google Scholar] [CrossRef]
- Sharma, B.; Srivastava, S.; Singh, N.; Sachdev, V.; Kapur, S.; Saraya, A. Role of probiotics on gut permeability and endotoxemia in patients with acute pancreatitis: A double-blind randomized controlled trial. J. Clin. Gastroenterol. 2011, 45, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Kim, M.S.; Jung, M.J.; Roh, S.W.; Nam, Y.D.; Park, E.J.; Bae, J.W. Leucobacterceler sp. nov., isolated from Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2011, 61, 2353–2357. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacteriumprausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Furushiro, M.; Hirai, K.; Motoike, M.; Watanabe, T.; Yokokura, T. Purification and characterization of an antihypertensive compound from Lactobacillus casei. Agric. Biol. Chem. 1990, 54, 3211–3219. [Google Scholar] [CrossRef]
- Antunes, L.C.; Han, J.; Ferreira, R.B.; Lolić, P.; Borchers, C.H.; Finlay, B.B. Effect of antibiotic treatment on the intestinal metabolome. Antimicrob. Agents Chemother. 2011, 55, 1494–1503. [Google Scholar] [CrossRef]
- Kok, M.G.; Ruijken, M.M.; Swann, J.R.; Wilson, I.D.; Somsen, G.W.; de Jong, G.J. Anionic metabolic profiling of urine from antibiotic-treated rats by capillary electrophoresis-mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 2585–2594. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; Romero, M.; Jiménez, R.; Sánchez, M.; Pérez-Vizcaíno, F.; Duarte, J. Antihypertensive Effects of Probiotics. Curr. Hypertens. Rep. 2017, 19, 26. [Google Scholar] [CrossRef]
- Chaluvadi, S.; Hotchkiss, A.T.; Yam, K.L. Gut Microbiota. In Impact of Probiotics, Prebiotics, Synbiotics, Pharmabiotics, and Postbiotics on Human Health; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 515–523. [Google Scholar]
- Amores, G.; Virto, M. Total and Free Fatty Acids Analysis in Milk and Dairy Fat. Separations 2019, 6, 14. [Google Scholar] [CrossRef]
- Klemashevich, C.; Wu, C.; Howsmon, D.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Rational identification of diet-derived postbiotics for improving intestinal microbiota function. Curr. Opin. Biotechnol. 2014, 26, 85–90. [Google Scholar] [CrossRef]
- Li, W.; Ji, J.; Rui, X.; Yu, J.; Tang, W.; Chen, X.; Jiang, M.; Dong, M. Production of exopolysaccharides by Lactobacillus helveticus MB2–1 and its functional characteristics in vitro. LWT—Food Sci. Technol. 2014, 59, 732–739. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Mechnikov, I. Ilya Mechnikov—Nobel Lecture. In: NobelPrize.org. 1908. Available online: https://www.nobelprize.org/prizes/medicine/1908/mechnikov/lecture/ (accessed on 6 June 2023).
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food. Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Kranich, J.; Maslowski, K.M.; Mackay, C.R. Commensal flora and the regulation of inflammatory and autoimmune responses. Semin Immunol. 2011, 23, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Gordon, J.I. The core gut microbiome, energy balance and obesity. J. Physiol. 2009, 587, 4153–4158. [Google Scholar] [CrossRef] [PubMed]
- Kostić, T.; Sessitsch, A. Microbial diagnostic microarrays for the detection and typing of food-and water-borne (bacterial) pathogens. Microarrays 2011, 1, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Xuan, C.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef]
- Qiu, X.; Macchietto, M.G.; Liu, X.; Lu, Y.; Ma, Y.; Guo, H.; Saqui-Salces, M.; Bernlohr, D.A.; Chen, C.; Shen, S.; et al. Identification of gut microbiota and microbial metabolites regulated by an antimicrobial peptide lipocalin 2 in high fat diet-induced obesity. Int. J. Obes. 2021, 45, 143–154. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Tlaskalová-Hogenová, H.; Stěpánková, R.; Kozáková, H.; Hudcovic, T.; Vannucci, L.; Tučková, L.; Rossmann, P.; Hrnčíř, T.; Kverka, M.; Zákostelská, Z.; et al. The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: Contribution of germ-free and gnotobiotic animal models of human diseases. Cell Mol. Immunol. 2011, 8, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; de Vos, W.M. The First 1000 Cultured Species of the Human Gastrointestinal Microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, P.; Bobe, A.; Dolan, K.; Leone, V.; Martinez, K. Nutritional modulation of gut microbiota—The impact on metabolic disease pathophysiology. J. Nutr. Biochem. 2016, 28, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Ben-Zeev Brik, R.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Tompkins, T.; Heimbach, J.T.; Kolida, S. Weight of evidence needed to substantiate a health effect for probiotics and prebiotics: Regulatory considerations in Canada, E.U., and U.S. Eur. J. Nutr. 2005, 44, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Yelin, I.; Flett, K.B.; Merakou, C.; Mehrotra, P.; Stam, J.; Snesrud, E.; Hinkle, M.; Lesho, E.; McGann, P.; McAdam, A.J.; et al. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat. Med. 2019, 25, 1728–1732. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Exosomal vesicles enhance immunosuppression in chronic inflammation: Impact in cellular senescence and the aging process. Cell Signal. 2020, 75, 109771. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Gilani, N.; Abbasi, A.; Khani, N. Postbiotics, as dynamic biomolecules, and their promising role in promoting food safety. Biointerface Res. Appl. Chem. 2021, 11, 14529–14544. [Google Scholar]
- Humam, A.M.; Loh, T.C.; Foo, H.L.; Izuddin, W.I.; Zulkifli, I.; Samsudin, A.A.; Mustapha, N.M. Supplementation of postbiotic RI11 improves antioxidant enzyme activity, upregulated gut barrier genes, and reduced cytokine, acute phase protein, and heat shock protein 70 gene expression levels in heat-stressed broilers. Poult. Sci. 2021, 100, 100908. [Google Scholar] [CrossRef] [PubMed]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Molaei, R.; Guimarães, J.T. A review on preparation and chemical analysis of postbiotics from lactic acid bacteria. Enzyme Microb Technol. 2021, 143, 109722. [Google Scholar] [CrossRef]
- Mirnejad, R.; Vahdati, A.R.; Rashidiani, J.; Erfani, M.; Piranfar, V. The antimicrobial effect of lactobacillus casei culture supernatant against multiple drug resistant clinical isolates of Shigella sonnei and Shigella flexneri in vitro. Iran Red Crescent Med. J. 2013, 15, 122–126. [Google Scholar] [CrossRef]
- Carmen, C.M.; Seppo, S.; Vinderola, G. Postbiotics: Defining the impact of inactivated microbes and their metabolites on promotion of health. In The Human Microbiome in Early Life; Academic Press: Cambridge, MA, USA, 2021; pp. 257–268. [Google Scholar]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef]
- Kim, H.G.; Lee, S.Y.; Kim, N.R.; Lee, H.Y.; Ko, M.Y.; Jung, B.J.; Kim, C.M.; Lee, J.M.; Park, J.H.; Han, S.H.; et al. Lactobacillus plantarum lipoteichoic acid down-regulated Shigella flexneri peptidoglycan-induced inflammation. Mol. Immunol. 2011, 48, 382–391. [Google Scholar] [CrossRef]
- Jensen, G.S.; Benson, K.F.; Carter, S.G.; Endres, J.R. GanedenBC30 cell wall and metabolites: Anti-inflammatory and immune modulating effects in vitro. BMC Immunol. 2010, 11, 15. [Google Scholar] [CrossRef]
- Wang, K.; Niu, M.; Song, D.; Song, X.; Zhao, J.; Wu, Y.; Lu, B.; Niu, G. Preparation, partial characterization and biological activity of exopolysaccharides produced from Lactobacillus fermentum S1. J. Biosci. Bioeng. 2020, 129, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536 on the healthy gut microbiota composition at phyla and species level: A preliminary study. World J. Gastroenterol. 2017, 23, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, E.; Rinninella, E.; Basilico, M.; Colomier, E.; Rasetti, C.; Larussa, T.; Santori, P.; Abenavoli, L. From Pre- and Probiotics to Post-Biotics: A Narrative Review. Int. J. Environ. Res. Public Health 2021, 19, 37. [Google Scholar] [CrossRef]
- Patel, A.; Prajapati, J.B. Food and health applications of exopolysaccharides produced by lactic acid bacteria. Adv. Dairy Res. 2013, 1, 1–7. [Google Scholar]
- Cabello-Olmo, M.; Araña, M.; Urtasun, R.; Encio, I.J.; Barajas, M. Role of Postbiotics in Diabetes Mellitus: Current Knowledge and Future Perspectives. Foods 2021, 10, 1590. [Google Scholar] [CrossRef]
- Zhao, M.; Tao, J.; Du, L.; Jiang, S.; Qian, D.; Duan, J. UPLC-Q-TOF/MS-Based Metabolic Profiling Comparison of Two Major Bioactive Components and Their Metabolites in Normal and CKD Rat Plasma, Urine and Feces Following Oral Administration of Fructus Corni Extract. J. Chromatogr. Sci. 2017, 55, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Barros, B.R.; Do Nascimento, D.K.D.; De Araújo, D.R.C.; Da Costa Batista, F.R.; De Oliveira Lima, A.M.N.; Da Cruz Filho, I.J.; De Oliveira, M.L.; De Melo, C.M.L. Phytochemical analysis, nutritional profile and immunostimulatory activity of aqueous extract from Malpighia emarginata DC leaves. Biocatal. Agric. Biotechnol. 2020, 23, 101442. [Google Scholar] [CrossRef]
- Sionek, B.; Szydłowska, A.; Zielińska, D.; Neffe-Skocińska, K.; Kołożyn-Krajewska, D. Beneficial Bacteria Isolated from Food in Relation to the Next Generation of Probiotics. Microorganisms 2023, 11, 1714. [Google Scholar] [CrossRef]
- Belizário, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef]
- Toda, K.; Hisata, K.; Satoh, T.; Katsumata, N.; Odamaki, T.; Mitsuyama, E.; Katayama, T.; Kuhara, T.; Aisaka, K.; Shimizu, T.; et al. Neonatal oral fluid as a transmission route for bifidobacteria to the infant gut immediately after birth. Sci. Rep. 2019, 9, 8692. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid. Based Complement Alternat. Med. 2018, 2018, 1756308. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Pyclik, M.; Srutkova, D.; Schwarzer, M.; Górska, S. Bifidobacteria cell wall-derived exo-polysaccharides, lipoteichoic acids, peptidoglycans, polar lipids and proteins—Their chemical structure and biological attributes. Int. J. Biol. Macromol. 2020, 147, 333–349. [Google Scholar] [CrossRef]
- Hernández-Granados, M.J.; Franco-Robles, E. Postbiotics in human health: Possible new functional ingredients? Food Res. Int. 2020, 137, 109660. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; De los Reyes-Gavilán, C.G. Invited review: Methods for the screening, isolation, and characterization of exopolysaccharides produced by lactic acid bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, H.Y.; Lee, S.; Lee, J.M.; Muthaiya, M.J.; Kim, B.S.; Oh, J.Y.; Song, C.K.; Jeon, E.J.; Ryu, H.S.; et al. Mass spectrometry-based metabolite profiling and bacterial diversity characterization of Korean traditional meju during fermentation. J. Microbiol. Biotechnol. 2012, 22, 1523–1531. [Google Scholar] [CrossRef]
- Garnier, L.; Penland, M.; Thierry, A.; Maillard, M.B.; Jardin, J.; Coton, M.; Valence, F.; Mounier, J. Antifungal activity of fermented dairy ingredients: Identification of antifungal compounds. Int. J. Food Microbiol. 2020, 322, 108574. [Google Scholar] [CrossRef]
- Lin, W.; Sun, F.; Zhang, Y.; Xu, X.; Lu, X.; Li, L.; Xu, R. Comparative transcriptome and metabolite profiling of four tissues from Alisma orientale (Sam.) Juzep reveals its inflorescence developmental and medicinal characteristics. Sci. Rep. 2019, 9, 12310. [Google Scholar] [CrossRef]
- Lim, J.H.; Lee, C.H.; Kim, K.Y.; Jung, H.Y.; Choi, J.Y.; Cho, J.H.; Park, S.H.; Kim, Y.L.; Baek, M.C.; Park, J.B.; et al. Novel urinary exosomal biomarkers of acute T cell-mediated rejection in kidney transplant recipients: A cross-sectional study. PLoS ONE 2018, 13, e0204204. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, G.; Zhou, J.; Pei, J.; Zhang, Q.; Song, X.; Sun, Z. Development and validation of a UPLC-MS/MS method for quantitation of droxidopa in human plasma: Application to a pharmacokinetic study. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1027, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Saharan, B.S.; Chauhan, N.; Procha, S.; Lal, S. Isolation and functional characterization of novel biosurfactant produced by Enterococcus faecium. Springerplus 2015, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, Y.; Wang, J.; Zhang, H.; Qi, W. Production and characterization of antifungal compounds produced by Lactobacillus plantarum IMAU10014. PLoS ONE 2012, 7, e29452. [Google Scholar] [CrossRef] [PubMed]
- Rühmann, B.; Schmid, J.; Sieber, V. Methods to identify the unexplored diversity of microbial exopolysaccharides. Front. Microbiol. 2015, 6, 565. [Google Scholar]
- Lin, T.H.; Pan, T.M. Characterization of an antimicrobial substance produced by Lactobacillus plantarum NTU 102. J. Microbiol. Immunol. Infec. 2019, 52, 409–417. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A.; Rejiniemon, T.S.; Lee, K.D.; Huxley, V.A.J.; Kim, D.H.; Duraipandiyan, V.; Karuppiah, P.; Choi, K.C. Identification and Characterization of Lactobacillus brevis P68 with Antifungal, Antioxidant and Probiotic Functional Properties. Indian J. Microbiol. 2015, 55, 19–28. [Google Scholar] [CrossRef]
- Ricci, A.; Levante, A.; Cirlini, M.; Calani, L.; Bernini, V.; Del Rio, D.; Galaverna, G.; Neviani, E.; Lazzi, C. The Influence of Viable Cells and Cell-Free Extracts of. Front. Microbiol. 2018, 9, 2784. [Google Scholar] [CrossRef]
- Li, X.; Zhu, L.; Wang, X.; Li, J.; Tang, B. Evaluation of irbiotyper for lactiplantibacillus plantarum typing and its application potential in probiotic preliminary screening. Front. Microbiol. 2022, 13, 823120. [Google Scholar]
- Malashree, L.; Vishwanath Angadi, K.; Yadav, S.; Prabha, R. “Postbiotics”—One Step Ahead of Probiotics. Int.J.Curr.Microbiol.App.Sci. 2019, 8, 2049–2053. [Google Scholar] [CrossRef]
- Sugahara, M.; Tanaka, T.; Nangaku, M. Prolyl hydroxylase domain inhibitors as a novel therapeutic approach against anemia in chronic kidney disease. Kidney Int. 2017, 92, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic Activity of Lactobacillus reuteri Strains on the Adhesion Characteristics of Selected Pathogens. Front.Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [PubMed]
- Kapustian, V.; Namazov, A.; Yaakov, O.; Volodarsky, M.; Anteby, E.Y.; Gemer, O. Is intrauterine device a risk factor for failure of conservative management in patients with tubo-ovarian abscess? An observational retrospective study. Arch. Gynecol. Obstet. 2018, 297, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr. Opin. Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, Y.; Kim, J.I.; Lee, H.Y.; Moon, G.S.; Kang, C.H. Improvements in Human Keratinocytes and Antimicrobial Effect Mediated by Cell-Free Supernatants Derived from Probiotics. Fermentation 2022, 8, 332. [Google Scholar] [CrossRef]
- Mani-López, E.; Arrioja-Bretón, D.; López-Malo, A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr. Rev. Food. Sci. Food Saf. 2022, 21, 604–641. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Kim, S.H. Anti-inflammatory and Anti-osteoporotic Potential of Lactobacillus plantarum A41 and L. fermentum SRK414 as Probiotics. Probiotics Antimicrob. Proteins 2020, 12, 623–634. [Google Scholar] [CrossRef]
- Hamad, G.M.; Abdelmotilib, N.M.; Darwish, A.M.G.; Zeitoun, A.M. Commercial probiotic cell-free supernatants for inhibition of Clostridium perfringens poultry meat infection in Egypt. Anaerobe 2020, 62, 102181. [Google Scholar] [CrossRef]
- Okafor, U.G.; Nwachukwu, M.; Anosike, E.M. Screening and identification of predominant Lactobacillus spp. from fermented milled-mixed corn-soybean wastes. Int. J. Sci. Eng. Res. 2018, 9, 2050–2060. [Google Scholar]
- Moll, G.N.; Konings, W.N.; Driessen, A.J. Bacteriocins: Mechanism of membrane insertion and pore formation. Antonie Van Leeuwenhoek 1999, 76, 185–198. [Google Scholar] [CrossRef]
- Hartmann, H.A.; Wilke, T.; Erdmann, R. Efficacy of bacteriocin-containing cell-free culture supernatants from lactic acid bacteria to control Listeria monocytogenes in food. Int. J. Food Microbiol. 2011, 146, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, D.L.M.; Manzo, R.M.; Tonarelli, G.G.; Simonetta, A.C. Characterisation of a cell-free supernatant obtained from cultures of Enterococcus faecalis DBFIQ E24 with antagonistic activity against bacteria, yeasts and moulds. Int. J. Dairy Technol. 2012, 65, 568–577. [Google Scholar] [CrossRef]
- Khodaii, Z.; Ghaderian, S.M.H.; Natanzi, M.M. Probiotic Bacteria and their Supernatants Protect Enterocyte Cell Lines from Enteroinvasive. Int. J. Mol. Cell Med. 2017, 6, 183–189. [Google Scholar]
- Yoon, I.; Lee, K.H.; Cho, J. Gossypin protects primary cultured rat cortical cells from oxidative stress- and beta-amyloid-induced toxicity. Arch. Pharm. Res. 2004, 27, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, X.; Fu, Z.; Wang, Z.; Zhang, J. Sulphated modification of a polysaccharide obtained from fresh persimmon (Diospyros kaki L.) fruit and antioxidant activities of the sulphated derivatives. Food Chem. 2011, 127, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Van der Es, D.; Hogendorf, W.F.; Overkleeft, H.S.; Van der Marel, G.A.; Codée, J.D. Teichoic acids: Synthesis and applications. Chem. Soc. Rev. 2017, 46, 1464–1482. [Google Scholar] [CrossRef]
- Saeui, C.T.; Mathew, M.P.; Liu, L.; Urias, E.; Yarema, K.J. Cell Surface and Membrane Engineering: Emerging Technologies and Applications. J. Funct. Biomater. 2015, 6, 454–485. [Google Scholar] [CrossRef]
- Jung, B.J.; Kim, H.; Chung, D.K. Differential Immunostimulatory Effects of Lipoteichoic Acids Isolated from Four Strains of Lactiplantibacillus plantarum. Appl. Sci. 2022, 12, 954. [Google Scholar] [CrossRef]
- Brown, S.; Santa Maria, J.P.; Walker, S. Wall teichoic acids of gram-positive bacteria. Annu. Rev. Microbiol. 2013, 67, 313–336. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.; Vanderleyden, J. Anti-inflammatory potential of probiotics: Lipoteichoic acid makes a difference. Trends Microbiol. 2012, 20, 5–10. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef] [PubMed]
- Dinić, M.; Pecikoza, U.; Djokić, J.; Stepanović-Petrović, R.; Milenković, M.; Stevanović, M.; Filipović, N.; Begović, J.; Golić, N.; Lukić, J. Exopolysaccharide Produced by Probiotic Strain Lactobacillus paraplantarum BGCG11 Reduces Inflammatory Hyperalgesia in Rats. Front. Pharmacol. 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Human-derived probiotic Lactobacillus reuteri strains differentially reduce intestinal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1087–G1096. [Google Scholar] [CrossRef] [PubMed]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Moscovici, M. Present and future medical applications of microbial exopolysaccharides. Front. Microbiol. 2015, 6, 1012. [Google Scholar] [CrossRef] [PubMed]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Gezginç, Y.; Karabekmez-Erdem, T.; Tatar, H.D.; Ayman, S.; Ganiyusufoğlu, E.; Dayisoylu, K.S. Health promoting benefitsof postbiotics produced by lactic acid bacteria: Exopolysaccharide. Biotech Stud. 2022, 31, 61–70. [Google Scholar] [CrossRef]
- Khalil, E.S.; Abd Manap, M.Y.; Mustafa, S.; Alhelli, A.M.; Shokryazdan, P. Probiotic properties of exopolysaccharide-producing Lactobacillus strains isolated from tempoyak. Molecules 2018, 23, 398. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Pandey, A.; Ganansounou, E. Genetic modification: A tool for enhancing beta-glucosidase production for biofuel application. Bioresour. Technol. 2017, 245, 1352–1361. [Google Scholar] [CrossRef]
- Kumar, B.; Gupta, V.P.; Kumar, V. A Perspective on Monoamine Oxidase Enzyme as Drug Target: Challenges and Opportunities. Curr. Drug Targets 2017, 18, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, K.J.; Cho, K.S.; Nam, S.W.; Park, T.J.; Bajpai, R. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Contesini, F.J.; Melo, R.R.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef] [PubMed]
- De Moreno de LeBlanc, A.; LeBlanc, J.G.; Perdigón, G.; Miyoshi, A.; Langella, P.; Azevedo, V.; Sesma, F. Oral administration of a catalase-producing Lactococcus lactis can prevent a chemically induced colon cancer in mice. J. Med. Microbiol. 2008, 57, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Friesen, L.; Park, J.; Kim, H.M.; Kim, C.H. Microbial metabolites, short-chain fatty acids, restrain tissue bacterial load, chronic inflammation, and associated cancer in the colon of mice. Eur. J. Immunol. 2018, 48, 1235–1247. [Google Scholar] [CrossRef]
- Takagi, R.; Sasaki, K.; Sasaki, D.; Fukuda, I.; Tanaka, K.; Yoshida, K.; Kondo, A.; Osawa, R. A Single-Batch Fermentation System to Simulate Human Colonic Microbiota for High-Throughput Evaluation of Prebiotics. PLoS ONE 2016, 11, e0160533. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics-A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Kasahara, Y.; Sakurai, T.; Matsuda, R.; Narukawa, M.; Yasuoka, A.; Mori, N.; Watanabe, H.; Okabe, T.; Kojima, H.; Abe, K.; et al. Novel indole and benzothiophene ring derivatives showing differential modulatory activity against human epithelial sodium channel subunits, ENaC β and γ. Biosci. Biotechnol. Biochem. 2019, 83, 243–250. [Google Scholar] [CrossRef]
- Higashi, B.; Mariano, T.B.; De Abreu Filho, B.A.; Gonçalves, R.A.C.; De Oliveira, A.J.B. Effects of fructans and probiotics on the inhibition of Klebsiella oxytoca and the production of short-chain fatty acids assessed by NMR spectroscopy. Carbohydr. Polym. 2020, 248, 116832. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Bird, A.R.; Conlon, M.A.; Christophersen, C.T.; Topping, D.L. Resistant starch, large bowel fermentation and a broader perspective of prebiotics and probiotics. Benef. Microbes 2010, 1, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Rowland, C.; Hanratty, B.; Pilling, M.; van den Berg, B.; Grande, G. The contributions of family care-givers at end of life: A national post-bereavement census survey of cancer carers’ hours of care and expenditures. Palliat. Med. 2017, 31, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Fact. 2014, 13, S3. [Google Scholar] [CrossRef]
- Belguesmia, Y.; Bendjeddou, K.; Kempf, I.; Boukherroub, R.; Drider, D. Heterologous Biosynthesis of Five New Class II Bacteriocins From Lactobacillus paracasei CNCM I-5369 With Antagonistic Activity Against Pathogenic Escherichia coli Strains. Front. Microbiol. 2020, 11, 1198. [Google Scholar] [CrossRef]
- Kim, S.W.; Kang, S.I.; Shin, D.H.; Oh, S.Y.; Lee, C.W.; Yang, Y.; Son, Y.K.; Yang, H.S.; Lee, B.H.; An, H.J.; et al. Potential of Cell-Free Supernatant from Lactobacillus plantarum NIBR97, Including Novel Bacteriocins, as a Natural Alternative to Chemical Disinfectants. Pharmaceuticals 2020, 13, 266. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; De Giori, G.S.; Sesma, F.; Van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Mikkelse, K.; Prakash, M.D.; Kuol, N.; Nurgali, K.; Stojanovska, L.; Apostolopoulos, V. Anti-tumor Effects of Vitamin B2, B6, and B9 in Promonocytic Lymphoma Cells. Int. J. Mol. Sci. 2019, 20, 3763. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production Potency of Folate, Vitamin B(12), and Thiamine by Lactic Acid Bacteria Isolated from Japanese Pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef]
- Martín, R.; Olivares, M.; Marín, M.L.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Characterization of a Reuterin-Producing Lactobacillus coryniformis Strain Isolated from a Goat’s Milk Cheese. Int. J. Food Microbiol. 2005, 104, 267–277. [Google Scholar] [CrossRef]
- De Angelis, M.; Bottacini, F.; Fosso, B.; Kelleher, P.; Calasso, M.; Di Cagno, R.; Ventura, M.; Picardi, E.; van Sinderen, D.; Marco Gobbetti, M. Lactobacillus rossiae, a Vitamin B12 Producer, Represents a Metabolically Versatile Species within the Genus Lactobacillus. PLoS ONE 2014, 9, e107232. [Google Scholar] [CrossRef] [PubMed]
- Camelo-Castillo, A.; Rivera-Caravaca, J.M.; Orenes-Piñero, E.; Ramírez-Macías, I.; Roldán, V.; Lip, G.Y.H.; Marín, F. Gut Microbiota and the Quality of Oral Anticoagulation in Vitamin K Antagonists Users: A Review of Potential Implications. J. Clin. Med. 2021, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, e1900952. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hou, Y.; Wang, G.; Zheng, X.; Hao, H. Gut Microbial Metabolites of Aromatic Amino Acids as Signals in Host-Microbe Interplay. Trends Endocrinol. Metab. 2020, 31, 818–834. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.D.; Turnbaugh, P.J. Microbial Determinants of Biochemical Individuality and Their Impact on Toxicology and Pharmacology. Cell Metab. 2014, 20, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi Badi, S.; Moshiri, A.; Fateh, A.; Rahimi Jamnani, F.; Sarshar, M.; Vaziri, F.; Siadat, S.D. Microbiota-Derived Extracellular Vesicles as New Systemic Regulators. Front. Microbiol. 2017, 8, 1610. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles Influence Gut Permeability through the Regulation of Tight Junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- de Jong, B.; Barros, E.R.; Hoenderop, J.G.J.; Rigalli, J.P. Recent Advances in Extracellular Vesicles as Drug Delivery Systems and Their Potential in Precision Medicine. Pharmaceutics 2020, 12, 1006. [Google Scholar] [CrossRef]
- Molina-Tijeras, J.A.; Gálvez, J.; Rodríguez-Cabezas, M.E. The Immunomodulatory Properties of Extracellular Vesicles Derived from Probiotics: A Novel Approach for the Management of Gastrointestinal Diseases. Nutrients 2019, 11, 1038. [Google Scholar] [CrossRef]
- Damms-Machado, A.; Louis, S.; Schnitzer, A.; Volynets, V.; Rings, A.; Basrai, M.; Bischoff, S.C. Gut Permeability Is Related to Body Weight, Fatty Liver Disease, and Insulin Resistance in Obese Individuals Undergoing Weight Reduction. Am. J. Clin. Nutr. 2017, 105, 127–135. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and Intestinal Epithelium Controls Diet-induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Ashrafian, F.; Keshavarz Azizi Raftar, S.; Lari, A.; Shahryari, A.; Abdollahiyan, S.; Moradi, H.R.; Masoumi, M.; Davari, M.; Khatami, S.; Omrani, M.D.; et al. Extracellular Vesicles and Pasteurized Cells Derived from Akkermansia muciniphila Protect against High-Fat-Induced Obesity in Mice. Microb. Cell Fact. 2021, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz Azizi Raftar, S.; Ashrafian, F.; Yadegar, A.; Lari, A.; Moradi, H.R.; Shahriary, A.; Azimirad, M.; Alavifard, H.; Mohsenifar, Z.; Davari, M.; et al. The Protective Effects of Live and Pasteurized Akkermansia muciniphila and Its Extracellular Vesicles against HFD/CCl4-Induced Liver Injury. Microbiol. Spectr. 2021, 9, e0048421. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Thangaraj, P.; Das, J.; Kim, J.H. Antibacterial and Antibiofilm Effects of Pseudomonas aeruginosa-Derived Outer Membrane Vesicles against Streptococcus mutans. Heliyon 2023, 9, e22606. [Google Scholar] [CrossRef]
- Rad, A.H.; Abbasi, A.; Kafil, H.S.; Ganbarov, K. Potential Pharmaceutical and Food Applications of Postbiotics: A Review. Curr. Pharm. Biotechnol. 2020, 21, 1576–1587. [Google Scholar] [CrossRef]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef]
- Shin, H.S.; Park, S.Y.; Lee, D.K.; Kim, S.A.; An, H.M.; Kim, J.R.; Kim, M.J.; Cha, M.G.; Lee, S.W.; Kim, K.J.; et al. Hypocholesterolemic Effect of Sonication-killed Bifidobacterium longum Isolated from Healthy Adult Koreans in High Cholesterol Fed Rats. Arch. Pharm. Res. 2010, 33, 1425–1431. [Google Scholar] [CrossRef]
- Martorell, P.; Alvarez, B.; Llopis, S.; Navarro, V.; Ortiz, P.; Gonzalez, N.; Balaguer, F.; Rojas, A.; Chenoll, E.; Ramon, D.; et al. Heat-Treated Bifidobacterium longum CECT-7347: A Whole-Cell Postbiotic with Antioxidant, Anti-Inflammatory, and Gut-Barrier Protection Properties. Antioxidants 2021, 10, 536. [Google Scholar] [CrossRef]
- Balaguer, F.; Enrique, M.; Llopis, S.; Barrena, M.; Navarro, V.; Alvarez, B.; Chenoll, E.; Ramon, D.; Tortajada, M.; Martorell, P. Lipoteichoic Acid from Bifidobacterium animalis subsp. lactis BPL1: A Novel Postbiotic that Reduces Fat Deposition via IGF-1 Pathway. Microb. Biotechnol. 2021, 15, 805–816. [Google Scholar] [CrossRef]
- Inturri, R.; Molinaro, A.; Di Lorenzo, F.; Blandino, G.; Tomasello, B.; Hidalgo-Cantabrana, C.; De Castro, C.; Ruas-Madiedo, P. Chemical and Biological Properties of the Novel Exopolysaccharide Produced by a Probiotic Strain of Bifidobacterium longum. Carbohydr. Polym. 2017, 174, 1172–1180. [Google Scholar] [CrossRef]
- Inturri, R.; Mangano, K.; Santagati, M.; Intrieri, M.; Di Marco, R.; Blandino, G. Immunomodulatory Effects of Bifidobacterium longum W11 Produced Exopolysaccharide on Cytokine Production. Curr. Pharm. Biotechnol. 2017, 18, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, E.; Gleinser, M.; Molloy, E.; Groeger, D.; Frei, R.; Ferstl, R.; Rodriguez-Perez, N.; Ziegler, M.; Grant, R.; Moriarty, T.F.; et al. The Surface-Associated Exopolysaccharide of Bifidobacterium longum 35624 Plays an Essential Role in Dampening Host Proinflammatory Responses and Repressing Local T(H)17 Responses. Appl. Environ. Microb. 2016, 82, 7185–7196. [Google Scholar] [CrossRef] [PubMed]
- Sungur, T.; Aslim, B.; Karaaslan, C.; Aktas, B. Impact of Exopolysaccharides (EPSs) of Lactobacillus gasseri Strains Isolated from Human Vagina on Cervical Tumor Cells (HeLa). Anaerobe 2017, 47, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Bhat, B.; Bajaj, B.K. Hypocholesterolemic Potential and Bioactivity Spectrum of an Exopolysaccharide from a Probiotic Isolate Lactobacillus paracasei M7. Bioact. Carbohydr.Diet. Fibre 2019, 19, 100191. [Google Scholar] [CrossRef]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the Immunomodulatory Properties of Lactobacillus rhamnosus CRL1505 Peptidoglycan Common for All Lactobacilli during Respiratory Infection in Malnourished Mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.E.; Kim, H.; Chung, D.K. Lipoteichoic Acid Isolated from Lactobacillus plantarum Maintains Inflammatory Homeostasis through Regulation of Th1- and Th2-Induced Cytokines. J. Microbiol. Biotechnol. 2019, 29, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Matsuguchi, T.; Takagi, A.; Matsuzaki, T.; Nagaoka, M.; Ishikawa, K.; Yokokura, T.; Yoshikai, Y. Lipoteichoic Acids from Lactobacillus Strains Elicit Strong Tumor Necrosis Factor Alpha-Inducing Activities in Macrophages through Toll-Like Receptor 2. Clin. Diagn. Lab Immun. 2003, 10, 259–266. [Google Scholar]
- Wang, S.H.; Ahmadi, S.; Nagpal, R.; Jain, S.; Mishra, S.P.; Kavanagh, K.; Zhu, X.W.; Wang, Z.; McClain, D.A.; Kritchevsky, S.B.; et al. Lipoteichoic Acid from the Cell Wall of a Heat-Killed Lactobacillus paracasei D3-5 Ameliorates Aging-Related Leaky Gut, Inflammation, and Improves Physical and Cognitive Functions: From C. elegans to Mice. Geroscience 2020, 42, 333–352. [Google Scholar] [CrossRef]
- Osman, A.; El-Gazzar, N.; Almanaa, T.N.; El-Hadary, A.; Sitohy, M. Lipolytic Postbiotic from Lactobacillus paracasei Manages Metabolic Syndrome in Albino Wistar Rats. Molecules 2021, 26, 472. [Google Scholar] [CrossRef]
- Wang, Y.H.; Liu, Y.L.; Sidhu, A.; Ma, Z.H.; McClain, C.; Feng, W.K. Lactobacillus rhamnosus GG Culture Supernatant Ameliorates Acute Alcohol-Induced Intestinal Permeability and Liver Injury. Am. J. Physiol. Gastr. L 2012, 303, G32–G41. [Google Scholar] [CrossRef]
- Compare, D.; Rocco, A.; Coccoli, P.; Angrisani, D.; Sgamato, C.; Iovine, B.; Salvatore, U.; Nardone, G. Lactobacillus casei DG and Its Postbiotic Reduce the Inflammatory Mucosal Response: An Ex-Vivo Organ Culture Model of Post-Infectious Irritable Bowel Syndrome. BMC Gastroenterol. 2017, 17, 53. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, Y.B.; Wan, Y.; Hu, T.T.; Liu, L.T.; Yang, S.J.; Gong, Z.L.; Zeng, Q.; Wei, Y.; Yang, W.J.; et al. A Novel Postbiotic from Lactobacillus rhamnosus GG with a Beneficial Effect on Intestinal Barrier Function. Front. Microbiol. 2019, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, F.; Ishida, Y.; Sawada, D.; Ashida, N.; Sugawara, T.; Sakai, M.; Goto, T.; Kawada, T.; Fujiwara, S. Fragmented Lactic Acid Bacterial Cells Activate Peroxisome Proliferator-Activated Receptors and Ameliorate Dyslipidemia in Obese Mice. J. Agric. Food Chem. 2016, 64, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Zhang, X.; Tong, L.; Liu, Q.; Liang, X.; Bu, Y.; Gong, P.; Liu, T.; Zhang, L.; Xia, Y.; et al. Effect of Extracellular Vesicles Derived from Lactobacillus plantarum Q7 on Gut Microbiota and Ulcerative Colitis in Mice. Front. Immunol. 2021, 12, 777147. [Google Scholar] [CrossRef] [PubMed]
- Maghsood, F.; Mirshafiey, A.; Farahani, M.W.; Modarressi, M.H.; Jafari, P.; Motevaseli, E. Dual Effects of Cell-Free Supernatants from Lactobacillus acidophilus and Lactobacillus rhamnosus GG in Regulation of MMP-9 by Up-regulating TIMP-1 and Down-regulating CD147 in PMA-differentiated THP-1 Cells. Cell J. 2018, 19, 559. [Google Scholar] [PubMed]
- Barros, C.P.; Guimaraes, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and Postbiotics: Concepts and Potential Applications in Dairy Products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Mi, X.J.; Tran, T.H.M.; Park, H.R.; Xu, X.Y.; Subramaniyam, S.; Choi, H.S.; Kim, J.A.; Koh, S.C.; Kim, Y.J. Immune-Enhancing Effects of Postbiotic Produced by Bacillus velezensis Kh2-2 Isolated from Korea Foods. Food Res. Int. 2021, 152, 110911. [Google Scholar] [CrossRef]
- Ghoneim, M.A.M.; Hassan, A.I.; Mahmoud, M.G.; Asker, M.S. Effect of Polysaccharide from Bacillus subtilis sp. on Cardiovascular Diseases and Atherogenic Indices in Diabetic Rats. BMC Complem. Altern Med. 2016, 16, 112. [Google Scholar]
- Dahech, I.; Belghith, K.S.; Hamden, K.; Feki, A.; Belghith, H.; Mejdoub, H. Antidiabetic Activity of Levan Polysaccharide in Alloxan-Induced Diabetic Rats. Int. J. Biol. Macromol. 2011, 49, 742–746. [Google Scholar] [CrossRef]
- Gurunathan, S.; Ajmani, A.; Kim, J.H. Extracellular Nanovesicles Produced by Bacillus licheniformis: A Potential Anticancer Agent for Breast and Lung Cancer. Microb. Pathog. 2023, 185, 106396. [Google Scholar] [CrossRef]
- Jensen, G.S.; Cash, H.A.; Farmer, S.; Keller, D. Inactivated Probiotic Bacillus coagulans GBI-30 Induces Complex Immune Activating, Anti-Inflammatory, and Regenerative Markers In Vitro. J. Inflamm. Res. 2017, 10, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.Z.A.; Liu, S.Q. Fortifying Foods with Synbiotic and Postbiotic Preparations of the Probiotic Yeast, Saccharomyces boulardii. Curr. Opin. Food Sci. 2022, 43, 216–224. [Google Scholar] [CrossRef]
- Jensen, G.S.; Hart, A.N.; Schauss, A.G. An Antiinflammatory Immunogen from Yeast Culture Induces Activation and Alters Chemokine Receptor Expression on Human Natural Killer Cells and B Lymphocytes In Vitro. Nutr. Res. 2007, 27, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Jin, D.; Huang, S.; Wu, J.; Xu, M.; Liu, T.; Wang, S.; Zhong, W.; Liu, Y.; Jiang, R.; et al. Clostridium butyricum, a Butyrate-Producing Probiotic, Inhibits Intestinal Tumor Development through Modulating Wnt Signaling and Gut Microbiota. Cancer Lett. 2020, 469, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Laharie, D.; Asensio, C.; Vidal-Martinez, T.; Candalh, C.; Rullier, A.; Zerbib, F.; Mégraud, F.; Matysiak-Budnik, T.; Heyman, M. Bifidobacterium breve and Streptococcus thermophilus Secretion Products Enhance T Helper 1 Immune Response and Intestinal Barrier in Mice. Exp. Biol. Med. 2005, 230, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Tseng, S.C.; Wang, S.L. Production and Characterization of Antioxidant Properties of Exopolysaccharide(s) from Peanibacillusmucilaginosus TKU032. Mar. Drugs 2016, 14, 40. [Google Scholar] [CrossRef]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; De Vos, M.; Boon, N.; Van de Wiele, T. Butyrate-Producing Bacteria Supplemented In Vitro to Crohn’s Disease Patient Microbiota Increased Butyrate Production and Enhanced Intestinal Epithelial Barrier Integrity. Sci. Rep. 2017, 7, 11450. [Google Scholar] [CrossRef]
- Bhat, B.; Bajaj, B.K. Hypocholesterolemic and Bioactive Potential of Exopolysaccharide from a Probiotic Enterococcus faecium K1 Isolated from Kalarei. Bioresour. Technol. 2018, 254, 264–267. [Google Scholar] [CrossRef]
- Depommier, C.; Van Hul, M.; Everard, A.; Delzenne, N.M.; De Vos, W.M.; Cani, P.D. Pasteurized Akkermansia muciniphila Increases Whole-Body Energy Expenditure and Fecal Energy Excretion in Diet-Induced Obese Mice. Gut. Microbes 2020, 11, 1231–1245. [Google Scholar] [CrossRef]
- Jensen, B.A.; Holm, J.B.; Larsen, I.S.; von Burg, N.; Derer, S.; Sonne, S.B.; Pærregaard, S.I.; Damgaard, M.V.; Indrelid, S.A.; Rivollier, A.; et al. Lysates of Methylococcuscapsulatus Bath Induce a Lean-Like Microbiota, Intestinal FoxP3+ RORγt+ IL-17+ Tregs and Improve Metabolism. Nat. Commun. 2021, 12, 1093. [Google Scholar] [CrossRef]
- Durant, L.; Stentz, R.; Noble, A.; Brooks, J.; Gicheva, N.; Reddi, D.; O’Connor, M.J.; Hoyles, L.; McCartney, A.L.; Man, R.; et al. Bacteroides thetaiotaomicron-Derived Outer Membrane Vesicles Promote Regulatory Dendritic Cell Responses in Health but Not in Inflammatory Bowel Disease. Microbiome 2020, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Rong, X.; Zhao, G.; Zhou, Y.; Xiao, Y.; Ma, D.; Jin, X.; Wu, Y.; Yan, Y.; Yang, H.; et al. The Microbial Metabolite Trimethylamine N-Oxide Promotes Antitumor Immunity in Triple-Negative Breast Cancer. Cell Metab. 2022, 34, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Simon, M.V.; Lee, J.H.; Ustunol, Z.; Pestka, J.J. Ingestion of Yogurt Containing Lactobacillus acidophilus and Bifidobacterium to Potentiate Immunoglobulin A Responses to Cholera Toxin in Mice. J. Dairy Sci. 1999, 82, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Arshad, F.; Mehmood, R.; Hussain, S.; Khan, M.A.; Khan, M. Lactobacilli as Probiotics and Their Isolation from Different Sources. Br. J. Res. 2018, 5, 43. [Google Scholar] [CrossRef]
- Vidal, K.; Donnet-Hughes, A.; Granato, D. Lipoteichoic Acids from Lactobacillus johnsonii Strain La1 and Lactobacillus acidophilus Strain La10 Antagonize the Responsiveness of Human Intestinal Epithelial HT29 Cells to Lipopolysaccharide and Gram-Negative Bacteria. Infect. Immun. 2002, 70, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Nakagawa, E.; Nara, T.; Yajima, T.; Kuwata, T. Effects of Orally Ingested Bifidobacterium longum on the Mucosal IgA Response of Mice to Dietary Antigens. Biosci. Biotechnol. Biochem. 1998, 62, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hoover, D.G. Bacteriocins and Their Food Applications. Compr. Rev. Food Sci. Food Saf. 2003, 2, 82–100. [Google Scholar]
- Ahmad Rather, I.; Seo, B.J.; Rejish Kumar, V.J.; Choi, U.H.; Choi, K.H.; Lim, J.H.; Park, Y.H. Isolation and Characterization of a Proteinaceous Antifungal Compound from Lactobacillus plantarum YML007 and Its Application as a Food Preservative. Lett. Appl. Microbiol. 2013, 57, 69–76. [Google Scholar] [CrossRef]
- Tomasik, P.; Tomasik, P. Probiotics, Non-Dairy Prebiotics and Postbiotics in Nutrition. Appl. Sci. 2020, 10, 1470. [Google Scholar] [CrossRef]
- Arora, S.; Jood, S.; Khetarpaul, N. Effect of Germination and Probiotic Fermentation on Nutrient Profile of Pearl Millet Based Food Blends. Br. Food J. 2011, 113, 470–481. [Google Scholar] [CrossRef]
- Capozzi, V.; Russo, P.; Dueñas, M.T.; López, P.; Spano, G. Lactic Acid Bacteria Producing B-Group Vitamins: A Great Potential for Functional Cereal Products. Appl. Microbiol. Biotechnol. 2012, 96, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In Vitro Investigation of Bioactivities of Solid-State Fermented Lupin, Quinoa, and Wheat Using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Abbasi, A. Molecular Mechanisms of Postbiotics in Colorectal Cancer Prevention and Treatment. Crit. Rev. Food Sci. Nutr. 2021, 61, 1787–1803. [Google Scholar] [CrossRef]
- İncili, G.K.; Karatepe, P.; Akgöl, M.; Güngören, A.; Koluman, A.; İlhak, O.; Kanmaz, H.; Kaya, B.; Hayaloğlu, A.A. Characterization of Lactic Acid Bacteria Postbiotics, Evaluation of In-Vitro Antibacterial Effect, Microbial and Chemical Quality on Chicken Drumsticks. Food Microbiol. 2022, 104, 104001. [Google Scholar] [CrossRef] [PubMed]
- Chaney, W.E.; Naqvi, S.A.; Gutierrez, M.; Gernat, A.; Johnson, T.J.; Petry, D. Dietary Inclusion of A. Microorganisms 2022, 10, 544. [Google Scholar] [CrossRef] [PubMed]
- de Lima, A.L.; Guerra, C.A.; Costa, L.M.; de Oliveira, V.S.; Lemos Junior, W.J.F.; Luchese, R.H.; Guerra, A.F. A Natural Technology for Vacuum-Packaged Cooked Sausage Preservation with Potentially Postbiotic-Containing Preservative. Fermentation 2022, 8, 106. [Google Scholar] [CrossRef]
- Rad, A.H.; Hosseini, S.; Pourjafar, H. Postbiotics as Dynamic Biological Molecules for Antimicrobial Activity: A Mini-Review. Biointerface Res.Appl. Chem. 2022, 12, 6543–6556. [Google Scholar]
- Fong, W.; Li, Q.; Yu, J. Gut Microbiota Modulation: A Novel Strategy for Prevention and Treatment of Colorectal Cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef]
- Torino, M.I.; Font de Valdez, G.; Mozzi, F. Biopolymers from Lactic Acid Bacteria. Novel Applications in Foods and Beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Metabolism and Interaction with Food Components. Int. J. Mol. Sci. 2020, 21, 3688. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Abdali, D.; Samson, S.E.; Grover, A.K. How Effective Are Antioxidant Supplements in Obesity and Diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, S.; Abe, F.; Ishibashi, N.; Miyakawa, H.; Yaeshima, T.A.; Tomita, M. Relationship Between Oxygen Sensitivity and Oxygen Metabolism of Bifidobacterium Species. J. Dairy Sci. 1992, 75, 3296–3306. [Google Scholar] [CrossRef] [PubMed]
- Tomusiak-Plebanek, A.; Heczko, P.; Skowron, B.; Baranowska, A.; Okoń, K.; Thor, P.J.; Strus, M. Lactobacilli with Superoxide Dismutase-Like or Catalase Activity Are More Effective in Alleviating Inflammation in an Inflammatory Bowel Disease Mouse Model. Drug Des. Dev. Ther. 2018, 12, 3221–3233. [Google Scholar] [CrossRef] [PubMed]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two Antioxidative Lactobacilli Strains as Promising Probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Cousin, F.J.; Jouan-Lanhouet, S.; Dimanche-Boitrel, M.T.; Corcos, L.; Jan, G. Milk Fermented by Propionibacterium freudenreichii Induces Apoptosis of HGT-1 Human Gastric Cancer Cells. PLoS ONE 2012, 7, e31892. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from Probiotic Lactobacillus casei and Lactobacillus rhamnosus GG Decrease Colon Cancer Cell Invasion In Vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Chuah, L.O.; Foo, H.L.; Loh, T.C.; Mohammed Alitheen, N.B.; Yeap, S.K.; Abdul Mutalib, N.E.; Yusoff, K. Postbiotic Metabolites Produced by Lactobacillus plantarum Strains Exert Selective Cytotoxicity Effects on Cancer Cells. BMC Complement. Altern. Med. 2019, 19, 1–12. [Google Scholar]
- Shida, K.; Kiyoshima-Shibata, J.; Kaji, R.; Nagaoka, M.; Nanno, M. Peptidoglycan from Lactobacilli Inhibits Interleukin-12 Production by Macrophages Induced by Lactobacillus casei through Toll-Like Receptor 2-Dependent and Independent Mechanisms. Immunology 2009, 128, e858–e869. [Google Scholar] [CrossRef]
- Dahech, I.; Belghith, K.S.; Hamden, K.; Feki, A.; Belghith, H.; Mejdoub, H. Oral administration of levan polysaccharide reduces the alloxan-induced oxidative stress in rats. Int. J. Biol. Macromol. 2011, 49, 942–947. [Google Scholar] [CrossRef]
- Mutavdzin, S.; Gopcevic, K.; Stankovic, S.; Jakovljevic Uzelac, J.; LabudovicBorovic, M.; Djuric, D. The Effects of Folic Acid Administration on Cardiac Oxidative Stress and Cardiovascular Biomarkers in Diabetic Rats. Oxid. Med. Cell. Longev. 2019, 2019, 1342549. [Google Scholar] [CrossRef] [PubMed]
- Salimi, F.; Farrokh, P. Recent advances in the biological activities of microbial exopolysaccharides. World J. Microbiol. Biotechnol. 2023, 39, 213. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.D.; Shi, G.; Jiang, Y.; Yao, C.; Zhu, C. A Review on the Potential of Resveratrol in Prevention and Therapy of Diabetes and Diabetic Complications. Biomed. Pharmacother. 2020, 125, 109767. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zheng, J.; Wang, S.; Lau, H.K.; Fathi, A.; Wang, Q. Cardiovascular Benefits of Native GLP-1 and Its Metabolites: An Indicator for GLP-1-Therapy Strategies. Front. Physiol. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Heeney, D.D.; Zhai, Z.; Bendiks, Z.; Barouei, J.; Martinic, A.; Slupsky, C.; Marco, M.L. Lactobacillus plantarum Bacteriocin Is Associated with Intestinal and Systemic Improvements in Diet-Induced Obese Mice and Maintains Epithelial Barrier Integrity In Vitro. Gut. Microbes 2019, 10, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, J.F.; Schertzer, J.D. Intestinal Microbiota Contributes to Energy Balance, Metabolic Inflammation, and Insulin Resistance in Obesity. J. Obes. Metab. Syndr. 2017, 26, 161. [Google Scholar] [CrossRef] [PubMed]
- Hedl, M.; Li, J.; Cho, J.H.; Abraham, C. Chronic Stimulation of Nod2 Mediates Tolerance to Bacterial Products. Proc. Natl. Acad. Sci. USA 2007, 104, 19440–19445. [Google Scholar] [CrossRef]
- Denou, E.; Lolmède, K.; Garidou, L.; Pomie, C.; Chabo, C.; Lau, T.C.; Fullerton, M.D.; Nigro, G.; Zakaroff-Girard, A.; Luche, E.; et al. Defective NOD2 Peptidoglycan Sensing Promotes Diet-Induced Inflammation, Dysbiosis, and Insulin Resistance. EMBO Mol. Med. 2015, 7, 259–274. [Google Scholar] [CrossRef]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nuñez, G.; Flavell, R.A. Nod2-Dependent Regulation of Innate and Adaptive Immunity in the Intestinal Tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef]
- Huo, Y.; Lu, X.; Wang, X.; Wang, X.; Chen, L.; Guo, H.; Zhang, M.; Li, Y. Bifidobacterium animalis subsp. lactis A6 Alleviates Obesity Associated with Promoting Mitochondrial Biogenesis and Function of Adipose Tissue in Mice. Molecules 2020, 25, 1490. [Google Scholar]
- Gill, H.S.; Rutherfurd, K.J. Probiotic Supplementation to Enhance Natural Immunity in the Elderly: Effects of a Newly Characterized Immunostimulatory Strain Lactobacillus rhamnosus HN001 (DR20™) on Leucocyte Phagocytosis. Nutr. Res. 2001, 21, 183–189. [Google Scholar] [CrossRef]
- Ou, C.C.; Lin, S.L.; Tsai, J.J.; Lin, M.Y. Heat-Killed Lactic Acid Bacteria Enhance Immunomodulatory Potential by Skewing the Immune Response toward Th1 Polarization. J. Food Sci. 2011, 76, M260–M267. [Google Scholar] [CrossRef]
- Wu, S.; He, X.; Wu, X.; Qin, S.; He, J.; Zhang, S.; Hou, D.X. Inhibitory Effects of Blue Honeysuckle (Lonicera caerulea L.) on Adjuvant-Induced Arthritis in Rats: Crosstalk of Anti-Inflammatory and Antioxidant Effects. J. Funct. Foods 2015, 17, 514–523. [Google Scholar] [CrossRef]
- Marcial, G.; Villena, J.; Faller, G.; Hensel, A.; de Valdéz, G.F. Exopolysaccharide-producing Streptococcus thermophilus CRL1190 Reduces the Inflammatory Response Caused by Helicobacter pylori. Benef. Microbes 2017, 8, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Generoso, S.V.; Viana, M.L.; Santos, R.G.; Arantes, R.M.; Martins, F.S.; Nicoli, J.R.; Cardoso, V.N. Protection against Increased Intestinal Permeability and Bacterial Translocation Induced by Intestinal Obstruction in Mice Treated with Viable and Heat-Killed Saccharomyces boulardii. Eur. J. Nutr. 2011, 50, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zagato, E.; Mileti, E.; Massimiliano, L.; Fasano, F.; Budelli, A.; Penna, G.; Rescigno, M. Lactobacillus paracasei CBA L74 Metabolic Products and Fermented Milk for Infant Formula Have Anti-Inflammatory Activity on Dendritic Cells In Vitro and Protective Effects against Colitis and an Enteric Pathogen In Vivo. PLoS ONE 2014, 9, e87615. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Liu, F.; Chen, X.; Chen, M.; Deng, Q.; Zou, X.; Guo, H. Crocetin Protects against Fulminant Hepatic Failure Induced by Lipopolysaccharide/D Galactosamine by Decreasing Apoptosis, Inflammation and Oxidative Stress in a Rat Model. Exp. Ther. Med. 2019, 18, 3775–3782. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus casei CCFM419 Attenuates Type 2 Diabetes via a Gut Microbiota Dependent Mechanism. Food Funct. 2017, 8, 3155–3164. [Google Scholar] [CrossRef]
- Kamilya, D.; Baruah, A.; Sangma, T.; Chowdhury, S.; Pal, P. Inactivated Probiotic Bacteria Stimulate Cellular Immune Responses of Catla, Catla (Hamilton) In Vitro. Probiotics Antimicrob. Proteins 2015, 7, 101–106. [Google Scholar]
- Chen, Y.; Liu, W.; Sun, T.; Huang, Y.; Wang, Y.; Deb, D.K.; Yoon, D.; Kong, J.; Thadhani, R.; Li, Y.C. 1, 25-Dihydroxyvitamin D Promotes Negative Feedback Regulation of TLR Signaling via Targeting MicroRNA-155–SOCS1 in Macrophages. J. Immunol. 2013, 190, 3687–3695. [Google Scholar] [CrossRef]
- Akimova, T.; Beier, U.H.; Liu, Y.; Wang, L.; Hancock, W.W. Histone/Protein Deacetylases and T-Cell Immune Responses. BloodJ. Am. Soc. Hematol. 2012, 119, 2443–2451. [Google Scholar] [CrossRef]
- Griet, M.; Zelaya, H.; Mateos, M.V.; Salva, S.; Juarez, G.E.; de Valdez, G.F.; Villena, J.; Salvador, G.A.; Rodriguez, A.V. Soluble Factors from Lactobacillus reuteri CRL1098 Have Anti-Inflammatory Effects in Acute Lung Injury Induced by Lipopolysaccharide in Mice. PLoS ONE 2014, 9, e110027. [Google Scholar] [CrossRef]
- Sougioultzis, S.; Foukas, P.G.; Tzivras, M.; Kourtessas, D.; Gorgoulis, V.G.; Davaris, P.; Archimandritis, A.J. Alterations in the Proliferating Compartment of Gastric Mucosa during Helicobacter pylori Infection: The Putative Role of Epithelial Cells Expressing p27kip1. Mod. Pathol. 2003, 16, 1076–1085. [Google Scholar] [CrossRef]
- Baud, D.; Dimopoulou Agri, V.; Gibson, G.R.; Reid, G.; Giannoni, E. Using Probiotics to Flatten the Curve of Coronavirus Disease COVID-2019 Pandemic. Front. Public Health 2020, 8, 186. [Google Scholar] [CrossRef]
- Sundararaman, A.; Ray, M.; Ravindra, P.V.; Halami, P.M. Role of Probiotics to Combat Viral Infections with Emphasis on COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 8089–8104. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host Cell Death and Inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef]
- Wu, F.; Wang, A.; Liu, M.; Wang, Q.; Chen, J.; Xia, S.; Ling, Y.; Zhang, Y.; Xun, J.; Lu, L.; et al. Neutralizing Antibody Responses to SARS-CoV-2 in a COVID-19 Recovered Patient Cohort and Their Implications. MedRxiv 2020. [Google Scholar] [CrossRef]
- Ansari, F.; Pourjafar, H.; Tabrizi, A.; Homayouni, A. The Effects of Probiotics and Prebiotics on Mental Disorders: A Review on Depression, Anxiety, Alzheimer, and Autism Spectrum Disorders. Curr. Pharm. Biotechnol. 2020, 21, 555–565. [Google Scholar] [CrossRef]
- Negreira-Caamaño, M.; Piqueras-Flores, J.; Martínez-DelRio, J.; Nieto-Sandoval-Martin-DeLaSierra, P.; Aguila-Gordo, D.; Mateo-Gomez, C.; Negreira-Caamaño, M. Impact of Treatment with Renin–Angiotensin System Inhibitors on Clinical Outcomes in Hypertensive Patients Hospitalized with COVID-19. High Blood Press. Cardiovasc. Prev. 2020, 27, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Kalinichenko, S.V.; Melentyeva, K.V.; Manee, H.; Dubinina, N.V.; Zvereva, N.V.; Toryanik, I.I.; Popova, N.G.; Pakhomov, O.V. Study of Antivirus Actions of Metabolites of Lactobacteria. Wiad Lek. 2020, 73, 1484–1488. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M. Identification of Potential SARS-CoV-2 Entry Inhibitors by Targeting the Interface Region Between the Spike RBD and Human ACE2. J. Infect. Public Health 2021, 14, 227–237. [Google Scholar] [CrossRef]
- Mali, J.K.; Sutar, Y.B.; Pahelkar, A.R.; Verma, P.M.; Telvekar, V.N. Novel Fatty Acid-Thiadiazole Derivatives as Potential Antimycobacterial Agents. Chem. Biol. Drug Des. 2020, 95, 174–181. [Google Scholar] [CrossRef]
- Elnagdy, S.; AlKhazindar, M. The Potential of Antimicrobial Peptides as an Antiviral Therapy Against COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 780–782. [Google Scholar] [CrossRef]
- Mani-López, E.; García, H.; López-Malo, A. Organic Acids as Antimicrobials to Control Salmonella in Meat and Poultry Products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Gálvez, A.; Abriouel, H.; López, R.L.; Omar, N.B. Bacteriocin-Based Strategies for Food Biopreservation. Int. J. Food Microbiol. 2007, 120, 51–70. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, H.W. Innate Immunity Induced by Fungal β-Glucans via Dectin-1 Signaling Pathway. Int. J. Med. Mushrooms 2014, 16, 1–16. [Google Scholar] [CrossRef]
- O’Connor, P.M.; Kuniyoshi, T.M.; Oliveira, R.P.; Hill, C.; Ross, R.P.; Cotter, P.D. Antimicrobials for Food and Feed; A Bacteriocin Perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostalova, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and Remarkable Specificity of Antimicrobial Peptides In Vivo Using a Systematic Knockout Approach. eLife 2019, 8, e44341. [Google Scholar] [CrossRef]
- Karimi, N.; Jabbari, V.; Nazemi, A.; Ganbarov, K.; Karimi, N.; Tanomand, A.; Karimi, S.; Abbasi, A.; Yousefi, B.; Khodadadi, E.; et al. Thymol, Cardamom, and Lactobacillus plantarum Nanoparticles as a Functional Candy with High Protection against Streptococcus mutans and Tooth Decay. Microb. Pathog. 2020, 148, 104481. [Google Scholar] [CrossRef]
- Nowak, A.; Zakłos-Szyda, M.; Rosicka-Kaczmarek, J.; Motyl, I. Anticancer Potential of Post-Fermentation Media and Cell Extracts of Probiotic Strains: An In Vitro Study. Cancers 2022, 14, 1853. [Google Scholar] [CrossRef]
- Jan, G.B.A.S.; Belzacq, A.S.; Haouzi, D.; Rouault, A.; Metivier, D.; Kroemer, G.; Brenner, C. Propionibacteria Induce Apoptosis of Colorectal Carcinoma Cells via Short-Chain Fatty Acids Acting on Mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef]
- Lee, J.W.; Shin, J.G.; Kim, E.H.; Kang, H.E.; Yim, I.B.; Kim, J.Y.; Joo, H.G.; Woo, H.J. Immunomodulatory and Antitumor Effects In Vivo by the Cytoplasmic Fraction of Lactobacillus casei and Bifidobacterium longum. J. Vet. Sci. 2004, 5, 41–48. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, K.; Ding, J.; Xu, H.; Zhu, L.; Zhang, K.; Li, X.; Sun, W. Harmine Induces Apoptosis and Inhibits Tumor Cell Proliferation, Migration, and Invasion Through Down-Regulation of Cyclooxygenase-2 Expression in Gastric Cancer. Phytomedicine 2014, 21, 348–355. [Google Scholar] [CrossRef]
- Casanova, M.R.; Azevedo-Silva, J.; Rodrigues, L.R.; Preto, A. Colorectal Cancer Cells Increase the Production of Short Chain Fatty Acids by Propionibacterium freudenreichii Impacting on Cancer Cells Survival. Front. Nutr. 2018, 5, 44. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Goudarzi, H.; Ghalavand, Z.; Hajikhani, B.; Rafeieiatani, Z.; Hakemi-Vala, M. Anti-Proliferative Effects of Cell Wall, Cytoplasmic Extract of Lactococcus lactis and Nisin Through Down-Regulation of Cyclin D1 on SW480 Colorectal Cancer Cell Line. Iran. J. Microbiol. 2020, 12, 424. [Google Scholar] [CrossRef]
- An, B.C.; Ahn, J.Y.; Kwon, D.; Kwak, S.H.; Heo, J.Y.; Kim, S.; Ryu, Y.; Chung, M.J. Anti-Cancer Roles of Probiotic-Derived P8 Protein in Colorectal Cancer Cell Line DLD-1. Int. J. Mol. Sci. 2023, 24, 9857. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, S.; Yun, H.S.; Oh, S.; Kim, S.H. Cell-Bound Exopolysaccharide from Probiotic Bacteria Induces Autophagic Cell Death of Tumor Cells. Lett. Appl. Microbiol. 2010, 51, 123–130. [Google Scholar]
- Kim, S.; Kim, G.H.; Cho, H. Postbiotics for Cancer Prevention and Treatment. Microb. Cell 2021, 57, 142–153. [Google Scholar]
- Nakkarach, A.; Foo, H.L.; Song, A.A.L.; Mutalib, N.E.A.; Nitisinprasert, S.; Withayagiat, U. Anti-Cancer and Anti-Inflammatory Effects Elicited by Short Chain Fatty Acids Produced by Escherichia coli Isolated from Healthy Human Gut Microbiota. Microb. Cell Fact. 2021, 20, 36. [Google Scholar] [CrossRef] [PubMed]
- Kotani, Y.; Kunisawa, J.; Suzuki, Y.; Sato, I.; Saito, T.; Toba, M.; Kohda, N.; Kiyono, H. Role of Lactobacillus pentosus Strain b240 and the Toll-like Receptor 2 Axis in Peyer’s Patch Dendritic Cell-Mediated Immunoglobulin A Enhancement. PLoS ONE 2014, 9, e91857. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Ye, K.; Lu, J.; Wang, X.; Zhang, S.; Liu, L.; Yang, B.; Nassar, K.; Xu, X.; Pang, X.; et al. Probiotic Strain Lactobacillus plantarum YYC-3 Prevents Colon Cancer in Mice by Regulating the Tumour Microenvironment. Biomed. Pharmacother. 2020, 127, 110159. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Walia, S.; Kondepudi, K.K.; Shukla, G. Administration of Indigenous Probiotics Modulate High-Fat Diet-Induced Metabolic Syndrome in Sprague Dawley Rats. Antonie Van Leeuwenhoek 2020, 113, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Molochek, N.; Savchuk, O.; Kyriienko, D.; Komisarenko, I. Probiotic and Omega-3 Polyunsaturated Fatty Acids Supplementation Reduces Insulin Resistance, Improves Glycemia, and Obesity Parameters in Individuals with Type 2 Diabetes: A Randomised Controlled Trial. Obes. Med. 2020, 19, 100248. [Google Scholar] [CrossRef]
- Hu, F.; Niu, Y.; Xu, X.; Hu, Q.; Su, Q.; Zhang, H. Resistant Dextrin Improves High-Fat-High-Fructose Diet Induced Insulin Resistance. Nutr. Metab. 2020, 17, 1–11. [Google Scholar] [CrossRef]
- Li, H.Y.; Zhou, D.D.; Gan, R.Y.; Huang, S.Y.; Zhao, C.N.; Shang, A.; Xu, X.Y.; Li, H.B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients 2021, 13, 3211. [Google Scholar] [CrossRef]
- Isozaki, S.; Konishi, H.; Fujiya, M.; Tanaka, H.; Murakami, Y.; Kashima, S.; Ando, K.; Ueno, N.; Moriichi, K.; Okumura, T. Probiotic-Derived Polyphosphate Accelerates Intestinal Epithelia Wound Healing through Inducing Platelet-Derived Mediators. Mediat. Inflamm. 2021, 2021, 5582943. [Google Scholar] [CrossRef]
- Cheng, D.; Xu, J.H.; Li, J.Y.; Wang, S.Y.; Wu, T.F.; Chen, Q.K.; Yu, T. Butyrate Ameliorated-NLRC3 Protects the Intestinal Barrier in a GPR43-Dependent Manner. Exp. Cell Res. 2018, 368, 101–110. [Google Scholar] [CrossRef]
- Ye, Y.; Barghouth, M.; Luan, C.; Kazim, A.; Zhou, Y.; Eliasson, L.; Zhang, E.; Hansson, O.; Thevenin, T.; Renström, E. The TCF7L2-Dependent High-Voltage Activated Calcium Channel Subunit α2δ-1 Controls Calcium Signaling in Rodent Pancreatic Beta-Cells. Mol. Cell. Endocrinol. 2020, 502, 110673. [Google Scholar] [CrossRef]
- McDaniel, K.; Hall, C.; Sato, K.; Lairmore, T.; Marzioni, M.; Glaser, S.; Meng, F.; Alpini, G. Lin28 and Let-7: Roles and Regulation in Liver Diseases. Am. J. Physiol.-Gastrointest. Liver Physiol. 2016, 310, G757–G765. [Google Scholar] [CrossRef] [PubMed]
- Atreya, R.; Neurath, M.F. Signaling Molecules: The Pathogenic Role of the IL-6/STAT-3 Trans Signaling Pathway in Intestinal Inflammation and in Colonic Cancer. Curr. Drug Targets 2008, 9, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.V.; Griet, M. NF-κB in Anti-inflammatory Activity of Probiotics: An Update. Curr. Immunol. Rev. 2016, 12, 74–82. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.; Zhang, H.; Yu, J.; Yao, Z. Oral Probiotic Administration during Pregnancy Prevents Autism-Related Behaviors in Offspring Induced by Maternal Immune Activation via Anti-inflammation in Mice. Autism Res. 2019, 12, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Jones, M.L.; Ganopolsky, J.G.; Labbé, A.; Prakash, S. A Novel Nitric Oxide Producing Probiotic Patch and Its Antimicrobial Efficacy: Preparation and In Vitro Analysis. Appl. Microbiol. Biotechnol. 2010, 87, 509–516. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Immune System Stimulation by Probiotic Microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Uchinaka, A.; Azuma, N.; Mizumoto, H.; Nakano, S.; Minamiya, M.; Yoneda, M.; Aoyama, K.; Komatsu, Y.; Yamada, Y.; Murohara, T.; et al. Anti-inflammatory Effects of Heat-killed Lactobacillus plantarum L-137 on Cardiac and Adipose Tissue in Rats with Metabolic Syndrome. Sci. Rep. 2018, 8, 8156. [Google Scholar] [CrossRef]
- Petrof, E.O.; Kojima, K.; Ropeleski, M.J.; Musch, M.W.; Tao, Y.; De Simone, C.; Chang, E.B. Probiotics Inhibit Nuclear Factor-κB and Induce Heat Shock Proteins in Colonic Epithelial Cells through Proteasome Inhibition. Gastroenterology 2004, 127, 1474–1487. [Google Scholar] [CrossRef]
- Heuvelin, E.; Lebreton, C.; Bichara, M.; Cerf-Bensussan, N.; Heyman, M. A Bifidobacterium Probiotic Strain and Its Soluble Factors Alleviate Chloride Secretion by Human Intestinal Epithelial Cells. J. Nutr. 2010, 140, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Kaci, G.; Lakhdari, O.; Doré, J.; Ehrlich, S.D.; Renault, P.; Blottière, H.M.; Delorme, C. Inhibition of the NF-κB Pathway in Human Intestinal Epithelial Cells by Commensal Streptococcus salivarius. Appl. Environ. Microbiol. 2011, 77, 4681–4684. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In Vitro Immunomodulatory Activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two Probiotic Strains Isolated from Human Breast Milk. Immunobiology 2010, 215, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Shin, H.; Park, J.Y.; Rho, J.G.; Son, D.H.; Kim, K.W.; Seong, J.K.; Yoon, S.H.; Kim, W. A Novel Peripheral Cannabinoid 1 Receptor Antagonist, AJ5012, Improves Metabolic Outcomes and Suppresses Adipose Tissue Inflammation in Obese Mice. FASEB J. 2019, 33, 4314–4326. [Google Scholar] [CrossRef] [PubMed]
- Palani Kumar, M.K.; Halami, P.M.; Serva Peddha, M. Effect of Lactobacillus fermentum MCC2760-Based Probiotic Curd on Hypercholesterolemic C57BL6 Mice. ACS Omega 2021, 6, 7701–7710. [Google Scholar] [CrossRef]
- Kim, H.; Hwang, J.S.; Woo, C.H.; Kim, E.Y.; Kim, T.H.; Cho, K.J.; Kim, J.H. TNF-α-Induced Up-regulation of Intercellular Adhesion Molecule-1 Is Regulated by a Rac-ROS-Dependent Cascade in Human Airway Epithelial Cells. Exp. Mol. Med. 2008, 40, 167–175. [Google Scholar] [CrossRef]
- Claes, I.J.J.; Lebeer, S.; Shen, C.; Verhoeven, T.L.A.; Dilissen, E.; De Hertogh, G.; Bullens, D.M.; Ceuppens, J.L.; Van Assche, G.; Vermeire, S.; et al. Impact of Lipoteichoic Acid Modification on the Performance of the Probiotic Lactobacillus rhamnosus GG in Experimental Colitis. Clin. Exp. Immunol. 2010, 162, 306–314. [Google Scholar] [CrossRef]
- Grangette, C.; Nutten, S.; Palumbo, E.; Morath, S.; Hermann, C.; Dewulf, J.; Pot, B.; Hartung, T.; Hols, P.; Mercenier, A. Enhanced Antiinflammatory Capacity of a Lactobacillus plantarum Mutant Synthesizing Modified Teichoic Acids. Proc. Natl. Acad. Sci. USA 2005, 102, 10321–10326. [Google Scholar] [CrossRef]
- Granato, D.; Bergonzelli, G.E.; Pridmore, R.D.; Marvin, L.; Rouvet, M.; Corthésy-Theulaz, I.E. Cell Surface-Associated Elongation Factor Tu Mediates the Attachment of Lactobacillus johnsonii NCC533 (La1) to Human Intestinal Cells and Mucins. Infect. Immun. 2004, 72, 2160–2169. [Google Scholar] [CrossRef]
- Bergonzelli, G.E.; Granato, D.; Pridmore, R.D.; Marvin-Guy, L.F.; Donnicola, D.; Corthésy-Theulaz, I.E. GroEL of Lactobacillus johnsonii La1 (NCC 533) Is Cell Surface Associated: Potential Role in Interactions with the Host and the Gastric Pathogen Helicobacter pylori. Infect. Immun. 2006, 74, 425–434. [Google Scholar] [CrossRef]
- Qi, S.R.; Cui, Y.J.; Liu, J.X.; Luo, X.; Wang, H.F. Lactobacillus rhamnosus GG Components, SLP, gDNA and CpG, Exert Protective Effects on Mouse Macrophages upon Lipopolysaccharide Challenge. Lett. Appl. Microbiol. 2020, 70, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Hee Kim, C.; Geun Kim, H.; Yun Kim, J.; Ra Kim, N.; Jun Jung, B.; Hye Jeong, J.; Kyun Chung, D. Probiotic Genomic DNA Reduces the Production of Pro-inflammatory Cytokine Tumor Necrosis Factor-alpha. FEMS Microbiol. Lett. 2012, 328, 13–19. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram illustrating the major historical milestones in the development of prebiotics, probiotics, synbiotics, and postbiotics.
Figure 1.
Schematic diagram illustrating the major historical milestones in the development of prebiotics, probiotics, synbiotics, and postbiotics.

Figure 2.
Scheme illustrating the prominent technical advantages, safety profile, and health benefits of utilizing postbiotics.
Figure 2.
Scheme illustrating the prominent technical advantages, safety profile, and health benefits of utilizing postbiotics.

Figure 3.
Isolation of postbiotics from probiotics and characterization methods.

Figure 4.
A scheme illustrating the possible mechanism and application of antidiabetic, antiviral, anti-inflammatory, antibacterial, and anticancer effects of postbiotics. (A) Postbiotics increase antidiabetic activity by increasing insulin production, WAT Browning, glucose homeostasis, and insulin resistance. (B) Postbiotics inhibit viral activity by blocking the receptor-binding site of a virus in the host system. (C) Anti-inflammatory activity by suppressing the production of proinflammatory cytokines. (D) Antibacterial activity by oxidizing biomolecules and forming pores in the membrane. (E) Anticancer activity by caspase activation, cell cycle arrest, and apoptosis.
Figure 4.
A scheme illustrating the possible mechanism and application of antidiabetic, antiviral, anti-inflammatory, antibacterial, and anticancer effects of postbiotics. (A) Postbiotics increase antidiabetic activity by increasing insulin production, WAT Browning, glucose homeostasis, and insulin resistance. (B) Postbiotics inhibit viral activity by blocking the receptor-binding site of a virus in the host system. (C) Anti-inflammatory activity by suppressing the production of proinflammatory cytokines. (D) Antibacterial activity by oxidizing biomolecules and forming pores in the membrane. (E) Anticancer activity by caspase activation, cell cycle arrest, and apoptosis.


{kind=link}
{kind=link}
{kind=link}
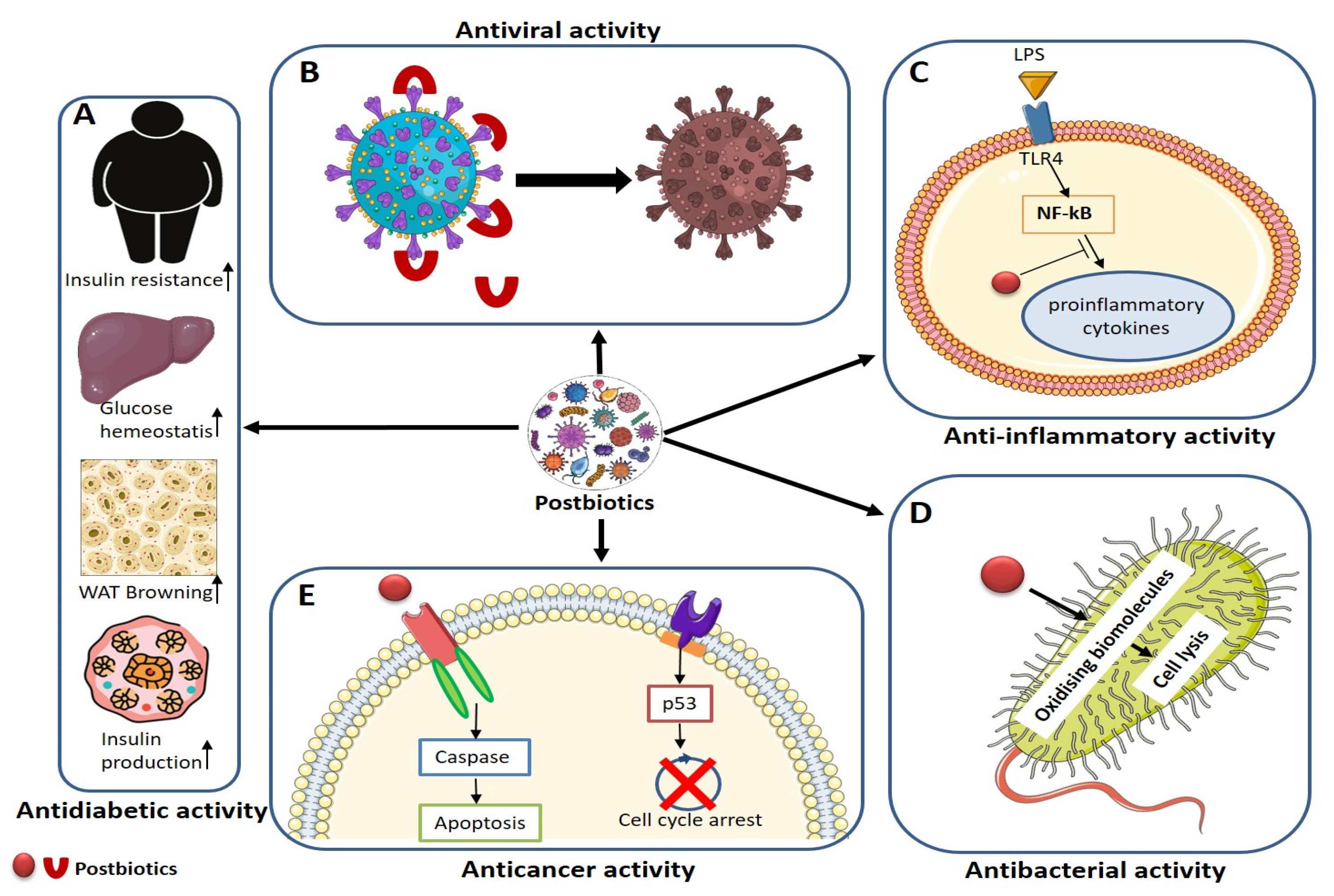{kind=link}
Table 1.
Source, components, types, and functional aspects of postbiotics.
| Bacteria | Derived Postbiotics | Functional Effects | References |
|---|---|---|---|
| Bifidobacteriumlongum | Bacterial lysates | Reduce cholesterol | Shin et al. [170] |
| Anti-inflammation and antibacterial activity | Martorell et al. [171] | ||
| Lipoteichoic acid | Anti-obesity | Balaguer et al. [172] | |
| Exopolysaccharides | Antioxidant | Inturri et al. [173] | |
| Immunomodulation | Inturri et al. [174] | ||
| Anti-inflammation | Schiavi et al. [175] | ||
| Lactobacillus sp. | Anti-inflammation | Sungur et al. [176] | |
| Lactobacillus paracasei | Reduce cholesterol | Bhat and Bajaj [177] | |
| Lacticaseibacillus rhamnosus | Peptidoglycan | Immunomodulation | Kolling et al. [178] |
| Lactobacillus plantarum | Lipoteichoic acid | Immunomodulation | Kim et al. [179] |
| Lactobacillus acidophilu, Lactobacillus reuteri, Lactobacillus plantarum | Matsuguchi et al. [180] | ||
| Lactobacillus paracasei | Anti-inflammation | Wang et al. [181] | |
| Lactobacillus paracasei | Bacterial lysates | Anti-obesity and reduce cholesterol | Osman et al. [182] |
| Lacticaseibacillus rhamnosus | Preventing alcoholic liver disease | Wang et al. [183] | |
| Lactobacillus casei | Immunomodulation and anti-inflammation | Compare et al. [184] | |
| Lacticaseibacillus rhamnosus | Antibacterial activity | Gao et al. [185] | |
| Lactobacillus amylovorus | Anti-obesity and reduce cholesterol | Nakamura et al. [186] | |
| Lactobacillus plantarum | Extracellular vesicles | Anti-inflammation | Haoet al., 2021 [187] |
| Lacticaseibacillus rhamnosus, Lactobacillus acidophilu | Cell-free supernatants | Anti-inflammation | Maghsood et al., 2018 [188] |
| Lactobacillus sp., Bifidobacteriumsp. | Cytoflora (Brand) | Immunomodulation | Barros et al., 2020 [189] |
| Lactobacillus sps., Nyaditumresea, Mycobacterium sp. | Lacteol (Brand) | ||
| Bacillus velezensis | Bacterial lysates | Immunomodulation and antibacterial activity | Mi et al. [190] |
| Bacillus subtilis | Polysaccharides | Anti-diabetic activity | Ghoneim et al. [191] |
| Bacillus licheniformis | Dahech et al. [192] | ||
| Bacillus licheniformis | Extra cellular vesicles | Anticancer activity | Gurunathan et al., 2023 [193] |
| Bacillus coagulans | Metabolites | Antioxidant | Jensen et al., 2017 [194] |
| Saccharomyces cerevisiae | Health advantages | Chan and Liu, 2022 [195] | |
| Saccharomyces cerevisiae | Anti-inflammation | Jensen et al., 2007 [196] | |
| Faecalibacterium prausnitzii | Bacterial lysates | Anti-inflammation | Sokol et al., 2008 [33] |
| Clostridium butyricum | Bacterial lysates | Anticancer activity | Chen et al., 2020 [197] |
| Bifidobacterium breve, Streptococcus thermophilus | Metabolites | Anti-inflammation | Menard et al., 2005 [198] |
| Peanibacillus mucilaginosus | Exopolysaccharides | Antioxidant | Liang et al., 2016 [199] |
| Butyricicoccus pullicaecorum | Butyric acid | Anti-inflammation | Geirnaert et al., 2017 [200] |
| Enterococcus faecium | Exopolysaccharides | Reduce cholesterol | Bhat and Bajaj, 2018 [201] |
| Akkermansia muciniphila | Inactivated bacteria | Anti-obesity | Depommier et al., 2020 [202] |
| Methylococcus capsulatus | Bacterial lysates | Anti-diabetic and immunomodulation | Jensenet al., 2021 [203] |
| Bacteroides thetaiotaomicron | Outer membrane vesicles | Anti-inflammation | Durant et al., 2020 [204] |
| TMAO (Brand) | Metabolites | Anticancer activity | Wang et al., 2022 [205] |
| Pseudomonas aeruginosa | Outer membrane vesicles | Antibacterial and anti-biofilm activity | Gurunathan et al., 2023 [167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gurunathan, S.; Thangaraj, P.; Kim, J.-H. Postbiotics: Functional Food Materials and Therapeutic Agents for Cancer, Diabetes, and Inflammatory Diseases. Foods 2024, 13, 89. https://doi.org/10.3390/foods13010089
AMA Style
Gurunathan S, Thangaraj P, Kim J-H. Postbiotics: Functional Food Materials and Therapeutic Agents for Cancer, Diabetes, and Inflammatory Diseases. Foods. 2024; 13(1):89. https://doi.org/10.3390/foods13010089
Chicago/Turabian StyleGurunathan, Sangiliyandi, Pratheep Thangaraj, and Jin-Hoi Kim. 2024. "Postbiotics: Functional Food Materials and Therapeutic Agents for Cancer, Diabetes, and Inflammatory Diseases" Foods 13, no. 1: 89. https://doi.org/10.3390/foods13010089
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.