
Effects of Viable and Heat-Inactivated Bifidobacterium longum D42 on Proliferation and Apoptosis of HT-29 Human Colon Cancer Cells
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Culture, Cell Culture and Materials
2.2. Proliferation Analysis of HT-29 Cells
2.3. Measurement of Lactate Dehydrogenase Release in HT-29 Cells
2.4. Morphological Analysis of Apoptosis in HT-29 Cells
2.5. Apoptosis Analysis of HT-29 Cells
2.6. Determination of Reactive Oxygen Species Level in HT-29 Cells
2.7. Determination of Changes in Mitochondrial Membrane Potential in HT-29 Cells
2.8. Analysis of Expression Level of Apoptosis-Related Genes in HT-29 Cells
2.9. Analysis of Expression Level of Apoptosis-Related Proteins in HT-29 Cells
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of Viable and Heat-Inactivated B. longum D42 on the HT-29 Cells
3.2. Effect of Viable and Heat-Inactivated B. longum D42 on the Membrane Permeability
3.3. Effect of Viable and Heat-Inactivated B. longum D42 on Apoptosis of HT-29 Cells
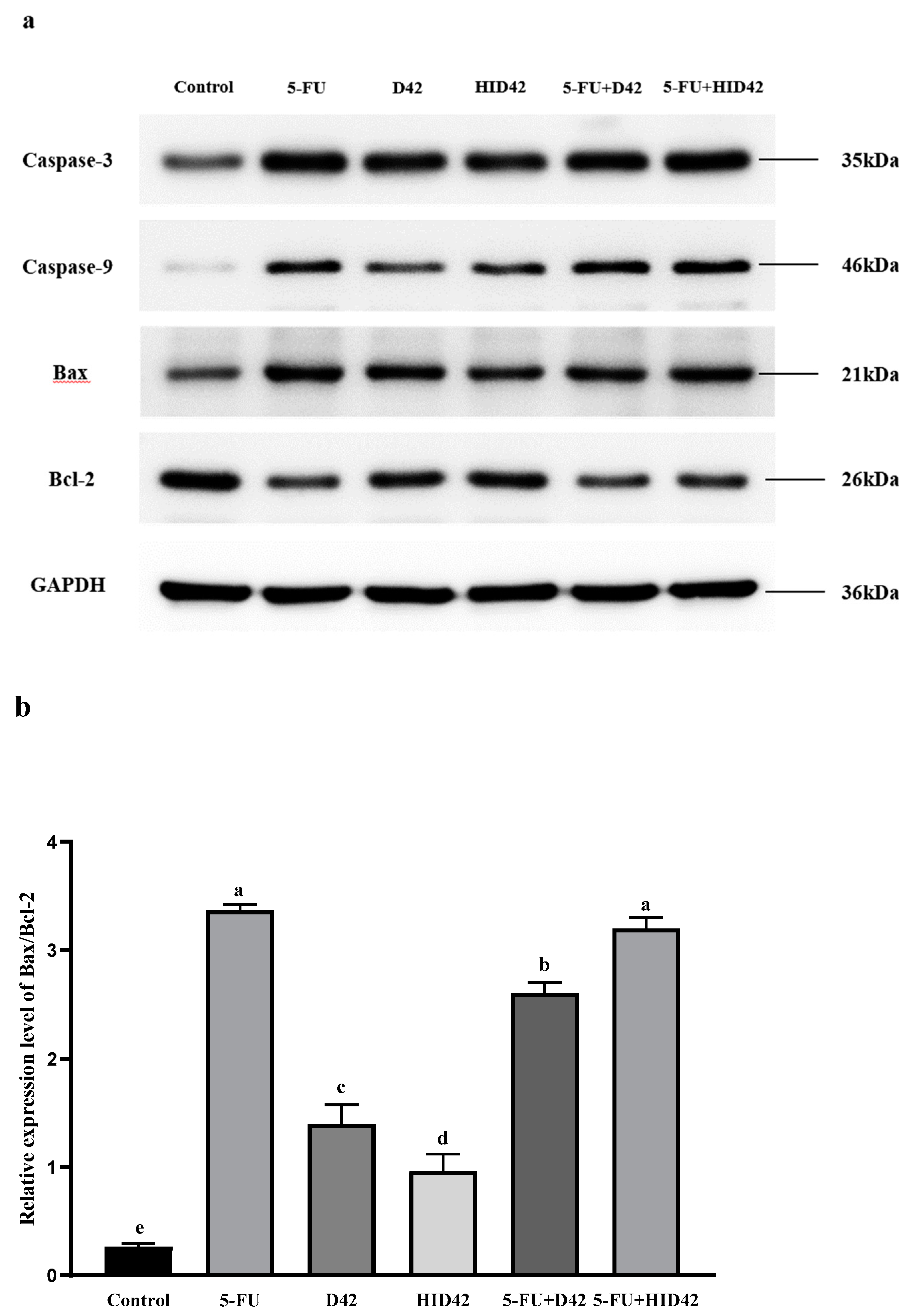
3.4. Further Study on the Apoptotic Mechanism of Viable and Heat-Inactivated B. longum D42 towards HT-29 Cells
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alrushaid, N.; Khan, F.A.; Al-Suhaimi, E.; Elaissari, A. Progress and Perspectives in Colon Cancer Pathology, Diagnosis, and Treatments. Diseases 2023, 11, 148. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, Y.K.; Lee, D.J.; Kim, K.M. Colonoscopy procedural skills and training for new beginners. World J. Gastroenterol. 2014, 20, 16984–16995. [Google Scholar] [CrossRef]
- Deng, X.; Yang, J.; Zhang, Y.; Chen, X.Y.; Wang, C.; Suo, H.Y.; Song, J.J. An Update on the Pivotal Roles of Probiotics, Their Components, and Metabolites in Preventing Colon Cancer. Foods 2023, 12, 3706. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Zhu, R.Q.; Lang, T.Q.; Yan, W.L.; Zhu, X.; Huang, X.; Yin, Q.; Li, Y.P. Gut Microbiota: Influence on Carcinogenesis and Modulation Strategies by Drug Delivery Systems to Improve Cancer Therapy. Adv. Sci. 2021, 8, 2003542. [Google Scholar] [CrossRef]
- Dienstmann, R.; Salazar, R.; Tabernero, J. Personalizing Colon Cancer Adjuvant Therapy: Selecting Optimal Treatments for Individual Patients. J. Clin. Oncol. 2015, 33, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Sieow, B.F.-L.; Wun, K.S.; Yong, W.P.; Hwang, I.Y.; Chang, M.W. Tweak to Treat: Reprograming Bacteria for Cancer Treatment. Trends Cancer 2021, 7, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Belguesmia, Y.; Bendali, F.; Spano, G.; Seal, B.S.; Drider, D. Lactobacillus fermentum: A bacterial species with potential for food preservation and biomedical applications. Crit. Rev. Food Sci. Nutr. 2020, 60, 3387–3399. [Google Scholar] [CrossRef]
- Dasari, S.; Kathera, C.; Janardhan, A.; Kumar, A.P.; Viswanath, B. Surfacing role of probiotics in cancer prophylaxis and therapy: A systematic review. Clin. Nutr. 2017, 36, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Akedo, I.; Otani, T.; Suzuki, T.; Nakamura, T.; Takeyama, I.; Ishiguro, S.; Miyaoka, E.; Sobue, T.; Kakizoe, T. Randomized trial of dietary fiber and Lactobacillus casei administration for prevention of colorectal tumors. Int. J. Cancer 2005, 116, 762–767. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef]
- Hashempour-Baltork, F.; Hosseini, H.; Shojaee-Aliabadi, S.; Torbati, M.; Alizadeh, A.M.; Alizadeh, M. Drug Resistance and the Prevention Strategies in Food Borne Bacteria: An Update Review. Adv. Pharm. Bull. 2019, 9, 335–347. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Abbasi, A. Molecular mechanisms of postbiotics in colorectal cancer prevention and treatment. Crit. Rev. Food Sci. Nutr. 2021, 61, 1787–1803. [Google Scholar] [CrossRef]
- Ardestani, S.K.; Tafvizi, F.; Ebrahimi, M.T. Heat-killed probiotic bacteria induce apoptosis of HT-29 human colon adenocarcinoma cell line via the regulation of Bax/Bcl2 and caspases pathway. Hum. Exp. Toxicol. 2019, 38, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Shamekhi, S.; Abdolalizadeh, J.; Ostadrahimi, A.; Mohammadi, S.A.; Barzegari, A.; Lotfi, H.; Bonabi, E.; Zarghami, N. Apoptotic Effect of Saccharomyces cerevisiae on Human Colon Cancer SW480 Cells by Regulation of Akt/NF-κB Signaling Pathway. Probiotics Antimicrob. Proteins 2020, 12, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.C.; OuYang, C.N.; Yuan, S.N.; Lin, H.C.; Huang, K.Y.; Wu, P.S.; Liu, C.Y.; Tsai, K.J.; Loi, L.K.; Chen, Y.J.; et al. Pretreatment with a Heat-Killed Probiotic Modulates the NLRP3 Inflammasome and Attenuates Colitis-Associated Colorectal Cancer in Mice. Nutrients 2019, 11, 516. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, X.; Ho, C.L. Recent Development of Probiotic Bifidobacteria for Treating Human Diseases. Front. Bioeng. Biotechnol. 2021, 9, 770248. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, C.A.; Gamal-Eldeen, A.M.; El-Hussieny, E.A.; Raafat, B.M.; Mehanna, N.S.; Talaat, R.M.; Shaaban, M.T. Bifidobacterium longum Suppresses Murine Colorectal Cancer through the Modulation of oncomiRs and Tumor Suppressor miRNAs. Nutr. Cancer-Int. J. 2019, 71, 688–700. [Google Scholar] [CrossRef]
- Singh, J.; Rivenson, A.; Tomita, M.; Shimamura, S.; Ishibashi, N.; Reddy, B.S. Bifidobacterium longum, a lactic acid-producing intestinal bacterium inhibits colon cancer and modulates the intermediate biomarkers of colon carcinogenesis. Carcinogenesis 1997, 18, 833–841. [Google Scholar] [CrossRef]
- Yue, Y.X.; Wang, S.; Shi, J.L.; Xie, Q.G.; Li, N.; Guan, J.Q.; Evivie, S.E.; Liu, F.; Li, B.L.; Huo, G.C. Effects of Lactobacillus acidophilus KLDS1.0901 on Proliferation and Apoptosis of Colon Cancer Cells. Front. Microbiol. 2022, 12, 788040. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, S.M.; Zhang, L.W.; Fan, R.B.; Han, X.; Yi, H.X.; Zhang, L.L.; Xue, C.H.; Li, H.B.; Zhang, Y.H.; Shigwedha, N. Induction of HT-29 cells apoptosis by lactobacilli isolated from fermented products. Res. Microbiol. 2014, 165, 202–214. [Google Scholar] [CrossRef]
- Alizadeh, S.; Esmaeili, A.; Omidi, Y. Anti-cancer properties of Escherichia coil Nissle 1917 against HT-29 colon cancer cells through regulation of Bax/Bcl-xL and AKT/PTEN signaling pathways. Iran. J. Basic Med. Sci. 2020, 23, 886–893. [Google Scholar] [CrossRef]
- Ramadoss, D.P.; Sivalingam, N. Vanillin extracted from proso and barnyard millets induces cell cycle inhibition and apoptotic cell death in MCF-7 cell line. J. Cancer Res. Ther. 2021, 17, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Dong, T. Effects of propofol on invasion and migration of colon cancer cells and JAK2/STAT3 signaling pathway. Zhong Nan Da Xue Xue Bao Yi Xue Ban J. Cent. South Univ. Med. Sci. 2020, 45, 290–296. [Google Scholar] [CrossRef]
- Fesik, S.W. Promoting apoptosis as a strategy for cancer drug discovery. Nat. Rev. Cancer 2005, 5, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, L.L.; Hong, D.; Liu, Y.L.; Han, P.P.; Li, S.T.; Jia, Y.M. Broad-spectrum cytotoxicity to cancer cells of Brevilaterin C from Brevibacillus laterosporus and its specific mechanism on human epidermal cancer cells. J. Cell. Biochem. 2022, 123, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Rogoff, H.A. Implications of reactive oxygen species on cancer formation and its treatment. Semin. Oncol. 2021, 48, 238–245. [Google Scholar] [CrossRef]
- Lin, S.S.; Li, Y.Y.; Zamyatnin, A.A.; Werner, J.; Bazhin, A.V. Reactive oxygen species and colorectal cancer. J. Cell. Physiol. 2018, 233, 5119–5132. [Google Scholar] [CrossRef]
- Hu, P.P.; Song, W.; Shan, Y.J.; Du, M.; Huang, M.H.; Song, C.; Zhang, L.W. Lactobacillus paracasei subsp paracasei M5L induces cell cycle arrest and calreticulin translocation via the generation of reactive oxygen species in HT-29 cell apoptosis. Food Funct. 2015, 6, 2257–2265. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial dysfunction and oxidative stress in heart disease. Exp. Mol. Med. 2019, 51, 162. [Google Scholar] [CrossRef]
- Guo, Y.X.; Zhang, T.; Gao, J.J.; Jiang, X.X.; Tao, M.X.; Zeng, X.Q.; Wu, Z.; Pan, D.D. Lactobacillus acidophilus CICC 6074 inhibits growth and induces apoptosis in colorectal cancer cells in vitro and in HT-29 cells induced-mouse model. J. Funct. Foods 2020, 75, 104290. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.L.; Wang, X.T.; Yang, X.Z.; Li, X.P.; Yuan, M.L. Selection of reference genes for qRT-PCR and expression analysis of high-altitude-related genes in grassland caterpillars (Lepidoptera: Erebidae: Gynaephora) along an altitude gradient. Ecol. Evol. 2017, 7, 9054–9065. [Google Scholar] [CrossRef]
- Cheresh, P.; Kim, S.J.; Tulasiram, S.; Kamp, D.W. Oxidative stress and pulmonary fibrosis. Biochim. Biophys. Acta-Mol. Basis Dis. 2013, 1832, 1028–1040. [Google Scholar] [CrossRef]
- Zhong, L.; Zhang, X.; Covasa, M. Emerging roles of lactic acid bacteria in protection against colorectal cancer. World J. Gastroenterol. 2014, 20, 7878–7886. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Hsieh, Y.M.; Huang, C.C.; Tsai, C.C. Inhibitory Effects of Probiotic Lactobacillus on the Growth of Human Colonic Carcinoma Cell Line HT-29. Molecules 2017, 22, 107. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.T.; Chu, F.J.; Chou, C.C.; Yu, R.C. Antiproliferative and anticytotoxic effects of cell fractions and exopolysaccharides from Lactobacillus casei 01. Mutat. Res.-Genet. Toxicol. Environ. Mutagen. 2011, 721, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Nozari, S.; Faridvand, Y.; Etesami, A.; Beiki, M.A.K.; Mazrakhondi, S.A.M.; Abdolalizadeh, J. Potential anticancer effects of cell wall protein fractions from Lactobacillus paracasei on human intestinal Caco-2 cell line. Lett. Appl. Microbiol. 2019, 69, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.C.; Lin, S.L.; Tsai, J.J.; Lin, M.Y. Heat-Killed Lactic Acid Bacteria Enhance Immunomodulatory Potential by Skewing the Immune Response toward Th1 Polarization. J. Food Sci. 2011, 76, M260–M267. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| Caspase-3 | TGCCTGTAACTTGAGAGTAGATGG | CTTCACTTTCTTACTTGGCGATGG |
| Caspase-9 | TGCTGCGTGGTGGTCATTCTC | CCGACACAGGGCATCCATCTG |
| Bax | CCCGAGAGGTCTTTTTCCGAG | CCAGCCCATGATGGTTCTGAT |
| Bcl-2 | GGTGGGGTCATGTGTGTGG | CGGTTCAGGTACTCAGTCATCC |
| GAPDH | AAGCTCATTTCCTGGTATGACAACG | TCTTCCTCTTGTGCTCTTGCTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Cao, T.; Wang, Y.; Yang, R.; Han, Y.; Li, S.; Liu, D.; Yue, Y.; Cao, Y.; Li, B.; et al. Effects of Viable and Heat-Inactivated Bifidobacterium longum D42 on Proliferation and Apoptosis of HT-29 Human Colon Cancer Cells. Foods 2024, 13, 958. https://doi.org/10.3390/foods13060958
Zhang Y, Cao T, Wang Y, Yang R, Han Y, Li S, Liu D, Yue Y, Cao Y, Li B, et al. Effects of Viable and Heat-Inactivated Bifidobacterium longum D42 on Proliferation and Apoptosis of HT-29 Human Colon Cancer Cells. Foods. 2024; 13(6):958. https://doi.org/10.3390/foods13060958
Chicago/Turabian StyleZhang, Yifan, Ting Cao, Yuqi Wang, Rui Yang, Yu Han, Shuang Li, Deyu Liu, Yingxue Yue, Yun Cao, Bailiang Li, and et al. 2024. "Effects of Viable and Heat-Inactivated Bifidobacterium longum D42 on Proliferation and Apoptosis of HT-29 Human Colon Cancer Cells" Foods 13, no. 6: 958. https://doi.org/10.3390/foods13060958