4.1. Effects of Different Aquaculture Modes on the Physical Properties of Grass Carp Muscle
Muscles’ capacity to retain moisture under external conditions is known as water-holding capability (WHC), which is influenced by their unique chemical makeup and physical characteristics. It is a physicochemical index for evaluating muscle quality, significantly affecting muscle texture [
19]. A higher WHC indicates a slower loss of nutrients and flavor substances, longer storage times, and better muscle quality. WHC can be measured by indicators such as drip loss, centrifugal loss, liquid loss, stored loss, frozen leakage, and cooking loss, all of which are inversely related to WHC [
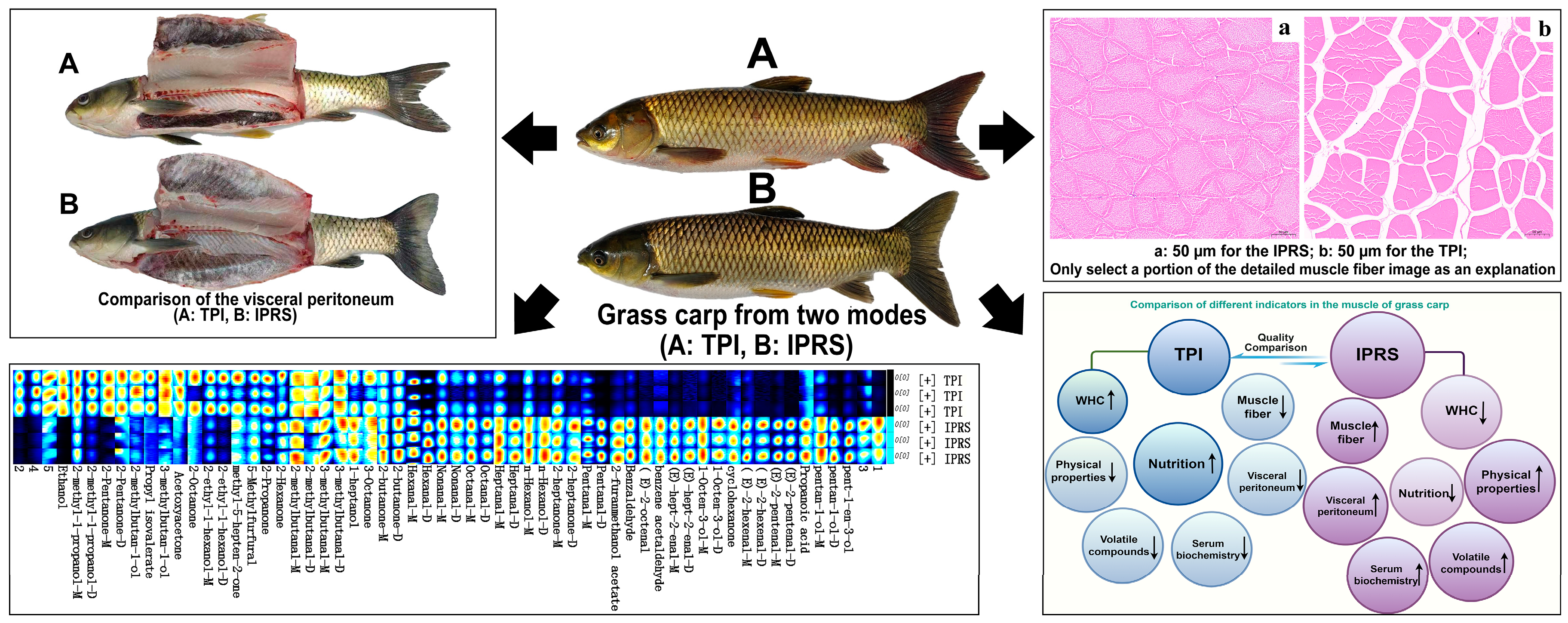
11]. The TPI group had notably reduced liquid loss compared to the IPRS group in the experimental findings (
p < 0.05), with an even more significant decrease in cooking loss (
p < 0.01) (
Table 2). This indicates that the TPI group has a higher WHC, related to the connective tissue between muscle fibers. Studies have shown that an increase in connective tissue in muscles can enhance the ability to retain moisture and prevent juice loss [
20]; when muscle is compressed or cooked and contracts, a certain amount of connective tissue reduces water mobility, retaining water within the muscle tissue [
21]. The significantly lower WHC in the IPRS group is linked to its low connective tissue content, differing from the general assumption that high muscle fiber density correlates with high WHC, proving that WHC is also constrained by muscle connective tissue, forming a prerequisite for water retention that includes both muscle fiber density and connective tissue elements [
22].
Muscle fiber properties are important physical indicators for evaluating meat quality, closely related to muscle meat quality. Muscle fiber properties are mainly influenced by diameter and density; smaller fiber diameters at constant volume lead to higher density, resulting in firmer muscle with better texture. This research revealed that the size of muscle fibers in the IPRS group (
Table 3), both long and short diameters, were notably smaller compared to the TPI group (
p < 0.01), while also showing a higher fiber density (
p < 0.01). Research suggests that exercise training promotes fish growth by regulating muscle fiber hyperplasia, possibly due to enhanced movement in pond raceway systems promoting fiber proliferation and reducing inter-fiber cell spacing, leading to shorter fiber diameters and increased overall density [
23,
24].
Muscle fiber structure and texture properties are closely related, affecting the textural properties of fish meat [
22]. The characteristics of fish flesh texture, which are crucial for evaluating the quality of food, encompass attributes like hardness, chewiness, cohesiveness, springiness, resilience, and gumminess, with hardness playing a key role in determining the quality of the meat [
24,
25,
26]. The IPRS group in the grass carp raw flesh category exhibited notably elevated levels of hardness, chewiness, cohesiveness, springiness, resilience, gumminess, and shearing in comparison to the TPI group (
p < 0.05) (
Figure 2), consistent with the results reported by Zhang but with slightly different aquaculture modes [
27]. The cooked flesh group showed similar results, with the IPRS group showing significantly higher values in the same parameters (
p < 0.05); the texture property indicators of both groups of cooked meat were significantly lower than in the raw meat group (
p < 0.05). This study demonstrates that smaller muscle fiber diameters and higher muscle density correlate with increased hardness and chewiness, thereby enhancing meat quality [
26,
28]. This suggests a positive correlation between muscle fiber density and indicators such as hardness and chewiness, similar to findings from Jia’s study [
11]. Conversely, the lower fiber density and higher connective tissue content in the TPI group reduce texture properties such as hardness and springiness while inhibiting moisture loss, resulting in a softer texture and higher moisture content in TPI-group meat [
21,
25]. According to Luo’s result, reduced muscle hardness impacts meat quality, while improvements in hardness, chewiness, and springiness enhance meat quality, indicating that IPRS plays a positive role in improving muscle texture properties and meat quality [
29].
4.2. Impact of Various Aquaculture Modes on the Nutritional Composition of Grass Carp Flesh
Muscle typically contains moisture, ash, crude protein, and crude lipid as its primary nutritional components, with protein and lipid being the most concentrated. The muscle quality is greatly influenced by the protein and lipid content. Excessive lipid in cultivated fish can negatively impact taste [
30]. Our experimental findings indicate that there were no notable variances in moisture and crude protein levels in grass carp muscles between the two cultivation methods (
Table 4). However, the ash content in the IPRS group was notably lower than in the TPI group (
p < 0.01), while the crude lipid content was significantly higher in the IPRS group compared to the TPI group (
p < 0.05). These findings are consistent with Gharti’s result, who reported increased crude lipid levels in grass carp raised in raceways, which is also in line with the results of Hai-Shan, where the crude lipid content in gibel carp (
Carassius auratus gibelio) muscle significantly increased after swimming training [
16,
31]. The fundamental reason for the differences in nutritional content between aquaculture modes lies in the variation in food and environment [
32]. Multiple research studies indicate that swimming activity at specific levels can boost the crude lipid content in fish muscles, as raceway cultivation promotes moderate exercise, enhancing muscle growth and indirectly leading to fat deposition. However, when the exercise exceeds a certain threshold, it leads to direct fat breakdown for energy, decreasing fat content and leading to reduced body weight and condition factor [
33], consistent with the findings of Li, where fat levels initially rise, then decrease, and finally stabilize [
34]. During TPI mode, grass carp consume grass and other natural food, leading to reduced muscle fat levels and increased moisture levels. The decreased fat levels are linked to the heightened activity and vigilance of grass carp in pond intercropping, as stated in Gharti’s study [
16]. The higher fat content in the IPRS group leads to smoother muscle; if the fat content is lower, the muscle becomes coarser, reducing palatability [
30]. The higher ash content in the TPI group is closely related to its living environment, where still-water ponds contain rich sediment and aquatic plants, providing abundant inorganic salts and minerals.
Fatty acids play a crucial role in evaluating the nutritional value of fish muscle, necessary for overall health, with their nutritional content mainly determined by the levels of Σn-3, Σn-6, EPA, and DHA. Aquaculture in raceway systems primarily enhances the quality of muscle nutrition by regulating muscle fiber texture and nutritional components through exercise [
35,
36,
37,
38]. The findings from the experiment indicated that the level of saturated fatty acids in the IPRS group was notably greater compared to the TPI group (
p < 0.01) (
Table 5). Additionally, the levels of PUFAs, EPA, DHA, and Σn-3 were significantly lower in the IPRS group (
p < 0.01), which contradicted the prevailing belief in various studies like Zhu’s that moderate swimming can greatly boost the levels of PUFAs, EPA, and DHA in fish muscle [
39]. This is also in contrast to the results of Yuan’s study, which found that the IPRS method resulted in higher levels of MUFAs, EPA, and DHA [
14]. Studies have indicated that the levels of PUFAs, EPA, and DHA rise during low-intensity swimming but decline during high-intensity swimming [
23,
40]. This explains why the IPRS group, despite exercising, had lower levels of PUFAs, EPA, and DHA, likely due to excessive exercise intensity [
41]. The reason why the TPI group had higher contents of PUFAs, Σn-3, EPA, and DHA is that TPI grass carp feed on grass and plankton, with plankton being a major source of Σn-3 polyunsaturated fatty acids like EPA and DHA [
42]. The significantly higher ΣSFA content in the IPRS group is similar to findings by Wang and Jia [
11,
43]. SFAs, as high-energy fatty acids, can supply the necessary energy for swimming in the IPRS group, and being composed of lipid substances, SFAs are closely related to the significantly higher crude fat content in the IPRS group [
44]. The IPRS group requires more energy for movement, leading to a relative abundance of energy-related saturated fatty acids; the TPI group, being a static cultivation mode that feeds on natural food like grass, consumes less energy yet achieves a superior muscle nutritional composition compared to IPRS-fed grass carp, aligning with the result of Zhao’s study [
45]. This indirectly suggests that fish meat quality post-exercise is not necessarily superior to that of fish without continuous exercise, providing new insights for TPI mode.
Amino acids serve as a crucial metric for assessing the nutritional value of muscle, evaluated through seventeen different types of amino acids, which include nine essential amino acids (EAAs), four non-essential amino acids (NEAAs), and four umami amino acids (UAAs). The presence of EAAs and UAAs is essential for both the nutritional quality and taste of muscle tissue [
23]. Our findings showed that the IPRS group exhibited notably elevated amounts of valine and histidine compared to the TPI group (
p < 0.05), whereas the content of lysine and the total umami amino acids were significantly higher in the TPI group (
p < 0.05), with no notable differences in ΣEAAs and ΣNEAAs (
Table 6). Studies suggest that valine is crucial for muscle production and maintenance, improving the physical flavor characteristics of fish slices and preventing muscle atrophy [
46]; histidine is a key element in muscle composition [
47]. Research has shown that exercise in fish can enhance muscle characteristics, possibly due to an increase in amino acids that maintain muscle, ensuring quality. Lysine is crucial for the growth of TPI-group grass carp, serving as the primary amino acid that limits their development [
48]. Certain research suggests that moderate exercise may boost the levels of EAAs and UAAs in muscle, ultimately improving its taste [
49]. This is in opposition to the lower ΣUAA levels in the IPRS group found in this experiment. This discrepancy may result from the exercise intensity in IPRS mode not favoring the synthesis of umami amino acids, or the intercropping aquaculture environment may be better for creating umami amino acids in grass carp muscle. This could be attributed to the grass carp’s natural diet and other natural foods, which may enhance the levels of UAAs in the muscles, as seen in the study by Zhao, who found that grass carp feeding on natural food like grass had higher fatty acids and protein [
45]. Glutamate, the primary source of meaty flavor [
50], showed no significant difference between the two aquaculture modes (
p > 0.05), suggesting that pond intercropping leads to an overall increase in UAAs without significantly altering meat flavor and texture. In summary, the TPI-group grass carp had higher ΣUAA content, while the IPRS group had more amino acids that help to generate and maintain muscle tissue, with both aquaculture modes positively impacting muscle quality.
4.3. Impact of Different Aquaculture Modes on Volatile Compounds in Grass Carp Muscle
In the IPRS group, the total amount of volatile compounds was higher compared to the TPI group, with 32 types showing significantly higher levels. Conversely, the TPI group exhibited only 12 significantly higher types. The IPRS group exhibited notably elevated levels of nonanal, hexanal, heptanal, hexanol, 1-octen-3-ol, 2-heptanone, 2-butanone, 2-methylbutanal, and 3-methylbutanal compared to the TPI group (
p < 0.05); conversely, the TPI group had significantly higher ethanol and 2-ethylhexanol contents than the IPRS group (
p < 0.01) (
Table S1). Aldehydes are considered key to fish meat flavor, with nonanal, hexanal, and heptanal identified as markers of fish meat flavor, with nonanal being a primary flavor compound [
51,
52]. Nonanal has a distinct aroma of fresh green grass and melon [
20], while hexanol has a grassy flavor [
53]; 1-octen-3-ol is described as having a mushroom-like flavor and a potent vegetal scent [
54,
55]. 2-Heptanone is considered to have fruity, vanilla, and floral flavors [
56]; 2-butanone is associated with a buttery flavor and freshness identification [
57,
58]. 2-Methylbutanal and 3-methylbutanal are considered to have mushroom and vegetal grass flavors, respectively [
59]. Overall, the IPRS group exhibited a variety of flavor aldehydes with intense multiple aromas. Aldehydes are thought to be degradation products of unsaturated fatty acids under the action of enzymes and microbes, closely related to lipid content. Exercise may increase fat content, and in conjunction with the combined action of exercise and lipids, leads to the production of some aromatic aldehydes and other volatile substances to enhance the aroma of fish meat [
60]. The TPI group has a high ethanol content, which was different from the results of Ma’s study [
12], possibly due to the presence of wild algae and floating plants in the intercropping ponds that can serve as raw materials for ethanol synthesis, resulting in a significantly higher ethanol content in grass carp muscle compared to the IPRS group [
61]. The TPI group also exhibited a high content of 2-ethylhexanol, imparting a strong earthy aroma, attributed to the typically sediment-rich environment of TPI ponds, where grass carp inhabit the middle-to-lower layers of water with high soil content, leading to an increase in the volatile compound content imparting an earthy aroma to the muscles. In summary, the volatile aroma of grass carp meat in the IPRS group is primarily influenced and regulated by exercise, with exercise significantly modulating the meat’s aroma, resulting in multidimensional changes in aroma, which is relatively fresh and pleasant. The meat aroma of the TPI group is easily influenced by the environment, characterized by a strong wine aroma but also containing a pronounced earthy aroma, indicating that sediment-rich soil and abundant algae in intercropping ponds have a significant impact on the flavor of the meat.
4.4. Impact of Different Aquaculture Modes on Serum Biochemical Indices of Grass Carp
Serum biochemical indicators are closely related to the physiological status of fish, and their changes are accompanied by changes in substances such as proteins, lipids, and sugars in the body. The serum biochemical indicators typically remain within normal range. Abnormal conditions in a fish’s body, including lesions, injuries, or other irregularities, can lead to abnormalities in serum biochemical indicators. The total protein, albumin, and alkaline phosphatase in serum play important roles in fish metabolism and immune function, with albumin being the most abundant protein in plasma, closely related to fish immunity and the health index [
62,
63]. The findings indicated that the levels of serum total protein and alkaline phosphatase were notably elevated in the IPRS group compared to the TPI group (
p < 0.01), with the albumin content also being significantly higher in the IPRS group than the TPI group (
p < 0.05) (
Table 7), consistent with the findings of Ma’s study [
12]. This is related to the continuous exercise of the IPRS group. According to Zhang’s study, exercise can improve fish health, relying on the proteasome system in the fish body, accelerating protein turnover, regulating the dynamic changes of various enzyme activities in the serum, and enhancing fish metabolism and immune function, specifically reflected in increasing the plasma protein level of the IPRS group [
64].
Triglycerides and total cholesterol can reflect the content of blood lipids and the lipid metabolism ability of fish [
65]. Research indicates that moderate aerobic exercise can lower levels of TG and TCHO [
14]. The results of this experiment are similar to those of Wang’s result [
66]. The findings indicated a significant increase in TG levels in the IPRS group compared to the non-exercise TPI group (
p < 0.01), along with a significant decrease in TCHO levels compared to the TPI group (
p < 0.01) (
Table 7). The continuous exercise of the IPRS group continuously promotes aerobic metabolism in fish, thereby increasing their feed intake and feeding frequency, causing an increase in TG and fat content, and participating in regulating the conversion of TG to SFA in the IPRS group [
33,
44,
67,
68]. TG can store energy in fish, and the increase in its content indicates that endogenous fat transport is active [
32]. The content of TCHO can reflect the liver metabolic ability, indicating that the liver of grass carp in the IPRS group has a strong ability to metabolize cholesterol, which is related to the regulation of protease activity in the fish body by exercise. By improving the immune health of the fish body, the liver metabolic ability can be improved.
Serum levels of ALT and AST are frequently utilized as biomarkers to assess liver conditions. An increase in ALT and AST indicates varying degrees of damage such as liver and pancreatic dysfunction, necrosis, tissue degeneration, and changes in protein metabolism [
65,
69,
70]. ALT and AST levels were notably reduced in the IPRS group compared to the TPI group (
Table 7), aligning with the findings of Kharat and Yuan [
14,
71]. The experimental findings follow a pattern comparable to Zhang’s result, but the AST content of both groups in this experiment is higher than the normal value, with the TPI group significantly exceeding the standard [
72]. Various research has indicated that insufficient oxygen levels in water can harm the liver of grass carp and elevate AST levels. Grass carp ingesting a certain amount of selenium will effectively reduce the content of ALT and AST and alleviate liver damage, and grass carp ingesting enough alanine and glycine will improve the growth performance of grass carp larvae and reduce ALT and AST levels [
72,
73,
74]. The high AST content in the IPRS group may be related to the high-density hypoxic runway aquaculture environment, but due to the fact that consuming selenium-containing artificial feed can alleviate the rise of ALT and AST, the content is lower than that in the TPI group. However, grass carp in the TPI group were not artificially fed, and their food sources lacked sufficient selenium, alanine, and glycine. Furthermore, they usually had high stress levels, which may be important causes of liver damage in the TPI group. Carbohydrates in the blood are an important source of energy and are related to the body’s metabolic levels [
75]. Bony fish are usually glucose-intolerant and exhibit persistent high blood sugar levels, so the blood sugar levels in the fish can indirectly reflect their metabolic levels [
76]. Both groups exhibited decreased GLC levels, with those of the IPRS group being even lower (
Table 7), which was different from the results of Ma’s study and similar to the results of Su’s study [
12,
77]. The fast glucose clearance rate of grass carp plasma leads to lower GLC levels than other fish, which is related to the strong digestion, gluconeogenesis, and glucose metabolism ability of grass carp. According to West’s study, fish in the swimming exercise group exhibited a glucose metabolism rate three times higher than those in the non-swimming exercise group [
78]. The swimming exercise group exhibited increased metabolic activity and improved glucose tolerance. Swimming can lower blood sugar, improve metabolic function, and alleviate the hyperglycemic state caused by glucose formation in the body.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}