3.1. Acidity and Microbiological and Chromaticity Characteristics of Lentil Wholemeal Samples
The pH values, chromaticity characteristics, and LAB viable counts of lentil samples are presented in
Table 1. Comparing the pH values of non-fermented LR and L sample groups (SM and SS), higher pH values were observed in LR sample groups. However, after 24 and 48 h of SMF, the opposite tendencies were established; i.e., L samples showed lower pH values than LR groups. SSF conditions were more effective in reducing the pH values of L samples; however, when comparing SSF L and LR sample groups, lower pH values were attained in L sample groups (after 24 and 48 h of SSF). In both sample groups (LR and L) under both conditions (SMF and SSF), the pH values decreased between 24 and 48 h of fermentation.
Despite the pH differences in non-fermented samples (between C
LRSM and C
LSM; C
LRSS and C
LSS), significant differences in LAB viable counts were found only between C
LRSM and C
LSM samples. Contrasting non-fermented samples (C
LRSM, C
LRSS, C
LSM, and C
LSS), LAB viable counts were, on average, 4.67 log
10 CFU/g. In comparison, for LAB numbers in different lentil sample groups (LR and L) with the same treatment, significant differences were also inexistent, and, in both sample groups (SMF and SSF), higher LAB viable counts were detected after 48 h of fermentation, with an average value of 8.23 log
10 CFU/g. The analysed factors (type of fermentation and fermentation duration) and the interaction of growing conditions * fermentation duration were significant (
p = 0.005,
p < 0.001, and
p = 0.003, respectively) for the LAB viable counts in lentil samples (see
supplementary file: Table S1). Also, a strong negative correlation (r = −0.886,
p < 0.001) between lentil pH values and LAB numbers was found (see
supplementary file: Table S2).
The fermentation of leguminous crops can provide probiotic benefits in foods [
13]. Moreover, it was reported that SSF is more commonly used for various biological conversion of substrates, because of its higher efficiency [
40]. Our study showed that all the analysed factors (type of fermentation, growing conditions, and fermentation duration) and their interactions were significant (
p < 0.001) on the pH values of lentil samples (see
supplementary file: Table S1). The results also showed that, in all of the fermented lentil samples, LAB viable counts were higher than 6.0 log
10 CFU/g; such a number of desirable microorganisms may ensure the probiotic properties of foodstuffs, if the strains used for fermentation are probiotics (which is the case here).
The carbohydrate metabolism of
P. acidilactici strain was studied in 47 different carbon sources, and this strain demonstrated fermentation activity for 20 out of 47 tested carbohydrates [
34]. Additionally, the optimal growth temperature for this strain was 30 °C. This characteristic can explain the differences in the fermentation activity of the
P. acidilactici strain reported in the study of Byanju et al. [
13], who found that, for
P. acidilactici (provided by Lallemand Animal Nutrition–North America, Milwaukee, WI, USA), the exponential growth in lentil flour was observed between 6 and 24 h in SMF conditions. In addition, the pH decreased significantly during the first 24 h of fermentation, and the microbial population attained the highest value during this time period [
13].
Throughout fermentation, the low pH of the substrate can damage both the non-desirable and desirable microorganism’s cell wall and cell membrane [
41]. LAB adaptation to low pH conditions depends on their phenotype characteristics and other conditions under which cells are exposed to stress [
42]. The molecular mechanisms of LAB adaptation and habituation to stress may overlap to a certain degree, but they are not completely identical [
34]. This may explain the different results reported in distinct LAB species for the fermentation effectiveness parameters. However, the pH values and the number of LAB are the main characteristics of fermentation efficiency. This study showed that
P. acidilactici is a suitable strain for lentil fermentation; furthermore, a 48 h fermentation duration was shown to be most suitable for lentil fermentation with this particular LAB strain, because statistically significant lower pH and higher LAB viable counts after 48 h of SMF and SSF were obtained, when compared with 24 h fermented samples.
In comparison, chromaticity characteristics of the non-fermented lentils (LR samples: C
LRSM and C
LRSS) showed, respectively, on average, 1.03 and 10.6% lower brightness (L*) and yellowness (b*), as well as, on average, 42.3% higher redness (a*), in comparison with L samples. In all cases, the same tendencies were found: with an increasing duration of fermentation, the L* and b* values of the lentils decreased and the a* values increased. Lentil L*, a*, and b* values showed statistically significant correlations with pH and LAB viable counts (with pH: r = 0.763,
p < 0.001; r = −0.679,
p < 0.001; and r = 0.839,
p < 0.001, respectively; with LAB viable counts: r = −0.710,
p < 0.001; r = 0.552,
p < 0.001; and r = −0.799,
p < 0.001, respectively). Analysed factors (fermentation conditions, lentil growing conditions, and fermentation duration) and their interactions were statistically significant (
p < 0.001) for the colour coordinate values of lentil samples, except for the interaction between growing conditions, type of fermentation, and fermentation durations on a* values (see
supplementary file: Table S1).
The colour of the lentil is a key quality parameter due to its impact on the acceptability of the legume products. The lentil colour ranges from light tan to dark brown, and the latter is often referred as being a lower quality product. Furthermore, a lighter colour can be linked to a loss of nutrients or other secondary metabolites, such as polyphenolic compounds [
2]. Our study discovered that the changes in lentil colour can be found during the fermentation process. It was reported that the anthocyanins can be hydrolysed by bacterial
β-glucosidase enzymes and converted to small phenolic compounds [
43]. Likewise, other flavonoid pigments (flavonols, flavones isoflavones, etc.) may be degraded by bacterial enzymes [
43]. Excreted bacterial enzymes break glycosidic bonds to form aglycones and, then, the latter, after prolonged fermentation, may be further be converted into smaller phenolic compounds [
43,
44]. Moreover, yellow to orange pigments—carotenoids [
45]—can degrade under LAB fermentation conditions, producing volatile low-molecular weight carotenoid derivatives, such as β-ionone and β-damascenone [
46]. Finally, further studies are needed to explain, in detail, the colour changes in lentils during fermentation but we found correlations between the lentil colour characteristics and the main fermentation parameters (pH and LAB viable counts)—exposing that the changes occurring during lentil fermentation are very complex and should be taken into consideration during its processing.
3.2. Free Amino Acid Profile and γ-Aminobutyric Acid (GABA) Concentration in Lentil Wholemeal Samples
The non-essential free amino acid concentrations in lentil samples are presented in
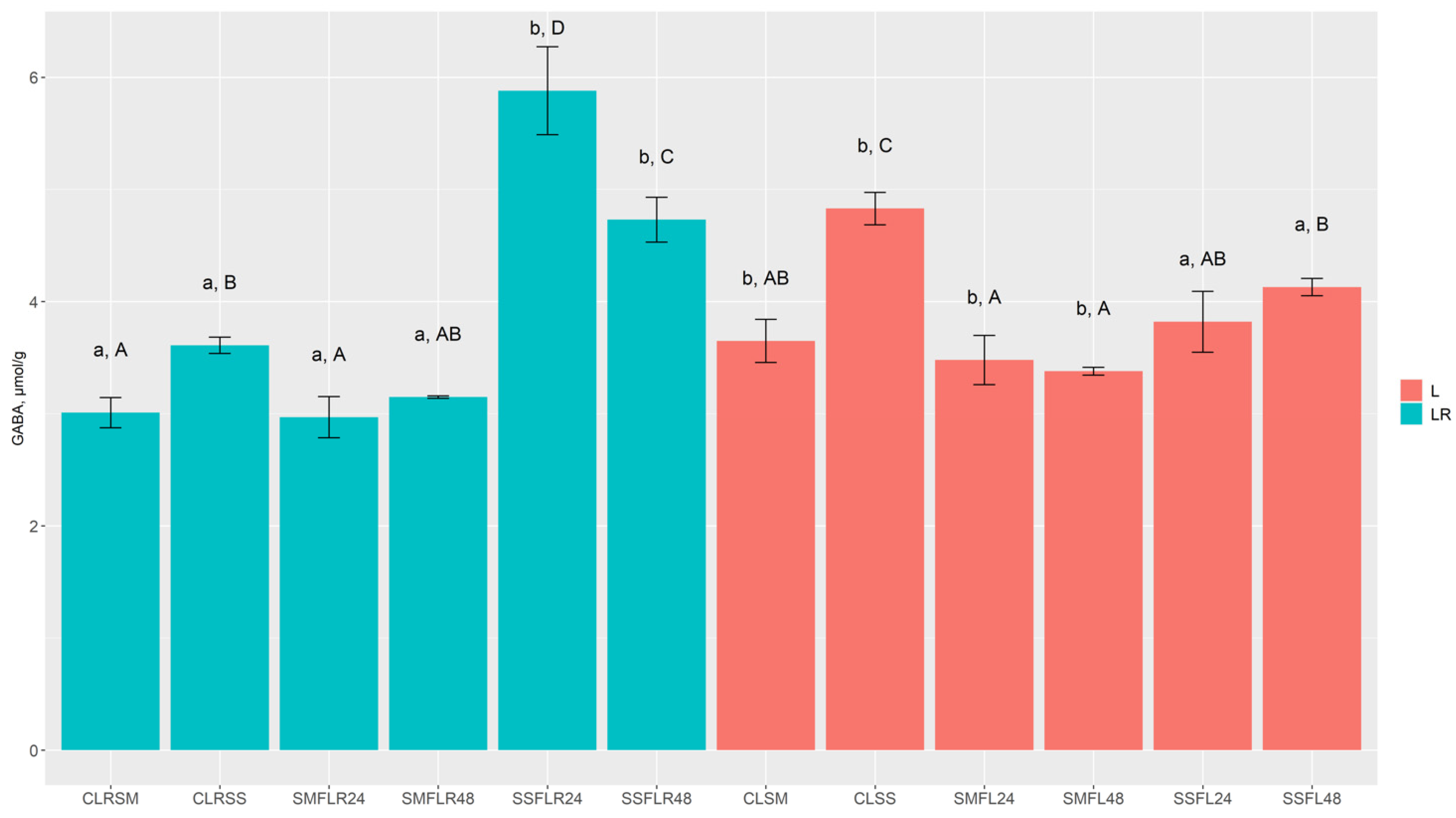
Table 2, and GABA concentrations in lentil samples are presented in
Figure 2. The fermentation process proved to be a significant factor of reduction in arginine, asparagine, glutamine, and aspartic acid concentrations in most of the cases, while glutamic acid, glycine, alanine, and proline concentrations in fermented lentil samples were significantly higher in most cases, in comparison to respective control samples. Moreover, serine concentration in SMF samples (SMF
LR24, SMF
LR48, SMF
L24, and SMF
L48) was significantly lower, while its concentration in SSF samples (SSF
LR24, SSF
LR48, SSF
L24, and SSF
L48) was significantly higher than the respective control sample groups. Additionally, SMF
L24, SMF
L48, SSF
L24, and SSF
L48 samples contained significantly lower concentration of GABA, while its concentration was significantly higher in most of SMF and SSF LR samples, in comparison with the respective control samples.
L control samples (CLSM and CLSS) contained significantly higher concentrations of arginine, asparagine, and alanine than the LR control samples. Meanwhile, LR control samples (CLRSM and CLRSS) contained significantly higher concentrations of glutamic acid, in comparison with L control samples. Arginine concentration in almost all fermented samples was reduced to undetectable levels. L SMF samples (SMFL24 and SMFL48) contained significantly higher concentrations of glutamine, asparagine, serine, aspartic acid, glycine, alanine, proline, and GABA, in comparison to LR SMF samples (SSFLR24 and SSFLR48), while glutamic acid was significantly higher only in SMFL48 samples. Moreover, L SSF samples (SSFL24 and SSFL48), in comparison with SSFLR24 and SSFLR48, contained significantly higher concentrations of asparagine and aspartic acid, while glutamine and proline showed significantly higher concentrations only in SSFL48 samples. Furthermore, significantly higher concentrations of GABA were observed in LR SSF samples (SSFLR24 and SSFLR48), in comparison with SSFL24 and SSFL48, while only SSFLR24 samples contained significantly higher concentrations of glutamine, alanine, and tyrosine.
Correlations between FAA and GABA concentrations and pH and LAB count are presented in
supplementary file: Table S4. Only glutamine and arginine showed positive correlations with samples’ pH (r = 0.784,
p < 0.001 and r = 0.479,
p = 0.003, respectively) and negative correlations with LAB count (r = −0.892,
p < 0.001 and r = −0.593,
p < 0.001, respectively). On the other hand, glutamic acid, aspartic acid, glycine, alanine, proline, and tyrosine showed negative correlations with lentil samples’ pH (r = −0.828,
p < 0.001; r = −0.758,
p < 0.001; r = −0.431,
p = 0.009; r = −0.360,
p = 0.031; r = −0.608,
p < 0.001; and r = −0.442,
p = 0.007, respectively) and negative correlations with LAB count (r = 0.869,
p < 0.001; r = 0.794,
p < 0.001; r = 0.369,
p = 0.027; r = 0.329,
p = 0.050; r = 0.472,
p = 0.004; and r = 0.433,
p = 0.008, respectively).
The significance of analysed factors and their interactions on FAA and GABA concentrations are presented in
supplementary file: Table S3. The growing conditions factor was statistically significant for almost all nonessential FAA concentrations, except for glycine and alanine. The type of fermentation factor was statistically significant for all nonessential FAA concentrations. The fermentation duration factor was significant for almost all FAA concentrations, except for asparagine. The interaction of growing conditions * type of fermentation was significant for almost all nonessential FAAs, except for glutamic acid, aspartic acid, proline, and tyrosine. The interaction of growing conditions * fermentation duration was statistically significant for arginine, glutamine, serine, GABA, and tyrosine, while the interaction of type of fermentation * fermentation duration was significant for almost all nonessential FAAs, except for asparagine, alanine, and proline. Moreover, the interaction of growing conditions * type of fermentation * fermentation duration was statistically significant for arginine, glutamine, GABA, and tyrosine.
The essential free amino acid concentrations in lentils are depicted in
Table 3. All of the fermented samples contained significantly higher concentrations of threonine, valine, phenylalanine, and leucine/isoleucine, while methionine and lysine were significantly higher for most of the fermented sample groups, in comparison to the respective control groups. However, histidine concentrations were not significantly higher in fermented sample groups, in comparison to respective control groups.
L SMF samples (SMFL24 and SMFL48) contained significantly higher concentrations of threonine, valine, and lysine, in comparison to SMFLR24 and SMFLR48, respectively, while methionine, phenylalanine, and leucine/isoleucine contained significantly higher concentrations only in SMFL48 samples. Furthermore, L SSF samples (SSFL24 and SSFL48), in comparison with SSFLR24 and SSFLR48, respectively, contained significantly higher concentrations of methionine, while valine, phenylalanine, leucine/isoleucine, and lysine concentrations were significantly higher only in SSFL48 samples. Moreover, histidine concentration in SSFLR24 samples was significantly higher than in the SSFL24 samples.
Threonine, valine, phenylalanine, leucine/isoleucine, and lysine showed negative correlations with pH (r = −0.670, p < 0.001; r = −0.709, p < 0.001; r = −0.740, p < 0.001; r = −0.808, p < 0.001; and r = −0.661, p < 0.001, respectively) and positive correlations with LAB count (r = 0.609, p < 0.001; r = 0.693, p < 0.001; r = 0.714, p < 0.001; r = 0.783, p < 0.001; and r = 0.537, p < 0.001, respectively).
Growing conditions, the type of fermentation, and fermentation duration factors were statistically significant for all essential FAAs, except histidine. The interaction of growing conditions * type of fermentation was significant for threonine, leucine/isoleucine, lysine, and histidine, while the interaction of growing conditions * fermentation duration was statistically significant for all essential FAAs, except for threonine and methionine. Moreover, the interaction of type of fermentation * fermentation duration was significant for all essential FAAs, except for lysine.
Microbial protease enzymes can effectively break down protein to produce polypeptides and free amino acids [
22]. Moreover, bacteria can produce essential and non-essential amino acids by utilizing various products from the glycolysis pathway [
47]. Therefore, most the changes in free amino acid composition after fermentation can be linked to the activity of bacterial proteases and other metabolic processes. The different proteolytic activities of LAB are the primary cause of the variation in the degree of protein hydrolysis [
22]. Varied proteolytic activities and the ability to produce extracellular amino acids was reported for
Pediococcus acidilactici and
Pediococcus pentosaceus [
48].
Furthermore, it has been noted that SSF, in opposite to SMF, is often associated with higher yields of fermentation products and, despite the exact reasoning not being well understood, in the literature, it is often associated with SSF conditions being similar to the natural conditions [
49,
50,
51].
Arginine consumption during fermentation can be explained by BA production [
52]. Asparagine and glutamine reduction could be explained by the activity of bacterial asparaginases and glutaminases in converting to aspartic and glutamic acid, respectively [
53]. Glutamic acid is regarded as an intermediary amino acid that is often involved as an amino group donor for aminotransferases, while it is also consumed by the bacteria to produce GABA via decarboxylation [
54]. Some species of LAB, e.g.,
Lactobacillus and
Leuconostoc, with glutamate decarboxylase activity, also contribute to GABA production [
55]. GABA is an important compound towards the inhibition of neurotransmitters, and, when implemented into the diet, can reduce sleeplessness, depression, and anxiety, enhance immunity, regulate blood pressure, etc. [
54,
55].
3.3. The Concentration of Biogenic Amines in Lentil Wholemeal Samples
Biogenic amine concentration in lentil samples is tabulated in
Table 4. Tryptamine (TRIP), cadaverine (CAD), histamine (HIS), and tyramine (TYR) were not detected in any lentil groups. Phenylethylamine (PHE) was found only in SMF
LR24 samples. In most of the cases, fermented samples contained higher concentrations of other detected biogenic amines—chiefly, putrescine (PUT), spermidine (SPRMD), and spermine (SPRM)—in comparison to the respective sample control groups. In this regard, total BA concentration in L and LR SMF and SSF sample groups were significantly higher in than the respective control sample groups.
All factors and their interactions were statistically significant on PHE concentration in lentil samples (see
supplementary file: Table S6). Moreover, all factors and the growing conditions * type of fermentation interaction were statistically significant for PUT concentration. Almost all factors and the interactions thereof, except type of fermentation, were statistically significant for SPRMD concentration. The type of fermentation, the growing conditions * type of fermentation interaction, and the growing conditions * fermentation duration interaction were statistically significant for SPRM concentration in lentil samples. The correlation analysis (see
supplementary file: Table S5) discovered that only PUT and SPRM possessed strong and weak positive correlations with LAB viable counts (r = 0.761,
p < 0.001 and r = 0.331,
p = 0.049, respectively), while only PUT showed a significant strong positive correlation with the pH of the lentils (r = 0.719,
p < 0.001). Strong and weak negative correlations of PUT with arginine and glutamine (r = −0.686,
p < 0.001 and r = −0.341,
p = 0.042, respectively) were established. Moreover, strong positive correlations of PUT with glutamic and aspartic acids, valine, phenylalanine, leucine/isoleucine, and lysine (r = 0.734,
p < 0.001; r = 0.881,
p < 0.001; r = 0.614,
p < 0.001; r = 0.639,
p < 0.001; r = 0.697,
p < 0.001; and r = 0.583,
p < 0.001, respectively), and moderate positive correlations with threonine, glycine, and proline (r = 0.594,
p < 0.001; r = 0.349,
p = 0.037; and r = 0.468,
p = 0.004, respectively) were found. SPRMD showed moderate positive correlations with GABA and tyrosine (r = 0.424,
p = 0.010; and r = 0.534,
p < 0.001, respectively). Also, a weak positive correlation of SPRMD with methionine (r = 0.356,
p = 0.033) was observed. Strong positive correlations were found between SPRM and alanine, proline, glycine, phenylalanine, and histidine (r = 0.787,
p < 0.001; r = 0.604,
p < 0.001; r = 0.644,
p < 0.001; r = 0.610,
p < 0.001; and r = 0.668,
p < 0.001, respectively). Furthermore, moderate positive correlations between SPRM and glutamic acid, serine, threonine, GABA, leucine/isoleucine, tyrosine, valine, and lysine (r = 0.461,
p = 0.005; r = 0.430,
p = 0.009; r = 0.563,
p < 0.001; r = 0.566,
p < 0.001; r = 0.425,
p = 0.010; r = 0.466,
p = 0.004; r = 0.520,
p = 0.001; and r = 0.575,
p < 0.001, respectively) were established. Yet, a weak positive correlation between SPRM and glutamine (r = 0.354,
p = 0.034) was observed.
Aromatic and heterocyclic biogenic amines (TRIP, PHE, HIS, and TYR) are produced from the respective amino acids (i.e., tryptophan, phenylalanine, histidine, and tyrosine, respectively) by decarboxylation with enzymes that are produced by bacteria [
52]. Certain LAB, e.g.,
Lactococci, Pediococcus, and
Lactobacilli, which are decarboxylase-positive, may result in biogenic amine synthesis during fermentation processes [
56]. Some strains of
Pediococcus spp. have been found to accumulate cadaverine and tyramine in alcoholic beverages at low levels [
52]. The aliphatic amine PUT can be formed from arginine via multiple pathways, e.g., arginine decarboxylation to agmatine, which is converted to PUT via agmatine deaminase and PUT carbamoyl transferase; arginine deaminization to citrulline and conversion to PUT via ornithine carbamoyl transferase; arginine conversion to ornithine via arginase/arginine ureohydrolase and conversion to PUT via ornithine decarboxylase; and decarboxylation to agmatine and conversion to PUT via agmatinase/agmatine ureohydrolase. CAD is formed from lysine via lysine decarboxylase [
57]. SPRMD is formed from PUT via spermidine synthase and SPRM is formed from SPRMD via spermine synthase [
58]. The Food and Drug Administration (FDA) of the United Sates of America (USA) and the Food and Agriculture Organization (FAO) of the United Nations (UN), with the World Health Organization (WHO), proposed a maximum acceptable histamine limit of 50 mg/kg in fish and food products [
59]. On the other hand, according to a rat model, the no observed adverse effect level (NOAEL) was set at 2000 ppm for tyramine, putrescine, and cadaverine, 1000 ppm for spermidine, and 200 ppm for spermine [
60].
3.4. Fatty Acid Composition in Lentil Wholemeal Samples
The results of fatty acid profiles in lentil samples are presented in
Table 5. Linoleic acid (C18:2) was the most abundant fatty acid in all of the lentil samples, ranging, on average, from 41.3 to 50.3%. The least abundant fatty acid detected in all samples was saturated stearic acid (C18:0), ranging, on average, from 1.63 to 3.29%.
Palmitic acid’s (C16:0) relative concentration in the SMF and SSF LR groups (SMF
LR24, SMF
LR48, SSF
LR24, and SSF
LR48) was not significantly different from the respective control groups, while SMF
L24, SSF
L24, and SSF
L48 samples contained significantly higher percentages of C16:0, in comparison with respective control. Relative concentrations of C18:0 in SMF and SSF LR samples were not significantly different from the respective control groups (except in SMF
LR48 samples). Moreover, C18:0 relative concentrations in SMF and SSF L samples were significantly higher only in the SMF
LR48, SMF
L24, and SSF
L48 samples, in comparison to respective control groups. Only the SMF
LR24 samples contained significantly lower relative percentages of 9-octadecenoic acid (C18:1), in comparison with the respective control group, while other fermented samples did not show significant differences. Similarly, the C18:1 concentration in fermented L samples did not show significant differences when compared to the respective control groups. Furthermore, only SMF
L24 samples contained a significantly different relative concentration of C18:2, in comparison to the respective controls. α-linolenic acid’s (C18:3 α) relative concentration in SMF and SSF LR samples was not different from the respective controls. On the other hand, its relative concentrations in SMF
L24, SMF
LR24, SSF
L24, and SSF
L48 sample groups were lower than those in the respective controls. Tetradecanoic acid (C14:0), eicosanoic acid (C20:0), and
cis-11-eicosenoic acid (C20:1) were only detected in fermented sample groups. The factor of growing conditions was statistically significant in almost all the content of fatty acids in lentils but C16:0 and C18:0 (see
supplementary file: Table S8). The type of fermentation was a statistically significant factor for almost all fatty acids content except C18:0 and C18:2. Fermentation duration was a statistically significant factor for most fatty acids content except C18:0, C18:1, and C18:2. The factor interactions (growing conditions * type of fermentation and type of fermentation * fermentation duration) were shown to be statistically significant for the most of the fatty acids, but not for C18:1 and C18:2, while the growing conditions * fermentation duration factor’s interaction was only not statistically significant for C18:1 content in lentils. Furthermore, the interaction of the factors growing conditions * type of fermentation * fermentation duration was statistically significant for all fatty acid content in lentils. A weak positive correlation was found between C18:0 concentration and LAB viable counts (r = 0.363,
p = 0.030) in lentils (see
supplementary file: Table S7).
Saturated (SFA), monounsaturated (MUFA), and polyunsaturated (PUFA) fatty acids and omega-3, omega-6, and omega-9 fatty acid concentrations (% of the total fat content) of lentils are displayed in
Table 6. As expected, the predominant groups of fatty acids in lentils were PUFA and omega-6. Only L fermented samples often showed significantly higher SFA contents than in the respective control groups. Other fatty acid groups were not significantly different, in comparison to the non-treated and fermented lentil groups.
It has been reported that C16:0, C18:0, C18:1, C18:2, and C18:3 α are the main fatty acids detected in lentils (
Lens culinaris). The major fatty acid detected in lentil samples was C18:2, as noted in the literature, and, furthermore, driving PUFA were described to be most abundant group of fatty acids in lentils [
5,
61,
62]. Most of the established differences observed between the L and LR sample groups (fermented and non-fermented) can be explained by differences in type of cultivar, genotype, or growing conditions [
63,
64].
3.5. Volatile Compound Profile in Lentil Wholemeal Samples
The volatile compound profiles (% of the total identified VC content) are presented in
Figure 3 (see
supplementary file: Table S11). The most abundant VCs (the total area percentage of which was at least 10% in one of the sample groups) were 1-hexanol, hexanal, hexanoic acid, β-damascenone, pentadecanal, 11-hexadecyn-1-ol, 14-methyl-8-hexadecenal, and eugenol. The most abundant VCs presented as various classes of chemical compounds: alcohols (1-hexanol and 11-hexadecyn-1-ol), aldehydes (hexanal, pentadecanal, and 14-methyl-8-hexadecenal), organic acids (hexanoic acid), ketones (β-damascenone), and allylbenzenes (eugenol). Most of the identified VCs with lower abundances (i.e., an area percentage under 10%) in lentils are aldehydes. Furthermore, in almost all cases, identified VCs with lower abundances in the sample control groups had significantly higher abundances in fermented sample groups. Moreover, the complexity of VC composition in the SSF fermented sample groups (for example, there were 30, 35, 28, and 39 VCs detected in the SSF
LR24, SSF
L24, SSF
LR48, and SSF
L48 samples, respectively) was higher than in the respective control groups (i.e., 25 and 22 VCs were detected in the C
LRSS and C
LSS samples, respectively). In addition, the SSF
LR24 and SSF
LR48 sample groups contained nine newly formed VCs, while the SSF
L24 and SSF
L48 samples contained thirteen and seventeen newly formed VCs, respectively. Moreover, between 38.5 and 55.6% of newly formed VCs detected in SSF samples were aldehydes. However, in SMF sample groups, only three, five, and two newly formed VCs were detected in the SMF
LR24, SMF
L24, and SMF
L48 sample groups, while, in the SMF
LR48 sample group, the VC profile was similar to the non-fermented samples. Most of the SMF samples contained VCs belonging to aldehydes and alkanes for the SMF
LR24 and SMF
L24 sample groups, while alcohols and ketones were the largest newly formed VC classes in the SMF
L48 sample group.
Most of VCs detected in lentils are known to be associated with specific flavour and aroma characteristics. The 1-hexanol is described having light apple, sweet, fruity, ethereal, and herbal flavour notes with herbal and oily aroma characteristics [
65,
66]. Some of the most abundant aldehydes—hexanal and pentadecanal—have similar flavour and aroma traits and are described as fatty, grassy, fresh, and green [
65,
66,
67]. Organic acids, such as hexanoic acid, are described as having sour, fatty, and cheesy flavours and aromas [
65]. β-Damascenone contributes to woody, fruity, stewed apple, and sweet aroma and flavour notes [
65]. Eugenol is known to contribute to flavour and aroma with clove, spicy, and honeylike notes [
65].
The aliphatic aldehydes and ketones, such as hexanal or oct-3-en-2-one, are associated with fatty acid oxidation [
68]. The detected aromatic aldehydes and alcohols, benzaldehyde phenylacetaldehyde and phenylethyl alcohol, showed a statistically significant correlation with the aromatic amino acid phenylalanine (see
supplementary file: Table S11). Aromatic aldehydes and alcohols are associated with the catabolism of aromatic amino acids; in this case, phenylalanine [
69]. The respective amino acid is converted to α-keto acid by the respective aminotransferase, which can be converted to aldehyde by decarboxylases [
53,
69]. Furthermore, the respective aldehyde can be oxidised to form carboxylic acids or reduced to form alcohols [
53,
68]. The ester formation has four main biosynthesis pathways, as follows: alcohol and acid conversion to ester via esterase; hemiacetal, which is formed via spontaneous reaction between aldehyde and alcohol, with dehydrogenation via its dehydrogenases; ketone conversion to ester via Baeyer–Villiger monooxygenases; and alcohol and acyl-CoA conversion to ester via alcohol acyltransferases [
70]. The production of hydrocarbons, such as tetradecane, tridecane, or 1-tetradecene, is associated with fatty acid catabolism [
71]. Finally, all the factors and their interactions were statistically significant for most of the VCs in lentils (see
supplementary file: Table S12).
3.6. Micro- and Macro-Elements Concentration in Lentil Wholemeal Samples
The concentrations of micro- and macro-elements in lentil samples are presented in
Table 7. Almost all concentrations of macro-elements were significantly higher in L samples, in comparison with LR ones, except magnesium. Moreover, of all essential micro-elements, only phosphorus, manganese, and selenium concentrations in the L sample group were significantly higher than in the LR group. Moreover, iron and nickel concentrations in the LR samples were significantly higher than in the L samples.
Lentils are defined as a leguminous crop, containing significant amounts of macro-elements and essential micro-elements, such as phosphorus, potassium, iron, and zinc [
72,
73]. Furthermore, the concentration of essential microelements, such as iron, is high enough for the addition of lentils in a diet to prevent iron deficiency anaemia [
5]. The other minerals that are plentiful in lentils are crucial for important biological functions in humans [
74]. These functions include, but are not limited to, the following: muscle and nerve functioning (Mg and Ca); cell growth, function, and energy production (P); electrolyte balance and muscle contraction (Na); the metabolism of fat and carbohydrates (Cr); the structural components of many enzymes (Mn, Zn, and Cu); etc. [
74,
75].
According to various sources in the literature, the concentrations of various essential microelements can vary within the ranges of 2000–5300 mg/kg, 0.12–0.60 mg/kg, 10.5–29.0 mg/kg, 49–81.4 mg/kg, 1.1–1.8 mg/kg, 9.4–14.3 mg/kg, 36.7–64.2 mg/kg, and 0.18–1.6 mg/kg for phosphorus, chromium, manganese, iron, nickel, copper, zinc, and selenium, respectively [
1,
72,
76,
77,
78]. The mineral composition of lentils can be affected by differences in growing conditions (e.g., phosphorus and other mineral fertilization, water availability, and environmental conditions), plant origin (e.g., genotype and cultivar), or other parameters (e.g., harvesting time) [
73,
79,
80,
81,
82,
83].
However, the aforementioned micronutrients have their bioavailability restrained due to the existing antinutrient factors (phytates, oxalates, and phenolic compounds) [
72,
84]. Mineral bioavailability can be enhanced by employing certain strategies, such as genetic manipulation, dehulling, soaking, enzymatic treatment, or fermentation [
5,
72,
84,
85]. The bioavailability enhancement of minerals via fermentation can be explained by the disruption of plant cell wall structures, production of phytase enzymes, or even the activation of endogenous phytase enzymes, due to the existing low pH environment [
26,
86,
87].

 ,
, 




{kind=link}
{kind=link}
{kind=link}