
Changes in the Quality and Microbial Communities of Precooked Seasoned Crayfish Tail Treated with Microwave and Biological Preservatives during Room Temperature Storage
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Sample Preparation, and Storage
2.2. Determination of Total Viable Count
2.3. Chemical Analyses
2.4. Volatile Compounds
2.5. DNA Extraction and 16S rRNA Sequencing
2.6. Data Processing and Microbial Community Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Changes in the TVC during Room Temperature Storage of Precooked Crayfish Tails
3.2. Changes in pH and TVB-N Values during Room Temperature Storage of Precooked Crayfish Tails
3.3. Changes in Volatile Compounds during Room Temperature Storage of Precooked Crayfish Tails
3.4. Changes in Microbial Community during Room Temperature Storage of Precooked Crayfish Tails
3.5. Correlation of Microorganisms and Flavor Compounds during Room Temperature Storage of Precooked Crayfish Tails
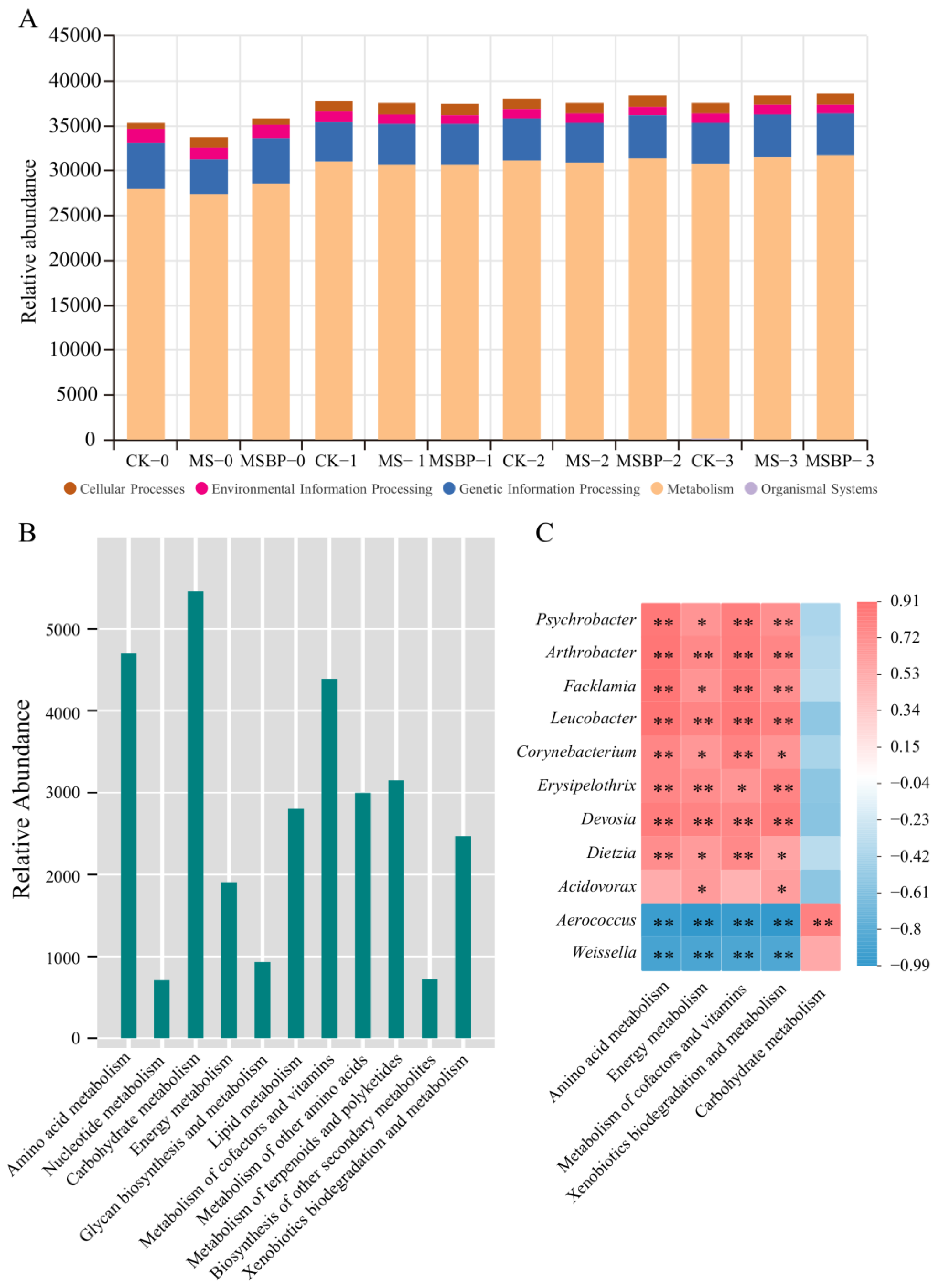
3.6. Predicted Functional Analysis of the Microbial Communities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Odeyemi, O.A.; Dabadé, D.S.; Amin, M.; Dewi, F.; Waiho, K.; Kasan, N.A. Microbial diversity and ecology of crustaceans: Influencing factors and future perspectives. Curr. Opin. Food Sci. 2021, 39, 140–143. [Google Scholar] [CrossRef]
- Yu, X.J.; Hao, X.J.; Yang, L.K.; Dang, Z.Q.; Wang, X.G.; Zhang, Y.H.; Zhang, X. China crayfish industry development report. China Fish 2023, 7, 26–31. (In Chinese) [Google Scholar]
- Cremades, O.; ÁLvarez-Ossorio, C.; Gutierrez-Gil, J.F.; Parrado, J.; Bautista, J. Quality changes of cooked crayfish (Procambarus clarkii) tails without additives during storage under protective atmospheres. J. Food Process. Preserv. 2011, 35, 898–906. [Google Scholar] [CrossRef]
- Soedarini, B.; van Gestel, C.A.M.; van Straalen, N.M.; Widianarko, B.; Röling, W.F.M. Interactions between accumulated copper, bacterial community structure and histamine levels in crayfish meat during storage. J. Sci. Food Agric. 2014, 94, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.C.; Feng, X.P.; Li, W.W.; Yuan, L.; Ge, J.; Lu, D.L.; Chen, B.; Yu, G. Changes in properties of white shrimp (Litopenaeus vannamei) protein during thermal denaturation. Food Sci. Biotechnol. 2016, 25, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.M.; Li, D.Y.; Liu, Z.Q.; Liu, Y.X.; Zhou, J.Z.; Zhang, M.; Zhou, D.Y.; Zhu, B.W. Effects of heat treatments on texture of abalone muscles and its mechanism. Food Biosci. 2021, 44, 101402. [Google Scholar] [CrossRef]
- Jiang, Q.X.; Zhang, J.W.; Gao, P.; Yu, D.W.; Yang, F.; Xu, Y.S.; Xia, W.S.; Chen, N.; Jiao, T.T. Effects of cooking temperature and time on physicochemical, textural, structural, and microbiological features of fresh crayfish (Procambarus clarkii). J. Food Meas. Charact. 2022, 17, 1208–1217. [Google Scholar] [CrossRef]
- Guo, Q.S.; Sun, D.W.; Cheng, J.H.; Han, Z. Microwave processing techniques and their recent applications in the food industry. Trends Food Sci. Technol. 2017, 67, 236–247. [Google Scholar] [CrossRef]
- Jouquand, C.; Tessier, F.J.; Bernard, J.; Marier, D.; Woodward, K.; Jacolot, P.; Gadonna-Widehem, P.; Laguerre, J.-C. Optimization of microwave cooking of beef burgundy in terms of nutritional and organoleptic properties. LWT—Food Sci. Technol. 2015, 60, 271–276. [Google Scholar] [CrossRef]
- Surówka, K.; Rzepka, M.; Özoğul, F.; Özoğul, Y.; Surówka, B.; Ligaszewski, M. Nucleotide degradation, biogenic amine level and microbial contamination as quality indicators of cold-stored rainbow trout (Oncorhynchus mykiss) gravad. Food Chem. 2021, 346, 128904. [Google Scholar] [CrossRef]
- Yan, S.J.; Yu, D.W.; Tang, C.H.; Shen, J.D.; Xu, Y.S.; Xia, W.S.; Jiang, Q.X.; Yang, F. Physicochemical and microbiological changes in postmortem crayfish (Procambarus clarkii) stored at 4 °C and 25 °C. Int. J. Food Sci. Technol. 2022, 57, 2992–3000. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, J.; Zhao, X.R.; Wang, Z.Y. Effects of different freezing rates on the quality changes in cooked crayfish (Procambarus clarkia) meat during frozen storage (−20 °C). Adv. Mater. Res. 2014, 1033, 673–676. [Google Scholar] [CrossRef]
- Li, D.M.; Zhu, Z.W.; Sun, D.W. Effects of freezing on cell structure of fresh cellular food materials: A review. Trends Food Sci. Technol. 2018, 75, 46–55. [Google Scholar] [CrossRef]
- Wu, J.J.; Zang, M.W.; Wang, S.W.; Zhao, B.; Bai, J.; Xu, C.C.; Shi, Y.X.; Qiao, X.L. Nisin: From a structural and meat preservation perspective. Food Microbiol. 2023, 111, 104207. [Google Scholar] [CrossRef] [PubMed]
- Ghomi, M.R.; Nikoo, M.; Heshmatipour, Z.; Jannati, A.A.; Ovissipour, M.; Benjakul, S.; Hashemi, M.; Langroudi, H.F.; Hasandoost, M.; Jadiddokhani, D. Effect of sodium acetate and nisin on microbiological and chemical changes of cultured grass carp (Ctenopharyngodon idella) during refrigerated storage. J. Food Saf. 2011, 31, 169–175. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, B.H.; Yu, J.X.; Guo, Q.Y.; Zheng, Y.; Hu, H.G. Effects of microwave sterilization combined with Nisin and sodium lactate on microorganisms and qualities of seasoned crayfish tails stored at room temperature. Food Ferment. Ind. 2023, 037360. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, J.F.; Xie, J. Progress on odor deterioration of aquatic products: Characteristic volatile compounds, analysis methods, and formation mechanisms. Food Biosci. 2023, 53, 102666. [Google Scholar] [CrossRef]
- Wang, J.Y.; Li, J.; Zhang, Y.; Ai, Y.W.; Wang, H.X.; Han, Y.H.; Hou, W.F. Changes in physicochemical properties, moisture distribution and volatile flavor compounds of yellow catfish: The roles of dominant spoilage bacteria. Food Biosci. 2023, 56, 103331. [Google Scholar] [CrossRef]
- Xu, X.D.; Sun, C.X.; Liu, B.; Zhou, Q.L.; Xu, P.; Liu, M.Y.; Wang, A.M.; Tian, H.Y.; Luo, W.Z.; Jiang, Q. Flesh flavor of red swamp crayfish (Procambarus clarkii Girard, 1852) processing by GS-IMS and electronic tongue is changed by dietary animal and plant protein. Food Chem. 2022, 373, 131453. [Google Scholar] [CrossRef]
- GB 4789.2-2022; National Standards for Food Safety-Food Microbiology Examination-Determination of Aerobic Plate Count. Standards Press of China: Beijing, China, 2022.
- Rajkumar, U.; Muthukumar, M.; Haunshi, S.; Niranjan, M.; Raju, M.V.L.N.; Rama Rao, S.V.; Chatterjee, R.N. Comparative evaluation of carcass traits and meat quality in native Aseel chickens and commercial broilers. Br. Poult. Sci. 2016, 57, 339–347. [Google Scholar] [CrossRef]
- GB 5009.228-2016; National Standards for Food Safety-Determination of Food Total Volatile Basic Nitrogen. Standards Press of China: Beijing, China, 2016.
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 669–673. [Google Scholar] [CrossRef]
- GB 10136-2015; National Standards for Food Safety-Aquatic Product. Standards Press of China: Beijing, China, 2015.
- Scherer, K.M.; Spille, J.-H.; Sahl, H.-G.; Grein, F.; Kubitscheck, U. The lantibiotic nisin induces lipid II aggregation, causing membrane instability and vesicle budding. Biophys. J. 2015, 108, 1114–1124. [Google Scholar] [CrossRef]
- Feng, X.; Moon, S.H.; Lee, H.Y.; Ahn, D.U. Effect of irradiation on the parameters that influence quality characteristics of raw turkey breast meat. Radiat. Phys. Chem. 2017, 130, 40–46. [Google Scholar] [CrossRef]
- Huang, X.X.; You, Y.; Zeng, X.F.; Liu, Q.Y.; Dong, H.; Qian, M.; Xiao, S.L.; Yu, L.M.; Hu, X. Back propagation artificial neural network (BP-ANN) for prediction of the quality of gamma-irradiated smoked bacon. Food Chem. 2024, 437, 137806. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, A.E.-D.A.; Holman, B.W.B.; Giteru, S.G.; Hopkins, D.L. Total volatile basic nitrogen (TVB-N) and its role in meat spoilage: A review. Trends Food Sci. Technol. 2021, 109, 280–302. [Google Scholar] [CrossRef]
- Dresow, J.F.; Bohm, H. The influence of volatile compounds of the flavour of raw, boiled and baked potatoes: Impact of agricultural measures on the volatile components. Landbauforsch. Volkenrode 2009, 59, 309–338. [Google Scholar]
- Houicher, A.; Bensid, A.; Regenstein, J.M.; Ozogul, F. Control of biogenic amine production and bacterial growth in fish and seafood products using phytochemicals as biopreservatives: A review. Food Biosci. 2021, 39, 100807. [Google Scholar] [CrossRef]
- Wang, M.R.; Kong, Q.; Yu, H.; Jiang, M.Q.; Han, Z.Y. Identification of dominant spoilage bacteria, bacterial diversity analysis, and antibacterial exploration of chilled monkfish. Food Biosci. 2023, 56, 103121. [Google Scholar] [CrossRef]
- Huang, J.Y.; Zhou, Y.Q.; Chen, M.Y.; Huang, J.Q.; Li, Y.; Hu, Y.Q. Evaluation of negative behaviors for single specific spoilage microorganism on little yellow croaker under modified atmosphere packaging: Biochemical properties characterization and spoilage-related volatiles identification. LWT 2021, 140, 110741. [Google Scholar] [CrossRef]
- Cocchi, M.; Biancolillo, A.; Marini, F. Chemometric methods for classification and feature selection. Compr. Anal. Chem. 2018, 82, 265–299. [Google Scholar] [CrossRef]
- Wang, S.Q.; Chen, H.T.; Sun, B.G. Recent progress in food flavor analysis using gas chromatography-ion mobility spectrometry (GC-IMS). Food Chem. 2020, 315, 126158. [Google Scholar] [CrossRef]
- Zang, J.H.; Xu, Y.S.; Xia, W.S.; Regenstein, J.M.; Yu, D.W.; Yang, F.; Jiang, Q.X. Correlations between microbiota succession and flavor formation during fermentation of Chinese low-salt fermented common carp (Cyprinus carpio L.) inoculated with mixed starter cultures. Food Microbiol. 2020, 90, 103487. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, Y.F.; Xu, Y.J.; Zhou, H.; Zhou, K.; Li, C.; Xu, B.C. Microbiota dynamics and volatile metabolite generation during sausage fermentation. Food Chem. 2023, 423, 136297. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Mallouchos, A.; Haroutounian, S.A.; Boziaris, I.S. Microbiological spoilage and investigation of volatile profile during storage of sea bream fillets under various conditions. Int. J. Food Microbiol. 2014, 189, 153–163. [Google Scholar] [CrossRef]
- Liu, S.L.; Chen, S.S.; Shao, L.T.; Ding, Z.X.; Xu, X.L.; Wang, H.H. Spoilage bacteria growth reduction and microbial community variation of chilled chicken packaged in PA/PE treated with pulsed light. Food Control 2024, 157, 110196. [Google Scholar] [CrossRef]
- Yin, H.Y.; Xu, L.B.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Lei, Y.L.; Ai, M.Y.; Lu, S.F.; Xu, H.L.; Wang, L.; Zhang, J.; Xiong, S.B.; Hu, Y. Effect of raw material frozen storage on physicochemical properties and flavor compounds of fermented mandarin fish (Siniperca chuatsi). Food Chem. X 2023, 20, 101027. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.Y.; Xiao, S.T.; Ruan, Q.F.; Gao, Q.; An, Y.Q.; Hu, Y.; Xiong, S.B. Differences in flavor characteristics of frozen surimi products reheated by microwave, water boiling, steaming, and frying. Food Chem. 2022, 372, 131260. [Google Scholar] [CrossRef]
- Sun, D.Y.; Qu, J.J.; Huang, Y.; Lu, J.; Yin, L. Analysis of microbial community diversity of muscadine grape skins. Food Res. Int. 2021, 145, 110417. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Ma, Y.H.; Deng, J.Y.; Ni, Y.S.; Xu, B.C. Monitoring and traceability of spoilage bacteria from three representative braised chickens based on culture-dependent and high-throughput sequencing. LWT—Food Sci. Technol. 2023, 182, 114824. [Google Scholar] [CrossRef]
- Gong, F.S.; Chen, Y.D.; Yang, F.; Jiang, Q.X.; Xu, Y.S.; Xia, W.S. Evaluation of food safety risk of frozen cooked crayfish based on microbial-related indicators. Food Sci. 2023, 44, 60–66. [Google Scholar] [CrossRef]
- Don, S.; Xavier, K.A.M.; Devi, S.T.; Nayak, B.B.; Kannuchamy, N. Identification of potential spoilage bacteria in farmed shrimp (Litopenaeus vannamei): Application of Relative Rate of Spoilage models in shelf life-prediction. LWT—Food Sci. Technol. 2018, 97, 295–301. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Burke, C.M.; Bolch, C.C.J.; Stanley, R. Seafood spoilage microbiota and associated volatile organic compounds at different storage temperatures and packaging conditions. Int. J. Food Microbiol. 2018, 280, 87–99. [Google Scholar] [CrossRef]
- Zhang, J.B.; Li, Y.; Liu, X.C.; Lei, Y.T.; Regenstein, J.M.; Luo, Y.K. Characterization of the microbial composition and quality of lightly salted grass carp (Ctenopharyngodon idellus) fillets with vacuum or modified atmosphere packaging. Int. J. Food Microbiol. 2019, 293, 87–93. [Google Scholar] [CrossRef]
- Rodriguez-Calleja, J.M.; Patterson, M.F.; Garcia-Lopez, I.; Santos, J.A.; Otero, A.; Garcia-Lopez, M.L. Incidence, radioresistance, and behavior of Psychrobacter spp. in rabbit meat. J. Food Prot. 2005, 68, 538–543. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Mallouchos, A.; Haroutounian, S.A.; Boziaris, I.S. Volatile organic compounds of microbial and non-microbial origin produced on model fish substrate un-inoculated and inoculated with gilt-head sea bream spoilage bacteria. LWT—Food Sci. Technol. 2017, 78, 54–62. [Google Scholar] [CrossRef]
- Das, J.; Mishra, H.N. A comprehensive review of the spoilage of shrimp and advances in various indicators/sensors for shrimp spoilage monitoring. Food Res. Int. 2023, 173, 113270. [Google Scholar] [CrossRef]
- Lou, X.W.; Hai, Y.W.; Le, Y.; Ran, X.L.; Yang, H.S. Metabolic and enzymatic changes of Shewanella baltica in golden pomfret broths during spoilage. Food Control 2023, 144, 109341. [Google Scholar] [CrossRef]
- Li, J.H.; Huang, J.; Li, C.L.; Zhang, Y.W.; Wang, Y.Y.; Hou, S.Q.; Cheng, Y.X.; Li, J.Y. Evaluation of the nutritional quality of edible tissues (muscle and hepatopancreas) of cultivated Procambarus clarkii using biofloc technology. Aquac. Rep. 2021, 19, 100586. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total Viable Count (log CFU/g) | |||
|---|---|---|---|---|
| Day 0 | Day 1 | Day 2 | Day 3 | |
| CK | 4.82 ± 0.11 a | 7.98 ± 0.01 a | 9.52 ± 0.06 a | 10.81 ± 0.02 a |
| MS | 1.43 ± 0.60 b | 2.40 ± 0.02 b | 6.62 ± 0.08 b | 7.82 ± 0.06 b |
| MSBP | 1.15 ± 0.21 b | 2.04 ± 0.05 b | 2.67 ± 0.08 c | 4.15 ± 0.01 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, B.; Gao, Y.; Zheng, Y.; Yu, J.; Fu, X.; Bao, H.; Guo, Q.; Hu, H. Changes in the Quality and Microbial Communities of Precooked Seasoned Crayfish Tail Treated with Microwave and Biological Preservatives during Room Temperature Storage. Foods 2024, 13, 1256. https://doi.org/10.3390/foods13081256
Wei B, Gao Y, Zheng Y, Yu J, Fu X, Bao H, Guo Q, Hu H. Changes in the Quality and Microbial Communities of Precooked Seasoned Crayfish Tail Treated with Microwave and Biological Preservatives during Room Temperature Storage. Foods. 2024; 13(8):1256. https://doi.org/10.3390/foods13081256
Chicago/Turabian StyleWei, Banghong, Yan Gao, Yao Zheng, Jinxiang Yu, Xuejun Fu, Hairong Bao, Quanyou Guo, and Huogen Hu. 2024. "Changes in the Quality and Microbial Communities of Precooked Seasoned Crayfish Tail Treated with Microwave and Biological Preservatives during Room Temperature Storage" Foods 13, no. 8: 1256. https://doi.org/10.3390/foods13081256