
Occurrence and Antioxidant Activity of C1 Degradation Products in Cocoa

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cocoa Samples
2.3. Preparation of Procyanidin Model Media
2.4. Extraction of Flavan-3-ols from Cocoa Beans/Chocolate
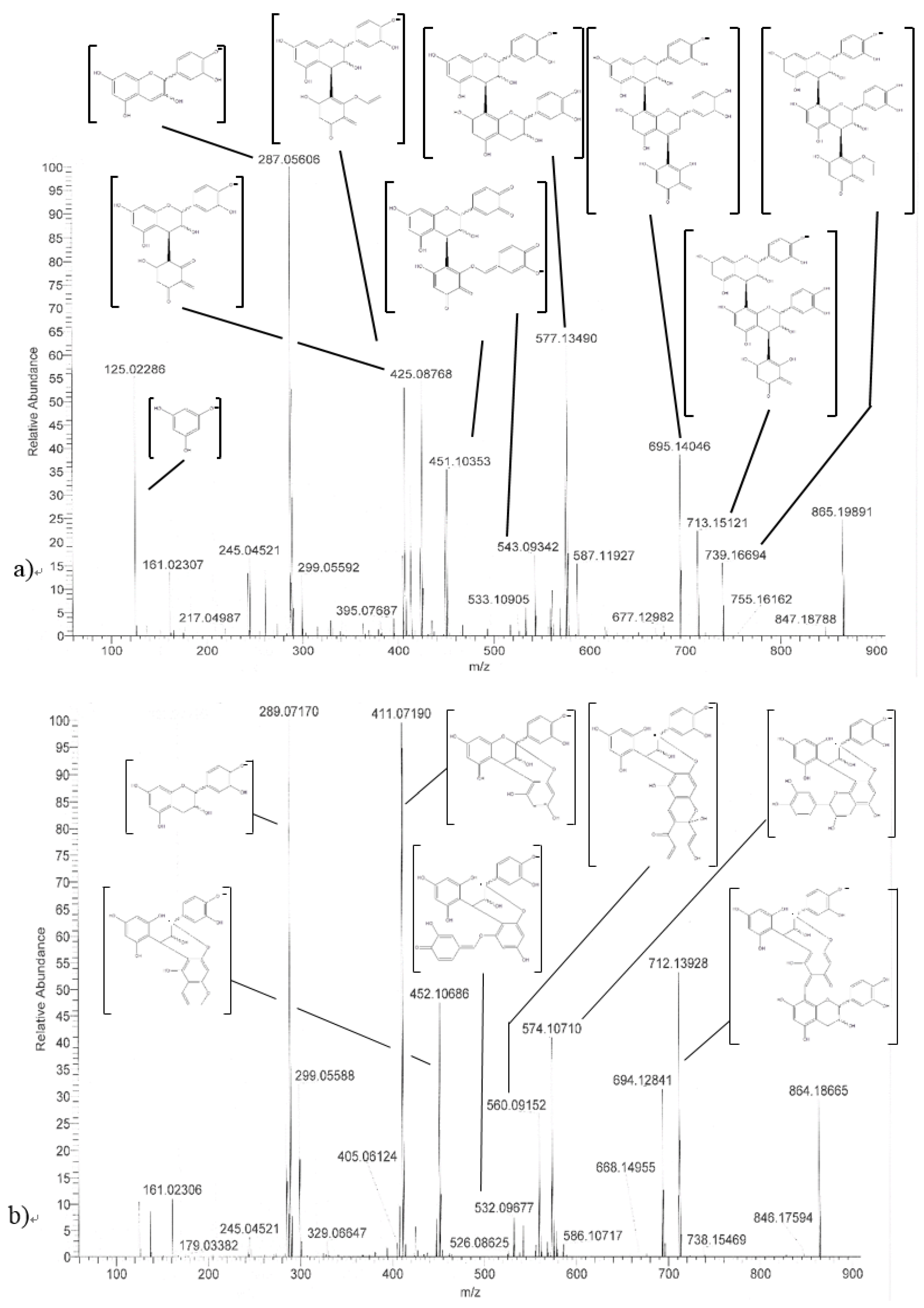
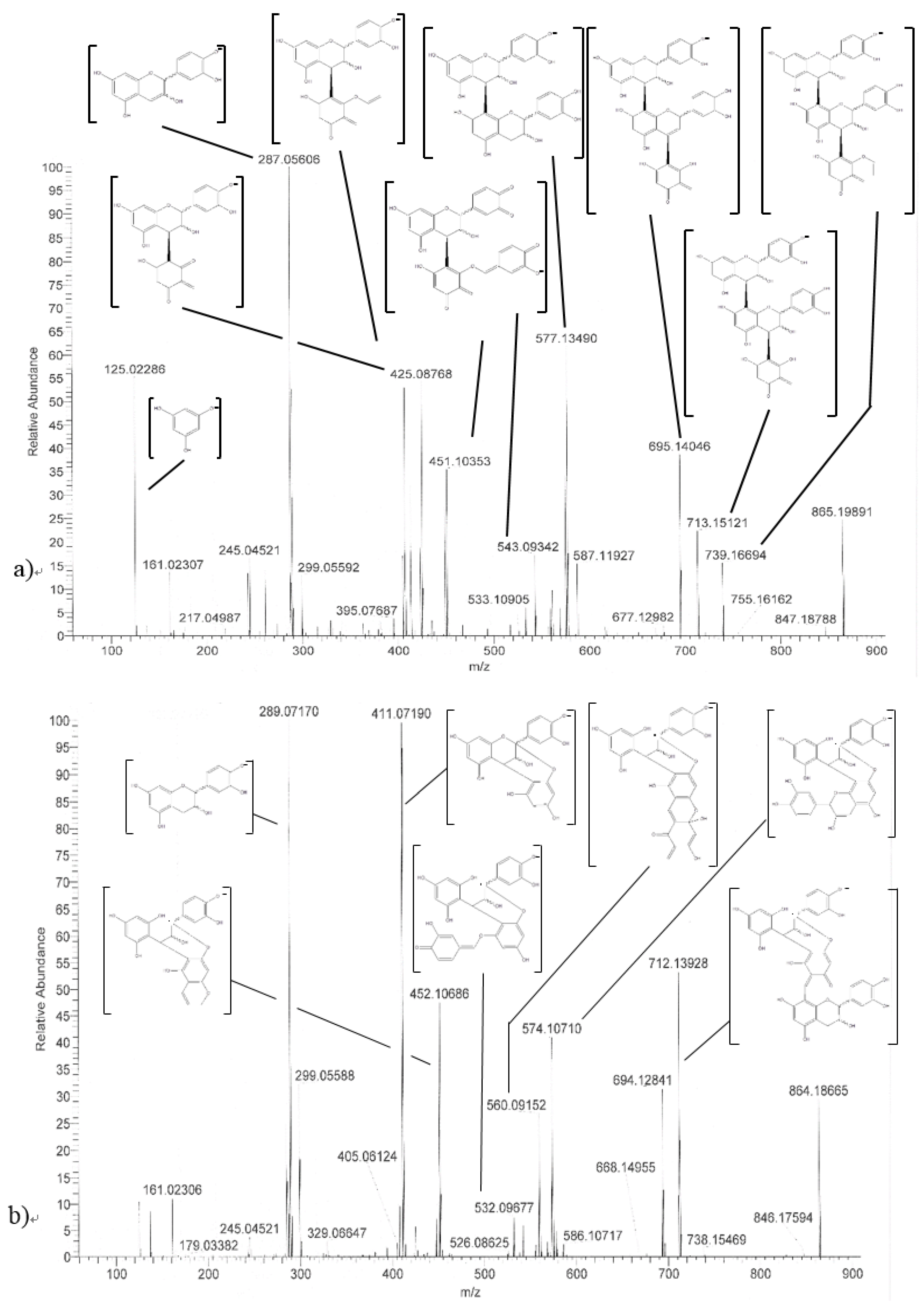
2.5. HRMS/MS Identification.
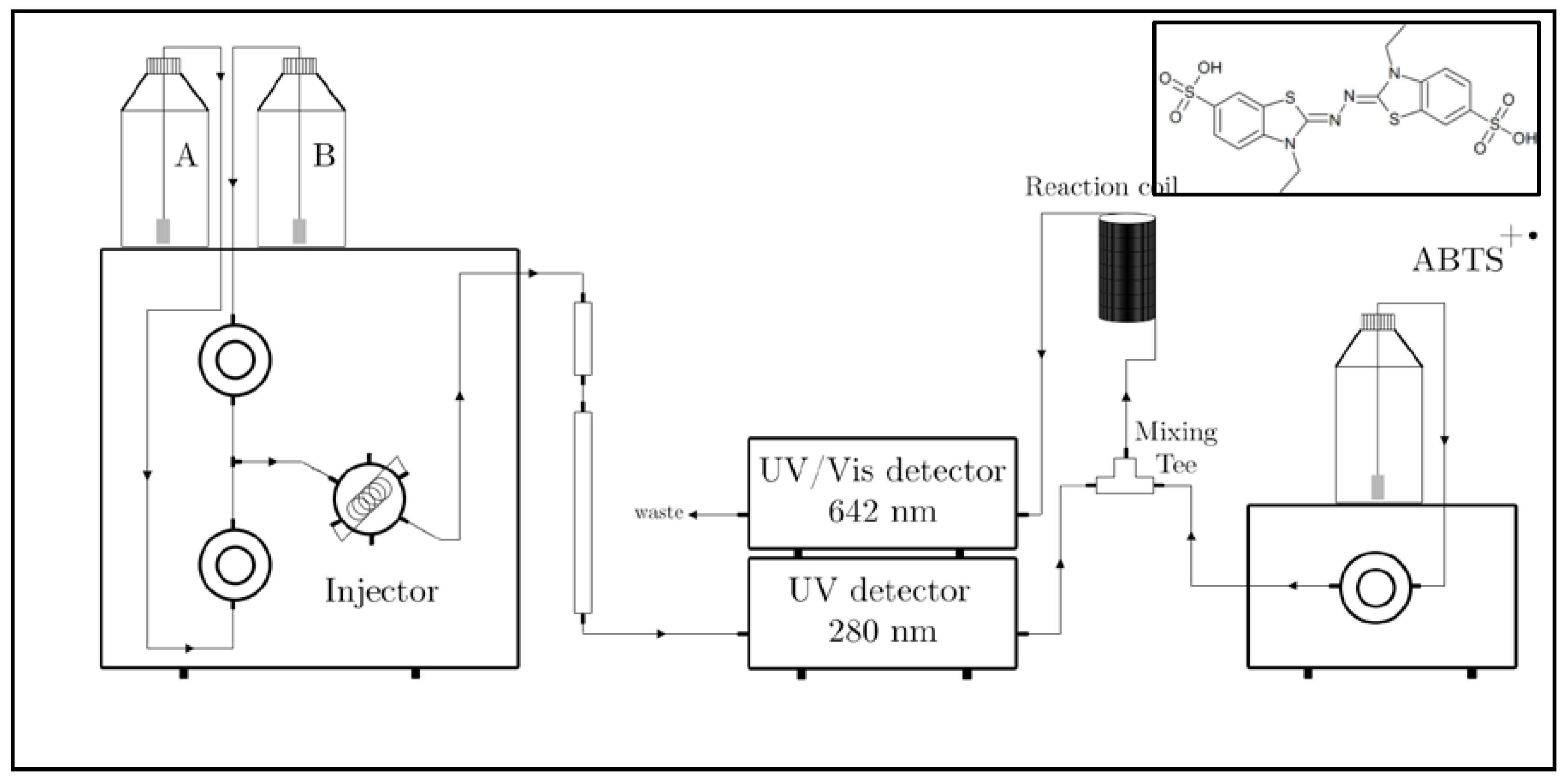
2.6. RP-HPLC-DAD-ESI(−)-MS/MS Semi Quantitation.
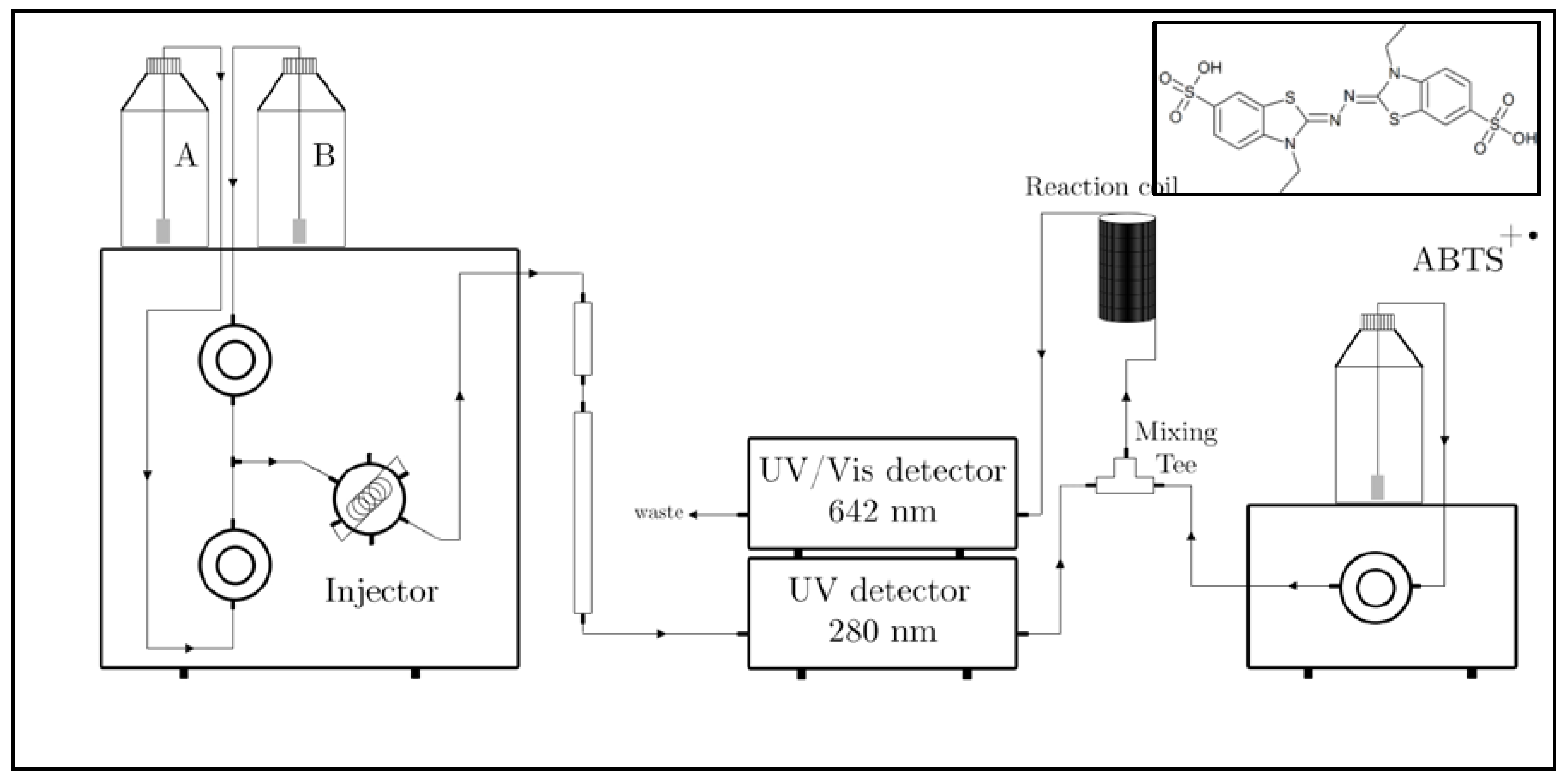
2.7. RP-HPLC-online TEAC
3. Results and Discussion
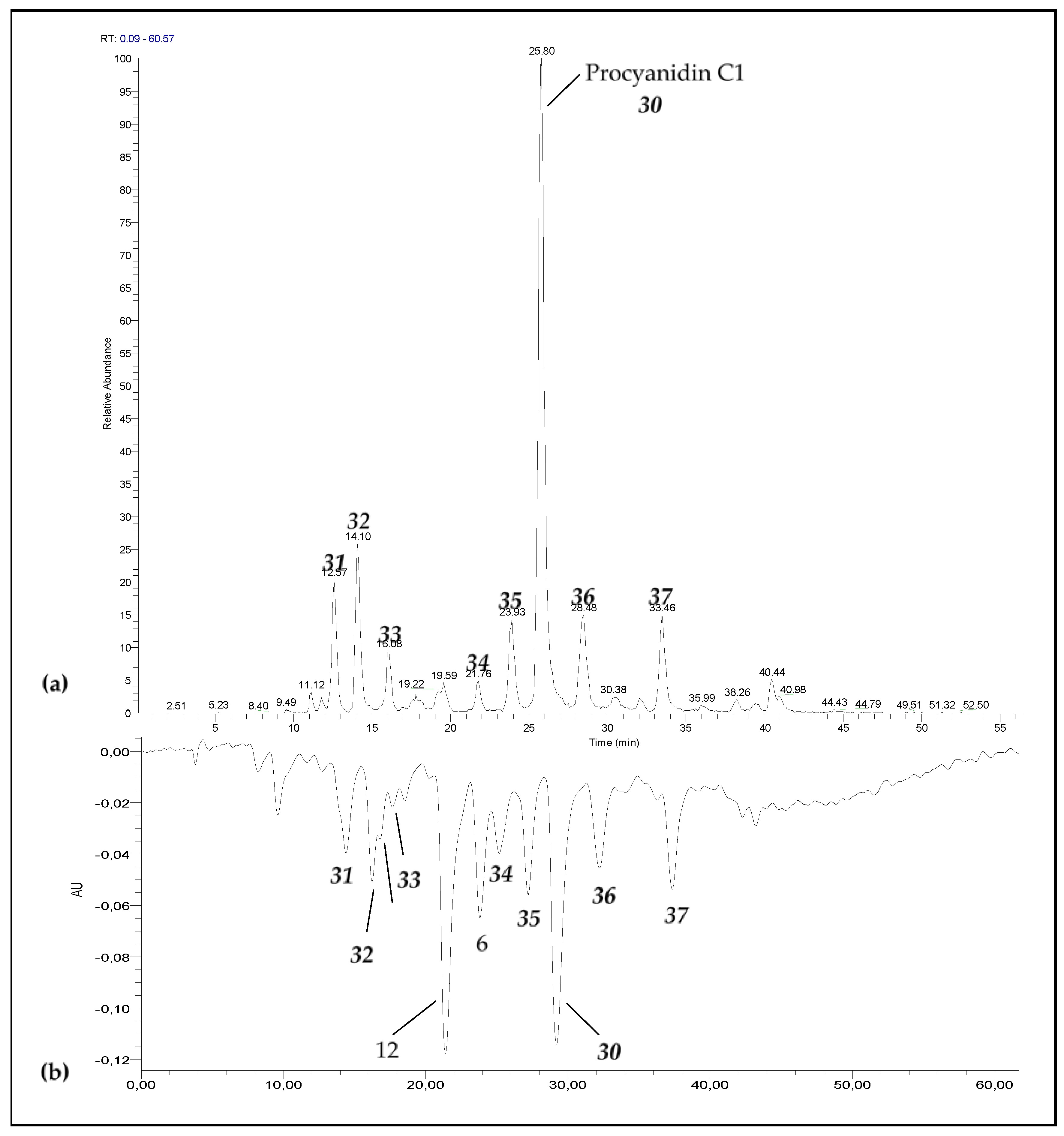
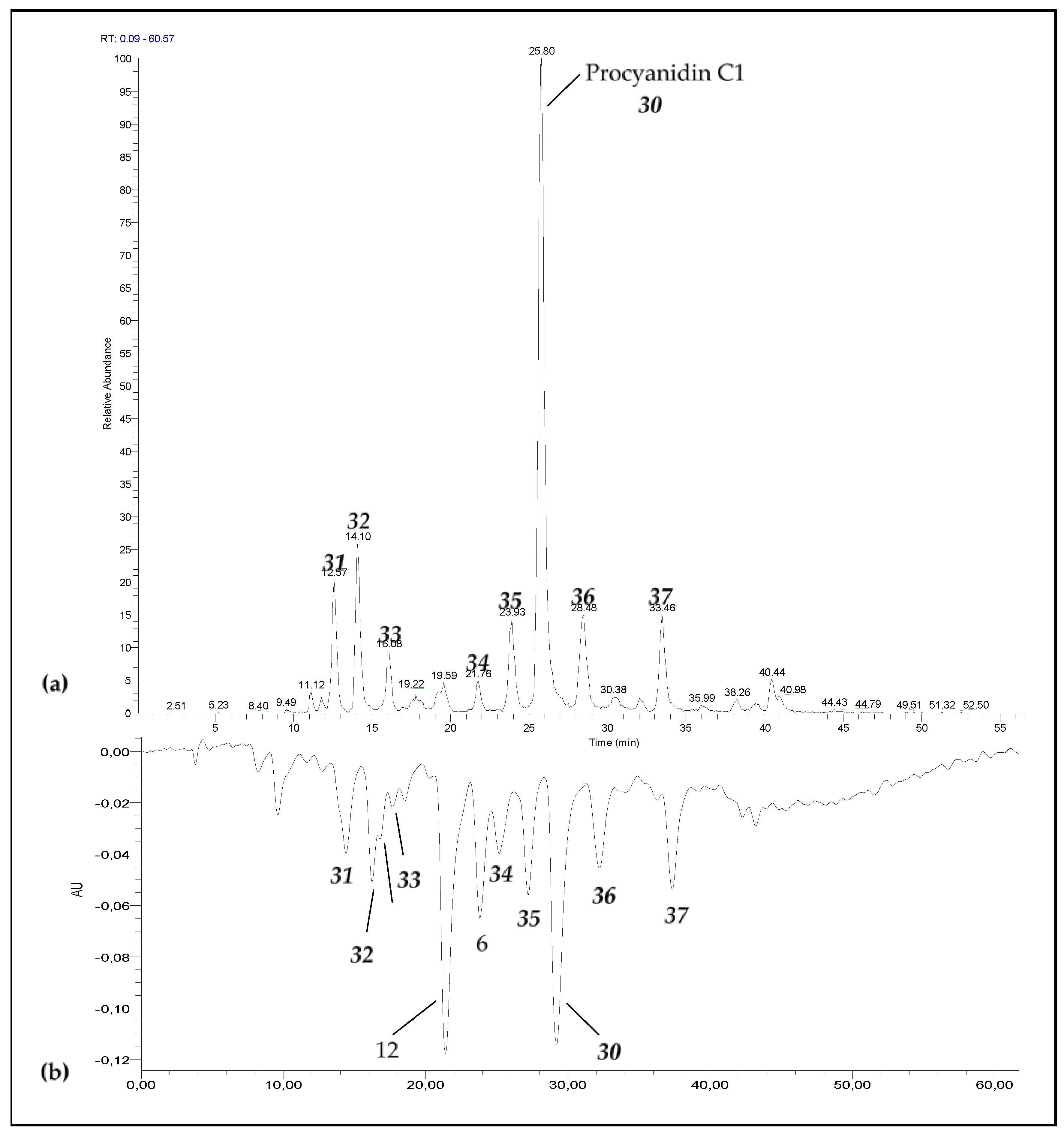
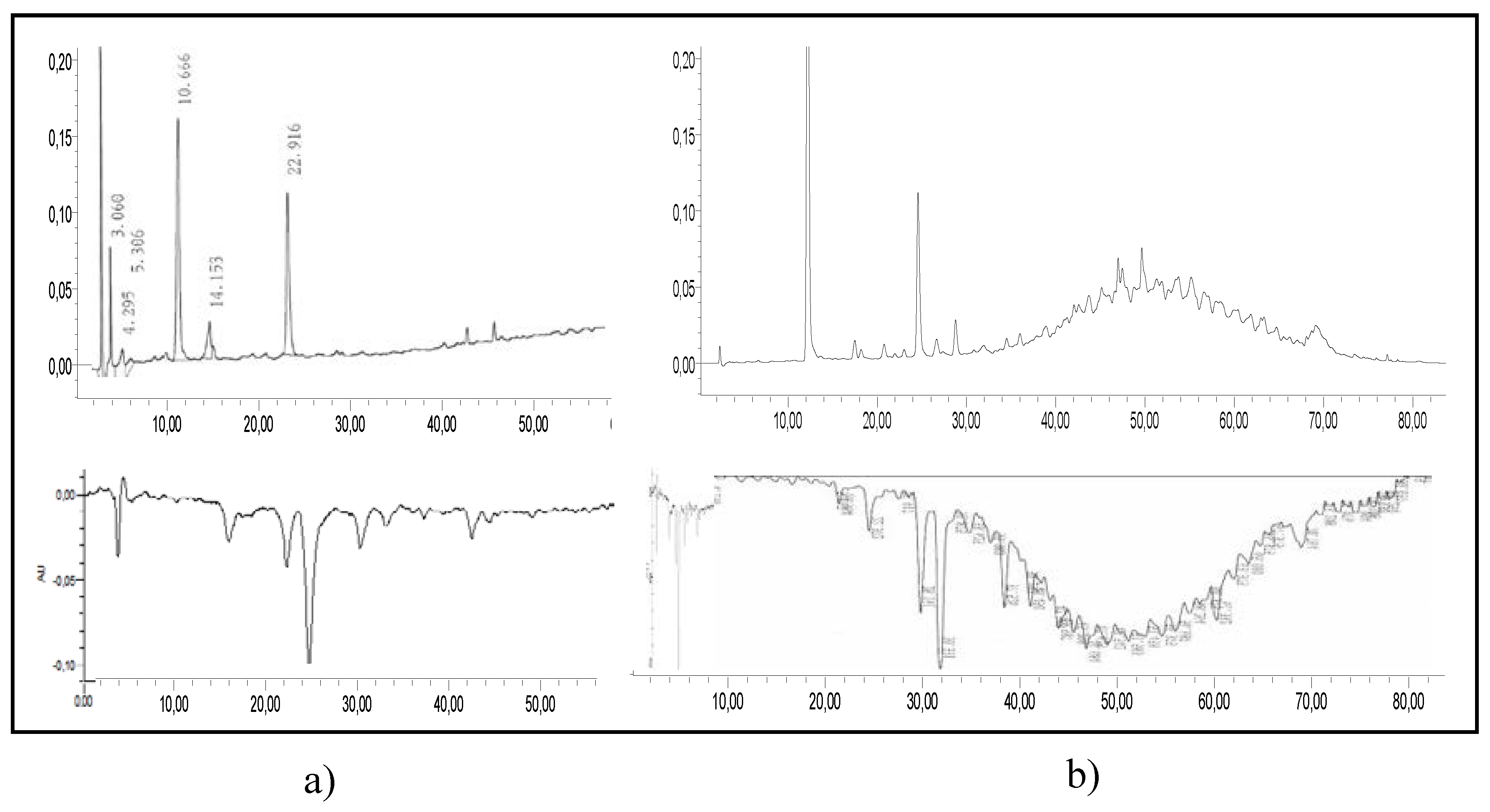
3.1. Degradation of Procyanidin C1 at 90 °C
3.2. Antioxidant Capacity of C1 Degradation Products
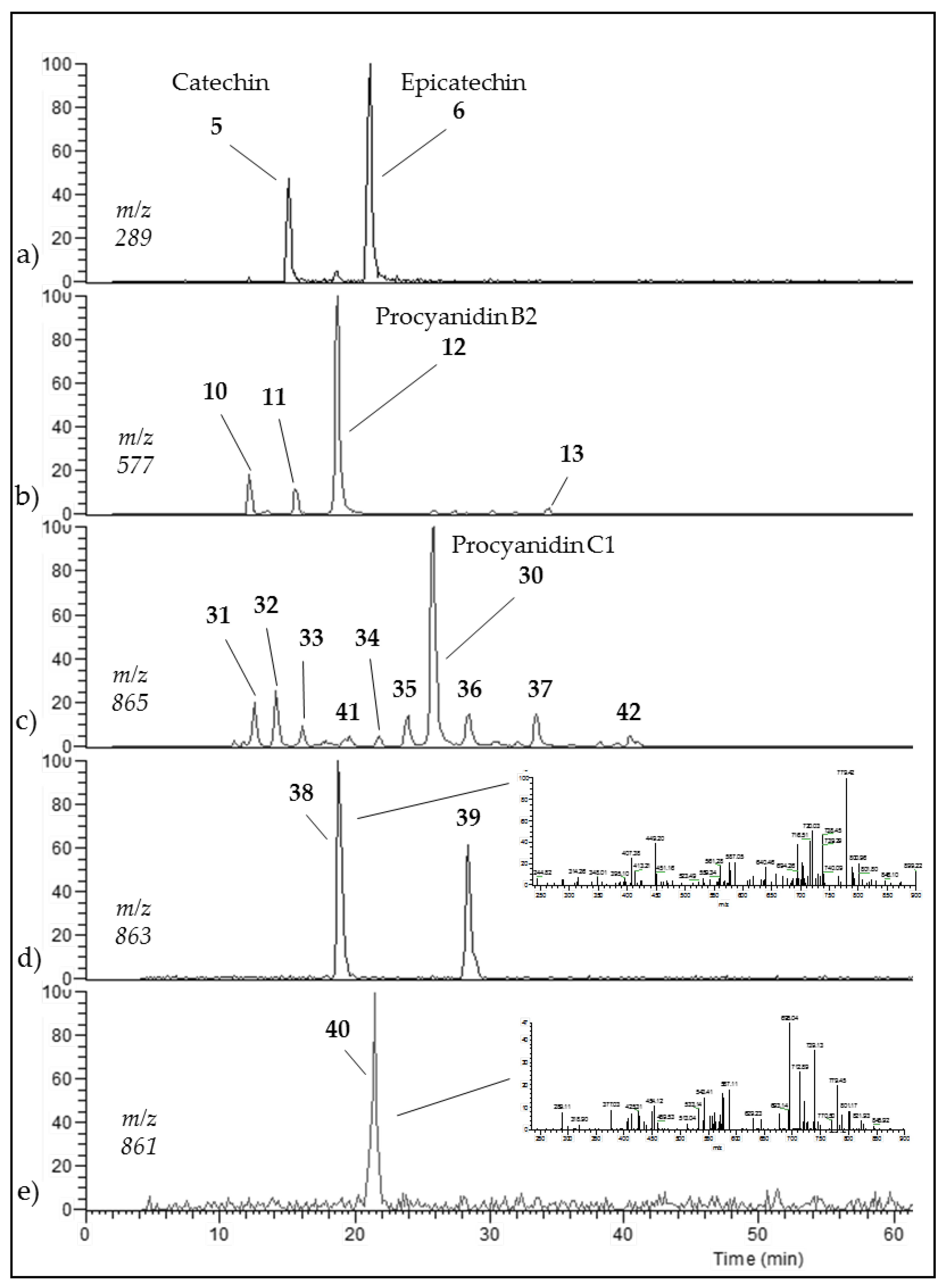
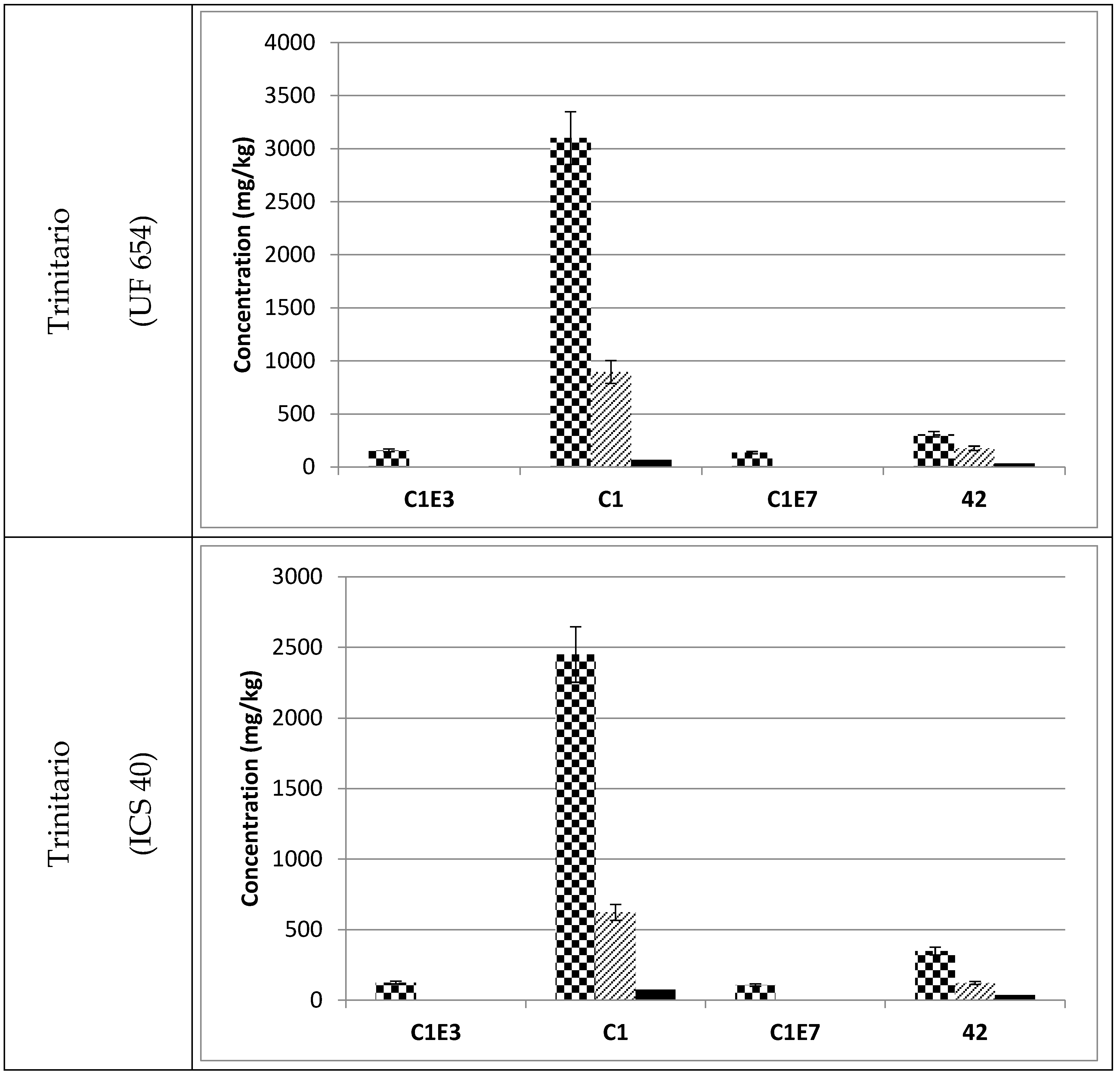
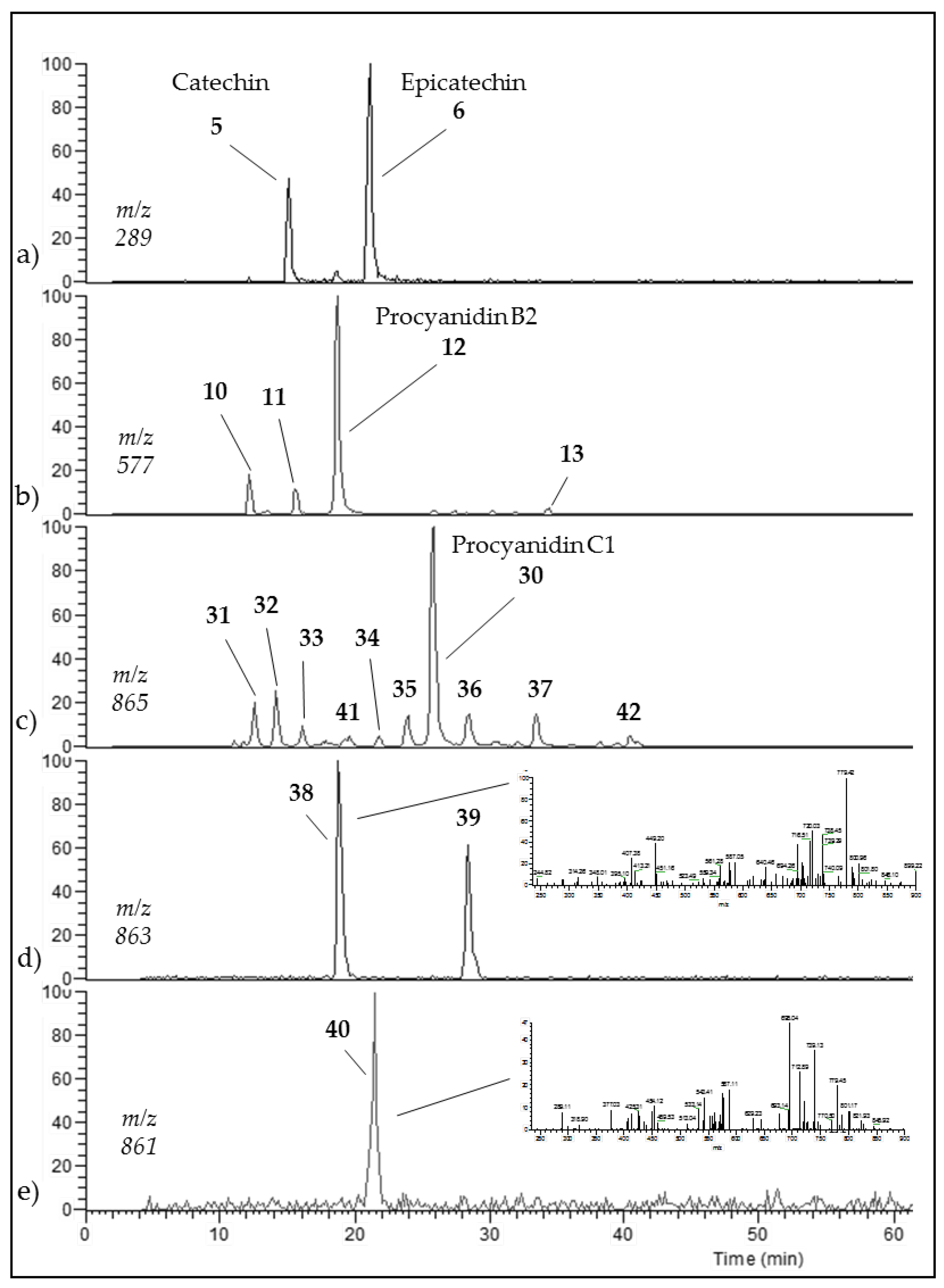
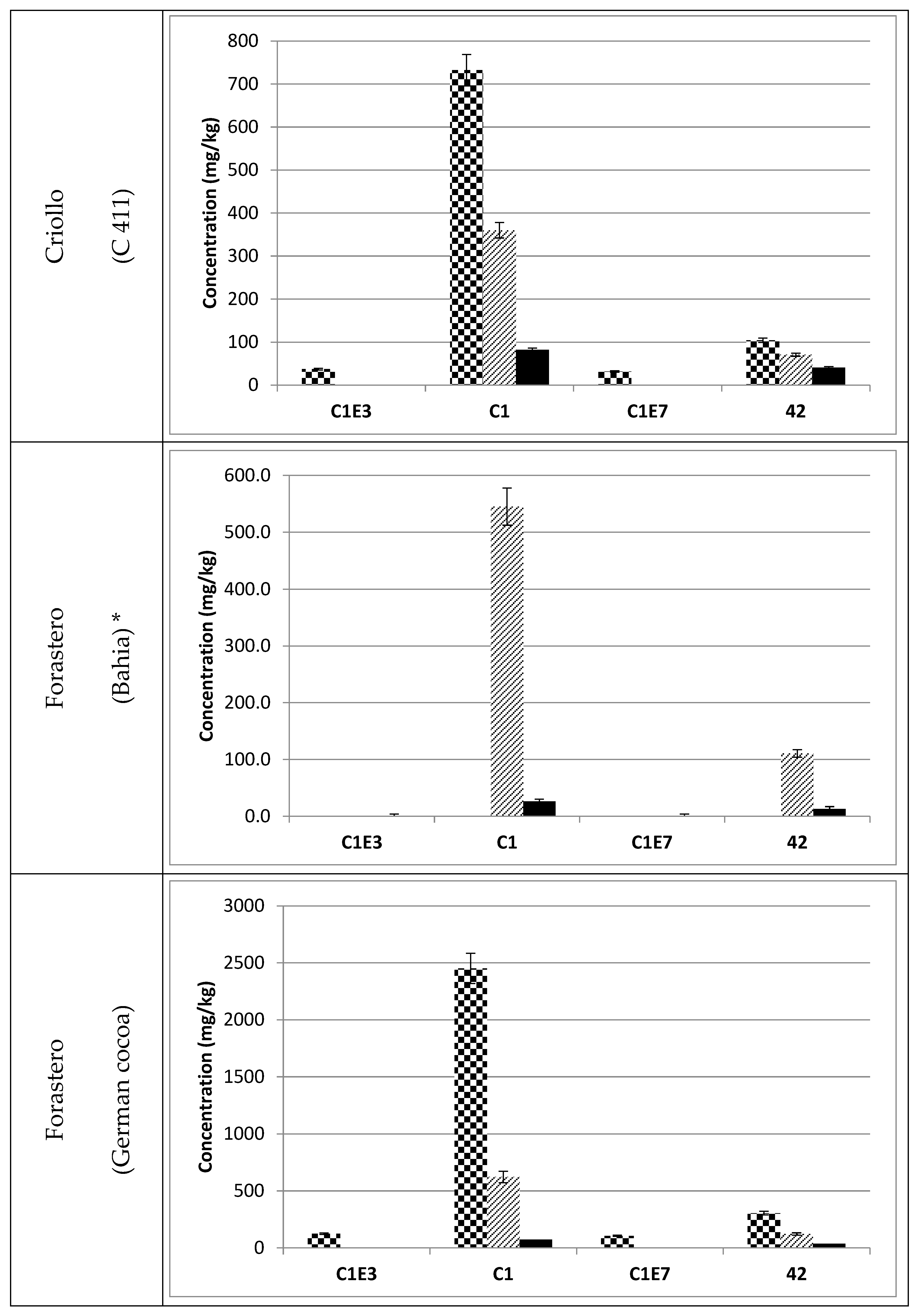
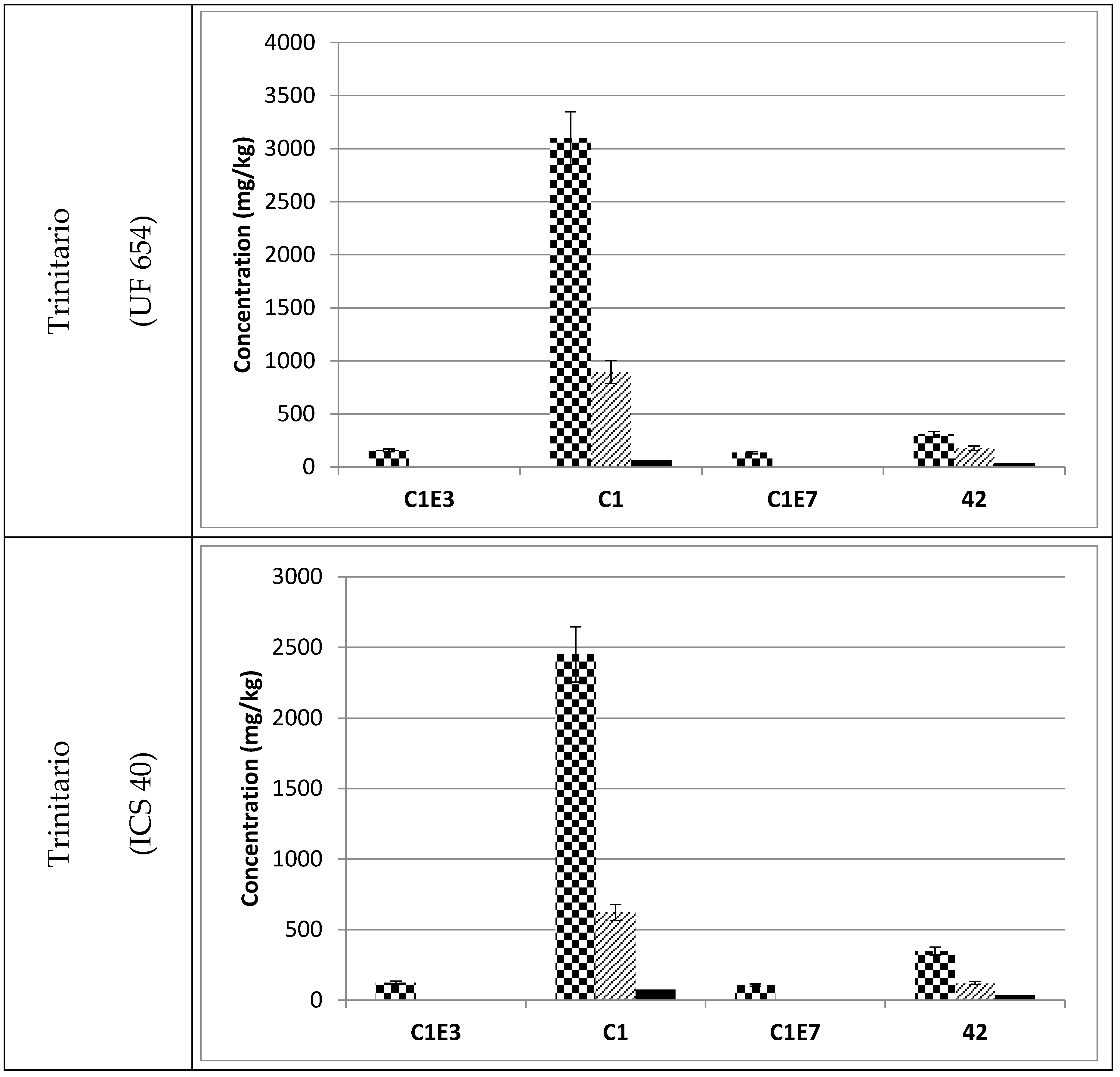
3.3. Occurrence of C1 Degradation Compounds in Cocoa Beans and Chocolate
4. Conclusions
Conflicts of Interest
Abbreviations
| RP-HPLC-HRMS/MS | reversed-phase high-performance liquid chromatographic |
| HRMS/MS | high-resolution mass pectrometry/mass pectrometry |
| ESI | electrospray ionization |
| TEAC | Trolox equivalent antioxidant capacity |
| DPPH | diphenylpicrylhydrazyl |
| SRM | selected reaction-monitoring |
References
- Sanbongi, N.S.; Sakane, T. Polyphenols in chocolate, which have antioxidant activity, modulate immune functions in humans in vitro. Cell. Immunol. 1997, 177, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Kenny, T.P.; Keen, C.L.; Schmitz, H.H.; Gershwin, M.E. Immune effects of cocoa procyanidin oligomers on peripheral blood mononuclear cells. Exp. Biol. Med. 2007, 232, 293–300. [Google Scholar]
- Ramiro-Puig, E.; Castell, M. Cocoa: Antioxidant and immunomodulator. Br. J. Nutr. 2009, 101, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Perez-Berezo, T.; Franch, A.; Ramos-Romero, S.; Castellote, S.; Perez-Cano, F.J.; Castell, M. Cocoa-enriched diets modulate intestinal and systemic humoral immune response in young adult rats. Mol. Nut. Food Res. 2011, 55, S56–S66. [Google Scholar] [CrossRef] [PubMed]
- Dryden, G.W.; Song, M.; McClain, C. Polyphenols and gastrointestinal diseases. Cur. Opin Gastroenterol. 2006, 22, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Andujar, I.; Recio, M.C.; Giner, R.M.; Cienfuegos-Jovellanos, E.; Laghi, S.; Muguerza, B.; Rios, J.L. Inhibition of ulcerative colitis in mice after oral administration of a polyphenol-enriched cocoa extract is mediated by the inhibition of STAT1 and STAT3 phosphorylation in colon cells. J. Agric. Food Chem. 2011, 59, 6474–6483. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Qiao, Z.; Wang, Q.; Zhu, L.; Zhang, L. Flavonoids: Promising anticancer agents. Med. Res. Rev. 2003, 23, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the Phenol-Explorer database. Eur. J. Clin Nutr. 2010, 64, 112–120. [Google Scholar] [CrossRef] [PubMed]
- De Pascual-Teresa, S.; Santos-Buelga, C.; Rivas-Gonzalo, J.C. Quantitative Analysis of Flavan-3-ols in Spanish Foodstuffs and Beverages. J. Agric. Food. Chem. 2000, 48, 5331–5337. [Google Scholar] [CrossRef] [PubMed]
- Lorrain, B.; Ky, I.; Pechamat, L.; Teissedre, P.-L. Identification of the condensed tannins content in grape and Bordeaux wine by means of standards of synthesis. Molecules 2013, 18, 1076–1100. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, K.N. Biosynthesis of proanthocyanidins in barley: Genetic control of the conversion of dihydroquercetin to catechin and procyanidins. Carlsberg Res. Commun. 1984, 49, 503–524. [Google Scholar] [CrossRef]
- Anderson, R.A.; Broadhurst, C.L.; Polansky, M.M.; Schmidt, W.F.; Khan, A.; Flanagan, V.P.; Schoene, N.W.; Graves, D.J. Isolation and characterization of polyphenol type-A polymers from cinnamon with insulin-like biological activity. J. Agric. Food Chem. 2004, 52, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yuan, P.; Chen, K.; Jia, Q.; Li, Y. Oxidative conversion of B- to A-type procyanidin trimer: Evidence for quinone methide mechanism. Food Chem. 2014, 154, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Eyamo Evina, V.J.; De Taeye, C.; Niemenak, N.; Youmbi, E.; Collin, S. Influence of acetic and lactic acids on cocoa flavan-3-ol degration through fermentation-like incubations. LWT Food Sci. Tech. 2016, 68, 514–522. [Google Scholar] [CrossRef]
- De Taeye, C.; Caullet, G.; Bodart, M.; Collin, S. Roasting conditions for preserving cocoa flavan-3-ol monomers and oligomers: Interesting behaviour of Criollo clones. J. Sci. Food Agric. Submitted. [CrossRef] [PubMed]
- Beheydt, B.; Ouwerx, C.; Collin, S.; Deledicque, C.; Nguyen, F. Method to Increase the Antioxidant Activity of Chocolate. U.S. Patent 20110059223 A1, 16 August 2007. [Google Scholar]
- Collin, S.; Callemien, D.; Nizet, S. Extraction et analyse des polyphénols. In Polyphénols et Procédés—Transformation des Polyphénols au Travers des Procédés Appliqués à L’agro-alimentaire; Collin, S., Crouzet, J., Eds.; Tec & Doc Lavoisier: Paris, France, 2011. (In French) [Google Scholar]
- De Taeye, C.; Cibaka, M.L.; Jerkovic, V.; Collin, S. Degradation of (−)-epicatechin and procyanidin B2 in aqueous and lipidic model systems. First evidence of “chemical” flavan-3-ol oligomers in processed cocoa. J. Agric. Food. Chem. 2014, 62, 9002–9016. [Google Scholar] [CrossRef] [PubMed]
- Counet, C.; Collin, S. Effect of the number of flavanol units on the antioxidant activity of procyanidin fractions isolated from chocolate. J. Agric. Food. Chem. 2003, 51, 6816–6822. [Google Scholar] [CrossRef] [PubMed]
- Leitao, C.; Marchioni, E.; Bergaentzlé, M.; Zhao, M.; Didierjean, L.; Taidi, B.; Ennahar, S. Effects of processing steps on the phenolic content and antioxidant activity of beer. J. Agric. Food Chem. 2011, 59, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Bröhan, M.; Jerkovic, V.; Wilmotte, R.; Collin, S. Catechins and derived procyanidins in red and white sorghum. Contribution of them to the antioxidant activity. J. Inst. Brew. 2011, 117, 600–607. [Google Scholar] [CrossRef]
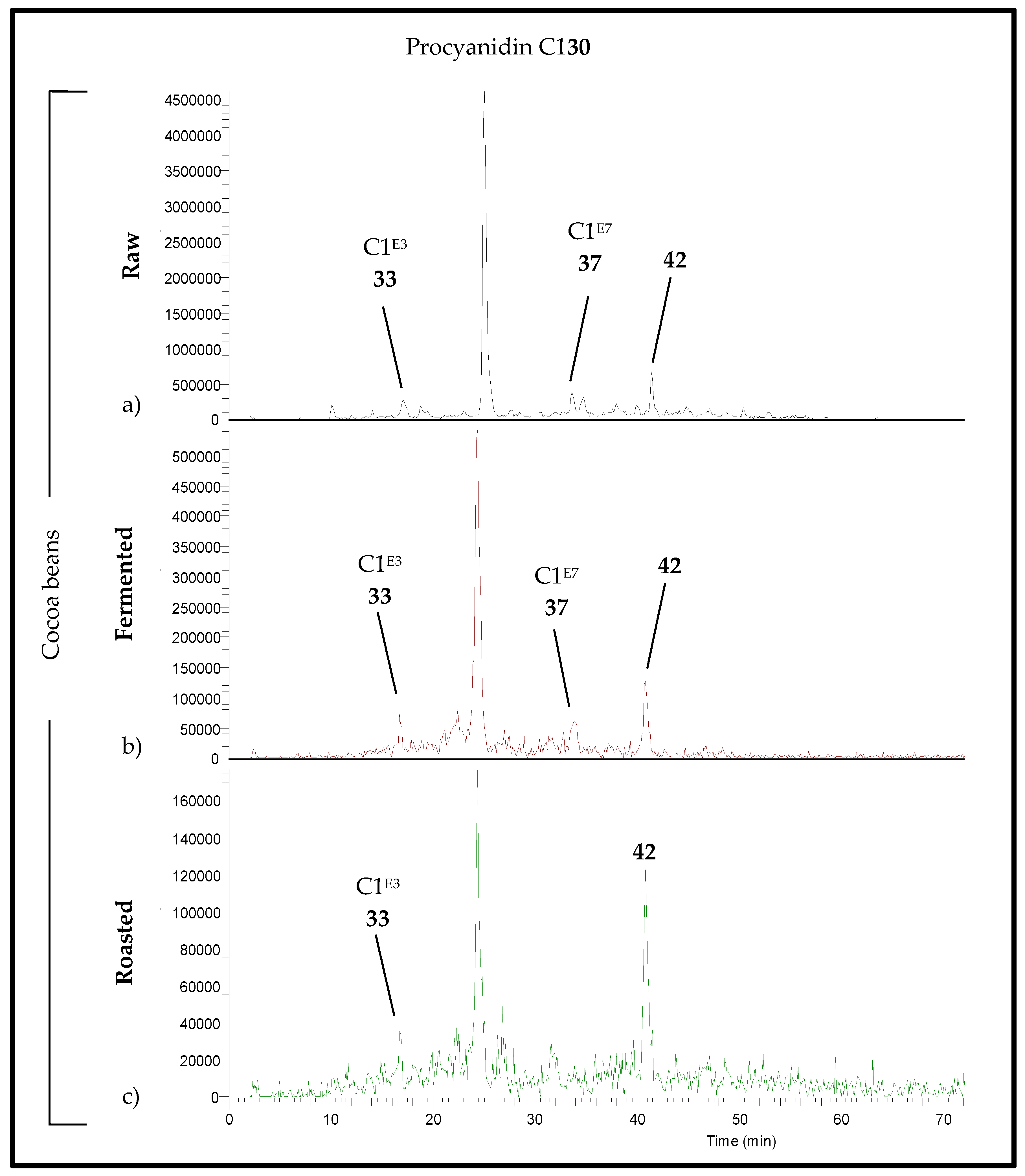
 , fermented
, fermented  and roasted (30 min—150 °C)
and roasted (30 min—150 °C)  cocoa beans. * Raw beans unavailable.
, fermented and roasted (30 min—150 °C) cocoa beans. * Raw beans unavailable.
cocoa beans. * Raw beans unavailable.
, fermented and roasted (30 min—150 °C) cocoa beans. * Raw beans unavailable.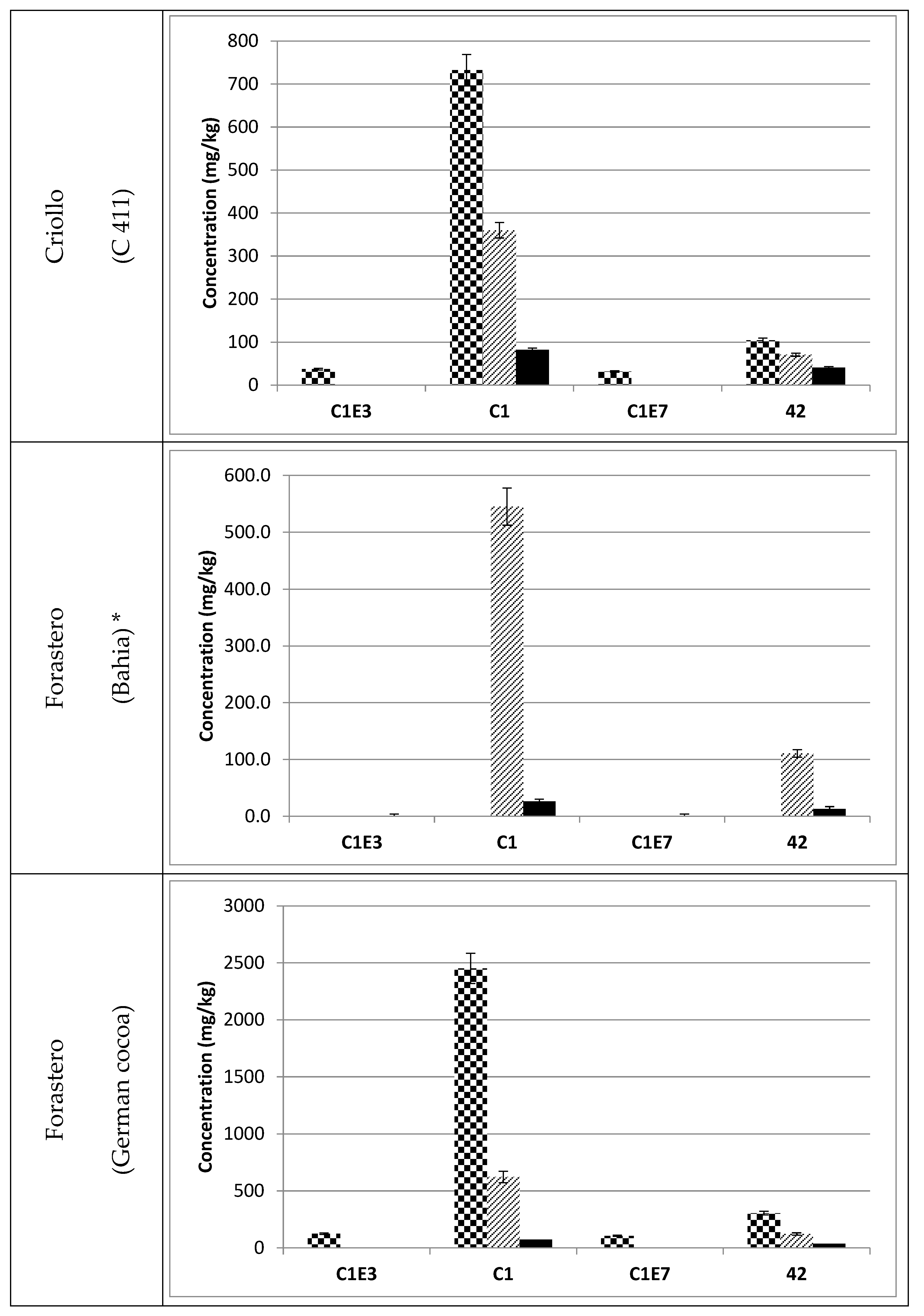

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (peak) | Retention Time (min) | (M−H)−1 | Structure | MS/MS |
|---|---|---|---|---|
| (−)-Catechin (5) | 15.0 | 289 |  | 289.1 (100) 245.1 (20) 179.1 (6) |
| (−)-Epicatechin (6) | 21.1 | 289 |  | 289.1 (100) 245.1 (20) 179.1 (6) |
| Procyanidin B2 epimer (10,11,13) | 12.2 15.2 25.7 | 577 |  | 425.1 (100) 451.1 (65) 407.2 (47) 289.1 (28) |
| Procyanidin B2 (12) | 18.7 | 577 |  | 425.1 (100) 451.1 (65) 407.2 (47) 289.1 (28) |
| Procyanidin C1 (30) | 25.8 | 865 |  | 695.1 (100) 577.1 (64) 739.1 (49) 713.1 (36) 407.2 (29) 575 (25) 425 (21) |
| Procyanidin C1 epimer (31–37) | 12.6 14.1 15.8 21.8 23.9 28.5 33.5 | 865 |  | 695.1 (100) 577.1 (64) 739.1 (49) 713.1 (36) 407.2 (29) 575 (25) 425 (21) |
| Once-oxidized trimer (38,39) | 18.7 28.4 | 863 |  or  | 845.1 (100) 575.1 (78) 289.2 (63) 287.1 (59) 125.1 (28) |
| Twice-oxidized trimer (40) | 21.5 | 861 |  | 843.1 (100), 691.2 (79), 649.1 (38), 575.1 (21). |
| C1 isomer (41,42) | 20.8 40.4 | 865 |  | 289.1 (100), 411.1 (98), 721.1 (53), 574.1 (41), 560.1 (31). |
| Compound (peak) | Source | TEAC | |
|---|---|---|---|
| µM TE/µM | µM TE/mg·kg−1 | ||
| Catechin (5) | a | 0.6 | 2.3 |
| Epicatehin (6) | a | 0.7 | 2.5 |
| Procyanidin B2 (12) | a | 1.3 | 2.4 |
| Procyanidin C1 (30) | a.b | 2.2 | 2.5 |
| C1E1 (31) | b | 2.1 | 2.5 |
| C1E2 (32) | b | 2.2 | 2.6 |
| C1E3 (33) | b | * | * |
| C1E4 (34) | b | 2.1 | 2.5 |
| C1E5 (35) | b | 2.3 | 2.6 |
| C1E6 (36) | b | 2.0 | 2.4 |
| C1E7 (37) | b | 2.2 | 2.5 |
| 38 and 39 | b | * | * |
| 40 | b | * | * |
| 41 and 42 | b | * | * |
| Trolox | a | 1 | 4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Taeye, C.; Cibaka, M.-L.K.; Collin, S. Occurrence and Antioxidant Activity of C1 Degradation Products in Cocoa. Foods 2017, 6, 18. https://doi.org/10.3390/foods6030018
De Taeye C, Cibaka M-LK, Collin S. Occurrence and Antioxidant Activity of C1 Degradation Products in Cocoa. Foods. 2017; 6(3):18. https://doi.org/10.3390/foods6030018
Chicago/Turabian StyleDe Taeye, Cédric, Marie-Lucie Kankolongo Cibaka, and Sonia Collin. 2017. "Occurrence and Antioxidant Activity of C1 Degradation Products in Cocoa" Foods 6, no. 3: 18. https://doi.org/10.3390/foods6030018