Comprehensive Description of Fusarium graminearum Pigments and Related Compounds


1
Laboratory of Food Process Engineering, Graduate School of Agriculture, Hokkaido University, Sapporo 060-0808, Hokkaido, Japan
2
Department of Biological Sciences, Faculty of Sciences, Eduardo Mondlane University, Av. Julius Nyerere, Maputo nr. 3453, Mozambique
Foods 2018, 7(10), 165; https://doi.org/10.3390/foods7100165
Submission received: 14 September 2018
/
Revised: 3 October 2018
/
Accepted: 4 October 2018
/
Published: 5 October 2018
(This article belongs to the Section Food Microbiology)
Abstract
:Several studies have explored in depth the biochemistry and genetics of the pigments present in Fusarium graminearum, but there is a need to discuss their relationship with the mold’s observable surface color pattern variation throughout its lifecycle. Furthermore, they require basic cataloguing, including a description of their major features known so far. Colors are a viable alternative to size measurement in growth studies. When grown on yeast extract agar (YEA) at 25 °C, F. graminearum initially exhibits a whitish mycelium, developing into a yellow-orange mold by the sixth day and then turning into wine-red. The colors are likely due to accumulation of the golden yellow polyketide aurofusarin and the red rubrofusarin, but the carotenoid neurosporaxanthin also possibly plays a major role in the yellow or orange coloration. Torulene might contribute to red tones, but it perhaps ends up being converted into neurosporaxanthin. Culmorin is also present, but it does not contribute to the color, though it was initially isolated in pigment studies. Additionally, there is the 5-deoxybostrycoidin-based melanin, but it mostly occurs in the teleomorph’s perithecium. There is still a need to chemically quantify the pigments throughout the lifecycle, and analyze their relationships and how much each impacts F. graminearum’s surface color.
1. Introduction
Fusarium graminearum (teleomorph: Gibberella zeae) is a pathogen of maize, wheat, rice, and barley responsible for the disease known as Fusarium head blight (FHB) and mycotoxin contamination [1,2]. FHB destroys the grain starch and protein and was responsible for losses of over $2.7 billion in the United States between 1998 and 2000 [2]. The mold’s most common mycotoxins are nivalenol (NIV) and deoxynivalenol (DON) [3], usually occurring together and frequently associated with gastrointestinal disorders, among other health impairments [4]. However, there are other relevant toxins, such as zearalenone (ZEA) [5], an estrogenic compound capable of causing abortion and other reproductive complications [6,7].
There are very few studies comprehensively describing and relating F. graminearum surface colors and its pigments, their properties, and biosynthetic or genetic origin, though some were isolated during the 1930s–1960s [8,9,10,11,12]. There was also considerable chemical analysis of Fusarium pigmentation in the late 1970s and early 1980s, but this never tried to relate the compounds with the mold’s observable biological phenomena [13]. Recent sequencing of the F. graminearum genome and development gene replacement tools allowed major progress in genetic and biochemical studies [13]. Now, it is known that the red pigmentation of F. graminearum is due to the deposition of aurofusarin in the walls [2,14], but it is likely to be the combination of several pigments [15,16].
Pigmentation is part of the mold growth process and it can be used as a tool for growth studies as an alternative to the expansion in size [17]. This approach can help overcome spatial constraints in fungal studies or applications, such as the limited size or particular shape of a Petri dish or bioreactor, or even predict toxin production solely by analyzing the mold surface color. For instance, mutants with absence of the pigment aurofusarin seem to produce an increased amount of ZEA [2], and histone H3 lysine 4 methylation (H3K4me) is important for the transcription of genes for the biosynthesis of both DON and aurofusarin [18]. Thus, there is some connection between the production of major Fusarium mycotoxins and pigments.
This review aims to identify the major F. graminearum pigments described in the literature and summarize what is known so far about them so that future researchers will be able to more comprehensively relate their dynamics and the mold’s color change.
2. F. graminearum Colors throughout Its Lifecycle
It is first important to know that there is no single set of colors to describe F. graminearum throughout its lifecycle. The surface colors change depending on several variables, such as strain, maturity, nutrients, temperature, pH, water activity, light exposure, and aeration [8,10,16,17]. Ashley et al. [8] mentioned early studies identifying pH as the main determinant of Fusarium colors, “so that one and the same culture may be orange or yellow colored at an acid reaction, the color changing to red or blue when the medium becomes alkaline”. Medentsev et al. [19] said that the biosynthesis of naphtoquinones (major secondary metabolites, including pigments) is the mold’s main response to stress. F. graminearum has different types of pigments, all with distinct properties [8,15,20,21,22], from which we have to expect numerous combinations and the resulting chromatic attributes. For instance, the teleomorph was found to have violet pigmentation in its perithecia [23]. Thus, it is impractical to summarize all possibilities. For this reason, this description will simply focus on the mold grown on yeast extract agar (YEA) at 25 °C as an example (Figure 1). The isolate was obtained from the Catalogue of the Japan Collection of Microorganisms (JCM), where it is registered as the teleomorph Giberella zeae (Schwabe) Petch, and it was isolated by Sugiura [24] from rice stubble in Hirosaki, Aomori Prefecture, Japan.
YEA is a highly nutritive medium containing agar as a solidifier, peptic digest of animal tissue, and yeast extract, and is thus rich in nitrogenous compounds, vitamin B, and other nutrients [25]. Furthermore, yeast extracts do not seem to affect the quality or level of aurofusarin, a major pigment, by any Fusarium species [26]. This is important because it is desirable to use the mold’s original coloration in the studies, without changing it much, because of the nutrients available.
F. graminearum colors change in a very consistent and predictable pattern [17]. At a glance, the mold germinates as a pale mycelium and starts to acquire a yellowish coloration between its third and fourth day. It attains its full orange tone on the sixth day and then shifts to dark wine red by the 16th day. The color distribution is heterogeneous: it forms a radial gradient, with the center more intensely colored and increasingly pale surroundings. Kim et al. [1] described the F. graminearum as a “yellow to tan mycelia with the white to carmine red margins”, certainly depending on the condition in which it grows. A dominant red color tends to become more evenly distributed as the fungus ages, and alternated concentric layers of white and red rings close to the center become increasingly more evident. The white rings are hairy and seem to be formed of colorless hyphae. The medium’s color change (Figure 1c) from pale to yellow is due to the accumulation of aurofusarin [14].
Recent analysis based on red, green, and blue (RGB) channels taken for F. graminearum photographs shows that the three color components are positively correlated and all exhibit a third-degree polynomial trend when measured throughout the first 20 days of its lifecycle, and it makes the colors a potential tool to replace size-based measurements for growth studies and to predict toxin production [17].
3. Major F. graminearum Pigments
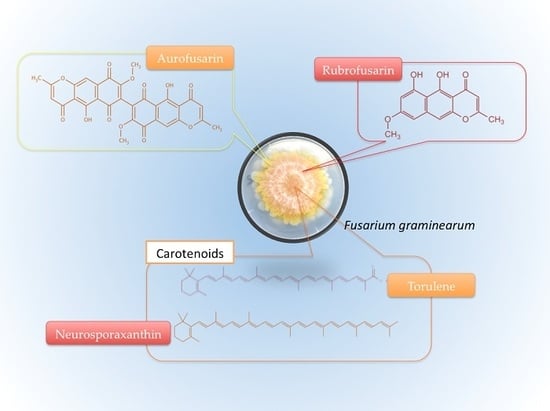
Most of what is known about the pigmentation of F. graminearum comes from studies on F. culmorum, F. aquaeductuum, F. fujikuroi, and F. oxysporum, and eventual confirmation that the pigments occur across species [15]. Such studies are aimed primarily at enhancing pigment production for the dye industry as an alternative to synthetic counterparts [27]. A pioneer study by Ashley et al. [8] identified aurofusarin, rubrofusarin, culmorin, and their derivatives among the pigments. Since then, others have been mentioned, including perithecial melanin [28] and carotenoids [15]. The most relevant carotenoids from F. graminearum are perhaps neurosporaxanthin and torulene [14,15,21].
F. graminearum pigmentation is very complex, but most pigments have similar colors, ranging from yellow and orange to red. Thus, it is perhaps difficult to know how much each pigment contributes to its color. Yet, the literature points towards aurofusarin and neurosporaxanthin, and possibly also rubrofusarin, as the ones impacting F. graminearum’s surface color the most. No source has simultaneously covered non-carotenoid and carotenoid pigments and the ones describing each of these classes showed the tendency to state the respective compounds as the main source of coloration, maybe because the pigments have similar colors. It would be a good idea to find out which contributes the most to the coloration, perhaps by experimentation based on distinctive properties of the pigments. For instance, carotenoids are expected to be reactive to light, but as far as the literature has shown, polyketides such as aurofusarin and rubrofusarin are not likely to change considerably in the presence and absence of illumination. By simple observation, aurofusarin appears to be predominant because F. graminearum specimens grown in dark and illuminated settings do not seem to present different coloration when maintained at the same temperature.
In any case, the color change has previously been demonstrated to follow a predictable trend, disregarding the pigments involved. Thus, whichever the dominant pigments, they follow a consistent pattern over time. It is still difficult to advocate if the changes are mostly due to variations in the proportion of different pigments or chemical reactions leading to changes of the same compound into its derivatives, just like the case of aurofusarin at different pH settings. It could even be simply the breakdown of aurofusarin into rubrofusarin molecules.
There are two more aspects to consider before listing F. graminearum pigments or related compounds. Culmorin is colorless, but it is included in this review because it was isolated together for the first time during studies of Fusarium pigmentation. Bikaverin and fusorubin are Fusarium pigments [19,29,30], but they are not included in the following list because there is very little evidence about their occurrence and impact on the coloration of F. graminearum.
3.1. Aurofusarin
Aurofusarin was obtained chemically and isolated from F. culmorum before Baker et al. [10] extracted and purified it from a strain of F. graminearum Schwabe. It is a dimeric metabolite belonging to the naphthoquinone group of polyketides [13,31], described as a golden yellow-orange or red micro-crystalline pigment in the form of a prism [8,13,20,32], with C30H18O12 as its molecular formula (Figure 2), 570.5 g/mol as its molecular weight, and a melting point over 360 °C [8,33]. It is now assigned as 13191-64-5 under the Chemical Abstracts Service (CAS) [33]. Aurofusarin is the only F. graminearum pigment produced under a deficiency of nitrogen, phosphorus, oxidative stress, and the inhibition of respiration [19].
Organic solvents can solve aurofusarin moderately, and it becomes yellow in acid and reddish in alkalis [12]. This is a plausible explanation for the color changes throughout F. graminearum’s lifecycle (Figure 1); perhaps the mold turns the medium increasingly alkaline and it causes aurofusarin to change its color to red. However, pH ≤ 4 seems to prevent F. graminearum from producing aurofusarin [19]. Indeed, pH is the most important regulator of aurofusarin production because most naphtoquinones tend to be cytostatic at a neutral pH level [14]. Yet, temperature and water activity (aw) also have an impact on aurofusarin biosynthesis, as they were found to be directly proportional [3].
The species F. acuminatum, F. avenaceum, F. crookwellens, F. culmorum, F. graminearum, F. poae, F. pseudograminearum, F. sambucinum, F. sporotrichioides, and F. tricinctum all produce aurofusarin [13]. Different strains of F. graminearum produce different quantities under similar conditions [34]. Besides Fusarium species, aurofusarin can also be isolated from Hypomyces rosellus and Dactylium dendroides [35]. It is negatively correlated with vegetative growth [1], thus the pigment is expected to be more abundant in differentiated structures. Aurofusarin certainly increases the organism’s competitive saprophytic ability due to its antibiotic properties, but it does not help the fungus colonizing the host crops and does not protect the fungus against radiation [2]. The pigment can be extracted using benzene-acetone (4:1) and purified by chromatography on silica gel impregnating with oxalic acid [12,20].
The aurofusarin biosynthetic pathway involves several genes in clusters and at least five enzymatic steps, with rubrofusarin as an intermediate [36]. For instance, its synthesis requires the intervention of polyketide synthase genes [37] in a 30 kb cluster (Figure 3), including PKS12, AurR1, aurJ, aurF, gip1, and gip2 [36,38] within at least 11 reading frames (FG02320.1–FG02330.1) plus a facilitator transporter gene (FG02331.1) [39]. Among the genes, AurR1 and gip2 are believed to be the cluster’s transcription factors [1,13], and the putative laccase gip1 has been described as potentially responsible for the dimerization of rubrofusarin into aurofusarin [36,40]. More recently, two more genes (gip3 and gip8) were also shown to be important for the dimerization [38]. Frandsen et al. [36] mentioned two more orphan genes (aurZ and aurS), and AurT, an aurofusarin pump, responsible for transporting aurofusarin and rubrofusarin across the plasma membrane.
Though aurofusarin was described before 1937 [8], it has only been regarded as a food and feed contaminant since start of the new millennium [41]. There is considerable literature about the issue [31,41]. It is frequently found in several commodities throughout virtually all climatic regions in the world, sometimes at concentrations as high as 2046 µg/kg to 10,200 µg/kg [31]. Beccari et al. [34] detected 10,400–140,000 µg/kg in Italian samples of durum wheat, Ezekiel et al. [42] mentioned concentrations above 800 µg/kg in chicken feed, and Nichea et al. [43] detected median concentrations of 71.4 µg/kg (2011) and 80.7 µg/kg (2014) in native grass of Argentina, intended for grazing cattle. The latter was found to co-occur with zearalenone.
Aurofusarin is bioactive and it is considered a neglected mycotoxin [31,44]. According to Medentsev et al. [32], it inhibits the growth of some molds and yeasts, and Tola et al. [45] also found it to impair the growth of Red Tilapia (Oreochromis niloticus × O. mossambicus). Moreover, it was found to be cytotoxic for colon adenocarcinoma cell line HT29 and the non-tumorigenic colon cells HCEC-1CT when at concentrations above 1 µM [31]. It has also shown toxicity for differentiated intestinal porcine epithelial cells (IPEC-J2) when combined with DON [46]. Dvorska, Surai and their colleagues [41,47,48,49] performed a series of studies demonstrating the detrimental effects of aurofusarin on Japanese quail eggs. They found aurofusarin to cause a significant decline of vitamins E and A, total carotenoid, lutein and zeaxanthin, and stimulated lipid peroxidation in the egg yolk. There are many details yet to be clarified regarding the way that aurofusarin affects these compounds. Regarding fats, the authors stated that aurofusarin exhibited an association with a “decrease in the docosahexaenoic acid proportion in the phospholipid, cholesteryl ester and free fatty acid fractions of the egg yolk” and a simultaneous increase in “the proportion of linoleic acid in the phospholipid, free fatty acid and triacylglycerol fractions”. Dvorska [41] also said that aurofusarin reduces the quality of chicken meat, though the mechanism of how it happens is presently unclear.
Regarding the relationship between pigmentation and mycotoxin contamination, there is solid yet scarce literature relating the genetic and biosynthetic origins of aurofusarin and both DON and ZEA [2,18]. As mentioned earlier, Malz et al. [2] demonstrated that mutants for the gene PKS12, unable to produce aurofusarin, produced an increased quantity of ZEA. It suggests that some inhibitory factor for ZEA is related to aurofusarin synthesis, but the mechanism is still to be identified. Furthermore, there seems to be some quantitative relationship between the three compounds, as the combined results of two studies suggest some correlation between them [3,34], though this possibility still requires further investigation. These studies are still in the preliminary stages, but they demonstrate that some genetic factors are common for aurofusarin and these mycotoxins. If such factors have a similar influence on aurofusarin and the toxins, variations in the quantity of aurofusarins might theoretically be used to predict the quantity of DON or ZEA. Thus, there is a possibility that F. graminearum’s surface color can be a good predictor of toxicity. Furthermore, aurofusarin is a toxin itself. It can surely be quantifiable through the mold’s surface color.
3.2. Rubrofusarin
Rubrofusarin (CAS: 3567-00-8) is a crystalline polyketide red-orange pigment [8,20] usually found in the form of needles [10,50]. Demicheli et al. [51] described it as a powder. Rubrofusarin belongs to the class of naphthopyrones and resembles the aurofusarin monomer [13], consisting of a mono methyl ether [8] (Figure 4). Indeed, the biosynthesis of both pigments seems connected because different levels of aeration produce distinct proportions of rubrofusarin and aurofusarin [22]. Thus, the logic behind aurofusarin as a predictor of F. graminearum toxicity mentioned above in theory is likely to be applicable to rubrofusarin.
Stout et al. [9] and Tanaka et al. [52] were the first investigators presenting its structure in 1962. According to them, rubrofusarin’s molecular formula is C15H12O5 and the melting point is 210–211 °C. The molecular weight is 272.3 g/mol [50]. Rubrofusarin is insoluble in water, but it is soluble in ethanol and dimethyl sulfoxide (DMSO) [51]. Its color does not respond to pH, unlike aurofusarin [8]. Furthermore, it has chelating properties, forming complexes with Mg2+, Al3+, Fe3+, Ni2+, and Cu2+ in a solid state and aqueous medium [53,54,55].
Several studies mention rubrofusarin or derivatives isolated from different sources, usually fungi [56], or plant roots or seed [55], for pharmacological purposes. Ashley et al. [8] published a pioneer report on the isolation of rubrofusarin from F. culmorum and F. graminearum. From the fungus Guanomyces polytrix, Mata et al. [57] isolated rubrofusarin B, a variation of the compound in which methyl ether replaces the hydroxy group at position 6 [58]. Another rubrofusarin producing mold is Aspegillus niger, also known for producing ochratoxins and consequently causing the Balkan nephropathy [59]. Regarding plants, Rangaswami [60] isolated the compound from Senna tora in India, and Oliveira et al. [61] obtained the rubrofusarin glycoside from the softwood of S. macranthera in Brazil. Senna comprises a diverse genus of native leguminous throughout the tropics [51]. Berchemia polyphylla var. leioclada, a woody deciduous plant abundant in China, produces at least three rubrofusarin glycosides [62]. Other rubrofusarin producing species are Paepalanthus bromelioides [63] and Flavoparmelia euplecta [64]. Moreover, it is reasonable to think that virtually all aurofusarin producing organisms also have the potential to produce rubrofusarin because the latter pigment is an intermediate of the aurofusarin biosynthetic pathway [36].
F. graminearum synthesizes rubrofusarin through a polyketide chain intermediate by condensing seven acetate units [65]. Deletion mutants of AurR1 and PKS12 cannot synthesize rubrofusarin, exactly as it happens to aurofusarin [13]. Furthermore, the FG12040 protein is also responsible for rubrofusarin synthesis, and the process is inhibited in mutants without the gene cch1, known as a calcium ion channel encoder [66]. Rugbjerg et al. [67] confirmed the intervention of the genes mentioned and the metabolic pathway by reconstructing it using Saccharomyces cerevisiae. They “paved the way” for industrial production of the pigment.
There is plenty of pharmaceutical potential of rubrofusarin and derivatives to be unlocked, and some of it has already been demonstrated. Rubrofusarin has antimycobacterial, antiallergic, and phytotoxic properties to herbs Amaranthus hypochondriacus and Echinochloa crus-galli [1,38,40,59]. According to Alqahtani et al. [68], rubrofusarin can enhance the cytotoxicity of paclitaxel (PTX) against the adriamycin-resistant breast cancer cell line MCF-7adr. Still regarding cancer research, rubrofusarin B was cytotoxic for the colon cancer cell line SW1116 [59], and rubrofusarin presented complete inhibitory ability against human DNA topoisomerase II-α, meaning that it can be used to develop potential anticancer and antiviral drugs [56]. Jing et al. [62] found rubrofusarin glycosides to have antioxidant properties, one of which was even stronger than vitamin C. The pigment was also found to exhibit estrogenic activity [69]. Rubrofusarin bioactivity has led Moreira et al. [55] to effectively demonstrate its fitness to be applied as a fluorescent probe.
3.3. Culmorin
Culmorin (CAS: 18374-83-9) is a natural colorless metabolite found in various Fusarium species [8,20,70,71], with C15H26O2 as its molecular structure (Figure 5), first isolated by Ashley et al. [8] and later established by Barton et al. [11]. Culmorin is defined as a longifolene sesquiterpene diol with a tricyclo-[6.3.0.0] undecane skeleton [72], and its molecular weight is 238.3 g/mol [73]. It seems to be biosynthesized from trans-farnesyl pyrophosphate [70] and compounds closely related include hydroxyculmorins, culmorone, and hydroxyculmorone [74,75,76]. Technically, it is not a pigment in the sense that it has no color, as already mentioned, but it was initially isolated during pigment studies together with aurofusarin and rubrofusarin [12], possibly because they share some chemical properties.
So far, the culmorin producing Fusarium species mentioned in the literature include F. graminearum, F. culmorum, F. crookwellense (F. cerealis), F. venenatum [79], and more recently, F. praegraminearum, a basal species of the F. graminearum complex [80]. Laraba et al. stated that 77% of F. culmorum genotypes were capable of producing culmorin in Algeria, and most were around the subtropical areas. Besides Fusarium, the marine fungi Leptosphaeria oreamaris [81] and Kallichroma tethys also synthesize culmorin [79].
The biosynthesis of culmorin requires the gene CLM1 (GenBank: GU123140.1), responsible for encoding a longiborneol synthase for the compound’s pathway [82,83]. The gene CLM2 encodes the cytochrome P450 and this is responsible for the subsequent hydroxylation of longiborneol. Culmorin was also synthesized in vitro using tetrahydroeucarvone [81]. Citric and lactic acids (5%) seem to attenuate the synthesis in feed, and this also happens to other Fusarium metabolites, including DON [84].
F. graminearum grown in durum wheat was found to produce culmorin at concentrations exceedingly high (2.5–14 g/kg) in central Italy, and there were considerable variations between strains [34]. It was also detected in Norway at median concentrations of 100 µg/kg (wheat), 292 µg/kg (barley), and 2000 µg/kg (oats) [85]. Similar results were found in an ensemble study from Austria, Denmark, and Hungary [44]. In Cameroon, Abia et al. [86] detected culmorin in cereals, nuts, and derivatives at a median concentration of 100 µg/kg. Generotti et al. [87] demonstrated that culmorin from contaminated wheat flour can endure an entire biscuit baking process, with final levels up to 92 µg/kg in baked biscuits, corresponding to a percentage of heat degradation between 25% and 80% at 180 °C. Their final products also exhibited 15-hydroxy-culmorin. Culmorin and derivatives also seem to resist the brewing process, even after the treatment of the substrates with the fungicide Prosaro® 250 [88].
Culmorin has mild [79] antifungal activity against several molds, particularly wheat and corn parasites [81]. It is also phytotoxic [72]. Regarding the fungal-plant interaction, culmorin presented a correlation with the amount of lutein in durum wheat contaminated with Fusarium [89], and this could be a way to empirically estimate the extent of contamination. Furthermore, it is frequently detected with DON [70,76,87], usually in a quantity three-fold higher, though some variables can influence this ratio [72]. However, since culmorin is colorless, theoretically, it cannot be directly measured through F. graminearum surface color. Culmorin and other F. graminearum secondary metabolites seem to enhance DON toxicity in caterpillars [90] and pigs [76,91]. Yet, culmorin itself is weakly toxic and it is negative to the Ames test of mutagenicity [79].
3.4. Black Perithecial Pigment
Fusarium is among the deuteromycota now known for having a teleomorphic ascomycota called Gibberella, and as such it develops a reproductive fruiting body called perithecium. The black perithecial pigment, sometimes described as dark blue, violet, or purple [28,92], is almost restricted to the fruiting body [93], though the producing gene is even present in Fusarium species with an unknown teleomorph [23]. The pigment’s nature is still under study and only recently has been related to fusorubins [94] and described as ″5-deoxybostrycoidin-based melanin″ [28], with C15H11NO4 as the molecular formula [95] (Figure 6).
Several Fusarium species, including F. graminearum, F. verticillioides, and F. fujikuroi [28,94], produce the blackish perithecial pigments, and appear to be related to an ancestral highly conserved gene cluster [96]. The fungus Nectria haematococca also showed the ability to produce 5-deoxybostrycoidin [95]. In F. graminearum, the six-gene PGL cluster, particularly the gene PGL1 or PKS3, seems to be associated with the production of the polyketide synthase responsible for biosynthesis of the black perithecial pigment [37,97]. PGL1 is related to a transcription-associated protein (TAP) cluster called TC3 [93]. Studt et al. [94] demonstrated an association between the dark pigments and fusarubin under the intervention of the so-called fsr gene cluster.
3.5. Carotenoids
Natural carotenoids comprise a family of more than 750 natural lipophilic terpenoids, several of which are produced by fungi, though they are not essential for these organisms [15]. Carotenoids are common in molds and they contribute to the yellow, orange, and reddish coloration [99]. They have been produced industrially and are widely used as food and feed additives [100].
An important characteristic of carotenoids worth mentioning is their sensitivity to light. As Avalos et al. [15] stated, illumination induces the synthesis of carotenoids through the transcriptional induction of structural genes in Fusarium. Jin et al. [21] identified neurosporaxanthin and torulene as the most relevant, but other carotenoids that likely play a minor role in F. graminearum’s color pattern are torularhodin, β-carotene, γ-carotene, ζ-carotene, and β-zeacarotene [101,102,103].
3.5.1. Torulene
Torulene (CAS: 547-23-9) is a natural carotenoid of importance for industrial application [104,105], with C40H54 as its molecular formula and 534.9 g/mol as its molecular weight [105]. As Figure 7 shows, it has 13 double bonds, a β-ionone, and a long polyene chain [104]. In petroleum ether solution, torulene has a pinkish-red color, depending on the concentration [103].
Neurospora crassa is the most well-known producer of torulene, but it is also synthesized by F. fujikuroi [107]; F. graminearum [21]; the red yeasts Sporidiobolus pararoseus and Rhodotorula glutinis [108] and related organisms; and the genera Cystofilobasidium, Dioszegia, Rhodosporium, and Sporobolomyces [103]. These organisms probably produce it as protection against photo-oxidation and free radicals [109].
Geranyl-geranyl diphosphate is the precursor of torulene, and it is transformed under the intervention of two enzymes called AL-2 and AL-1, produced by eponymous genes [107]. The biosynthesis is mostly influenced by nutrients, especially the sources of carbon and nitrogen, but it also responds to other factors such as aeration, temperature, acidity, exposure to radiation, and the presence of chemicals such as alcohols [103].
There is very little information on torulene’s bioactivity and nutritional value, perhaps because it is rare in food [110], but its nature, structure, and sparse evidence provide some hints. Animal studies demonstrated its safety to be used as a food additive [104], and Kot et al. [103] added that it can also be used as feedstock and a cosmetic additive. The presence of a non-substituted β-ionone ring makes torulene pro-vitamin A, which is likely to present higher antioxidant or free radical scavenging activity than β-carotene, higher reactivity in aqueous solutions, more efficient electron transfer-reactions than lycopene [108], and there is evidence of anti-prostate cancer activity [104].
3.5.2. Neurosporaxanthin
Neurosporaxanthin (CAS: 2468-88-4), or β-apo-4′-carotenoic acid, is carboxilic apocarotenoid xanthophyll [99,111], with C35H46O2 as its molecular formula (Figure 8) and 498.8 g/mol as its molecular weight [107,112]. Neurosporaxanthin carries the name of Neurospora crassa, from where it was originally isolated, but Fusarium, Verticillium, and Podospora species also synthesize it [99,111].
Prado-Cabrero et al. [114] described neurosporaxanthin as a “cleavage product of torulene”. The biosynthesis initiates from the condensation of two geranylgeranyl pyrophosphates into phytoene and following desaturations [99,100]. The cleavage of torulene into neurosporaxanthin requires the carotenoid oxygenase CAO-2, first resulting in β-apo-4’-carotenal, the aldehyde of neurosporaxanthin [107]. The major genes involved in the process are carRA, carB, carT, and carD [99]. The latter gene is in a different cluster.
4. Conclusion
In summary, it is reasonable to assume the possibility of using F. graminearum’s surface color to estimate how much toxin the mold is producing and to possibly estimate its potential bioactivity, but there is still a lot to be investigated in relation to F. graminearum and several other molds. It is necessary to catalog the pigments produced by each species and clarify their biosynthetic relationships to prevent information “gaps”. For instance, a trichothecene-producing mold is perhaps likely to also produce culmorin, and there are also parallelisms between aurofusarin and rubrofusarin or torulene and neurosporaxanthin. It is true that commercial demand is frequently a major driving force for research and sometimes it is not merely choice or curiosity leading investigators, but it is important to always try to build a very cohesive body of knowledge from which other researchers can craft their own contributions.
Funding
The Japanese Ministry of Education, Culture, Sports, Science and Technology (MEXT) funded this study.
Acknowledgments
Umi Ogawa, for helping to take the photos of the fungi.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, J.-E.; Jin, J.; Kim, H.; Kim, J.-C.; Yun, S.-H.; Lee, Y.-W. Gip2, a putative transcription factor that regulates the aurofusarin biosynthetic gene cluster in gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Malz, S.; Grell, M.N.; Thrane, C.; Maier, F.J.; Rosager, P.; Felk, A.; Albertsen, K.S.; Salomon, S.; Bohn, L.; Schäfer, W. Identification of a gene cluster responsible for the biosynthesis of aurofusarin in the fusarium graminearum species complex. Fungal Genet. Biol. 2005, 42, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cela, E.; Kiaitsi, E.; Medina, A.; Sulyok, M.; Krska, R.; Magan, N. Interacting environmental stress factors affects targeted metabolomic profiles in stored natural wheat and that inoculated with F. Graminearum. Toxins 2018, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Weidenbörner, M. Encyclopedia of Food Mycotoxins, 1st ed.; Springer-Verlag: Berlin Heidelberg: Berlin, Germany, 2001; p. 294. [Google Scholar]
- Yoshizawa, T. Thirty-five years of research on deoxynivalenol, a trichothecene mycotoxin: With special reference to its discovery and co-occurrence with nivalenol in Japan. Food Saf. 2013, 1, 2013002. [Google Scholar] [CrossRef]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysoe, E.; Klemsdal, S.S.; Bone, K.R.; Frandsen, R.J.; Johansen, T.; Thrane, U.; Giese, H. The PKS4 gene of Fusarium graminearum is essential for zearalenone production. Appl. Environ. Microbiol. 2006, 72, 3924–3932. [Google Scholar] [CrossRef] [PubMed]
- Ashley, J.N.; Hobbs, B.C.; Raistrick, H. Studies in the biochemistry of micro-organisms: The crystalline colouring matters of Fusarium culmorum (W.G. Smith) Sacc. and related forms. Biochem. J. 1937, 31, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Stout, G.H.; Jensen, L.H. Rubrofusarin: A structure determination using direct phase calculation. Acta Crystallogr. 1962, 15, 451–457. [Google Scholar] [CrossRef]
- Baker, P.M.; Roberts, J.C. Studies in mycological chemistry. Part XXI. The structure of aurofusarin, a metabolite of some fusarium species. J. Chem. Soc. C: Org. 1966, 2234–2237. [Google Scholar] [CrossRef]
- Barton, D.H.R.; Werstiuk, N.H. The constitution and stereochemistry of culmorin. Chem. Commun. (London) 1967, 1, 30–31. [Google Scholar] [CrossRef]
- Shibata, S.; Morishita, E.; Takeda, T.; Sakata, K. Metabolic products of fungi. XXVIII. The structure of aurofusarin. (1). Chem. Pharm. Bull. (Tokyo) 1968, 16, 405–410. [Google Scholar] [CrossRef]
- Frandsen, R.J.N.; Nielsen, N.J.; Maolanon, N.; Sørensen, J.C.; Olsson, S.; Nielsen, J.; Giese, H. The biosynthetic pathway for aurofusarin in fusarium graminearum reveals a close link between the naphthoquinones and naphthopyrones. Mol. Microbiol. 2006, 61, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Dufosse, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Pardo-Medina, J.; Parra-Rivero, O.; Ruger-Herreros, M.; Rodríguez-Ortiz, R.; Hornero-Méndez, D.; Limón, M.C. Carotenoid biosynthesis in Fusarium. J. Fungi (Basel) 2017, 3, 39. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Son, H.; Lee, Y.W. Effects of light on secondary metabolism and fungal development of Fusarium graminearum. J. Appl. Microbiol. 2014, 116, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Cambaza, E.; Koseki, S.; Kawamura, S. The use of colors as an alternative to size in Fusarium graminearum growth studies. Foods 2018, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, N.; Yin, Y.; Chen, Y.; Jiang, J.; Ma, Z. Histone H3K4 methylation regulates hyphal growth, secondary metabolism and multiple stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 4615–4630. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Arinbasarova, A.; Akimenko, V.K. Biosynthesis of naphthoquinone pigments by fungi of the genus fusarium. Appl. Biochem. Microbiol. 2005, 41, 503–507. [Google Scholar] [CrossRef]
- Shibata, S.; Morishita, E.; Takeda, T.; Sakata, K. The structure of Aurofusarin. Tetrahedron Lett. 1966, 7, 4855–4860. [Google Scholar] [CrossRef]
- Jin, J.-M.; Lee, J.; Lee, Y.-W. Characterization of carotenoid biosynthetic genes in the ascomycete gibberella zeae. FEMS Microbiol. Lett. 2009, 302, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Leeper, F.J.; Staunton, J. The biosynthesis of rubrofusarin, a polyketide naphthopyrone from Fusarium culmorum: 13C nmr assignments and incorporation of 13C-and 2H-labelled acetates. J. Chem. Soc., Perkin Trans. 1 1984, 2919–2925. [Google Scholar] [CrossRef]
- Proctor, R.H.; Butchko, R.A.; Brown, D.W.; Moretti, A. Functional characterization, sequence comparisons and distribution of a polyketide synthase gene required for perithecial pigmentation in some fusarium species. Food Addit. Contam. 2007, 24, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y. Gibberella zeae (schwabe) petch. In JCM Catalogue; Japan Collection of Microorganisms, Ed.; Microbe Division (JCM): Tsukuba, Japan, 1996. [Google Scholar]
- Sigma-Aldrich. 01497 yeast extract agar. Available online: https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma-Aldrich/Datasheet/1/01497dat.pdf (4 September 2018).
- Sorensen, J.L.; Sondergaard, T.E. The effects of different yeast extracts on secondary metabolite production in fusarium. Int. J. Food Microbiol. 2014, 170, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Dufossé, L.; Caro, Y.; Fouillaud, M. Fungal pigments: Deep into the rainbow of colorful fungi. J. Fungi 2017, 3, 45. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, R.J.N.; Rasmussen, S.A.; Knudsen, P.B.; Uhlig, S.; Petersen, D.; Lysøe, E.; Gotfredsen, C.H.; Giese, H.; Larsen, T.O. Black perithecial pigmentation in fusarium species is due to the accumulation of 5-deoxybostrycoidin-based melanin. Sci. Rep. 2016, 6, 26206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, N.N.; Ammar, M.S. New antibiotic pigments related to fusarubin from fusarium solani (Mart.) Sacc. II. Structure elucidations. J. Antibiot. (Tokyo) 1979, 32, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.S.; Gerber, N.N.; McDaniel, L.E. New antibiotic pigments related to fusarubin from fusarium solani (Mart.) Sacc. I. Fermentation, isolation, and antimicrobial activities. J. Antibiot. (Tokyo) 1979, 32, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Kotik, A.N.; Trufanova, V.A.; Akimenko, V.K. Identification of aurofusarin in fusarium graminearum isolates, causing a syndrome of worsening of egg quality in chickens. Appl. Biochem. Microbiol. 1993, 29, 542–546. [Google Scholar]
- Glentham Life Sciences. Ga7883-aurofusarin. In Product Datasheet; Glentham Life Sciences, Ed.; Glentham Life Sciences: Wiltshire, United Kingdom, 2018. [Google Scholar]
- Beccari, G.; Colasante, V.; Tini, F.; Senatore, M.T.; Prodi, A.; Sulyok, M.; Covarelli, L. Causal agents of Fusarium head blight of durum wheat (Triticum durum Desf.) in central Italy and their in vitro biosynthesis of secondary metabolites. Food Microbiol. 2018, 70, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Birchall, G.R.; Bowden, K.; Weiss, U.; Whalley, W.B. The chemistry of fungi. Part LVI. Aurofusarin. J. Chem. Soc. C: Org. 1966, 2237–2239. [Google Scholar] [CrossRef]
- Frandsen, R.J.; Schutt, C.; Lund, B.W.; Staerk, D.; Nielsen, J.; Olsson, S.; Giese, H. Two novel classes of enzymes are required for the biosynthesis of aurofusarin in fusarium graminearum. J. Biol. Chem. 2011, 286, 10419–10428. [Google Scholar] [CrossRef] [PubMed]
- Gaffoor, I.; Brown, D.W.; Plattner, R.; Proctor, R.H.; Qi, W.; Trail, F. Functional analysis of the polyketide synthase genes in the filamentous fungus gibberella zeae (anamorph Fusarium graminearum). Eukaryot. Cell 2005, 4, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Kim, J.-C.; Jin, J.-M.; Yun, S.-H.; Lee, Y.-W. Functional characterization of genes located at the aurofusarin biosynthesis gene cluster in gibberella zeae. Plant Pathol. J. 2008, 24, 8–16. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Han, K.-H.; Jin, J.; Kim, H.; Kim, J.-C.; Yun, S.-H.; Lee, Y.-W. Putative polyketide synthase and laccase genes for biosynthesis of aurofusarin in Gibberella zeae. Appl. Environ. Microbiol. 2005, 71, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H. Effect of the mycotoxin aurofusarin on the antioxidant composition and fatty acid profile of quail eggs. Br. Poult. Sci. 2001, 42, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Ezekiel, C.N.; Bandyopadhyay, R.; Sulyok, M.; Warth, B.; Krska, R. Fungal and bacterial metabolites in commercial poultry feed from nigeria. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Nichea, M.J.; Palacios, S.A.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Chulze, S.N.; Torres, A.M.; Ramirez, M.L. Presence of multiple mycotoxins and other fungal metabolites in native grasses from a wetland ecosystem in argentina intended for grazing cattle. Toxins 2015, 7, 3309–3329. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Tola, S.; Bureau, D.; Hooft, J.; Beamish, F.; Sulyok, M.; Krska, R.; Encarnação, P.; Petkam, R. Effects of wheat naturally contaminated with fusarium mycotoxins on growth performance and selected health indices of red tilapia (Oreochromis niloticus × O. mossambicus). Toxins 2015, 7, 1929–1944. [Google Scholar] [CrossRef] [PubMed]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of fusarium-derived metabolites on the barrier integrity of differentiated intestinal porcine epithelial cells (IPEC-J2). Toxins 2016, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Dvorska, J.E.; Surai, P.F. Yeast glucomannans prevent deterioration of quail egg quality during aurofusarinotoxicosis. In Proceedings of the XVI European Symposium on the Quality of Poultry Meat and the X European Symposium on the Quality of Eggs and Egg Products, Sint-Brieuc-Ploufragan, France, 23–26 September 2003; pp. 93–101. [Google Scholar]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H. Antioxidant systems of the developing quail embryo are compromised by mycotoxin aurofusarin. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 131, 197–205. [Google Scholar] [CrossRef]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H. Protective effect of modified glucomannans against aurofusarin-induced changes in quail egg and embryo. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 135C, 337–343. [Google Scholar] [CrossRef]
- BioViotica. Rubrofusarin. Available online: https://adipogen.com/productmanagement/resource/download/type/sheet/id/8299 (acessed on 10 August 2018).
- Marazzi, B.; Endress, P.K.; De Queiroz, L.P.; Conti, E. Phylogenetic relationships within Senna (Leguminosae, Cassiinae) based on three chloroplast DNA regions: Patterns in the evolution of floral symmetry and extrafloral nectaries. Am. J. Bot. 2006, 93, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Tamura, T. The chemical constitution of rubrofusarin, a pigment from fusarium graminearum. Agric. Biol. Chem. 1962, 26, 767–770. [Google Scholar] [CrossRef]
- Demicheli, C.; Bcraldo, H.; Tosi, L. Cu (II) and Ni (II) complexes of rubrofusarin and 6-galactosyl rubrofusarin. J. Braz. Chem. Soc. 1992, 52–54. [Google Scholar] [CrossRef]
- Pereira, E.; Demicheli, C.; Peixoto, L. A spectrometric study of the chelating properties of 6-galactosyl-rubrofusarin: Mg (II), Al (III), Fe (III), Ni (II) and Cu(II). J. Braz. Chem. Soc. 1995, 6, 381–386. [Google Scholar] [CrossRef]
- Moreira, L.M.; Lyon, J.P.; Lima, A.; Codognoto, L.; da Hora Machado, A.E.; de S. Tiago, F.; Araújo, D.M.S.; Silva, E.L.; Hioka, N.; Rodrigues, M.R.; et al. Quinquangulin and rubrofusarin: A. spectroscopy study. Orbital: Electron. J. Chem. 2017, 9. [Google Scholar] [CrossRef]
- Branco, A.; Pinto, A.C.; Braz-Filho, R.; Silva, E.F.; Grynberg, N.F.; Echevarria, A. Rubrofusarin, a natural polyketide as new human topoisomerase II-α inhibitor. Rev. Bras. Farmacogn 2008, 18, 703–708. [Google Scholar] [CrossRef]
- Mata, R.; Gamboa, A.; Macias, M.; Santillan, S.; Ulloa, M.; Gonzalez Mdel, C. Effect of selected phytotoxins from guanomyces polythrix on the calmodulin-dependent activity of the enzymes camp phosphodiesterase and NAD-kinase. J. Agric. Food Chem. 2003, 51, 4559–4562. [Google Scholar] [CrossRef] [PubMed]
- European Molecular Biology Laboratory. Rubrofusarin B (CHEBI:133805). Available online: https://www.ebi.ac.uk/chebi/searchId.do?chebiId=CHEBI:133805 (accessed on 4 September 2018).
- Song, Y.C.; Li, H.; Ye, Y.H.; Shan, C.Y.; Yang, Y.M.; Tan, R.X. Endophytic naphthopyrone metabolites are co-inhibitors of xanthine oxidase, SW1116 cell and some microbial growths. FEMS Microbiol. Lett. 2004, 241, 67–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangaswami, S. Crystalline chemical components of the seeds of Cassia Tora Linn. Identity of Tora Substance C with rubrofusarin and Tora Substance B with nor-rubrofusarin. Proc. Indian Acad. Sci. Sect. A 1963, 57, 88–93. [Google Scholar]
- Oliveira, A.; Fernandes, M.; Shaat, V.; Vasconcelos, L.; Gottlieb, O. Constituents of cassia species. Rev. Latinoamer. Quim. 1977, 8, 82–85. [Google Scholar]
- Jing, Y.; Yang, J.; Wu, L.; Zhang, Z.; Fang, L. Rubrofusarin glucosides of Berchemia Polyphylla var. Leioclada and their scavenging activities for DPPH radical. Zhongguo Zhong Yao Za Zhi 2011, 36, 2084–2087. [Google Scholar] [PubMed]
- Coelho, R.G.; Vilegas, W.; Devienne, K.F.; Raddi, M.S.G. A new cytotoxic naphthopyrone dimer from paepalanthus bromelioides. Fitoterapia 2000, 71, 497–500. [Google Scholar] [CrossRef]
- Ernst-Russell, M.A.; Chai, C.L.L.; Wardlaw, J.H.; Elix, J.A. Euplectin and coneuplectin, new naphthopyrones from the lichen flavoparmelia euplecta. J. Nat. Prod. 2000, 63, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Mock, B.H.; Robbers, J.E. Biosynthesis of rubrofusarin by Fusarium graminearum. J. Pharm. Sci. 1969, 58, 1560–1562. [Google Scholar] [CrossRef] [PubMed]
- Hallen, H.E.; Trail, F. The L-type calcium ion channel cch1 affects ascospore discharge and mycelial growth in the filamentous fungus gibberella zeae (anamorph Fusarium graminearum). Eukaryot. Cell 2008, 7, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Rugbjerg, P.; Naesby, M.; Mortensen, U.H.; Frandsen, R.J. Reconstruction of the biosynthetic pathway for the core fungal polyketide scaffold rubrofusarin in Saccharomyces cerevisiae. Microb. Cell Fact. 2013, 12, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alqahtani, S.D.; Assiri, H.A.; Al-Abbasi, F.A.; El-Halawany, A.M.; Al-Abd, A.M. Abstract 1205: Rubrofusarin and toralactone sensitize resistant mcf-7adr cell line to paclitaxel via inhibiting p-glycoprotein efflux activity. Cancer Res. 2017, 77, 1205. [Google Scholar] [CrossRef]
- El-Halawany, A.M.; Chung, M.H.; Nakamura, N.; Ma, C.M.; Nishihara, T.; Hattori, M. Estrogenic and anti-estrogenic activities of cassia tora phenolic constituents. Chem. Pharm. Bull. (Tokyo) 2007, 55, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Ghebremeskel, M.; Langseth, W. The occurrence of culmorin and hydroxy-culmorins in cereals. Mycopathologia 2001, 152, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Young, J.C.; Games, D.E. Supercritical fluid chromatography of fusarium mycotoxins. J. Chromatogr. 1992, 627, 247–254. [Google Scholar] [CrossRef]
- Weber, J.; Vaclavikova, M.; Wiesenberger, G.; Haider, M.; Hametner, C.; Frohlich, J.; Berthiller, F.; Adam, G.; Mikula, H.; Fruhmann, P. Chemical synthesis of culmorin metabolites and their biologic role in culmorin and acetyl-culmorin treated wheat cells. Org. Biomol. Chem. 2018, 16, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. Culmorin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Culmorin#section=Top (accessed on 5 September 2018).
- Miller, J.D.; MacKenzie, S. Secondary Metabolites of Fusarium venenatum Strains with Deletions in The Tri5 Gene Encoding Trichodiene Synthetase. Mycologia 2000, 92, 764–771. [Google Scholar] [CrossRef]
- Kasitu, G.C.; ApSimon, J.W.; Blackwell, B.A.; Fielder, D.A.; Greenhalgh, R.; Miller, J.D. Isolation and characterization of culmorin derivatives produced by Fusariumculmorum CMI 14764. Can. J. Chem. 1992, 70, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Langseth, W.; Ghebremeskel, M.; Kosiak, B.; Kolsaker, P.; Miller, D. Production of culmorin compounds and other secondary metabolites by Fusarium culmorum and F. Graminearum strains isolated from Norwegian cereals. Mycopathologia 2001, 152, 23–34. [Google Scholar] [CrossRef] [PubMed]
- iChemLabs. 2D Sketcher. Chemdoodle Web Components. Available online: https://web.chemdoodle.com/demos/sketcher/ (accessed on 9 September 2018).
- Nara Institute of Science and Technology. Knapsack metabolite information—culmorin. Available online: http://kanaya.naist.jp/knapsack_jsp/information.jsp?word=C00021971 (accessed on 2 September 2018).
- Pedersen, P.B.; Miller, J.D. The fungal metabolite culmorin and related compounds. Nat. Toxins 1999, 7, 305–309. [Google Scholar] [CrossRef]
- Grafenhan, T.; Johnston, P.R.; Vaughan, M.M.; McCormick, S.P.; Proctor, R.H.; Busman, M.; Ward, T.J.; O’Donnell, K. Fusarium praegraminearum sp. Nov., a novel nivalenol mycotoxin-producing pathogen from New Zealand can induce head blight on wheat. Mycologia 2016, 108, 1229–1239. [Google Scholar] [PubMed]
- Strongman, D.; Miller, J.; Calhoun, L.; Findlay, J.; Whitney, N. The biochemical basis for interference competition among some lignicolous marine fungi. Botanica Marina 1987, 30, 21–26. [Google Scholar] [CrossRef]
- McCormick, S.P.; Alexander, N.J.; Harris, L.J. CLM1 of Fusarium graminearum encodes a longiborneol synthase required for culmorin production. Appl. Environ. Microbiol. 2010, 76, 136–141. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology, I. Gibberella zeae strain GZ3639 longiborneol synthase (CLM1) gene, complete cds. Available online: https://www.ncbi.nlm.nih.gov/nuccore/GU123140 (accessed on 7 September 2018).
- Humer, E.; Lucke, A.; Harder, H.; Metzler-Zebeli, B.U.; Bohm, J.; Zebeli, Q. Effects of citric and lactic acid on the reduction of deoxynivalenol and its derivatives in feeds. Toxins 2016, 8, 285. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Eriksen, G.S.; Hofgaard, I.S.; Krska, R.; Beltran, E.; Sulyok, M. Faces of a changing climate: Semi-quantitative multi-mycotoxin analysis of grain grown in exceptional climatic conditions in Norway. Toxins 2013, 5, 1682–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.N.; Njobeh, P.B.; Dutton, M.F.; Moundipa, P.F. Determination of multi-mycotoxin occurrence in cereals, nuts and their products in cameroon by liquid chromatography tandem mass spectrometry (LC-MS/MS). Food Control 2013, 31, 438–453. [Google Scholar] [CrossRef]
- Generotti, S.; Cirlini, M.; Sarkanj, B.; Sulyok, M.; Berthiller, F.; Dall’Asta, C.; Suman, M. Formulation and processing factors affecting trichothecene mycotoxins within industrial biscuit-making. Food Chem. 2017, 229, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Mastanjevic, K.; Sarkanj, B.; Krska, R.; Sulyok, M.; Warth, B.; Mastanjevic, K.; Santek, B.; Krstanovic, V. From malt to wheat beer: A comprehensive multi-toxin screening, transfer assessment and its influence on basic fermentation parameters. Food Chem. 2018, 254, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.M.; Sulyok, M.; Jirsa, O.; Spitzer, T.; Krska, R.; Polisenska, I. Relationship between lutein and mycotoxin content in durum wheat. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2014, 31, 1274–1283. [Google Scholar] [CrossRef] [PubMed]
- Dowd, P.F.; Miller, J.D.; Greenhalgh, R. Toxicity and interactions of some Fusarium graminearum metabolites to caterpillars. Mycologia 1989, 81, 646–650. [Google Scholar] [CrossRef]
- Rotter, R.; Trenholm, H.; Prelusky, D.; Hartin, K.; Thompson, B.; Miller, J. A preliminary examination of potential interactions between deoxynivalenol (DON) and other selected Fusarium metabolites in growing pigs. Can. J. Anim. Sci. 1992, 72, 107–116. [Google Scholar] [CrossRef]
- Trail, F.; Common, R. Perithecial development by gibberella zeae: A light microscopy study. Mycologia 2000, 130–138. [Google Scholar] [CrossRef]
- Lawler, K.; Hammond-Kosack, K.; Brazma, A.; Coulson, R.M. Genomic clustering and co-regulation of transcriptional networks in the pathogenic fungus Fusarium graminearum. BMC Syst. Biol. 2013, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Studt, L.; Wiemann, P.; Kleigrewe, K.; Humpf, H.-U.; Tudzynski, B. Biosynthesis of fusarubins accounts for pigmentation of Fusarium fujikuroi perithecia. Appl. Environ. Microbiol. 2012, 78, 4468–4480. [Google Scholar] [CrossRef] [PubMed]
- Parisot, D.; Devys, M.; Barbier, M. Notizen: 5-Deoxybostrycoidin, a New Metabolite Produced by the Fungus Nectria haematococca (Berk. and Br.) Wr. Zeitschrift für Naturforschung B 1989, 44, 1473–1474. [Google Scholar] [CrossRef]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.; Lysoe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Dadachova, E.; Bryan, R.A.; Howell, R.C.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. The radioprotective properties of fungal melanin are a function of its chemical composition, stable radical presence and spatial arrangement. Pigm. Cell Melanoma Res. 2008, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Estrada, A.F. Regulation by light in fusarium. Fungal Genet. Biol. 2010, 47, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Prado-Cabrero, A.; Schaub, P.; Diaz-Sanchez, V.; Estrada, A.F.; Al-Babili, S.; Avalos, J. Deviation of the neurosporaxanthin pathway towards beta-carotene biosynthesis in fusarium fujikuroi by a point mutation in the phytoene desaturase gene. FEBS J. 2009, 276, 4582–4597. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Cerdà-Olmedo, E. Carotenoid mutants of gibberella fujikuroi. Curr. Genet. 1987, 11, 505–511. [Google Scholar] [CrossRef]
- Diaz-Sanchez, V.; Estrada, A.F.; Trautmann, D.; Al-Babili, S.; Avalos, J. The gene card encodes the aldehyde dehydrogenase responsible for neurosporaxanthin biosynthesis in fusarium fujikuroi. FEBS J. 2011, 278, 3164–3176. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “New” fungal carotenoids for industry? Microb. Cell Fact. 2018, 17, 49. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, N.; Li, B.; Xu, Q.; Song, J.; Wei, N.; Wang, W.; Zou, H. Increased torulene accumulation in red yeast sporidiobolus pararoseus ngr as stress response to high salt conditions. Food Chem. 2017, 237, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology, I. Torulene. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281253 (accessed on 12 September 2018).
- Royal Society of Chemistry. Torulene. Available online: http://www.chemspider.com/Chemical-Structure.4444665.html?rid=28fe93b0-090b-48cf-ba81-28141d575d21 (accessed on 2 September 2018).
- Saelices, L.; Youssar, L.; Holdermann, I.; Al-Babili, S.; Avalos, J. Identification of the gene responsible for torulene cleavage in the neurospora carotenoid pathway. Mol. Genet. Genomics 2007, 278, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wang, H.; Du, C.; Zhang, W.; Qian, H. Tentative identification of torulene cis/trans geometrical isomers isolated from sporidiobolus pararoseus by high-performance liquid chromatography-diode array detection-mass spectrometry and preparation by column chromatography. Anal. Sci. 2013, 29, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Zoz, L.; Carvalho, J.C.; Soccol, V.T.; Casagrande, T.C.; Cardoso, L. Torularhodin and torulene: Bioproduction, properties and prospective applications in food and cosmetics—A review. Braz. Arch. Biol. Technol. 2015, 58, 278–288. [Google Scholar] [CrossRef]
- Maldonade, I.R.; Rodriguez-Amaya, D.B.; Scamparini, A.R.P. Carotenoids of yeasts isolated from the brazilian ecosystem. Food Chem. 2008, 107, 145–150. [Google Scholar] [CrossRef]
- Martín, J.-F.; García-Estrada, C.; Zeilinger, S. Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites, 1st ed.; Springer-Verlag New York: New York, NY, USA, 2014. [Google Scholar]
- National Center for Biotechnology, I. Neurosporaxanthin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/637039 (accessed on 11 September 2018).
- Royal Society of Chemistry. Neurosporaxanthin. Available online: http://www.chemspider.com/Chemical-Structure.552701.html (accessed on 2 September 2018).
- Prado-Cabrero, A.; Scherzinger, D.; Avalos, J.; Al-Babili, S. Retinal biosynthesis in fungi: Characterization of the carotenoid oxygenase Carx from Fusarium fujikuroi. Eukaryot. Cell 2007, 6, 650–657. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
F. graminearum surface colors on its (a) 3rd day, (b) 6th day, and (c) 16th day.

Figure 2.
Structure of aurofusarin. Source: Glentham Life Sciences [33].
Figure 2.
Structure of aurofusarin. Source: Glentham Life Sciences [33].

Figure 3.
Biosynthetic gene cluster for aurofusarin in F. graminearum. Based on Hoffmeister and Keller [39].
Figure 3.
Biosynthetic gene cluster for aurofusarin in F. graminearum. Based on Hoffmeister and Keller [39].

Figure 4.
Structure of rubrofusarin. Source: BioViotica [50].
Figure 4.
Structure of rubrofusarin. Source: BioViotica [50].

Figure 5.
Structure of culmorin. Made on ChemDoodle [77], based on Nara Institute of Science and Technology [78].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 7.
Structure of torulene. Source: Royal Society of Chemistry [106].
Figure 7.
Structure of torulene. Source: Royal Society of Chemistry [106].

Figure 8.
Structure of neurosporaxanthin. Source: Royal Society of Chemistry [113].
Figure 8.
Structure of neurosporaxanthin. Source: Royal Society of Chemistry [113].

© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cambaza, E. Comprehensive Description of Fusarium graminearum Pigments and Related Compounds. Foods 2018, 7, 165. https://doi.org/10.3390/foods7100165
AMA Style
Cambaza E. Comprehensive Description of Fusarium graminearum Pigments and Related Compounds. Foods. 2018; 7(10):165. https://doi.org/10.3390/foods7100165
Chicago/Turabian StyleCambaza, Edgar. 2018. "Comprehensive Description of Fusarium graminearum Pigments and Related Compounds" Foods 7, no. 10: 165. https://doi.org/10.3390/foods7100165
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.