
Genistein Alleviates Intestinal Oxidative Stress by Activating the Nrf2 Signaling Pathway in IPEC-J2 Cells
by
 ,
,
Yanpin Li
1,†, 
Long Cai
1,†,
Qingyue Bi
1,2,
Wenjuan Sun
1,
Yu Pi
1,* ,
,
Xianren Jiang
1 and
Xilong Li
1,* 1
Key Laboratory of Feed Biotechnology of Ministry of Agriculture and Rural Affairs, Institute of Feed Research, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2
College of Agriculture, Yanbian University, Yanji 133000, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Vet. Sci. 2024, 11(4), 154; https://doi.org/10.3390/vetsci11040154
Submission received: 3 February 2024
/
Revised: 22 March 2024
/
Accepted: 28 March 2024
/
Published: 29 March 2024
(This article belongs to the Special Issue The Regulatory Mechanism of Nutrient Metabolism on Porcine Intestinal Health)
Abstract
:Simple Summary
Interest in using natural feed additives in animal diets to improve health and productivity has grown significantly over the past few decades. Genistein is an aglycone form of soybean isoflavones with higher antioxidant activity. However, whether genistein can alleviate oxidative stress in pig intestines and the precise mechanism behind this effect remains to be elucidated. In the present research, the hydrogen peroxide-stimulated IPEC-J2 cells oxidative stress model was employed to explore the antioxidant capacity of genistein and potential mechanisms. The results showed that genistein could exert a protective effect against hydrogen peroxide-stimulated oxidative stress by activating the Nrf2 signaling pathway in IPEC-J2 cells. These results could provide a novel nutritional intervention strategy to enhance the intestinal health of piglets under oxidative stress.
Abstract
In the weaning period, piglets often face oxidative stress, which will cause increased diarrhea and mortality. Genistein, a flavonoid, which is extracted from leguminous plants, possesses anti-inflammatory and antioxidative bioactivities. However, little is known about whether genistein could attenuate the oxidative stress that occurs in porcine intestinal epithelial cells (IPEC-J2). Herein, this experiment was carried out to investigate the protective effects of genistein in the IPEC-J2 cells oxidative stress model. Our results disclosed that H2O2 stimulation brought about a significant diminution in catalase (CAT) activity and cell viability, as well as an increase in the levels of reactive oxygen species (ROS) in IPEC-J2 cells (p < 0.05), whereas pretreating cells with genistein before H2O2 exposure helped to alleviate the reduction in CAT activity and cell viability (p < 0.05) and the raise in the levels of ROS (p = 0.061) caused by H2O2. Furthermore, H2O2 stimulation of IPEC-J2 cells remarkably suppressed gene level Nrf2 and CAT expression, in addition to protein level Nrf2 expression, but pretreating cells with genistein reversed this change (p < 0.05). Moreover, genistein pretreatment prevented the downregulation of occludin expression at the gene and protein level, and ZO-1 expression at gene level (p < 0.05). In summary, our findings indicate that genistein possesses an antioxidant capacity in IPEC-J2 cells which is effective against oxidative stress; the potential mechanism may involve the Nrf2 signaling pathway. Our findings could offer a novel nutritional intervention strategy to enhance the intestinal health of piglets during the weaning process.
1. Introduction
Oxidative stress arises from an imbalance within the oxidative and antioxidant systems and is related to an excessive yield of reactive oxygen species (ROS) [1]. It damages proteins, lipids, and DNA, resulting in tissue injury, cell death, and eventually causes the development of certain diseases [2]. The intestine is not only the main organ for digestion and absorption of nutrients, but it also acts as the most critical barrier against harmful pathogens, toxins, and antigens from the luminal environment [3,4]. However, as the boundary between the body and the environment, the intestine is more susceptible to oxidative stress due to its sustained exposure to the complex physiological or chemical environment [5,6]. Thus, protecting intestinal epithelial cells from oxidative stress is critical to the intestinal function, growth, and health of animals.
Based on the findings of our former research, it appears that soybean isoflavones play a crucial role in maintaining optimal antioxidant capacity and growth in pigs [7]. Excluding soybean isoflavones from their diet led to a reduction in growth performance and antioxidant properties, whereas re-adding soybean isoflavones prevented these negative effects [7]. However, soybean isoflavones in diets exist mainly in the form of glycosides, which are transformed into aglycones through deglycosylation under the action of enzymes in the intestine, and the aglycones are then absorbed by the intestine and circulated throughout the body [8,9]. In addition, the structure of glycosides is relatively complex and usually obtained by extraction, whereas the structure of aglycones is relatively simple and can be obtained by chemical synthesis, which not only significantly improves the purity but also greatly reduces the cost. Thus, we further investigated the application effect of daidzein (4′,7-dihydroxyisoflavone, an aglycone form of the soybean isoflavones) supplementation in basal diets containing soybean meal on weaned piglets, and the results showed that the basal diet added with daidzein (50 mg/kg) could efficaciously enhance antioxidant capacity and growth in weaned piglets [10]. Moreover, daidzein exerted protective effects in the porcine intestinal epithelial cells (IPEC-J2) oxidative stress model, and the underlying mechanism was possibly associated with activating the Nrf2 signaling pathway [10]. Genistein (4′,5,7-trihydroxyisoflavone), another main aglycone form of the soybean isoflavones, possess multiple bioeffects, including antioxidant, anticancer, and anti-inflammatory properties [11,12,13]. Several studies have reported the antioxidant properties of genistein [14,15]. However, only a limited number have explored the antioxidant properties of genistein, thus there is a lack of understanding regarding its positive effects on the alleviation of intestinal oxidative stress in pigs.
Therefore, the aim of our research was to estimate the antioxidant capacity and the potential mechanisms of genistein in IPEC-J2 cells. Our results provide insights for future applications of genistein as an antioxidant against intestinal oxidative stress in the pig industry.
2. Materials and Methods
2.1. Reagents
DMEM/F12, penicillin-streptomycin, fetal bovine serum, TRIzol reagent, SYBR Green, and RIPA buffer were supplied from Thermo Fisher Scientific (Waltham, MA, USA). Epidermal growth factor, and ITS (the mixture of insulin, transferrin, and selenious acids) were obtained from Corning Incorporated (New York, NY, USA), dimethyl sulfoxide, and genistein (synthetic product, purity ≥ 98%) were provided by Sigma-Aldrich (St. Louis, MO, USA). ROS assay kit was obtained from Beyotime Biotechnology (Shanghai, China). The cell counting kit (CCK-8) was bought from Med Chem Expression (Princeton, NJ, USA). The malondialdehyde (MDA), glutathione peroxidase (GSH-Px), catalase (CAT), and superoxide dismutase (SOD) assay kits were provided by Nanjing Jiancheng Bioengineering Institute (Nanjing, China). The Trans Script First-Strand cDNA Synthesis Kit was supplied by Trans Gen Biotech (Beijing, China). The VDF membranes, and ECL agent were provided by Bio-Rad Laboratories, Incorporated (Irvine, CA, USA).
2.2. Cell Culture
We thank the laboratory of Dr. Guoyao Wu at Texas A&M University for providing the IPEC-J2 cells, a well-established non-transformed porcine intestinal epithelial cell line derived from neonatal piglets’ mid-jejunum [16]. DMEM/F12 medium added with 0.01% epidermal growth factor, 0.1% ITS and 1% penicillin-streptomycin, and 5% fetal bovine serum was used for culture. The cells were cultured in a humidified incubator with 5% CO2 at 37 °C.
2.3. Selection of Genistein Concentration
The concentration (10 mg/mL) of genistein was dissolved using dimethyl sulfoxide. The 96-well plate was used to seed the IPEC-J2 cells with 1 × 104 cells per well for 24 h. Afterwards, the cells were pretreated with diverse concentrations of genistein (0, 10, 20, 40, 60, and 80 μM) for an additional 24 h, followed by treatment with or without 0.6 mM H2O2 for a further 1 h. The concentrations and treatment times of H2O2 are based on our prior research [10]. The CCK-8 kit was used to measure cell viability following the manufacturer’s instructions. In brief, following incubation for 3 h at 37 °C with 10% CCK-8 reagent, cell absorbance was recorded at 450 nm. The formula for calculating cell viability is as follows: Cell viability = [(the absorbance of the treatment group) − (the absorbance of the blank group)]/[(the absorbance of the control group) − (the absorbance of the blank group)] × 100%. We considered the control group’s cell viability to be 100%.
2.4. Determination of Intracellular ROS
The 96-well plate was used to seed the IPEC-J2 cells with 1 × 104 cells per well for 24 h. Then, the medium was changed with or without 20 μM genistein for an additional 24 h, followed by treating the cells with or without 0.6 mM H2O2 for another 1 h. The ROS assay kit was used to test the intracellular ROS levels based on the manufacturer’s instructions. The cells were cultured with DCFH-DA probes for 30 min, then washed 3 times using PBS. Subsequently, the fluorescence was recorded at 525 nm and 488 nm for emission and excitation, respectively.
2.5. Determination of Antioxidant Indices
The 6-well plate was used to seed the IPEC-J2 cells with 3 × 105 cells per well for 24 h. Then, the medium was changed with or without 20 μM genistein for an additional 24 h, followed by treatment of the cells with or without 0.6 mM H2O2 for a further 1 h. For subsequent lysis, we used RIPA buffer for 30 min, and centrifuged the supernatant for 30 min at 13,000× g. Then, we determined the malondialdehyde (MDA) levels, and CAT, SOD, GSH-Px activities using the MDA, CAT, SOD and GSH-Px assay kits as directed by the manufacturer. Briefly, MDA concentration was analyzed with 2-thiobarbituric acid, and the change in absorbance was read at 532 nm. CAT activity was measured with ammonium molybdate, and the change in absorbance was recorded at 405 nm. SOD activity was measured through a nonenzymatic NBT test, which measures the inhibition of the formation of superoxide anion free radicals that reduce the nitroblue tetrazolium of the sample. The change in absorbance at 450 nm was recorded. GSH-Px activity was measured with 5,50-dithiobis-p-nitrobenzoic acid, and the change in absorbance at 412 nm was recorded.
2.6. RNA Extraction, Reverse Transcription, and Quantitative Real-Time PCR (qPCR)
The 12-well plate was used to seed the IPEC-J2 cell with 1.5 × 105 cells per well for 24 h. Following this, the medium was changed with or without 20 μM genistein for an additional 24 h, followed by treatment of the cells with or without 0.6 mM H2O2 for another 1 h. TRIzol reagent was used to extract the total RNA from the cells according to the manufacturer’s instructions. The concentration and A260/A280 ratio of the total RNA was determined using an Epoch microplate spectrophotometer. An A260/280 ratio ranging between 1.8 and 2.0 was subjected to further analysis. Total RNA (1 μg) was reverse transcribed into cDNA using a Trans Script First-Strand cDNA Synthesis Kit. SYBR Green PCR Master Mix was used to perform qPCR analysis in the CFX96 Real-Time System. The qPCR reaction system was: 2 μL cDNA template, 0.5 μL forward primer, 0.5 μL reverse primer, 10 μL SYBR Green PCR Master Mix, and 7 μL DNase and RNase free water. The reaction procedure was as follows: denature: 50 °C for 2 min, 95 °C for 10 min; extension: 95 °C for 15 s, 60 °C for 1 min, 40 cycles; and melt curve: 95 °C for 15 s, 60 °C for 1 min, 95 °C for 15 s. The relative expressions of all the target genes were calculated using the 2−ΔΔCT method [17] and the housekeeping gene was glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Table 1 presents the sequences of primers.
2.7. Western Blotting
The 6-well plates were used to seed the IPEC-J2 cell with 3 × 105 cells per well for 24 h. Then, the medium was changed with or without 20 μM genistein for an additional 24 h, followed by treatment of the cells with or without 0.6 mM H2O2 for a further 1 h. For subsequent lysis, we used RIPA buffer for 30 min, and centrifuged the supernatant for 30 min at 13,000× g. The samples were transferred to PVDF membranes after being separated by the SDS-PAGE (12%). 5% skim milk in Tris-buffered saline with Tween 20 was used to block PVDF membranes at room temperature for 3 h, followed by overnight incubation at 4 °C in the presence of primary antibodies, and finally, they were incubated at 4 °C with secondary antibodies for 1 h. The ECL agent was used to detect the chemiluminescence signals, which were then visualized using the ChemiDoc MP Imaging System. The band intensity was quantified using Image J software (v1.8.0). The band of GAPDH was considered as the internal reference band. Table 2 lists detailed information on antibodies.
2.8. Statistical Analysis
Data analysis was performed using SPSS 20.0 software with the one-way ANOVA procedure. Treatment differences were assessed using Tukey’s post-hoc test. p < 0.05 was considered statistically significant, whereas 0.05 ≤ p < 0.10 indicated a trending treatment effect.
3. Results
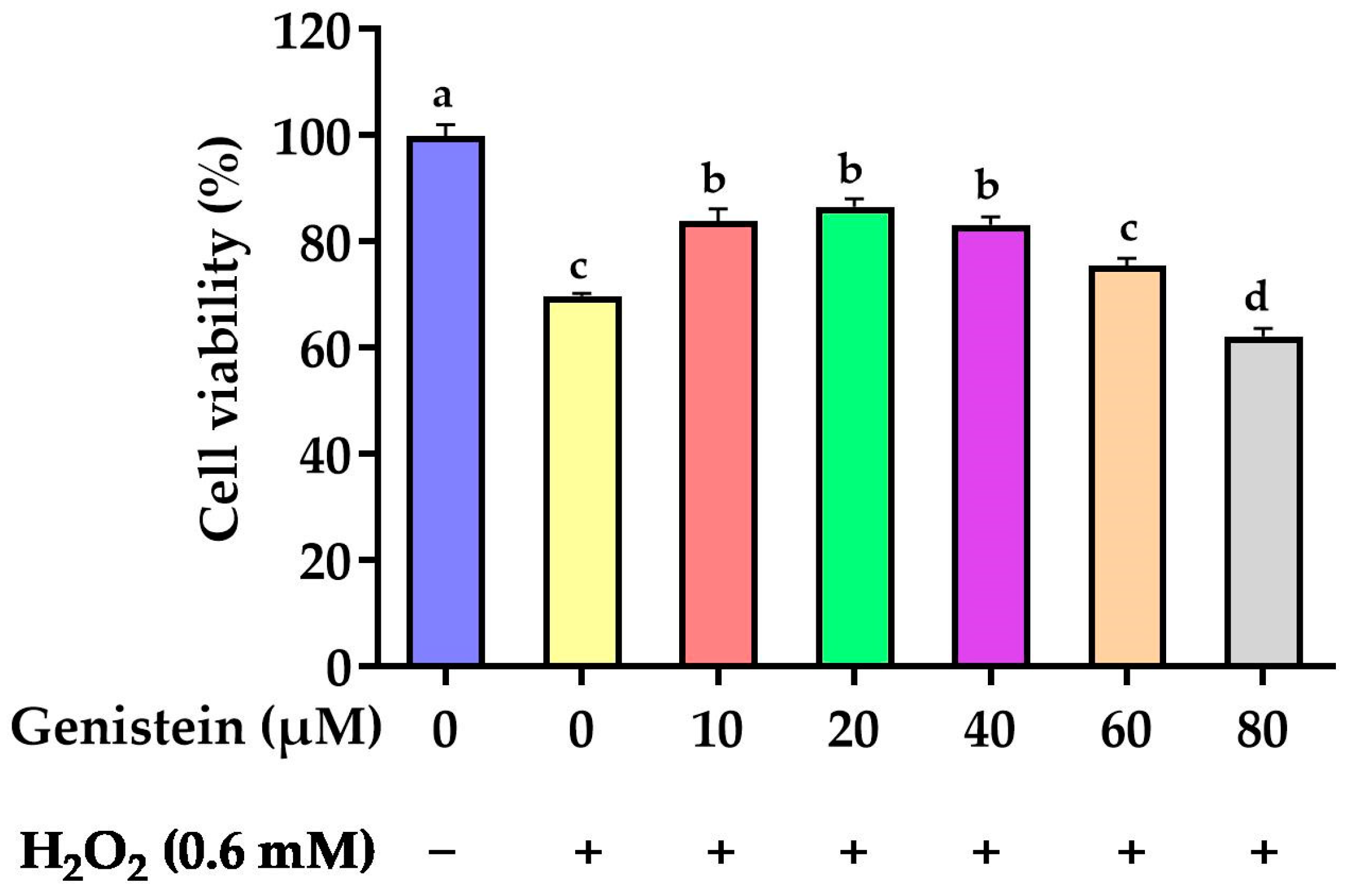
3.1. Protective Effect of Genistein on Cell Viability
As revealed in Figure 1, cell viability was significantly reduced from 100% to 69.8% in the H2O2-treated group compared to the control group (p < 0.05). However, in contrast to the H2O2-treated group, pretreating cells with 10, 20, and 40 μM genistein before H2O2 exposure enhanced cell viability from 69.8% to 83.8%, 86.5%, and 83.2%, respectively (p > 0.05). For subsequent experiments, we opted for a concentration of 20 μM genistein as it exhibited enhanced cell viability at this particular dosage.
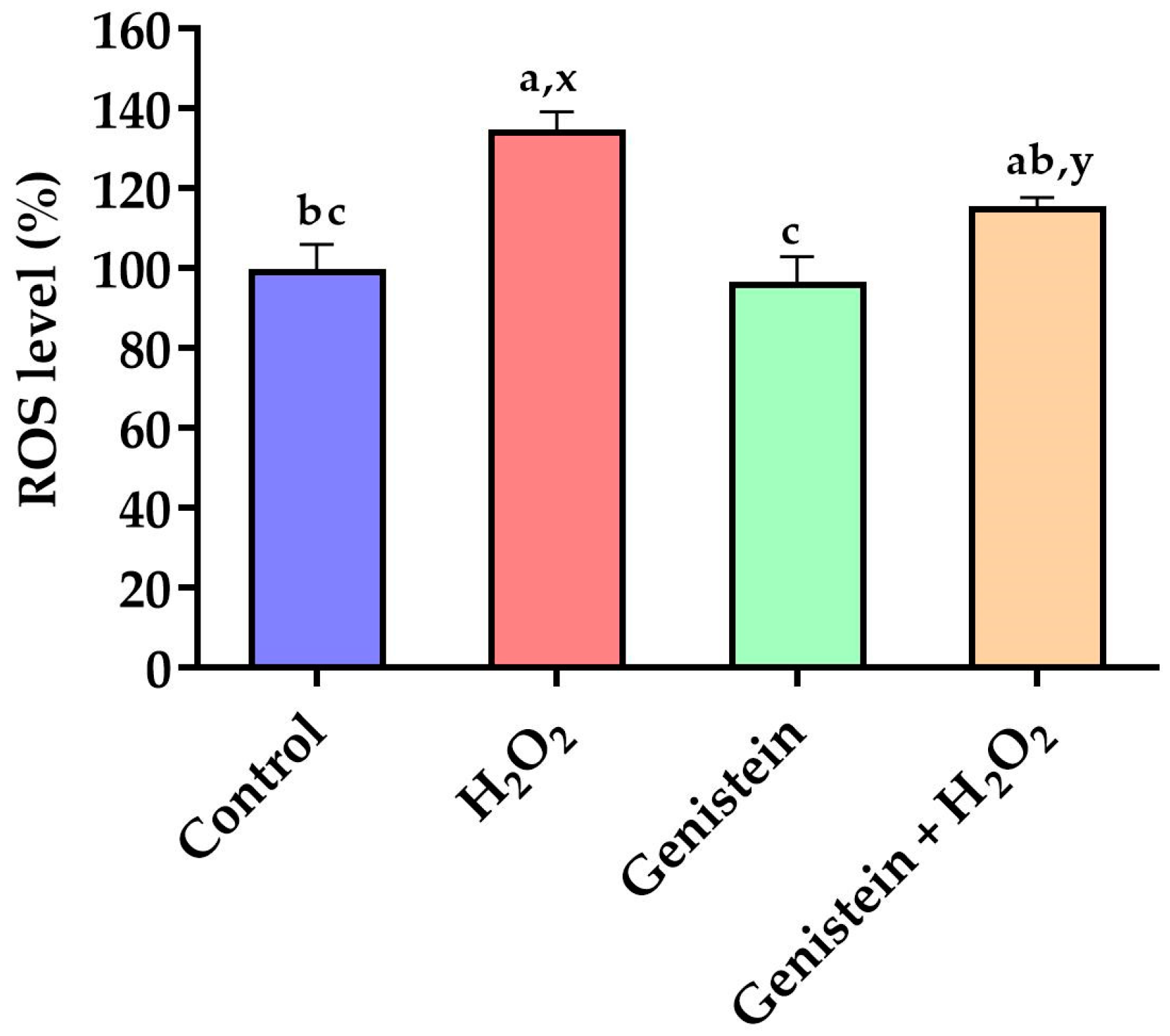
3.2. Intracellular ROS Levels
As demonstrated in Figure 2, a significant elevation in intracellular ROS levels was observed in the H2O2-treated group compared to the control group (p < 0.05). In contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure showed a tendency to reduce intracellular ROS levels (p = 0.061). Comparatively, the genistein treated group did not have elevated intracellular ROS levels (p > 0.05).
3.3. Antioxidant Enzyme Activities and MDA Level
According to Figure 3, H2O2 stimulation significantly reduced the CAT activity (p < 0.05) and showed a tendency to increase the MDA level (p = 0.06) compared to the control group. However, in contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure significantly elevated the CAT activity (p < 0.05). No significant differences in GSH-Px and SOD activities were observed (p > 0.05).
3.4. Expression of Key Genes in Nrf2 Signaling Pathway
As presented in Figure 4, a significant reduction in the gene expression of Nrf2 and CAT was observed in the H2O2-treated group compared to the control group (p < 0.05). However, in contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure evidently elevated the gene expression of SOD1, Nrf2, CAT, GPX1, NQO1, and HO-1. In addition, a significant increase in the gene expression of SOD1, CAT, Nrf2, NQO1, GPX1, and HO-1 was observed in the genistein treated group compared to the control group (p < 0.05).
3.5. Tight Junction Gene Expression
As displayed in Figure 5, a remarkable reduction in the gene expression of occludin and ZO-1 was observed in the H2O2-treated group compared to the control group (p < 0.05). However, in contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure led to a significant increase in the gene expression of occludin and ZO-1 (p < 0.05). In addition, the gene expression of ZO-1 and occludin in the genistein treated group was significantly increased in contrast to the control group (p < 0.05). Whereas, no significant differences in the gene expression of claudin 1 were observed (p > 0.05).
3.6. Nrf2 Protein Expression
According to Figure 6, a remarkable reduction in the relative protein abundance of Nrf2 was observed in the H2O2-treated group compared to the control group (p < 0.05). However, in contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure led to a significant increase in the relative protein abundance of Nrf2 (p < 0.05). Comparatively, the genistein treated group did not display the elevated relative protein abundance of Nrf2 (p > 0.05).
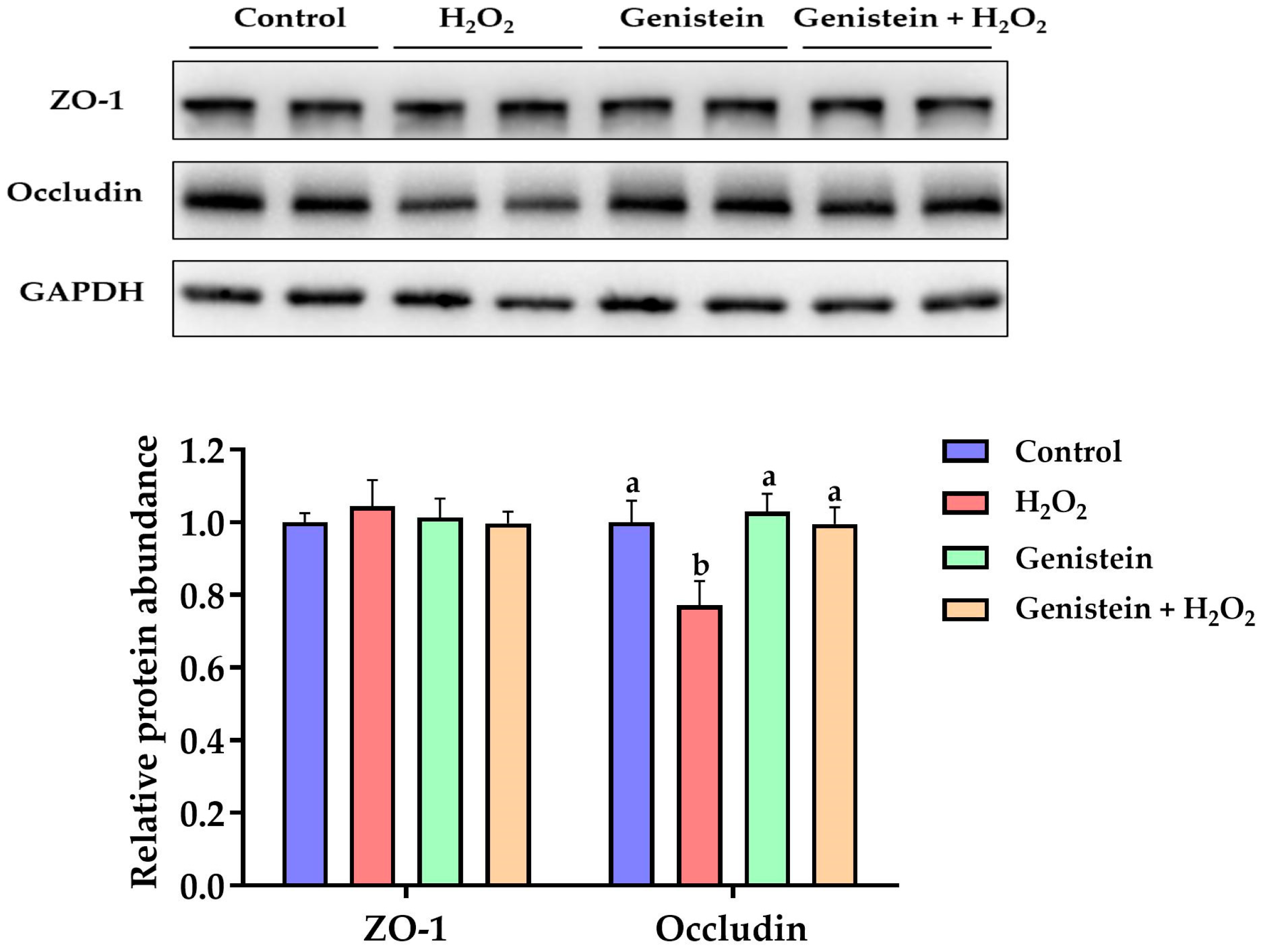
3.7. Tight Junction Protein Expression
As presented in Figure 7, a remarkable reduction in the relative protein abundance of occludin was observed in the H2O2-treated group compared to the control group (p < 0.05). However, in contrast to the H2O2-treated group, pretreating cells with 20 μM genistein before H2O2 exposure led to a significant increase in the relative protein abundance of occludin (p < 0.05). On the other hand, compared to the control group, the relative protein abundance of occludin in the genistein treatment group was not affected (p > 0.05). Furthermore, no significant differences in the relative protein abundance of ZO-1 were observed (p > 0.05).
4. Discussion
The level of intracellular ROS was maintained at a specific level, and any excessive ROS was typically eliminated by antioxidant enzymes under normal physiological conditions. Nevertheless, oxidative stress arises when the levels of ROS surpass the body’s antioxidant defense system, making it unable to remove them effectively [18]. Oxidative stress can damage proteins, lipids, and DNA, resulting in tissue injury and cell death, which can ultimately cause various diseases [19,20]. ROS production serves as a primary indicator of oxidative stress [21]. However, as the vital components in the antioxidant defense system, CAT, SOD, and GSH-Px can scavenge for ROS and reflect the body’s antioxidant capacity [22]. Whereas MDA is the final product of lipid peroxidation, the level of MDA reflects the degree of cell damage induced by oxidative stress. In the current research, the antioxidant capacity and underlying mechanisms of genistein were investigated through the use of the IPEC-J2 cell line as an in vitro cellular model. H2O2 was employed as the oxidant to induce oxidative stress as in previous research [23,24,25]. The findings of the current study indicated that H2O2 markedly decreased the IPEC-J2 cells’ viability, whereas pretreatment with genistein prior to H2O2 exposure markedly alleviated the decrease of cell viability caused by H2O2. To determine the antioxidant capacity of genistein, we analyzed ROS production, MDA levels, and the activities CAT, SOD, and GSH-Px. The results showed that genistein pretreatment decreased ROS levels, and enhanced CAT activity in H2O2-stimulated IPEC-J2 cells, indicating that genistein demonstrated protective properties and resists oxidative stress in IPEC-J2 cells. This result aligns with our previous study, in which daidzein (the aglycone form of the soybean isoflavones) significantly reduced MDA and ROS levels, and enhanced CAT activity when IPEC-J2 cells were stimulated by H2O2 [10]. Furthermore, our observations corresponded to other studies [26,27]. The isoflavonoid-enriched kudzu root extract recovered the downregulation of cell viability and alleviated the increase of ROS levels caused by rotenone in human umbilical vein endothelial cells (HUVECs) [26], and genistein pretreatment mitigated the decrease of SOD activity caused by oleic acid hydroperoxide in Caco-2 cells [27]. These results suggested that genistein had the potential to be used as an antioxidant against oxidative stress. In a future study, we will investigate whether dietary genistein improves the antioxidant capacity of pigs.
Demonstrations generally show that the Nrf2 signaling pathway protects against oxidative stress [28,29]. Nrf2 was bound to Kelch-like ECH-associated protein 1 (Keap1) and primarily located in the cytoplasm in an inactive state under normal physiological conditions. Upon exposure to oxidative stress, Nrf2 translocated from the cytoplasm to the nucleus after dissociating from Keap1, leading to the transcription of detoxifying enzymes [30,31]. In the current experiment, IPEC-J2 cells exposed to H2O2 had a significant lower gene expression of Nrf2 and CAT, whereas pretreating cells with genistein before H2O2 exposure resulted in a significant increase in the gene expression of Nrf2 and CAT. Furthermore, under the same conditions, the relative protein expression of Nrf2 correlated with the gene expression of Nrf2 and CAT, indicating that genistein demonstrated protective properties and resisted oxidative stress in IPEC-J2 cells, and this mechanism may be associated with the Nrf2 signaling pathway. This observation was consistent with our previous study, in which daidzein exerted protective properties and resisted oxidative stress in IPEC-J2 cells by regulating Nrf2 and its target gene expressions [10]. Similarly, in the research of Bai and Wang (2019), genistein protects against doxorubicin-induced cardiotoxic effects via activating the Nrf2 signaling pathway in mice models [11]. Miao et al. (2018) indicated that genistein could alleviate cerebral ischemia-induced oxidative stress injury in ovariectomized rats by promoting Nrf2 and NQO1 expression [32]. These findings demonstrated that genistein exerted an antioxidant capacity via the activation of the Nrf2 signaling pathway. However, the crucial role of the Nrf2 signaling pathway in genistein’s resistance to oxidative stress needs to be further confirmed by knocking out Nrf2 in our future research.
The intestinal epithelium serves a dual purpose: it facilitates the absorption of nutrients in addition to acting as a physical barrier preventing harmful pathogens, antigens, and toxins from the luminal environment permeating into the circulatory system [33]. Tight junctions, which are a key component of the intestinal physical barrier, have a strong association with intestinal permeability and play a vital role in maintaining gut health [34].
The structural composition of tight junctions consists of transmembrane proteins, such as zona occludens, occludin, and claudins [35], and ZO-1, occludin, and claudin 1 are three crucial tight junction proteins [36]. Previous studies have demonstrated that increased ZO-1 and occludin expression is associated with reduced intestinal permeability in weaned piglets [37,38]. In the current study, IPEC-J2 cells exposed to H2O2 had a significantly lower expressions of occludin at the gene and protein levels and ZO-1 at the gene level, whereas pretreating cells with genistein followed by H2O2 exposure reversed this change. Interestingly, we found H2O2 could reduce ZO-1 expression at the gene level rather than at the protein level, indicating that H2O2 may predominantly regulate its expression at the transcriptional level. We speculate that the protective properties of genistein on the intestinal barrier may be linked to its antioxidant capacity, and further study needs to be carried out to explore the underlying mechanism of genistein affecting the intestinal barrier under oxidative stress. Collectively, our findings indicated that genistein has the potential to maintain intestinal barrier function under oxidative stress.
5. Conclusions
In summary, the current study revealed that genistein exerted protective properties and resists oxidative stress in IPEC-J2 cells, and the potential mechanism may be associated with the Nrf2 signaling pathway. The in vitro study that was performed will be replicated in vivo to confirm the results. Our findings can provide a theoretical basis for improving intestinal oxidative stress in piglets through nutritional interventions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/vetsci11040154/s1, Figures S1 and S2: Original image of Figure 6; Figures S3 and S4: Original image of Figure 7.
Author Contributions
Conceptualization, Y.L. and X.L.; methodology, Y.L. and X.L.; software, Y.L.; validation, Y.L., Y.P. and X.J.; formal analysis, Y.L. and L.C.; investigation, Y.L. and L.C.; resources, X.L.; data curation, Y.L., X.J. and X.L.; writing—original draft preparation, Y.L. and L.C.; writing—review and editing, Y.L., L.C., Q.B., W.S., Y.P., X.J. and X.L.; visualization, Y.L., L.C. and Y.P.; supervision, Y.P. and X.L.; project administration, Y.P. and X.L.; funding acquisition, Y.P. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Elite Youth Program of the Chinese Academy of Agricultural Sciences (to X.L.), the China Postdoctoral Science Foundation (2023M730594), the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ZDRW202305).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used for the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank the graduate students and assistants in the Lab of Swine Nutrition and Feed of the Institute of Feed Research for their assistance in experimenting.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Jelic, M.D.; Mandic, A.D.; Maricic, S.M.; Srdjenovic, B.U. Oxidative stress and its role in cancer. J. Cancer Res. Ther. 2021, 17, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Catalioto, R.M.; Maggi, C.A.; Giuliani, S. Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr. Med. Chem. 2011, 18, 398–426. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, H.; Wang, S.; Tu, Z.; Zhang, L.; Wang, X.; Hou, Y.; Wang, C.; Chen, J.; Liu, Y. Flaxseed oil attenuates intestinal damage and inflammation by regulating necroptosis and TLR4/NOD signaling pathways following lipopolysaccharide challenge in a piglet model. Mol. Nutr. Food Res. 2018, 62, e1700814. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wu, H.; Wang, X.; He, J.; He, S.; Yin, Y. Resveratrol attenuates oxidative stress-induced intestinal barrier injury through PI3K/Akt-mediated Nrf2 Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 7591840. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Wang, S.; Wang, Y.; Chu, X.; Ji, H. Lactobacillus plantarum exhibits antioxidant and cytoprotective activities in porcine intestinal epithelial cells exposed to hydrogen peroxide. Oxid. Med. Cell. Longev. 2021, 2021, 8936907. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Jiang, X.R.; Wei, Z.X.; Cai, L.; Yin, J.D.; Li, X.L. Effects of soybean isoflavones on the growth performance, intestinal morphology and antioxidative properties in pigs. Animal 2020, 14, 2262–2270. [Google Scholar] [CrossRef] [PubMed]
- Walsh, K.R.; Haak, S.J.; Fastinger, N.D.; Bohn, T.; Tian, Q.; Mahan, D.C.; Schwartz, S.J.; Failla, M.L. Gastrointestinal absorption and metabolism of soy isoflavonoids in ileal-canulated swine. Mol. Nutr. Food Res. 2009, 53, 277–286. [Google Scholar] [CrossRef]
- Cools, S.; Van den Broeck, W.; Vanhaecke, L.; Heyerick, A.; Bossaert, P.; Hostens, M.; Opsomer, G. Feeding soybean meal increases the blood level of isoflavones and reduces the steroidogenic capacity in bovine corpora lutea, without affecting peripheral progesterone concentrations. Anim. Reprod. Sci. 2014, 144, 79–89. [Google Scholar] [CrossRef]
- Li, Y.P.; Jiang, X.R.; Cai, L.; Zhang, Y.L.; Ding, H.B.; Yin, J.D.; Li, X.L. Effects of daidzein on antioxidant capacity in weaned pigs and IPEC-J2 cells. Anim. Nutr. 2022, 11, 48–59. [Google Scholar] [CrossRef]
- Bai, Z.; Wang, Z. Genistein protects against doxorubicin-induced cardiotoxicity through Nrf-2/HO-1 signaling in mice model. Environ. Toxicol. 2019, 34, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Calvello, R.; Aresta, A.; Trapani, A.; Zambonin, C.; Cianciulli, A.; Salvatore, R.; Clodoveo, M.L.; Corbo, F.; Franchini, C.; Panaro, M.A. Bovine and soybean milk bioactive compounds: Effects on inflammatory response of human intestinal Caco-2 cells. Food Chem. 2016, 210, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y. Soy isoflavones and cancer prevention. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Yang, Z.Q.; Huang, J.C.; Wang, Y.S.; Guo, B.; Yue, Z.P. Genistein protects ovarian granulosa cells from oxidative stress via cAMP-PKA signaling. Cell Biol. Int. 2020, 44, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Pang, X.; Yu, H.; Zhou, H. Genistein suppresses ox-LDL-elicited oxidative stress and senescence in HUVECs through the SIRT1-p66shc-Foxo3a pathways. J. Biochem. Mol. Toxicol. 2022, 36, e22939. [Google Scholar] [CrossRef] [PubMed]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhu, Y.; Yan, S.; Xiao, H.; Yi, J.; Li, R.; Wu, J.; Wen, L. Phenethyl isothiocyanate induces IPEC-J2 cells cytotoxicity and apoptosis via S-G2/M phase arrest and mitochondria-mediated Bax/Bcl-2 pathway. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 226, 108574. [Google Scholar] [CrossRef] [PubMed]
- Kaschubek, T.; Mayer, E.; Rzesnik, S.; Grenier, B.; Bachinger, D.; Schieder, C.; König, J.; Teichmann, K. Effects of phytogenic feed additives on cellular oxidative stress and inflammatory reactions in intestinal porcine epithelial cells. J. Anim. Sci. 2018, 96, 3657–3669. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Wei, H.; Yu, H.; Xing, Q.; Zou, Y.; Zhou, Y.; Peng, J. Fish skin gelatin hydrolysate production by ginger powder induces glutathione synthesis to prevent hydrogen peroxide induced intestinal oxidative stress via the Pept1-p62-Nrf2 Cascade. J. Agric. Food Chem. 2018, 66, 11601–11611. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, C.; Yan, J.; Li, X.; Wen, J.; Hu, C. Curcumin ameliorates oxidative stress-induced intestinal barrier injury and mitochondrial damage by promoting Parkin dependent mitophagy through AMPK-TFEB signal pathway. Free Radic. Biol. Med. 2020, 147, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, X.; He, C. An isoflavonoid-enriched extract from Pueraria lobata (kudzu) root protects human umbilical vein endothelial cells against oxidative stress induced apoptosis. J. Ethnopharmacol. 2016, 193, 524–530. [Google Scholar] [CrossRef]
- Wijeratne, S.S.; Cuppett, S.L. Soy isoflavones protect the intestine from lipid hydroperoxide mediated oxidative damage. J. Agric. Food Chem. 2007, 55, 9811–9816. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.L.; Zhou, L.L.; Weng, Q.; Xiao, L.X.; Li, Q.Y. Curcumin analogues attenuate Aβ25-35-induced oxidative stress in PC12 cells via Keap1/Nrf2/HO-1 signaling pathways. Chem. Biol. Interact. 2019, 305, 171–179. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, M.; Li, J.; Shan, A. DL-Selenomethionine alleviates oxidative stress induced by zearalenone via Nrf2/Keap1 signaling pathway in IPEC-J2 cells. Toxins 2021, 13, 557. [Google Scholar] [CrossRef]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1-Nrf2-ARE pathway as a potential preventive and therapeutic target: An update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.Y.; Xia, X.; Che, L.; Song, Y.T. Genistein attenuates brain damage induced by transient cerebral ischemia through up-regulation of Nrf2 expression in ovariectomized rats. Neurol. Res. 2018, 40, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Role of flavonoids in intestinal tight junction regulation. J. Nutr. Biochem. 2011, 22, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- He, C.M.; Deng, J.; Hu, X.; Zhou, S.C.; Wu, J.T.; Xiao, D.; Darko, K.O.; Huang, Y.J.; Tao, T.; Peng, M.; et al. Vitamin A inhibits the action of LPS on the intestinal epithelial barrier function and tight junction proteins. Food Funct. 2019, 10, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
Figure 1.
Effect of genistein on the IPEC-J2 cells viability. The 96-well plate was used to seed the IPEC-J2 cells, then the medium was changed with various genistein concentrations for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–d.
Figure 1.
Effect of genistein on the IPEC-J2 cells viability. The 96-well plate was used to seed the IPEC-J2 cells, then the medium was changed with various genistein concentrations for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–d.

Figure 2.
Effect of genistein on ROS levels in IPEC-J2 cells. The 96-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h, and finally, the cells were cultured with DCFH-DA probes for 30 min. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–c; and there is a tendency (0.05 ≤ p < 0.10) between the means assigned the letters x, y.
Figure 2.
Effect of genistein on ROS levels in IPEC-J2 cells. The 96-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h, and finally, the cells were cultured with DCFH-DA probes for 30 min. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–c; and there is a tendency (0.05 ≤ p < 0.10) between the means assigned the letters x, y.
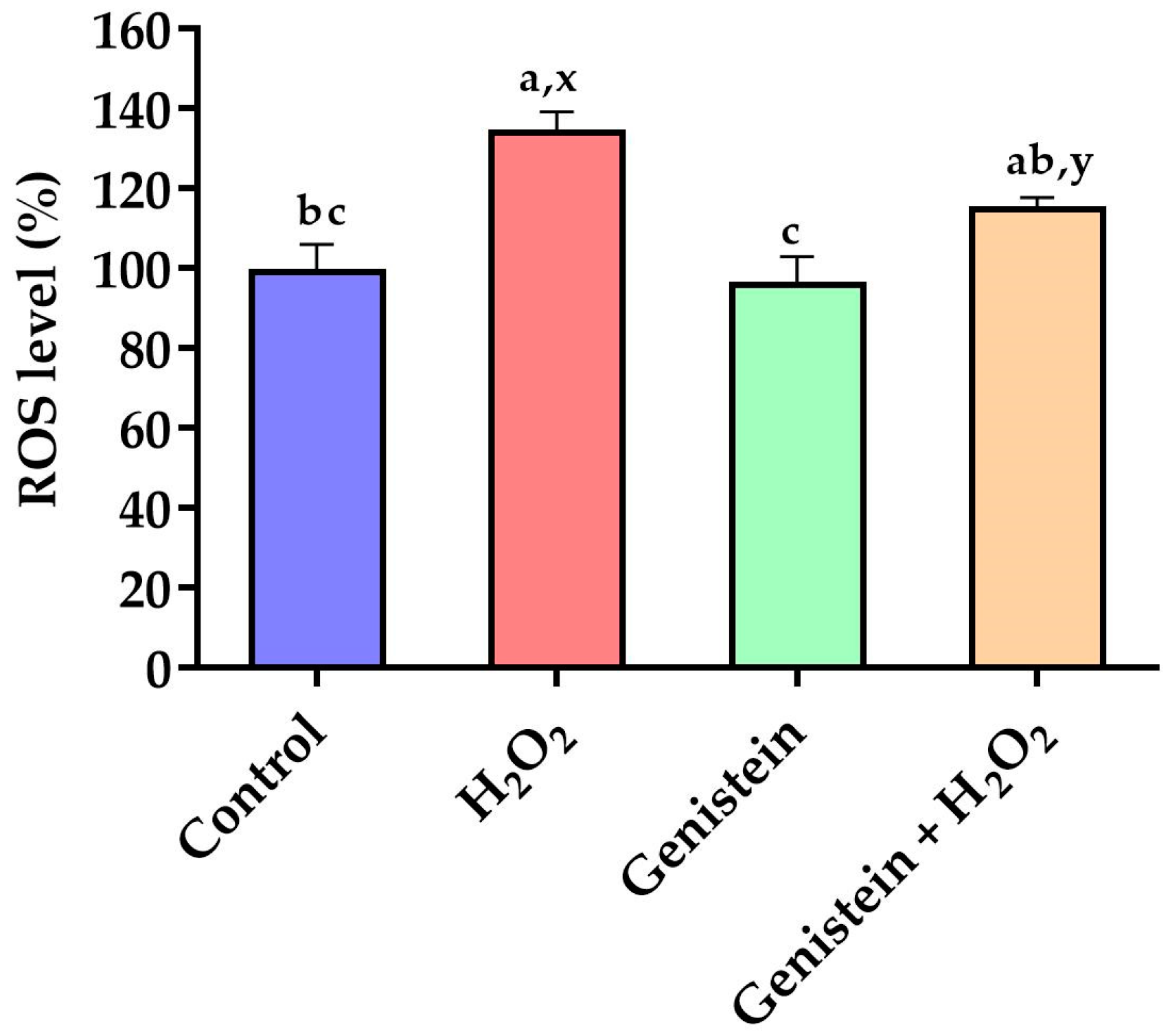
Figure 3.
Effects of genistein on MDA level and antioxidant enzyme activities in IPEC-J2 cells. (A) Antioxidant enzyme activities; (B) MDA level. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a, b; and there is a tendency (0.05 ≤ p < 0.10) between the means assigned the letters x, y.
Figure 3.
Effects of genistein on MDA level and antioxidant enzyme activities in IPEC-J2 cells. (A) Antioxidant enzyme activities; (B) MDA level. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a, b; and there is a tendency (0.05 ≤ p < 0.10) between the means assigned the letters x, y.
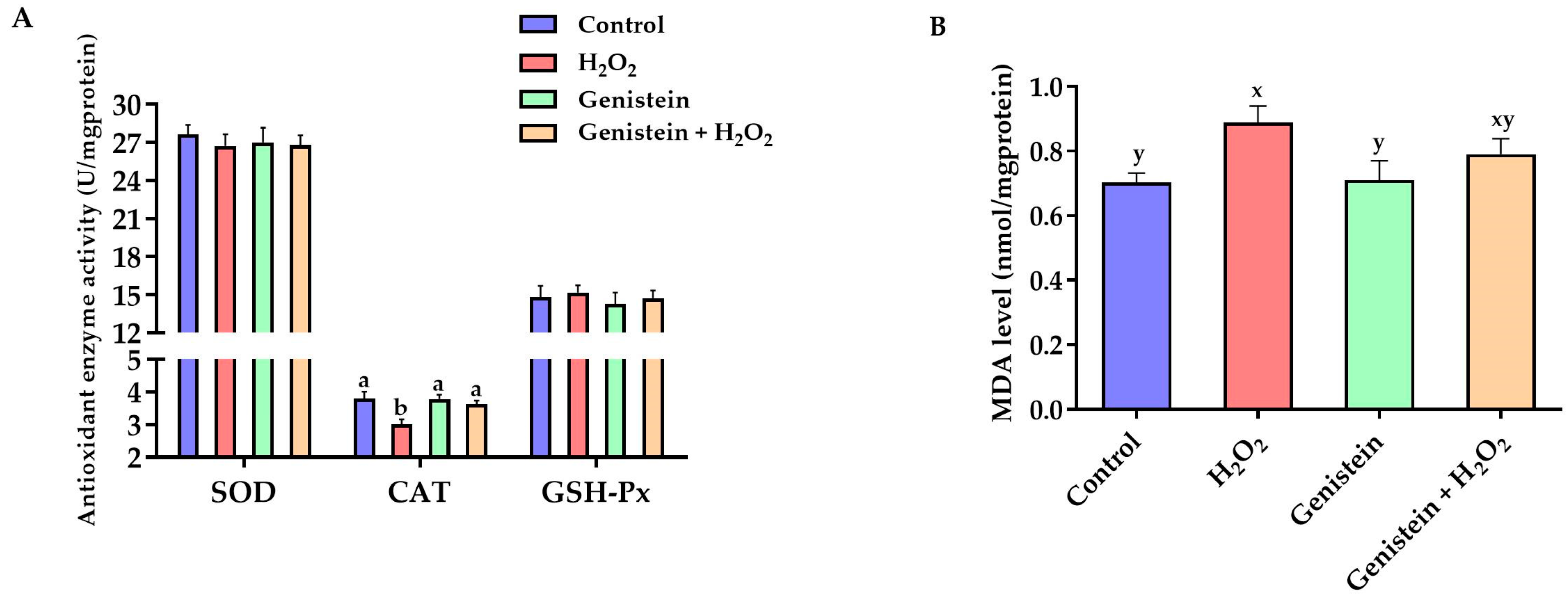
Figure 4.
Effect of genistein on the expression of key genes in the Nrf2 signaling pathway in IPEC-J2 cells. The 12-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–c.
Figure 4.
Effect of genistein on the expression of key genes in the Nrf2 signaling pathway in IPEC-J2 cells. The 12-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between the means assigned the letters a–c.
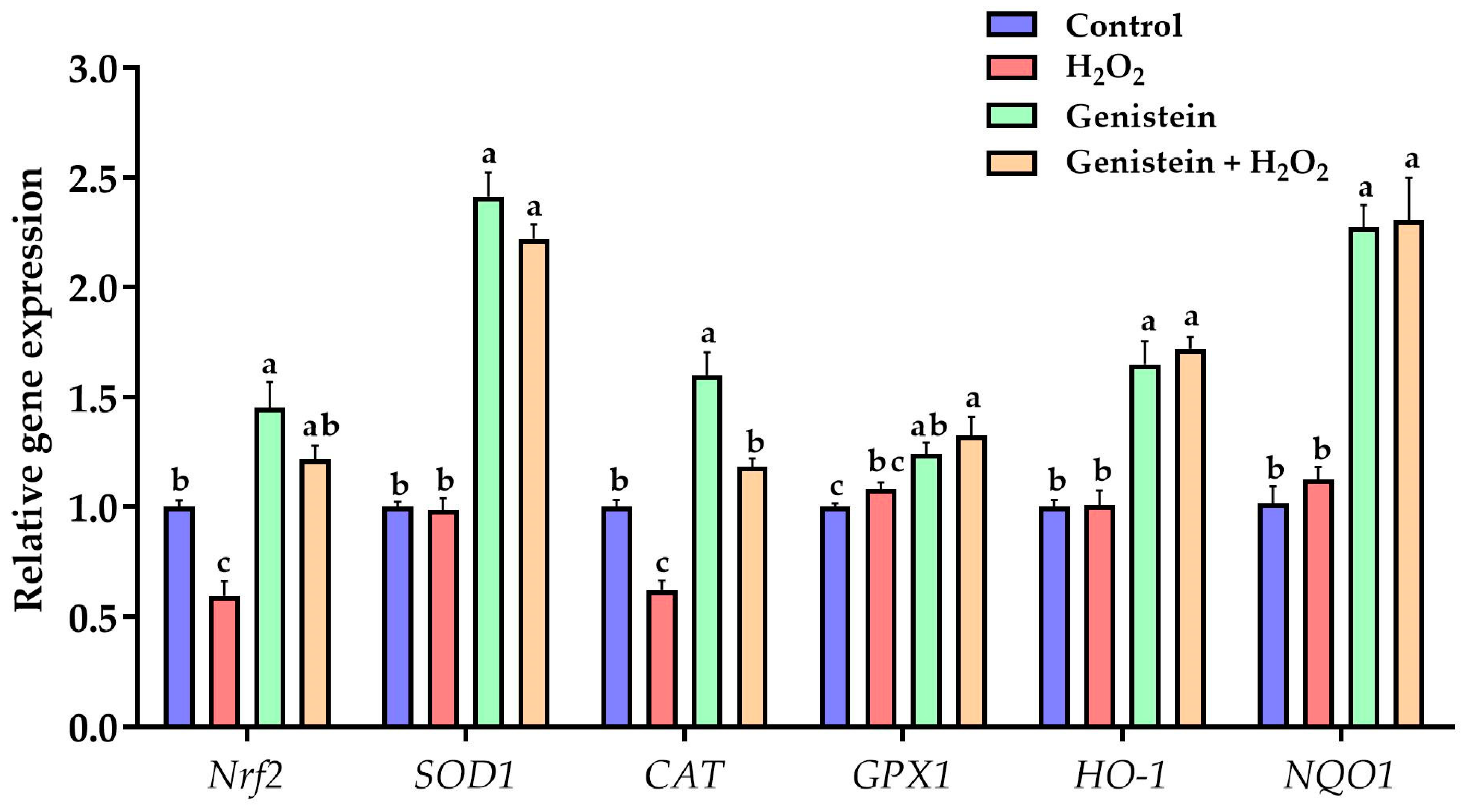
Figure 5.
Effect of genistein on the tight junction gene expression in H2O2-treated IPEC-J2 cells. The 12-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between means assigned the letters a–c.
Figure 5.
Effect of genistein on the tight junction gene expression in H2O2-treated IPEC-J2 cells. The 12-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 6. There are statistically significant differences (p < 0.05) between means assigned the letters a–c.
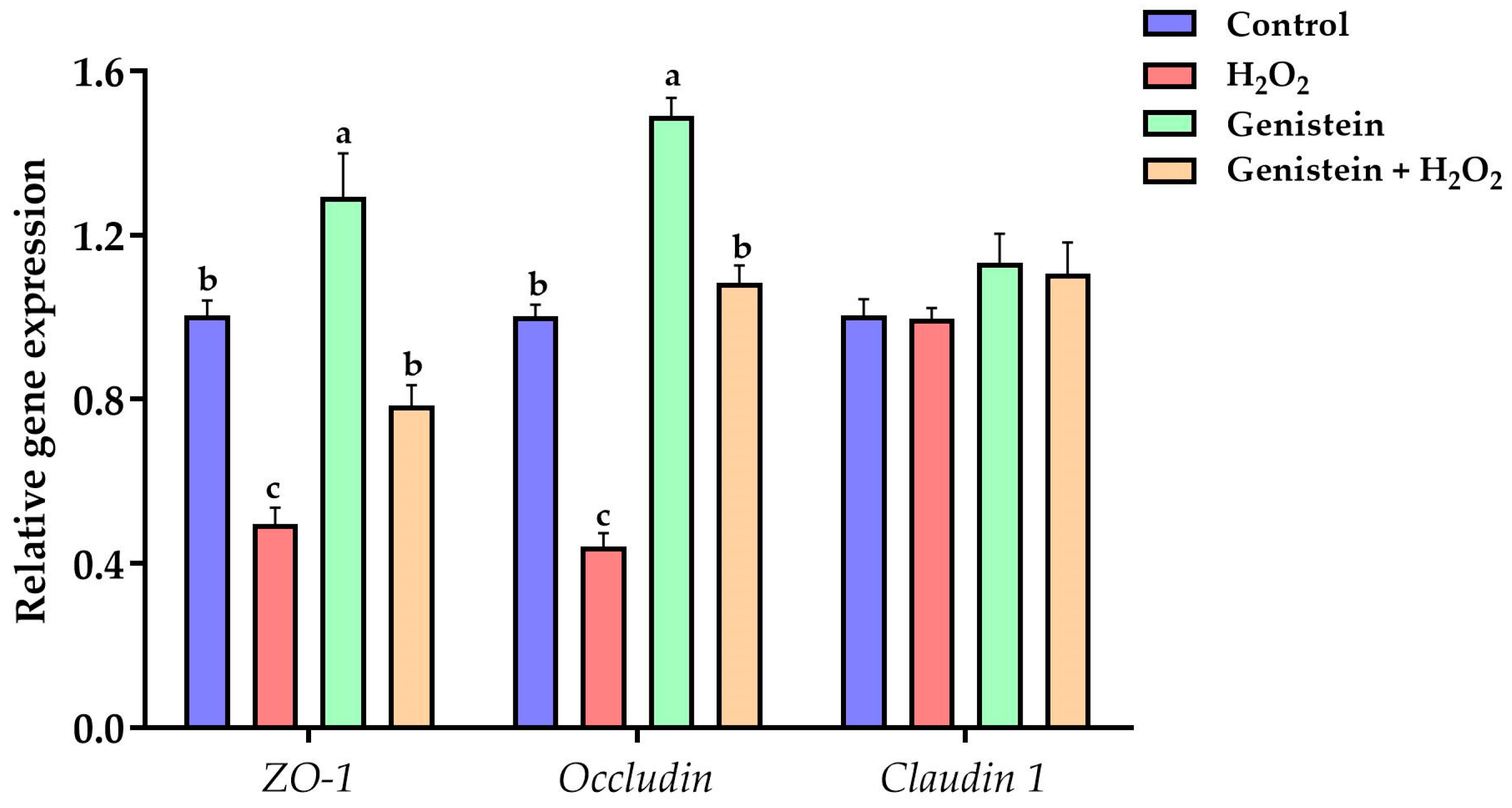
Figure 6.
Effect of genistein on the Nrf2 protein expression in IPEC-J2 cells. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 4. There are statistically significant differences (p < 0.05) between means assigned the letters a, b. (Figures S1 and S2).
Figure 6.
Effect of genistein on the Nrf2 protein expression in IPEC-J2 cells. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 4. There are statistically significant differences (p < 0.05) between means assigned the letters a, b. (Figures S1 and S2).
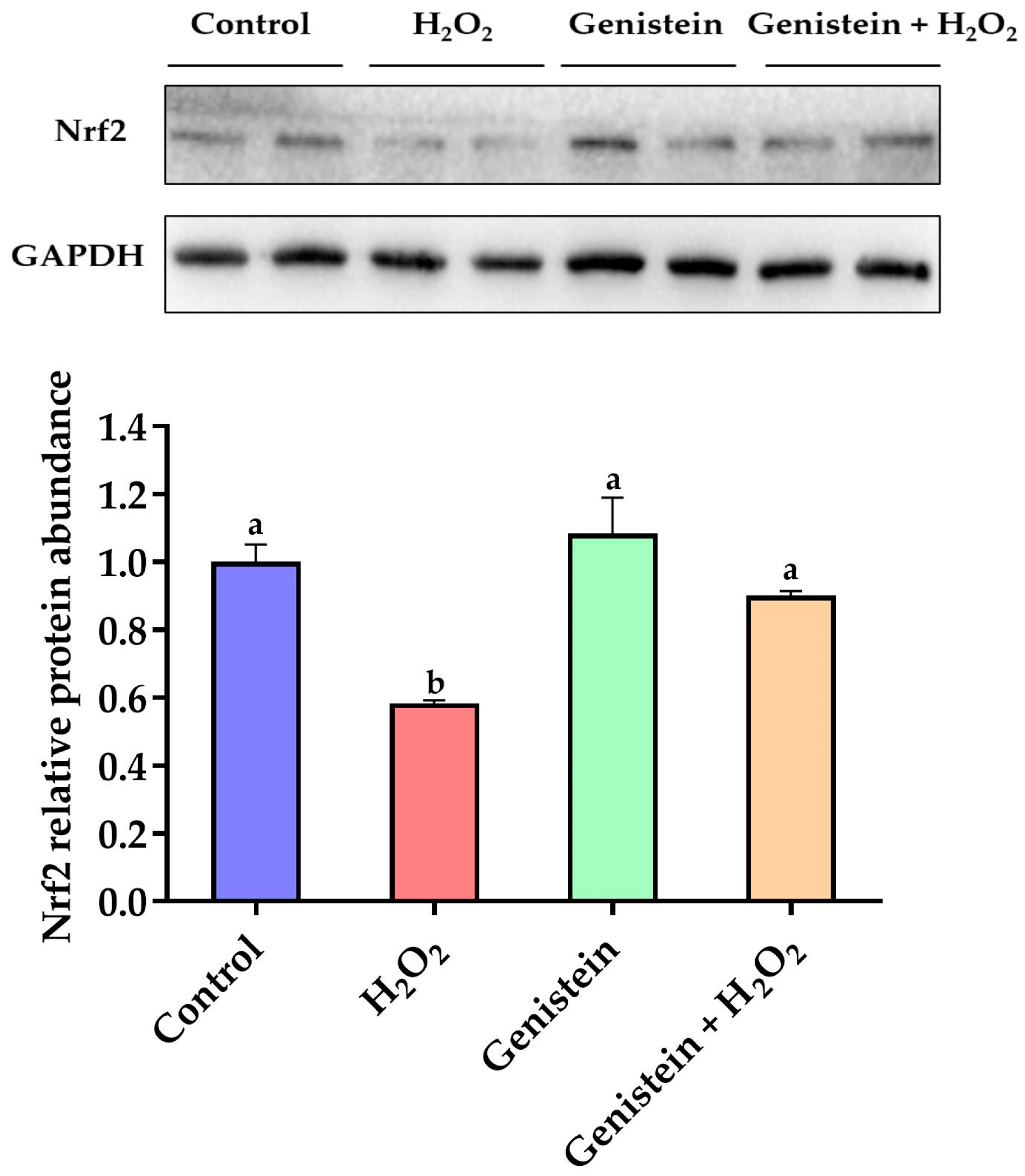
Figure 7.
Effect of genistein on the tight junction protein expression in H2O2-treated IPEC-J2 cells. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 4. There are statistically significant differences (p < 0.05) between means assigned the letters a, b. (Figures S3 and S4).
Figure 7.
Effect of genistein on the tight junction protein expression in H2O2-treated IPEC-J2 cells. The 6-well plate was used to seed the IPEC-J2 cells, then the medium was changed with or without 20 μM genistein for 24 h, followed by the treatment of cells with or without 0.6 mM H2O2 for 1 h. Values are means ± standard error, n = 4. There are statistically significant differences (p < 0.05) between means assigned the letters a, b. (Figures S3 and S4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer sequences used for quantitative real-time PCR.
| Gene | Forward (5′-3′) | Reverse (5′-3′) | Product Length, bp | Accession No. |
|---|---|---|---|---|
| GAPDH | GCTTGTCATCAATGGAAAGG | CATACGTAGCACCAGCATCA | 86 | NM_001206359.1 |
| SOD1 | GAAGACAGTGTTAGTAACGG | CAGCCTTGTGTATTATCTCC | 93 | NM_001190422.1 |
| CAT | CCTGCAACGTTCTGTAAGGC | GCTTCATCTGGTCACTGGCT | 72 | NM_214301.2 |
| GPX1 | TCTCCAGTGTGTCGCAATGA | TCGATGGTCAGAAAGCGACG | 104 | NM_214201.1 |
| Nrf2 | GACCTTGGAGTAAGTCGAGA | GGAGTTGTTCTTGTCTTTCC | 103 | XM_005671981.3 |
| HO-1 | GAGAAGGCTTTAAGCTGGTG | GTTGTGCTCAATCTCCTCCT | 74 | NM_001004027.1 |
| NQO1 | GGACATCACAGGTAAACTGA | TATAAGCCAGAGCAGTCTCG | 68 | NM_001159613.1 |
| Occludin | TCAGGTGCACCCTCCAGATT | TGGACTTTCAAGAGGCCTGG | 112 | NM_001163647.2 |
| ZO-1 | CGATCACTCCAGCATACAAT | CACTTGGCAGAAGATTGTGA | 111 | CV870309 |
| Claudin 1 | CCTCAATACAGGAGGGAAGC | CTCTCCCCACATTCGAGATGATT | 76 | NM_001244539.1 |
Table 2.
Antibodies used in western blotting.
| Antibody | Source | Dilution | Company | Cat# |
|---|---|---|---|---|
| Nrf2 | Rabbit | 1:1000 | Abcam, Cambridge, UK | ab92946 |
| ZO-1 | Rabbit | 1:1000 | Thermo Fisher Scientific, Waltham, MA, USA | 61-7300 |
| Occludin | Rabbit | 1:1000 | Abcam, Cambridge, UK | ab31721 |
| GAPDH | Rabbit | 1:2000 | Cell Signaling Technology, Danvers, MA, USA | 2118 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Cai, L.; Bi, Q.; Sun, W.; Pi, Y.; Jiang, X.; Li, X. Genistein Alleviates Intestinal Oxidative Stress by Activating the Nrf2 Signaling Pathway in IPEC-J2 Cells. Vet. Sci. 2024, 11, 154. https://doi.org/10.3390/vetsci11040154
AMA Style
Li Y, Cai L, Bi Q, Sun W, Pi Y, Jiang X, Li X. Genistein Alleviates Intestinal Oxidative Stress by Activating the Nrf2 Signaling Pathway in IPEC-J2 Cells. Veterinary Sciences. 2024; 11(4):154. https://doi.org/10.3390/vetsci11040154
Chicago/Turabian StyleLi, Yanpin, Long Cai, Qingyue Bi, Wenjuan Sun, Yu Pi, Xianren Jiang, and Xilong Li. 2024. "Genistein Alleviates Intestinal Oxidative Stress by Activating the Nrf2 Signaling Pathway in IPEC-J2 Cells" Veterinary Sciences 11, no. 4: 154. https://doi.org/10.3390/vetsci11040154
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.