In Full Force. Mechanotransduction and Morphogenesis during Homeostasis and Tissue Regeneration


Developmental Biology, Clinical, Experimental Surgery and Translational Research Center, Biomedical Research Foundation Academy of Athens, 11527 Athens, Greece
*
Authors to whom correspondence should be addressed.
J. Cardiovasc. Dev. Dis. 2020, 7(4), 40; https://doi.org/10.3390/jcdd7040040
Submission received: 7 July 2020
/
Revised: 17 September 2020
/
Accepted: 25 September 2020
/
Published: 1 October 2020
(This article belongs to the Special Issue Comparative Developmental Cardiovascular Biomechanics and Bioengineering)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The interactions of form and function have been the focus of numerous studies in the context of development and more recently regeneration. Our understanding on how cells, tissues and organs sense and interpret external cues, such as mechanical forces, is becoming deeper as novel techniques in imaging are applied and the relevant signaling pathways emerge. These cellular responses can be found from bacteria to all multicellular organisms such as plants and animals. In this review, we focus on hemodynamic flow and endothelial shear stress during cardiovascular development and regeneration, where the interactions of morphogenesis and proper function are more prominent. In addition, we address the recent literature on the role of extracellular matrix and fibrotic response during tissue repair and regeneration. Finally, we refer to examples where the integration of multi-disciplinary approaches to understand the biomechanics of cellular responses could be utilized in novel medical applications.
1. Mechanical Stimuli to Guide Biological Processes
During the late 1800s, scientists primarily working on bone and/or orthopedics put forward the notion that an extracellular stimulus, such as mechanical load, can instruct a specific tissue response, such as trabecular bone adaptation and remodeling [1,2,3]. We currently know that the series of events in which a physical force is converted into a cellular response, able to induce in turn a biological process, are regulated and described by the conserved principles of cellular mechanics and mechanotransduction. From bacteria that sense osmotic pressure changes through their membranes and adjust their cytoplasm [4], to plant stems that adapt their growth patterns in response to wind and rain, touching of passing animals [5] or gravity [6], to fracture healing after bone injury [7], all living organisms use stimuli originating from their environment to constantly adjust their behavior, respond to their surroundings and ensure survival.
With forces applied to a given cell from seconds to hours, ranging from pico- to nano- newtons and external stimuli spanning changes in i) the stiffness of extracellular matrix (ECM), ii) air and osmotic pressure, iii) compression, contraction, and stretch, iv) shear stress and v) fluid flow, cells respond to stimuli by adapting their cell function and, therefore, key cellular processes such as gene transcription, protein synthesis, proliferation, migration, differentiation and/or apoptosis. This renders mechanotransduction crucial to development, homeostasis, disease progression and regeneration (Figure 1).
In this review, we provide an overview of the basic principles that allow cells to sense and integrate mechanical cues and discuss how mechanotransduction shapes morphogenetic events during animal and tissue development with a specific focus on cardiogenesis. Furthermore, we summarize how different types of biomechanical signaling orchestrate tissue remodeling during homeostasis and regeneration.
2. Sensing and Integrating Mechanical Forces
Mechanotransduction is a complex, multi-step reaction cascade. The first steps involve mechanotransmission and mechanosensing: the transmission of an external, mechanical cue to a mechanosensitive cell component and its subsequent local, active perception by the sensor cell. During the next step, described as mechanocoupling, stimuli surpassing a certain threshold are transduced into an intracellular change; this might be a biochemical signal and/or electrochemical activity and is, therefore, referred to as biochemical coupling. The activation of such a signaling cascade induces the mechanoresponse of the sensor cell, which is eventually transferred to an effector cell and initiates its response.
So how does a mechanical stimulus transform into a subcellular, molecular signal with such high efficiency? Cells sense, control and interpret external cues, whether they come from the environment or a neighboring cell, by integrating changes in surface parameters through their membranes. Due to its lipid bilayer composition, besides its role as a physical barrier that selectively allows the movement of ions and molecules in and out of cells, the cell membrane provides an excellent dock for a variety of mechanosensitive molecules. A plethora of different biological structures, such as ion or cell–cell junctional channels and transmembrane receptors, have been suggested as mechanosensors and can be found in many different cell types. The integration of external cues originating from the ECM occurs through highly specific cell–matrix interactions. These take place in specialized sites, where cell membranes harbor the ECM in structures of specific molecular composition [8], named focal adhesions or focal contacts [9]. In turn, these form multi-protein complexes [10] that connect the extracellular space to the cell’s interior and the cytoskeleton, through transmembrane receptors such as those of the integrin family [11,12]. The extracellular part of the integrin receptor anchors defined ECM proteins, like various proteoglycans [13,14], while the cytoplasmic tail interacts with proteins of the cytoskeletal network, such as actin, myosin, tubulin, and paxillin [15]. The latter can further act as a scaffold and recruit other structural and signaling elements like vinculin [16,17] and/or Focal Adhesion Kinase (FAK) [18,19], one of many protein tyrosine kinases (PTKs), respectively. This creates a highly dynamic and tightly regulated complex that effectively creates a physical continuity between the extracellular space and the intracellular environment.
Conformational changes are the predominant mechanism through which these protein interactions can activate intracellular signaling cascades, mediating in turn the transmission of mechanical information into biological responses. For example, spatial rearrangements affect the folding landscape of macromolecules, the accessibility of their molecular recognition sites [20] and, therefore, their function. Different signaling pathways have been recognized as early effectors of mechanotransduction, with tyrosine kinase activities and small GTPases playing a crucial role in the regulation of the matrix–integrin–cytoskeleton complexes [21,22]. Once a tyrosine kinase, like FAK, is recruited to the focal adhesion complex, phosphorylation on its tyrosine residues modulates its catalytic activity [23]. This directs the creation of a high-affinity binding site for additional PTKs, such as those of the Src-family of protein tyrosine kinases (SFKs) and, thereby, of a kinase complex that can in turn regulate the activation of downstream signal transduction pathways, such as Ras [24,25,26]. Proteins of the Ras signaling pathway, also known as the Ras/Raf/Mitogen-activated protein kinase/ERK kinase (MEK)/extracellular-signal-regulated kinase (ERK), can act as the connecting elements between cellular compartments [27]. The solidity of this macromolecule complex is further strengthened by membrane protein-adaptors, such as caveolin-1, that connect integrins to the tyrosine kinases and provide a scaffold that ensures the efficient recruitment and separation of the appropriate signaling molecules [28,29]. Hence, mutations on tyrosine residues greatly affect the activation and subsequent stability of the complex, and, therefore, the eventual translocation of the initial stimulus to the nucleus (Figure 2).
2.1. From the Cell Surface to the Core
Signal propagation depends on the transport of the initial stimulus from the cell membrane to the DNA, in a chain-like reaction, such that information originating from cell surface-tethered, transmembrane receptors will be coupled to transcription factors that regulate and potentially alter gene expression. As discussed in the previous section, the transmission of information involves the propagation of biochemical signals through intracellular signaling components and secondary messengers. As signals need to be coupled to gene expression changes, the endpoint of the signaling cascade is the nucleus, where the translocation of certain co-factors activated in the pathway will activate the appropriate transcription factors. Alternatively, biochemical signaling can occur with the direct movement of transcription factors from the cell surface or cytoplasm to the nucleus. This is the case for the mechanosensitive proteins paxillin, β-catenin, and Notch and the cytoplasm-sequestered transcription nuclear factor kappa-light-chain-enhancer of activated B cells (NFkB), signal transducer and activator of transcription 3 (STAT3) and nuclear factor of activated T-cells (NFAT). Another option, would be the activation of a transcription factor shuttling host. Some of the most described mediators are the nuclear transducers Yes-associated Protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) [30]. YAP/TAZ dynamically respond to increasing applied forces by rapidly adjusting their nuclear to cytoplasmic localization [31]. As signals need to travel from the margins to the inner core of the cell either by diffusion or active transport, transmission of biochemical signals can range from seconds to minutes or even hours.
In the case of minimal physical forces, signal transduction can occur through the direct physical connections between the membrane, the cytoskeleton and the nucleus. In contrast, signals bearing a great amount of mechanical tension, require the remodeling of the cytoskeleton and the reorganization of perinuclear actin network, thus, allowing changes to nuclear shape and morphology. The magnitude of the force and the extent of cytoskeletal reorganization needed until the signal reaches the nucleus, defines the timescale of propagation which, in this mode, can reach sub-millisecond levels. The mechanical coupling and physical association of the cytoskeleton to the nucleus occurs through the Linker of Nucleoskeleton and Cytoskeleton (LINC) complex [32]. This comprises a protein network that not only ensures nuclear positioning and provides stability but also links the nuclear envelope to the cytoskeleton and the nuclear lamina [33]. Coupling connections occur through nesprins, proteins of the outer nuclear membrane [34,35] that bridge through their interactions the extracellular space to the nucleus and, subsequently, the genome. Towards the inner side of the nucleus, nesprins bind to SUN proteins, that are located in the inner nuclear membrane and interact with nuclear intermediate filaments, lamins [36,37]; towards the outer side, nesprins associate with actin and microtubule filaments [38]. As lamins anchor chromatin domains and actively take part in the assembly and organization of DNA machinery [39,40], this highly organized complex has the ability to instruct changes in the gene expression pattern of a given cell (Figure 3).
2.2. Here Comes the Nucleus
Mechanical stimuli reach the nucleus and become effective by means of transcription factors. These bind to regulatory DNA regions, enhancers or promoter sites, and control the up- or down- regulation of given genes. The spatial organization of chromosomes plays a crucial role in how such signals can guide transcriptional regulation and orchestrate nuclear mechanotransduction [41]. The density and compaction of the double-stranded DNA, represent a major obstacle that limits the accessibility of nuclear elements to such interactions. Recent accumulating evidence has shown that this is an active process. The nucleus, a mechanosensitive organelle itself, does not only respond to the downstream effectors of mechanical cues, but also to applied forces directly [42,43,44].
So how do transcription factors find their way through the highly packed chromatin landscape of the nucleus? The 3D organization of chromosomes plays a crucial role in the regulation of gene expression. To circumvent accessibility restraints placed on transcription factors by the highly dense, super-coiled DNA, it has been shown that chromosomal territories, representing sites of active transcription of co-regulated, functionally-related genes, are organized in a spatially defined, neighboring manner [45,46]. This chromosome intermingling allows the identification of distinct “mechanical hotspots for transcription” [47] and is strongly correlated with the common and efficient transcriptional control of functionally-clustered genes [48,49,50], as well as the maintenance of transcriptional memory to dividing cells.
3. Mechanical Forces Regulating Embryo and Tissue Development
The conversion of mechanical signals into gene expression regulation directs a plethora of fundamental biological processes that shape both embryonic development and adult tissue homeostasis. From the maturation of the ovary and throughout gametogenesis [51,52] to egg activation [53,54,55] and early asymmetric cell divisions, forces are generated and guide developmental patterning and tissue morphogenesis across species. In Arabidopsis, the expression of the homeobox gene SHOOT MERISTEMLESS (STM), key determinant of meristematic identity, is correlated to the curvature of the shoot apical meristem and induced by mechanical stress applied on the boundary domain [56], while the fertilization of the horseshoe crab (Limulus Polyphemus) egg requires the activation of the egg upon a spring force which is stored and applied by the sperm acrosome. The extension of an actin-based bundle [57], allows the sperm to penetrate the vitelline layer and triggers the cytoplasmic release of Ca2+ that powers fertilization [58]. Similarly, combined forces that occur by movements of the spindle pole and microtubule bending, contribute to the early asymmetric cell divisions shaping the Caenorhabditis elegans embryo [59], while maturation of the hemidesmosome structure into a junction that will later on direct the formation of body-wall muscle and the epidermis, occurs via the activation of a mechanotransduction pathway operating on a Rac GTPase and the phosphorylation of intermediate filaments [60]. In Drosophila, the stiffening of cells due to forces generated by myosin II contraction and exerted between neighbor cell relationships, i) sets the boundaries between different cellular/tissue compartments [61], ii) directs the patterning of anterior–posterior axis during germ band extension and formation of the embryonic thorax and abdomen [62,63], and iii) drives the sealing of the dorsal epidermis during dorsal closure [64]. Moreover, during Xenopus gastrulation, involution of the mesoderm requires the tension-dependent assembly of a fibronectin-based extracellular matrix. This guides the collective migration of cells towards the posterior side during convergence, a series of cellular movements that will eventually narrow and extend the embryo along the mediolateral body axis. Intriguingly, the migration of cells depends on their cadherin-mediated cell–cell contacts and can only take place when the mesoderm and the notochord remain stiff to prevent tissue collapsing or deformation [65,66]. Along the same lines, the mechanosensitive phosphorylation of β-catenin–tyrosine-667 and its subsequent nuclear translocation at the onset of gastrulation are essential for the expression of notail, a mesoderm-inducing gene in the zebrafish embryo [67], while regulation of actomyosin-dependent cell-cortex tension by Nodal and transforming growth factor β (TGFβ)-signaling drives progenitor-cell sorting and the organization of germ-layer formation [68]. Similarly, in the developing mouse embryo, the contractility-dependent localization of Yap drives positioning and lineage specification of blastomeres during formation of the inner cell mass (ICM) and the trophoectoderm [69], very much in the same way as human embryonic stem cells (hESCs) self-organize to generate all three germ layers under geometric confinement and in response to bone morphogenetic protein 4 (BMP4) [70].
Taken together, the paradigms discussed above highlight how physical and mechanical stimuli orchestrate embryogenesis and tissue patterning across species. Intriguingly, they illustrate how the continuity between cellular components like the membrane, the cytoskeleton and the nuclear envelope, allows the transduction of the stimuli to the effector endpoint, the DNA, and the transformation of the initial force into gene expression regulation. In fact, the ability of cells to sense and respond to mechanical forces, while controlling their mechanical properties, is highly dependent on this exact cellular architecture, described first in architectural and now also biological terms, by tensegrity (tensional integrity) [71,72,73]. This model views the complex cellular cytoskeletal network as an architectural structure in which stability is ensured by the tensile forces that its interacting components: the ECM, the focal adhesions, the filaments and the microtubules generate and oppose each other to achieve cellular balance and stability. This tensional prestress in which cells are found, allows them to not only control their shape and structure but also channel incoming forces into regulation of cell and whole-tissue behavior.
3.1. The (Mechano)Sensitive Heart
All organs are sensitive to mechanical signals. In vertebrates, mechanotransduction occurs in all tissues such as the central nervous system (CNS) [74] and the eyes [75], the inner ear [76] and the skin [77], the muscles [78], the respiratory [79] and intestinal tract [80], the kidneys [81], the liver [82], the vasculature [83] and the hematopoietic system [84]. The heart however is a paradigm where form and function can be best studied. Particularly, in the context of how mechanical forces shape cardiac development and morphogenesis [85].
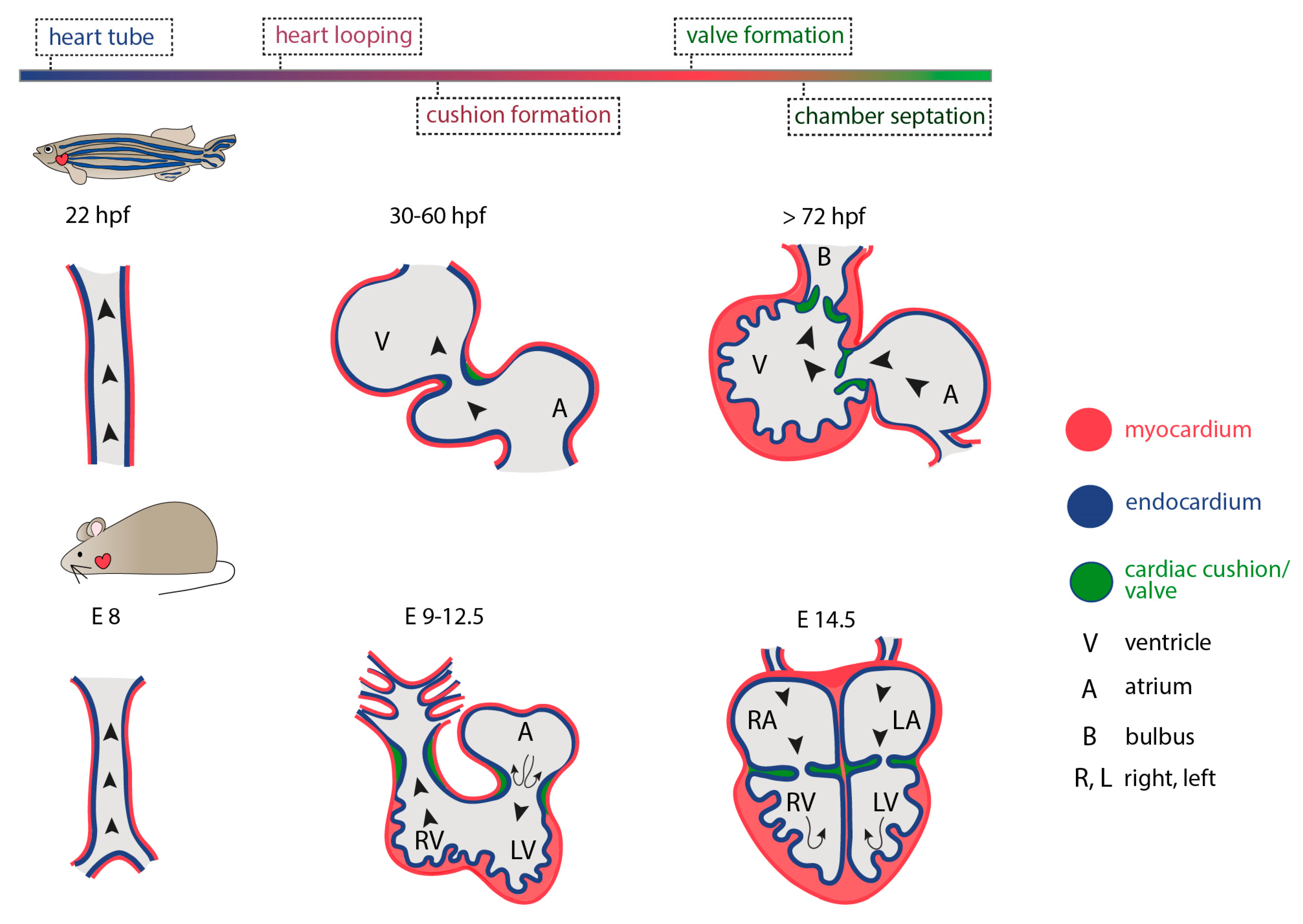
The heart is the first functional organ to develop during vertebrate embryogenesis. In humans, cardiogenesis begins very early, at around 18 to 19 days post fertilization with the initial formation of two cardiogenic regions arising from the lateral splanchnic mesoderm, one on either side of the neural plate, at the anterior end of the early embryo. As folding occurs, these two primary heart fields will transit from two oval-shaped structures to two endocardial tubes that will eventually converge towards each other and fuse together at the embryonic midline, forming the primitive heart. These initial steps of cardiac development involve conserved signaling pathways across vertebrate species. However, the timing of events differs significantly between them. In zebrafish, for example, the first cardiac contraction occurs 24 h post fertilization (hpf), while more than 8 and 24 days are required in mice and humans, respectively (reviewed in [86]). The cardiac tube, initially valve-less [87], will quickly bend, rotate and invert its spatial organization (cardiac looping) while differentiating into more complex and distinct parts. As the newly developed heart is required for meeting the circulatory oxygen and nutrient demands of all other developing embryonic tissues and organs, including the vasculature, peristaltic contraction begins during early looping events. Although it has been proposed that initiation of heart contraction is not coupled to the nutritional support of other developing tissues, rather than merely facilitating cardiac maturation and angiogenesis [88], the entity of morphogenetic events taking place throughout cardiogenesis are intermittently interwoven to hemodynamic changes. The latter have been shown to regulate cardiac cell proliferation, differentiation and growth, in close synergy to genetic factors [89].
3.2. Fluid Dynamics and the Role of Endocardium
As cardiac looping and remodeling evolve, the heart obtains its three-dimensional structure and becomes a curvature-based organ. Concomitantly, the vascular walls begin to accommodate an elevated degree of shear stress due to blood flow. Both the primitive heart tube as well as the mature cardiovascular system are lined by the endocardium, an inner layer of endothelial cells that comes in direct contact with the vascular wall and the blood flow. Hence, the endothelium acts as the sensor and signal transducer of the biomechanical stimuli generated by flow, including shear stress, stretch strain and hydrostatic pressure [90,91]. Interestingly, an increase in the amount of fluid shear stress placed on cultured monolayers of bovine endothelial cells has been shown to activate a K+ selective, shear-stress activated ionic current [92]. Moreover, this results in time-dependent marked changes in (i) cell shape and cytoskeletal assembly, (ii) basic endothelial functions such as fluid endocytosis [93] and (iii) differential regulation of endothelial-derived factors like endothelin-1 mRNA [94]; indicating that fluid mechanical forces can directly influence endothelial cell physiology. It has been shown that endothelial cells sense fluid dynamics through a mechanosensory complex involving (i) the immunoglobulin family receptor platelet endothelial cell adhesion molecule (PECAM)-1 that transmits the mechanical force through Src activation, (ii) the vascular endothelial cell cadherin (VE-cadherin) that acts as an adaptor element binding directly to the transmembrane domain of VEGFR2 [95] and (iii) the vascular endothelial growth factor receptor 2 (VEGFR2) that subsequently activates phosphatidylinositol-3-OH kinase (PI(3)K) [96]. Activation of the complex triggers the conformational activation of integrin αvβ3 which mediates its binding to ECM proteins and the cytoskeletal remodeling and alignment of endothelial cells in the direction of flow [97,98]. Interestingly, a certain threshold/set point of fluid stress is needed to activate the transduction cascade [99], indicating a fine balance between the cues that a cell receives and the output responses these will generate. Moreover, Piezo1 (Fam38a) channels have been also shown to act as integrators and sensors of frictional forces arising from fluid flow. When negative pressure was used to deliver a physical force in cell-attached membrane patches of mouse embryonic endothelial cells, unitary single channel events (with ionic characteristics representative of Piezo channels) were detected within less than 1s. Importantly, Piezo1 channel activation was coupled to elevated Ca2+ influx and the downstream activation of calpain-2 [100], a cysteine protease known to control the turnover of focal adhesion anchorage [101]. Piezo is also shown to regulate Kruppel-like factor (klf)2 activity in the endothelium as well as Yap1 localization in the smooth muscle cells in the context of zebrafish outflow tract development, providing an example of how mechanotransduction can occur between cell types during valvulogenesis [102]. Overall, these data suggest a strong correlation between hemodynamics and endothelial cell responses and argue in favor of an imminent role for mechanical forces and blood flow pattern in shaping cardiac development and homeostasis.
So far, a great amount of in vitro studies has elegantly shown how endothelial cell behavior is regulated by mechanical cues. Yet, there is an inherent difficulty in mapping intracardial flow dynamics in vivo. In the last few decades, the relevance of shear forces in cardiac function in vivo, has been greatly advanced with the use of the zebrafish (Danio rerio) as a model organism. Due to their (i) external fertilization, (ii) small size, (iii) high degree of conservation with the human genome [103] and (iv) their amenability to forward and reverse genetic approaches [51,104], zebrafish have emerged as an ideal model organism to study the cellular and molecular events orchestrating cardiogenesis in vivo [105]. Furthermore, because of their small size, zebrafish embryos receive sufficient oxygen by passive diffusion which, in contrast to other model organisms, allows them to develop normally even in the total absence of active blood circulation [106]. Consequently, the cellular and molecular basis of cardiovascular defects in which blood flow is perturbed, can be studied in great detail [107]. Finally, due to their high ability to regenerate many tissues [108], including their heart [109], zebrafish are widely used to unravel the mechanisms governing tissue repair and functional recovery in a regeneration-competent vertebrate.
3.3. What Can Fish Teach Us about Cardiogenesis
Together with their small size, a great advantage of zebrafish embryos for the investigation of cardiac morphogenesis, is their optical clarity that allows the in vivo visualization of intracardiac dynamics in the developing beating heart, in living animals. Moreover, understanding on how hemodynamics shape cardiac development, was also greatly advanced by the development of selective plane illumination microscopy (SPIM) [110,111]. The efficient optical sectioning of internal cardiac structures deep into the tissue and the generation of multidimensional images, enabled the fast, non-invasive and long-term imaging of live animals at single-cell resolution [112,113].
Indeed, high-speed imaging and digital particle image velocimetry (DPIV), revealed an extremely elevated wall shear stress, as a result of the highly viscous flow, and identified the presence of vortices in the circulation pattern during the development of the embryonic zebrafish heart [114]. Interestingly, interference with blood flow resulted in severe cardiac phenotypes that comprised the absence of cardiac looping and bulbus formation as well as defects in the development of inflow and outflow tracts [114]. Blood flow and shear stress promote embryonic haematopoiesis [115] and epithelial–mesenchymal transition (EMT) in the context of the tumor microenvironment by downregulating ERK and glycogen synthase kinase 3 beta (GSK3β) [116]. Zebrafish mutants and manipulations of circulation in vivo during embryogenesis showed that, in the absence of intracardiac sheer-stress, atrioventricular endocardial cells remain quiescent and fail to undergo a mesenchymal transition and become interstitial, resulting in cardiac valve defects [109,117]. Similarly, blood flow was shown to regulate the formation of the developing ventricle by affecting cardiomyocyte morphology. By utilizing a zebrafish weak atrium (wea) mutant, in which the lack of atrial sarcomeres results in impaired atrial function and decreased blood flow, Auman et al. showed that the reduction of circulation impairs cardiomyocyte elongation and enlargement, thus, impeding the formation of ventricle curvature, a key determinant to a mature and functional heart [118]. These results were strongly corroborated by Kalogirou et al., in which a wea homozygous mutant could be raised and investigated to adulthood [117]. The adult heart exhibited only one functional chamber, the ventricle, which was markedly enlarged, while the pattern of transvalvular flow through the atrioventricular (AV) valve was also altered. Interestingly, the AV valve presented fewer cells compared to controls and failed to mature from two to four cuspids, indicating the strong coupling of intracardiac flow dynamics to cardiac chamber formation and also valve morphogenesis (reviewed in [119]).
Furthermore, optical mapping of the silent heart (sihb109) zebrafish mutant, that fails to contract due to a mutation in the cardiac troponin T (tnnt2) gene and exhibits cardiac conduction defects, identified loss of contraction and also blood flow as the possible etiology behind the downregulation of Connexin-40 (Cx-40) and hence, an impairment in trabeculae formation [120]. Interestingly, the endocardium-specific, blood flow-dependent activation of Notch1 signaling and its downstream effectors ephrin b2a (efnb2a) and neuregulin 1 (nrg1) were also shown to direct trabeculation in the developing zebrafish heart [121,122]. Specifically, these data also demonstrated that detection of fluid shear stress by primary cilia is required for the response of endocardial cells to cardiac contraction.
A crucial step in cardiac development, is the formation of heart valves. As cardiac output is necessary to drive blood circulation both during development and adult stages, the precise formation of heart valves that prevent retrograde blood flow, plays a crucial role in a functional heart. During early cardiac development, valve leaflets originate from AV canal endocardial cells that experience high shear stress forces as a result of blood circulation and the presence of reversing-oscillatory flow patterns between the atrium and the ventricle [123,124]. To examine the relationship between hemodynamics and valve development, Vermot et al. manipulated blood flow by interfering with hematopoiesis and heart rate [125]. By altering blood flow patterns, authors identified a transcription factor from the Kruppel-like factor (Klf) family, klf2a, and confirmed its expression in valve precursors in response to oscillating fluid flows, thereby indicating its involvement in early events of valvulogenesis. It is now shown, that Trpv4, a mechanosensitive ion channel that is specifically expressed in the endocardium, regulates the release of endocardial Ca2+ and the expression of the flow-responsive klf2a promoter, while its absence results in prominent valve impairments [126]. This argues in favor of a prominent mechanotransduction pathway as key determinant to zebrafish valve formation. In 2013, Banjo et al. showed that the signaling cascade underlying the transmission of physical forces through klf2a activation during valve formation, is mediated by miRNAs. The authors pondered on previous studies demonstrating the contribution and involvement of miRNAs to fluid flow-dependent angiogenesis in zebrafish [127] and postulated that miR-21 microRNA directs valvulogenesis in the developing zebrafish. Indeed, they showed that mir-21 morphants do not develop heart valves. Further they demonstrated that flow-dependent expression of mir-21 controls cell proliferation of the valve-forming endocardium via inhibition of sprouty2 (spry2), programmed cell death 4b (pdcd4) and phosphatase and tensin homologue B (ptenb) targets, through the removal of suppression on the RTK/Ras/ERK pathway. Intriguingly, miR-21 was not essential to the stress-induced cardiomyocyte and/or fibroblast remodeling occurring in the mammalian heart [128], suggesting a potentially different mechanism through which microRNA-21 might act in different tissue and species contexts. Taken together, a long line of emerging evidence demonstrates that both morphogenesis and function of the developing and adult zebrafish heart are coupled to mechanical forces that are generated from blood circulation and can regulate the genetic basis of cell behavior within the cardiac tissue (Figure 4). But how do (cardiac) cells adjust their behavior in the absence of mechanical stimuli to maintain tissue homeostasis and remain quiescent?
4. Quiescence, Mechanical Stiffness and Tissue Regeneration
In zebrafish silent heart mutants, a basic embryonic angiogenic pattern is laid down even in the total absence of blood flow. However, endothelial cells remain quiescent and they do not form a proper lumen. Renz et al. suggested that in the zebrafish embryo, endothelial quiescence in the absence of blood flow-induced mechanical forces is safeguarded by cerebral cavernous malformation (CCM) proteins that act upstream of the Klf2 signaling pathway [129]. Using ccm mutants, the authors showed that CCM proteins, which are involved in cell–cell junctions and adhesion of endothelial cells to the extracellular matrix, prevent angiogenic overgrowth by negatively regulating the β1 integrin-dependent overexpression of klf2 and the consequent upregulation of epidermal growth factor-like domain 7 gene (egfl7) [130]. The latter, a secreted protein that is expressed by endothelial cells and associated with the blood vessel extracellular matrix [131]. CCM proteins are crucial for the mechanosensitive properties of the endocardium during valve development [132]. Intriguingly, restoring blood flow in a zebrafish model of CCM1 has been shown to suppress its pathological phenotypes and prevent cardiovascular anomalies [133]. For a detailed review on how blood flow affects vascular development, one of the paradigms to study the interactions of form and function we encourage the readers to also consider [134]. Such mechanotransduction feedback loops are particularly relevant in the context of tissue repair and regeneration, when cells need to rapidly adjust their properties and transition from quiescence to proliferating and differentiating states to ensure survival and functional recovery of the injured tissue.
In the adult mammalian body, a variety of tissues such as the epidermis, the blood or the intestine are able to regenerate even on a daily basis, while other tissues, mainly quiescent, like the lungs or the liver can rapidly respond in cases of injury or cellular loss and replace the lost part [135]. On the contrary, the adult heart and the adult CNS, consisting of the brain and the spinal cord, have a very low regenerative capacity [136,137,138]. Interestingly, it has been shown that cells of the CNS are able to regenerate when in the periphery [139]. This suggests that a combination of post-injury events renders the mammalian CNS (and potentially other tissues) as non-permissive to regeneration. It has been shown that the experimental activation of the YAP/TAZ signaling pathway in rodents can promote regeneration in organs with limited regenerative capacity [140], suggesting a prominent role of mechanotransduction in the regeneration of different adult tissues. YAP/TAZ integrates multiple signaling pathways and is activated by the Hippo pathway as well as biomechanical forces through a yet unidentified mediator/sensor. The downstream activators and regulation in regeneration as well as cancer progression appear to be cell and tissue specific (reviewed in [141]).
Both in the adult mammalian CNS and the heart, the formation of a fibrotic scar after injury is one of the key events impeding functional regeneration, representing a chemical but also physical barrier to cell regrowth [142,143]. In zebrafish, cardiac fibroblasts are transiently activated following cardiac injury to contribute ECM components to the injury site [144]. In addition, macrophages have been reported to contribute collagen in a cell-autonomous manner at the heart injury site, both in zebrafish and mice [145]. Modulating the time-window of fibrotic response, its levels and the ECM components contributing to the scar formation might hold some answers to the regenerative potential differences between organs as well as species [146]. Moreover, similarly to cardiogenesis, local substrate stiffness critically directs axon pathfinding during CNS development [74]. Hence, certain analogies can be drawn for the two systems. Intriguingly, using atomic force microscopy in the injured rat CNS, Moeendarbary et al. have shown that, contrary to other tissues, CNS significantly softens after injury, while alteration of tissue elastical properties correlates to the expression levels of scar elements like intermediate filaments and extracellular matrix components [147]. Further, in vitro data have shown that neuron branching decreases in response to increased substrate stiffness [148] while substrate stiffness over the same range, results in higher adherent area of astrocytes, key players of the fibrous scar [149]. Alterations to substrate stiffness were also observed upon injury in the regenerating zebrafish spinal cord [150]. Using AFM-enabled nano-identation to determine elastic moduli on living spinal cord sections of control and injured samples, the authors provided the first quantitative mapping of spinal cord tissue stiffness upon injury. Intriguingly, the authors showed that in contrast to mammals, spinal cord tissue transiently stiffened during regeneration, suggesting a contradictory role of environmental mechanical cues in the regeneration-permissive environment of zebrafish. Interestingly, the mechanical differences observed in the regenerating zebrafish spinal cord correlated with changes in tissue architecture and specifically cell number densities, axonal orientation and vascularization. Taken together, these studies suggest that the finely tuned interaction between cells and their environment defines important cellular functions and argues in favor of a key role of extracellular physical properties in promoting versus inhibiting regeneration. This information could be of great importance in the identification of novel targets to be used in new therapeutic interventions.
5. Perspectives: Biomaterials and Tissue Engineering to Promote Tissue Regeneration
Regeneration describes the regrowth and replacement of a lost cell, tissue, organ or body part, such that the full restoration of its previous biological function is achieved. It has been an attractive field of interest for scientists already from the 1700′s when the first scientific studies on crayfish, amphibian and hydra regeneration were described [151]. A recurrent theme in tissue regeneration is the re-activation of crucial developmental signaling pathways as shown with an inducible cardiac valve injury model [152,153]. Following cardiac valve injury, the intracardiac flow pattern is reminiscent of the immature, embryonic one and results in the induction of developmental pathways such as Notch [152]. In addition, cell cycle re-entry of valve endocardial cells and ECM production following decellularization of cardiac valves is dependent on TGF-β activation, revealing a pro-regenerative role for TGF signaling [153]. Yet, to date, the basic mechanisms governing tissue regeneration are still being explored and the extent to which these can be expanded in non-regenerating environments/tissues during adulthood remains unknown. Besides the presence of a regeneration-permissive environment, successful tissue regeneration requires either the activation of endogenous stem cells or the transplantation of stem and/or progenitor cells that will replace and repair the injured tissue. However, conventional in vitro methods used so far often lack the level of complexity needed to fully recapitulate the complex ECM network which cells would physically encounter in vivo. The field of biomaterial science and tissue engineering is furthering our understanding on how extracellular physical properties direct stem cell homeostatic and injury responses such as quiescence, proliferation and differentiation. In silico models to predict the behavior of tissue-engineered valves has been already successfully integrated in their design [154]. In addition, the use of porous collagen-based scaffolds is shown to enhance the regeneration in experimental animal models of spinal cord injury [155]. All these advances, coming from cross-disciplinary approaches, unravel the complex cell–matrix interactions we need to understand, in order to achieve optimal regeneration. Specifically, the emergence of novel cultured conditions comprising hydrogels with tunable stress properties, 2D and/or 3D synthetic matrices, microfluidic chambers and cellular scaffolds, allows the precise and timely control of the biomechanical properties of the stem cell niche microenvironment and, hence, of potential external cues determining cell lineage fate (also reviewed in [156]). Such approaches can unravel several of the necessary elements (cells, secreted factors, extracellular components) as well as the mechanical properties required for successful tissue repair in “difficult” to regenerate cellular environments.
Funding
DB acknowledges support from the Fondation Sante and VT from the Greek State Scholarships Foundation (ΙΚΥ) (co-financed by Greece and the European Union (European Social Fund—ESF) through the Operational Programme «Human Resources Development, Education and Lifelong Learning» in the context of the project “Reinforcement of Postdoctoral Researchersf—2nd Cycle” (MIS-5033021).
Acknowledgments
We apologize to all colleagues whose work could not be cited due to length limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wolff, J. The classic: On the theory of fracture healing. Clin. Orthop. Relat. Res. 2010, 468, 1052–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, J. Das Gesetz der Transformation der Knochen. DMW - Dtsch. Med. Wochenschr. 1893, 19, 1222–1224. [Google Scholar] [CrossRef] [Green Version]
- Duncan, R.L.; Turner, C.H. Mechanotransduction and the functional response of bone to mechanical strain. Calcif. Tissue Int. 1995, 57, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Persat, A. Bacterial mechanotransduction. Curr. Opin. Microbiol. 2017, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, M.J. Thigmomorphogenesis: The response of plant growth and development to mechanical stimulation. Planta 1973, 114, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Kramer, E.M.; Perry, P.; Knox, K.A.; Leyser, O.; Haseloff, J.; Beemster, G.T.S.; Bhalerao, R.; Bennett, M. Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nature 2005, 7, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ogawa, R. Mechanotransduction in bone repair and regeneration. FASEB J. 2010, 24, 3625–3632. [Google Scholar] [CrossRef]
- Jalali, S.; del Pozo, M.A.; Chen, K.; Miao, H.; Li, Y.; Schwartz, M.A.; Shyy, J.Y.; Chien, S. Integrin-mediated mechanotransduction requires its dynamic interaction with specific extracellular matrix (ECM) ligands. Proc. Natl. Acad. Sci. USA 2001, 98, 1042–1046. [Google Scholar] [CrossRef]
- Geiger, B.; Spatz, J.P.; Bershadsky, A.D. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell Biol. 2009, 10, 21–33. [Google Scholar] [CrossRef]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’Ayan, A.; Iyengar, R.; Geiger, B. Functional atlas of the integrin adhesome. Nature 2007, 9, 858–867. [Google Scholar] [CrossRef]
- Schwartz, M.A.; DeSimone, D.W. Cell adhesion receptors in mechanotransduction. Curr. Opin. Cell Biol. 2008, 20, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bershadsky, A.D.; Balaban, N.Q.; Geiger, B. Adhesion-dependent cell mechanosensitivity. Annu. Rev. Cell Dev. Biol. 2003, 19, 677–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burridge, K.; Chrzanowska-Wodnicka, M. Focal adhesions, contractility, and signaling. Annu. Rev. Cell Dev. Biol. 1996, 12, 463–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saoncella, S.; Echtermeyer, F.; Denhez, F.; Nowlen, J.K.; Mosher, D.F.; Robinson, S.D.; Hynes, R.O.; Goetinck, P.F. Syndecan-4 signals cooperatively with integrins in a Rhodependent manner in the assembly of focal adhesions and actin stress fibers. Proc. Natl. Acad. Sci. 1999, 96, 2805–2810. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.E. Molecules in focus Paxillin. Int. J. Biochem. Cell Biol. 1998, 30, 955–959. [Google Scholar] [CrossRef]
- Turner, C.E.; Glenney, J.R.; Burridge, K. Paxillin: A new vinculin-binding protein present in focal adhesions. J. Cell Biol. 1990, 111, 1059–1068. [Google Scholar] [CrossRef]
- Brown, M.C.; Perrotta, J.; Turner, C.E. Identification of LIM3 as the principal determinant of paxillin focal adhesion localization and characterization of a novel motif on paxillin directing vinculin and focal adhesion kinase binding. J. Cell Biol. 1996, 135, 1109–1123. [Google Scholar] [CrossRef] [Green Version]
- Parsons, J.T.; Martin, K.H.; Slack, J.K.; Taylor, J.M.; Weed, S.A. Focal Adhesion Kinase: A regulator of focal adhesion dynamics and cell movement. Oncogene 2000, 19, 5606–5613. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.; Brugge, J. Integrins and signal transduction pathways: The road taken. Science 1995, 268, 233–239. [Google Scholar] [CrossRef]
- Krammer, A.; Lu, H.; Isralewitz, B.; Schulten, K.; Vogel, V. Forced unfolding of the fibronectin type III module reveals a tensile molecular recognition switch. Proc. Natl. Acad. Sci. USA 1999, 96, 1351–1356. [Google Scholar] [CrossRef] [Green Version]
- Barry, S.T.; Critchley, D.R. The RhoA-dependent assembly of focal adhesions in Swiss 3T3 cells is associated with increased tyrosine phosphorylation and the recruitment of both pp125FAK and protein kinase C-delta to focal adhesions. J. Cell Sci. 1994, 107, 2033–2045. [Google Scholar] [PubMed]
- Hall, A. Rho GTPases and the Actin Cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calalb, M.B.; Polte, T.R.; Hanks, S.K. Tyrosine phosphorylation of focal adhesion kinase at sites in the catalytic domain regulates kinase activity: A role for Src family kinases. Mol. Cell. Biol. 1995, 15, 954–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Chen, H.C.; Nowlen, J.K.; Taylor, S.J.; Shalloway, D.; Guan, J.L. Direct interaction of v-Src with the focal adhesion kinase mediated by the Src SH2 domain. Mol. Biol. Cell 1994, 5, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Schlaepfer, D.D.; Hunter, T. Evidence for in vivo phosphorylation of the Grb2 SH2-domain binding site on focal adhesion kinase by Src-family protein-tyrosine kinases. Mol. Cell. Biol. 1996, 16, 5623–5633. [Google Scholar] [CrossRef] [Green Version]
- Schlaepfer, D.D.; Hanks, S.K.; Hunter, T.; Van Der Geer, P. Integrin-mediated signal transduction linked to Ras pathway by GRB2 binding to focal adhesion kinase. Nature 1994, 372, 786–791. [Google Scholar] [CrossRef]
- Chang, F.; Steelman, L.S.; Lee, J.T.; Shelton, J.G.; Navolanic, P.M.; Blalock, W.; Franklin, R.A.; McCubrey, J.A. Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: Potential targeting for therapeutic intervention. Leukemia 2003, 17, 1263–1293. [Google Scholar] [CrossRef]
- Wary, K.K.; Mariotti, A.; Zurzolo, C.; Giancotti, F.G. A Requirement for Caveolin-1 and Associated Kinase Fyn in Integrin Signaling and Anchorage-Dependent Cell Growth. Cell 1998, 94, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Goetz, J.G.; Joshi, B.; Lajoie, P.; Strugnell, S.S.; Scudamore, T.; Kojic, L.D.; Nabi, I.R. Concerted regulation of focal adhesion dynamics by galectin-3 and tyrosine-phosphorylated caveolin-1. J. Cell Biol. 2008, 180, 1261–1275. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, M.L.; Jaalouk, D.E.; Shanahan, C.M.; Burke, B.; Roux, K.J.; Lammerding, J. The Interaction between Nesprins and Sun Proteins at the Nuclear Envelope Is Critical for Force Transmission between the Nucleus and Cytoskeleton. J. Biol. Chem. 2011, 286, 26743–26753. [Google Scholar] [CrossRef] [Green Version]
- Maniotis, A.J.; Chen, C.S.; Ingber, D.E. Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 1997, 94, 849–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Skepper, J.N.; Yang, F.; Davies, J.D.; Hegyi, L.; Roberts, R.G.; Weissberg, P.L.; Ellis, J.A.; Shanahan, C.M. Nesprins: A novel family of spectrin-repeat-containing proteins that localize to the nuclear membrane in multiple tissues. J. Cell Sci. 2001, 114, 4485–4498. [Google Scholar] [PubMed]
- Rajgor, D.; Shanahan, C.M. Nesprins: From the nuclear envelope and beyond. Expert Rev. Mol. Med. 2013, 15, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.L.; Roux, K.J.; Burke, B. Blurring the Boundary: The Nuclear Envelope Extends Its Reach. Science 2007, 318, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, T.; Kim, E.; Riehl, B.D.; Esfahani, A.M.; Rosenbohm, J.; Yang, R.; Duan, B.; Lim, J.Y. The LINC complex, mechanotransduction, and mesenchymal stem cell function and fate. J. Biol. Eng. 2019, 13, 1–12. [Google Scholar] [CrossRef]
- Roux, K.J.; Crisp, M.L.; Liu, Q.; Kim, D.; Kozlov, S.; Stewart, C.L.; Burke, B. Nesprin 4 is an outer nuclear membrane protein that can induce kinesin-mediated cell polarization. Proc. Natl. Acad. Sci. USA 2009, 106, 2194–2199. [Google Scholar] [CrossRef] [Green Version]
- De Leeuw, R.; Gruenbaum, Y.; Medalia, O. Nuclear Lamins: Thin Filaments with Major Functions. Trends Cell Biol. 2018, 28, 34–45. [Google Scholar] [CrossRef]
- Burke, B.; Stewart, C.L. The nuclear lamins: Flexibility in function. Nat. Rev. Mol. Cell Biol. 2012, 14, 13–24. [Google Scholar] [CrossRef]
- Maurer, M.; Lammerding, J. The Driving Force: Nuclear Mechanotransduction in Cellular Function, Fate, and Disease. Annu. Rev. Biomed. Eng. 2019, 21, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Guilluy, C.; Osborne, L.D.; Van Landeghem, L.; Sharek, L.; Superfine, R.; Garcia-Mata, R.; Burridge, K. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nature 2014, 16, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.J.; Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nat. Cell Biol. 2018, 20, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Isermann, P.; Lammerding, J. Nuclear mechanics and mechanotransduction in health and disease. Curr. Biol. 2013, 23, R1113–R1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharana, S.; Iyer, K.V.; Jain, N.; Nagarajan, M.; Wang, Y.; Shivashankar, G.V. Chromosome intermingling-the physical basis of chromosome organization in differentiated cells. Nucleic Acids Res. 2016, 44, 5148–5160. [Google Scholar] [CrossRef] [Green Version]
- Thévenin, A.; Ein-Dor, L.; Ozery-Flato, M.; Shamir, R. Functional gene groups are concentrated within chromosomes, among chromosomes and in the nuclear space of the human genome. Nucleic Acids Res. 2014, 42, 9854–9861. [Google Scholar] [CrossRef] [Green Version]
- Uhler, C.; Shivashankar, G. Chromosome Intermingling: Mechanical Hotspots for Genome Regulation. Trends Cell Biol. 2017, 27, 810–819. [Google Scholar] [CrossRef]
- Branco, M.R.; Pombo, A. Intermingling of Chromosome Territories in Interphase Suggests Role in Translocations and Transcription-Dependent Associations. PLoS Biol. 2006, 4, e138. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chen, J.; Muir, L.A.; Ronquist, S.; Meixner, W.; Ljungman, M.; Ried, T.; Smale, S.; Rajapakse, I. Functional organization of the human 4D Nucleome. Proc. Natl. Acad. Sci. USA 2015, 112, 8002–8007. [Google Scholar] [CrossRef] [Green Version]
- Feuerborn, A.; Cook, P.R. Why the activity of a gene depends on its neighbors. Trends Genet. 2015, 31, 483–490. [Google Scholar] [CrossRef]
- Shah, J.S.; Sabouni, R.; Vaught, K.C.C.; Owen, C.M.; Albertini, D.F.; Segars, J.H. Biomechanics and mechanical signaling in the ovary: A systematic review. J. Assist. Reprod. Genet. 2018, 35, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Riechmann, V. The Role of the Actomyosin Cytoskeleton in Coordination of Tissue Growth during Drosophila Oogenesis. Curr. Biol. 2007, 17, 1349–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartain, C.V.; Wolfner, M.F. Calcium and egg activation in Drosophila. Cell Calcium 2012, 53, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horner, V.L.; Wolfner, M.F. Transitioning from egg to embryo: Triggers and mechanisms of egg activation. Dev. Dyn. 2008, 237, 527–544. [Google Scholar] [CrossRef]
- Cram, E.J. Mechanotransduction: Feeling the squeeze in the C. elegans reproductive system. Curr. Biol. 2015, 25, R74–R75. [Google Scholar] [CrossRef] [Green Version]
- Landrein, B.; Kiss, A.; Sassi, M.; Chauvet, A.; Das, P.; Cortizo, M.; Laufs, P.; Takeda, S.; Aida, M.; Traas, J.; et al. Mechanical stress contributes to the expression of the STM homeobox gene in Arabidopsis shoot meristems. eLife 2015, 4, e07811. [Google Scholar] [CrossRef]
- Schmid, M.F.; Sherman, M.B.; Matsudaira, P.; Chiu, W. Structure of the acrosomal bundle. Nature 2004, 431, 104–107. [Google Scholar] [CrossRef]
- Sanders, M.C.; Way, M.; Sakai, J.; Matsudaira, P. Characterization of the Actin Cross-linking Properties of the Scruin-Calmodulin Complex from the Acrosomal Process of Limulus Sperm. J. Biol. Chem. 1996, 271, 2651–2657. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, C.; Srayko, M.; Nédélec, F. Cortical Microtubule Contacts Position the Spindle in C. elegans Embryos. Cell 2007, 129, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Landmann, F.; Zahreddine, H.; Rodríguez, D.; Koch, M.; Labouesse, M. A tension-induced mechanotransduction pathway promotes epithelial morphogenesis. Nature 2011, 471, 99–103. [Google Scholar] [CrossRef]
- Monier, B.; Pelissier-Monier, A.; Brand, A.H.; Sanson, B. An actomyosin-based barrier inhibits cell mixing at compartmental boundaries in Drosophila embryos. Nature 2009, 12, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertet, C.; Sulak, L.; Lecuit, T. Myosin-dependent junction remodelling controls planar cell intercalation and axis elongation. Nature 2004, 429, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.C.; Fernández-González, R. Local mechanical forces promote polarized junctional assembly and axis elongation in Drosophila. eLife 2016, 5, e10757. [Google Scholar] [CrossRef] [PubMed]
- Jacinto, A.; Wood, W.; Balayo, T.; Turmaine, M.; Martinez-Arias, A.; Martin, P. Dynamic actin-based epithelial adhesion and cell matching during Drosophila dorsal closure. Curr. Biol. 2000, 10, 1420–1426. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.W.; E Keller, R.; A Koehl, M. The dorsal involuting marginal zone stiffens anisotropically during its convergent extension in the gastrula of Xenopus laevis. Development 1995, 121, 3131–3140. [Google Scholar]
- Weber, G.F.; Bjerke, M.A.; DeSimone, D.W. A Mechanoresponsive Cadherin-Keratin Complex Directs Polarized Protrusive Behavior and Collective Cell Migration. Dev. Cell 2012, 22, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Brunet, T.; Bouclet, A.; Ahmadi, P.; Mitrossilis, D.; Driquez, B.; Brunet, A.-C.; Henry, L.; Serman, F.; Béalle, G.; Menager, C.; et al. Evolutionary conservation of early mesoderm specification by mechanotransduction in Bilateria. Nat. Commun. 2013, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Krieg, M.; Arboleda-Estudillo, Y.; Puech, P.-H.; Kafer, J.; Graner, F.; Müller, D.J.; Heisenberg, C.-P. Tensile forces govern germ-layer organization in zebrafish. Nature 2008, 10, 429–436. [Google Scholar] [CrossRef]
- Maître, J.-L.; Turlier, H.; Illukkumbura, R.; Eismann, B.; Niwayama, R.; Nédélec, F.; Hiiragi, T. Asymmetric division of contractile domains couples cell positioning and fate specification. Nature 2016, 536, 344–348. [Google Scholar] [CrossRef]
- Warmflash, A.; Sorre, B.; Etoc, F.; Siggia, E.D.; Brivanlou, A.H. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat. Methods 2014, 11, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Ingber, D.E.; Wang, N.; Stamenović, D. Tensegrity, cellular biophysics, and the mechanics of living systems. Rep. Prog. Phys. 2014, 77, 046603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingber, D.E.; Madri, J.A.; Jamieson, J.D. Role of basal lamina in neoplastic disorganization of tissue architecture. Proc. Natl. Acad. Sci. USA 1981, 78, 3901–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, B. Tensegrity Portfolio Artnews Annual. 1961, 4, 112–127. Available online: http://www.rwgrayprojects.com/rbfnotes/fpapers/tensegrity/tenseg01.html (accessed on 2 July 2020).
- Koser, D.E.; Thompson, A.J.; Foster, S.K.; Dwivedy, A.; Pillai, E.K.; Sheridan, G.K.; Svoboda, H.; Viana, M.; Costa, L.D.F.; Guck, J.; et al. Mechanosensing is critical for axon growth in the developing brain. Nat. Neurosci. 2016, 19, 1592–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, N.; Conwell, M.D.; Chen, X.; Kettenhofen, C.I.; Westlake, C.J.; Cantor, L.B.; Wells, C.D.; Weinreb, R.N.; Corson, T.W.; Spandau, D.F.; et al. Primary cilia signaling mediates intraocular pressure sensation. Proc. Natl. Acad. Sci. USA 2014, 111, 12871–12876. [Google Scholar] [CrossRef] [Green Version]
- Rabbitt, R.D.; Boyle, R.; Highstein, S.M. Mechanical amplification by hair cells in the semicircular canals. Proc. Natl. Acad. Sci. USA 2010, 107, 3864–3869. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Safferling, K.; Rausch, S.; Grabe, N.; Boehm, H.; Spatz, J.P. A molecular mechanotransduction pathway regulates collective migration of epithelial cells. Nature 2015, 17, 276–287. [Google Scholar] [CrossRef]
- Palmisano, M.G.; Bremner, S.N.; Hornberger, T.A.; Meyer, G.; Domenighetti, A.A.; Shah, S.B.; Kiss, B.; Kellermayer, M.; Ryan, A.F.; Lieber, R.L. Skeletal muscle intermediate filaments form a stress-transmitting and stress-signaling network. J. Cell Sci. 2014, 128, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, K.; Woo, S.-H.; Chang, R.B.; Gillich, A.; Qiu, Z.; Francisco, A.G.; Ranade, S.S.; Liberles, S.D.; Patapoutian, A. Piezo2 senses airway stretch and mediates lung inflation-induced apnoea. Nature 2016, 541, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Nerurkar, N.L.; Mahadevan, L.; Tabin, C.J. BMP signaling controls buckling forces to modulate looping morphogenesis of the gut. Proc. Natl. Acad. Sci. USA 2017, 114, 2277–2282. [Google Scholar] [CrossRef] [Green Version]
- Nauli, S.M.; Alenghat, F.J.; Luo, Y.; Williams, E.; Vassilev, P.; Li, X.; Elia, A.E.H.; Lu, W.; Brown, E.M.; Quinn, S.; et al. Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat. Genet. 2003, 33, 129–137. [Google Scholar] [CrossRef]
- Lorenz, L.; Axnick, J.; Buschmann, T.; Henning, C.; Urner, S.; Fang, S.; Nurmi, H.; Eichhorst, N.; Holtmeier, R.; Bódis, K.; et al. Mechanosensing by β1 integrin induces angiocrine signals for liver growth and survival. Nature 2018, 562, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Lucitti, J.L.; Jones, E.A.V.; Huang, C.; Chen, J.; Fraser, S.; Dickinson, M.E. Vascular remodeling of the mouse yolk sac requires hemodynamic force. Development 2007, 134, 3317–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- North, T.E.; Goessling, W.; Peeters, M.; Li, P.; Ceol, C.; Lord, A.M.; Weber, G.J.; Harris, J.; Cutting, C.C.; Huang, P.L.; et al. Hematopoietic Stem Cell Development Is Dependent on Blood Flow. Cell 2009, 137, 736–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, A.H.; Raftrey, B.C.; D’Amato, G.; Surya, V.N.; Poduri, A.; Chen, H.I.; Goldstone, A.B.; Woo, J.; Fuller, G.G.; Dunn, A.R.; et al. DACH1 stimulates shear stress-guided endothelial cell migration and coronary artery growth through the CXCL12–CXCR4 signaling axis. Genes Dev. 2017, 31, 1308–1324. [Google Scholar] [CrossRef] [Green Version]
- Beis, D.; Kalogirou, S.; Tsigkas, N. Insights into Heart Development and Regeneration. In Introduction to Translational Cardiovascular Research; Springer: Cham, Switzerland, 2015; Chapter 2; pp. 17–30. [Google Scholar]
- Forouhar, A.S.; Liebling, M.; Hickerson, A.; Nasiraei-Moghaddam, A.; Tsai, H.-J.; Hove, J.R.; Fraser, S.; Dickinson, M.E.; Gharib, M. The Embryonic Vertebrate Heart Tube Is a Dynamic Suction Pump. Science 2006, 312, 751–753. [Google Scholar] [CrossRef] [Green Version]
- Burggren, W.W. What Is the Purpose of the Embryonic Heart Beat? or How Facts Can Ultimately Prevail over Physiological Dogma. Physiol. Biochem. Zool. 2004, 77, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Majkut, S.; Dingal, P.D.P.; Discher, D.E. Stress Sensitivity and Mechanotransduction during Heart Development. Curr. Biol. 2014, 24, R495–R501. [Google Scholar] [CrossRef] [Green Version]
- Haack, T.; Abdelilah-Seyfried, S. The force within: Endocardial development, mechanotransduction and signalling during cardiac morphogenesis. Development 2016, 143, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.F. Flow-Mediated Endothelial Mechanotransduction. Physiol. Rev. 1995, 75, 519–560. [Google Scholar] [CrossRef]
- Olesen, S.-P.; Clapham, D.E.; Davies, P.F. Haemodynamic shear stress activates a K+ current in vascular endothelial cells. Nature 1988, 331, 168–170. [Google Scholar] [CrossRef]
- Dewey, C.F.; Bussolari, S.R.; Gimbrone, M.A.; Davies, P.F. The Dynamic Response of Vascular Endothelial Cells to Fluid Shear Stress. J. Biomech. Eng. 1981, 103, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.M.; Izumo, S. Control of endothelial cell gene expression by flow. J. Biomech. 1995, 28, 1515–1528. [Google Scholar] [CrossRef]
- Coon, B.G.; Baeyens, N.; Han, J.; Budatha, M.; Ross, T.D.; Fang, J.S.; Yun, S.; Thomas, J.-L.; Schwartz, M.A. Intramembrane binding of VE-cadherin to VEGFR2 and VEGFR3 assembles the endothelial mechanosensory complex. J. Gen. Physiol. 2015, 145, 975–986. [Google Scholar] [CrossRef] [Green Version]
- Tzima, E.; Irani-Tehrani, M.; Kiosses, W.B.; Dejana, E.; Schultz, D.A.; Engelhardt, B.; Cao, G.; Delisser, H.; Schwartz, M.A. A mechanosensory complex that mediates the endothelial cell response to fluid shear stress. Nature 2005, 437, 426–431. [Google Scholar] [CrossRef]
- Tzima, E.; Del Pozo, M.A.; Shattil, S.J.; Chien, S.; Schwartz, M.A. Activation of integrins in endothelial cells by fluid shear stress mediates Rho-dependent cytoskeletal alignment. EMBO J. 2001, 20, 4639–4647. [Google Scholar] [CrossRef] [Green Version]
- Tzima, E.; Kiosses, W.B.; Del Pozo, M.A.; Schwartz, M.A. Localized Cdc42 Activation, Detected Using a Novel Assay, Mediates Microtubule Organizing Center Positioning in Endothelial Cells in Response to Fluid Shear Stress. J. Biol. Chem. 2003, 278, 31020–31023. [Google Scholar] [CrossRef] [Green Version]
- Baeyens, N.; Nicoli, S.; Coon, B.G.; Ross, T.D.; Dries, K.V.D.; Han, J.; Lauridsen, H.M.; O Mejean, C.; Eichmann, A.; Thomas, J.-L.; et al. Vascular remodeling is governed by a VEGFR3-dependent fluid shear stress set point. eLife 2015, 4. [Google Scholar] [CrossRef]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef]
- Lebart, M.-C.; Benyamin, Y. Calpain involvement in the remodeling of cytoskeletal anchorage complexes. FEBS J. 2006, 273, 3415–3426. [Google Scholar] [CrossRef] [Green Version]
- Duchemin, A.-L.; Vignes, H.; Vermot, J. Mechanically activated piezo channels modulate outflow tract valve development through the Yap1 and Klf2-Notch signaling axis. eLife 2019, 8. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haffter, P.; Granato, M.; Brand, M.; Mullins, M.C.; Hammerschmidt, M.; Kane, D.A.; Odenthal, J.; Van Eeden, F.J.; Jiang, Y.J.; Heisenberg, C.P.; et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 1996, 123, 1–36. [Google Scholar] [PubMed]
- Weber, M.; Huisken, J. In vivo imaging of cardiac development and function in zebrafish using light sheet microscopy. Swiss Med Wkly. 2015, 145, 5152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelster, B.; Burggren, W.W. Disruption of Hemoglobin Oxygen Transport Does Not Impact Oxygen-Dependent Physiological Processes in Developing Embryos of Zebra Fish (Danio rerio). Circ. Res. 1996, 79, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Sehnert, A.J.; Huq, A.; Weinstein, B.M.; Walker, C.; Fishman, M.; Stainier, D.Y. Cardiac troponin T is essential in sarcomere assembly and cardiac contractility. Nat. Genet. 2002, 31, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The zebrafish as a model for complex tissue regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart Regeneration in Zebrafish. Sciences 2002, 298, 2188–2190. [Google Scholar] [CrossRef]
- Huisken, J. Optical Sectioning Deep Inside Live Embryos by Selective Plane Illumination Microscopy. Science 2004, 305, 1007–1009. [Google Scholar] [CrossRef] [Green Version]
- Huisken, J.; Stainier, D.Y. Even fluorescence excitation by multidirectional selective plane illumination microscopy (mSPIM). Opt. Lett. 2007, 32, 2608–2610. [Google Scholar] [CrossRef]
- Mickoleit, M.; Schmid, B.; Weber, M.; Fahrbach, F.O.; Hombach, S.; Reischauer, S.; Huisken, J. High-resolution reconstruction of the beating zebrafish heart. Nat. Methods 2014, 11, 919–922. [Google Scholar] [CrossRef]
- Kaufmann, A.; Mickoleit, M.; Weber, M.; Huisken, J. Multilayer mounting enables long-term imaging of zebrafish development in a light sheet microscope. Development 2012, 139, 3242–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hove, J.R.; Köster, R.W.; Forouhar, A.S.; Acevedo-Bolton, G.; Fraser, S.; Gharib, M. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature 2003, 421, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Adamo, L.; Naveiras, O.; Wenzel, P.L.; McKinney-Freeman, S.; Mack, P.J.; Gracia-Sancho, J.; Suchy-Dicey, A.M.; Yoshimoto, M.; Lensch, M.W.; Yoder, M.C.; et al. Biomechanical forces promote embryonic haematopoiesis. Nature 2009, 459, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.Y.; Yang, G.-M.; Dayem, A.A.; Saha, S.K.; Kim, K.; Yoo, Y.; Hong, K.; Kim, J.-H.; Yee, C.; Lee, J.-Y.; et al. Hydrodynamic shear stress promotes epithelial-mesenchymal transition by downregulating ERK and GSK3β activities. Breast Cancer Res. 2019, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Kalogirou, S.; Malissovas, N.; Moro, E.; Argenton, F.; Stainier, D.Y.; Beis, D. Intracardiac flow dynamics regulate atrioventricular valve morphogenesis. Cardiovasc. Res. 2014, 104, 49–60. [Google Scholar] [CrossRef]
- Auman, H.J.; Coleman, H.; Riley, H.E.; Olale, F.; Tsai, H.-J.; Yelon, D. Functional modulation of cardiac form through regionally confined cell shape changes. PLoS Biol. 2007, 5, e53. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, N.; Ostwald, P.; Bark, D.; Garrity, D. Biomechanical Cues Direct Valvulogenesis. J. Cardiovasc. Dev. Dis. 2020, 7, 18. [Google Scholar] [CrossRef]
- Chi, N.C.; Shaw, R.M.; Jungblut, B.; Huisken, J.; Ferrer, T.; Arnaout, R.; Scott, I.C.; Beis, D.; Xiao, T.; Baier, H.; et al. Genetic and physiologic dissection of the vertebrate cardiac conduction system. PLoS Biol. 2008, 6, e109. [Google Scholar] [CrossRef] [Green Version]
- Peshkovsky, C.; Totong, R.; Yelon, D. Dependence of cardiac trabeculation on neuregulin signaling and blood flow in zebrafish. Dev. Dyn. 2011, 240, 446–456. [Google Scholar] [CrossRef]
- Samsa, L.A.; Givens, C.; Tzima, E.; Stainier, D.Y.; Qian, L.; Liu, J. Cardiac contraction activates endocardial Notch signaling to modulate chamber maturation in zebrafish. Development 2015, 142, 4080–4091. [Google Scholar] [CrossRef] [Green Version]
- Beis, D.; Bartman, T.; Jin, S.-W.; Scott, I.C.; D’Amico, L.A.; Ober, E.A.; Verkade, H.; Frantsve, J.; Field, H.A.; Wehman, A.M.; et al. Genetic and cellular analyses of zebrafish atrioventricular cushion and valve development. Development 2005, 132, 4193–4204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherz, P.J.; Huisken, J.; Sahai-Hernandez, P.; Stainier, D.Y. High-speed imaging of developing heart valves reveals interplay of morphogenesis and function. Development 2008, 135, 1179–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermot, J.; Forouhar, A.S.; Liebling, M.; Wu, D.; Plummer, D.; Gharib, M.; Fraser, S.E. Reversing Blood Flows Act through klf2a to Ensure Normal Valvulogenesis in the Developing Heart. PLoS Biol. 2009, 7, e1000246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heckel, E.; Boselli, F.; Roth, S.; Krudewig, A.; Belting, H.-G.; Charvin, G.; Vermot, J. Oscillatory Flow Modulates Mechanosensitive klf2a Expression through trpv4 and trpp2 during Heart Valve Development. Curr. Biol. 2015, 25, 1354–1361. [Google Scholar] [CrossRef] [Green Version]
- Nicoli, S.; Standley, C.; Walker, P.; Hurlstone, A.F.L.; Fogarty, K.E.; Lawson, N.D. MicroRNA-mediated integration of haemodynamics and Vegf signalling during angiogenesis. Nature 2010, 464, 1196–1200. [Google Scholar] [CrossRef] [Green Version]
- Patrick, D.M.; Montgomery, R.L.; Qi, X.; Obad, S.; Kauppinen, S.; Hill, J.A.; Van Rooij, E.; Olson, E.N. Stress-dependent cardiac remodeling occurs in the absence of microRNA-21 in mice. J. Clin. Investig. 2010, 120, 3912–3916. [Google Scholar] [CrossRef] [Green Version]
- Renz, M.; Otten, C.; Faurobert, E.; Rudolph, F.; Zhu, Y.; Boulday, G.; Duchene, J.; Mickoleit, M.; Dietrich, A.-C.; Ramspacher, C.; et al. Regulation of β1 Integrin-Klf2-Mediated Angiogenesis by CCM Proteins. Dev. Cell 2015, 32, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Faurobert, E.; Albiges-Rizo, C. Recent insights into cerebral cavernous malformations: A complex jigsaw puzzle under construction. FEBS J. 2010, 277, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Fitch, M.J.; Campagnolo, L.; Kuhnert, F.; Stuhlmann, H. Egfl7, a novel epidermal growth factor-domain gene expressed in endothelial cells. Dev. Dyn. 2004, 230, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Donat, S.; Lourenco, M.; Paolini, A.; Otten, C.; Renz, M.; Abdelilah-Seyfried, S. Heg1 and Ccm1/2 proteins control endocardial mechanosensitivity during zebrafish valvulogenesis. Elife 2018, 7, e28939. [Google Scholar] [CrossRef]
- Rödel, C.J.; Otten, C.; Donat, S.; Lourenco, M.; Fischer, D.; Kuropka, B.; Paolini, A.; Freund, C.; Abdelilah-Seyfried, S. Blood Flow Suppresses Vascular Anomalies in a Zebrafish Model of Cerebral Cavernous Malformations. Circ. Res. 2019, 125, e43–e54. [Google Scholar] [CrossRef] [PubMed]
- Campinho, P.; Vilfan, A.; Vermot, J. Blood Flow Forces in Shaping the Vascular System: A Focus on Endothelial Cell Behavior. Front. Physiol. 2020, 11, 552. [Google Scholar] [CrossRef]
- Tsonis, P.A. Regenerative biology: The emerging field of tissue repair and restoration. Differentiation 2002, 70, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Cajal, S.R.Y.; May, R.M. Cajal’s Degeneration and Regeneration of the Nervous System; Oxford University Press: New York, NY, USA, 1991. [Google Scholar]
- Horner, P.J.; Gage, F.H. Regenerating the damaged central nervous system. Nature 2000, 407, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Olson, E.N. A neonatal blueprint for cardiac regeneration. Stem Cell Res. 2014, 13, 556–570. [Google Scholar] [CrossRef] [Green Version]
- Richardson, P.M.; McGuinness, U.M.; Aguayo, A.J.; Richardson, U.M.M.P.M. Axons from CNS neurones regenerate into PNS grafts. Nature 1980, 284, 264–265. [Google Scholar] [CrossRef]
- Moya, I.M.; Halder, G. Hippo–YAP/TAZ signalling in organ regeneration and regenerative medicine. Nat. Rev. Mol. Cell Biol. 2018, 20, 211–226. [Google Scholar] [CrossRef]
- Pocaterra, A.; Romani, P.; Dupont, S. YAP/TAZ functions and their regulation at a glance. J. Cell Sci. 2020, 133, jcs230425. [Google Scholar] [CrossRef]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Gourdie, R.G.; Dimmeler, S.; Kohl, P. Novel therapeutic strategies targeting fibroblasts and fibrosis in heart disease. Nat. Rev. Drug Discov. 2016, 15, 620–638. [Google Scholar] [CrossRef] [Green Version]
- Iranzo, H.S.; Galardi-Castilla, M.; Sanz-Morejón, A.; González-Rosa, J.M.; Costa, R.; Ernst, A.; De Aja, J.S.; Langa, X.; Mercader, N. Transient fibrosis resolves via fibroblast inactivation in the regenerating zebrafish heart. Proc. Natl. Acad. Sci. USA 2018, 115, 4188–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, F.C.; Cahill, T.J.; Kenyon, A.; Gavriouchkina, D.; Vieira, J.M.; Sun, X.; Pezzolla, D.; Ravaud, C.; Masmanian, E.; Weinberger, M.; et al. Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair. Nat. Commun. 2020, 11, 600–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psarras, S.; Beis, D.; Nikouli, S.; Tsikitis, M.; Capetanaki, Y. Three in a Box: Understanding Cardiomyocyte, Fibroblast, and Innate Immune Cell Interactions to Orchestrate Cardiac Repair Processes. Front. Cardiovasc. Med. 2019, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Moeendarbary, E.; Weber, I.P.; Sheridan, G.K.; Koser, D.E.; Soleman, S.; Haenzi, B.; Bradbury, E.J.; Fawcett, J.; Franze, K. The soft mechanical signature of glial scars in the central nervous system. Nat. Commun. 2017, 8, 14787. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.A.; Ju, Y.-E.; Marg, B.; Osterfield, M.; Janmey, P.A. Neurite branching on deformable substrates. NeuroReport 2002, 13, 2411–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georges, P.C.; Miller, W.J.; Meaney, D.F.; Sawyer, E.S.; Janmey, P.A. Matrices with Compliance Comparable to that of Brain Tissue Select Neuronal over Glial Growth in Mixed Cortical Cultures. Biophys. J. 2006, 90, 3012–3018. [Google Scholar] [CrossRef] [Green Version]
- Möllmert, S.; Kharlamova, M.A.; Hoche, T.; Taubenberger, A.V.; Abuhattum, S.; Kuscha, V.; Kurth, T.; Brand, M.; Guck, J. Zebrafish Spinal Cord Repair Is Accompanied by Transient Tissue Stiffening. Biophys. J. 2020, 118, 448–463. [Google Scholar] [CrossRef]
- Maden, M. A history of regeneration research. Milestones in the evolution of a science. Cell 1992, 69, 723–724. [Google Scholar] [CrossRef]
- Kefalos, P.; Agalou, A.; Kawakami, K.; Beis, D. Reactivation of Notch signaling is required for cardiac valve regeneration. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Brito, A.B.; Ramkumar, S.; Boezio, G.L.; Guenther, S.; Kuenne, C.; Helker, C.S.; Sánchez-Iranzo, H.; Iloska, D.; Piesker, J.; Pullamsetti, S.; et al. TGF-β Signaling Promotes Tissue Formation during Cardiac Valve Regeneration in Adult Zebrafish. Dev. Cell 2020, 52, 9–20. [Google Scholar] [CrossRef]
- Emmert, M.Y.; Schmitt, B.; Loerakker, S.; Sanders, B.; Spriestersbach, H.; Fioretta, E.S.; Bruder, L.; Brakmann, K.; Motta, S.E.; Lintas, V.; et al. Computational modeling guides tissue-engineered heart valve design for long-term in vivo performance in a translational sheep model. Sci. Transl. Med. 2018, 10, eaan4587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourgiantaki, A.; Tzeranis, D.S.; Karali, K.; Georgelou, K.; Bampoula, E.; Psilodimitrakopoulos, S.; Yannas, I.V.; Stratakis, E.; Sidiropoulou, K.; Charalampopoulos, I.; et al. Neural stem cell delivery via porous collagen scaffolds promotes neuronal differentiation and locomotion recovery in spinal cord injury. npj Regen. Med. 2020, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, M.; Benedetti, E.; Tupone, M.G.; Catanesi, M.; Castelli, V.; Antonosante, A.; Cimini, A. The Role of Stiffness in Cell Reprogramming: A Potential Role for Biomaterials in Inducing Tissue Regeneration. Cells 2019, 8, 1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Examples of key mechanotransduction stimuli and occurring cellular events from bacteria to humans.
Figure 1.
Examples of key mechanotransduction stimuli and occurring cellular events from bacteria to humans.

Figure 2.
Integration of external stimuli. The perception and interpretation of external signals starts at the cell membrane. Cells sense changes in their surface parameters through transmembrane receptors, ion or cell–cell junctional channels. Sensing of external cues originating from the extracellular matrix (ECM), takes place in specialized sites of interaction between the ECM and the membrane.
Figure 2.
Integration of external stimuli. The perception and interpretation of external signals starts at the cell membrane. Cells sense changes in their surface parameters through transmembrane receptors, ion or cell–cell junctional channels. Sensing of external cues originating from the extracellular matrix (ECM), takes place in specialized sites of interaction between the ECM and the membrane.

Figure 3.
Signal propagation to the nucleus. Biochemical and mechanical stimuli are transferred to the nucleus and become effective through i) direct physical interactions of the membrane, the cytoskeleton and the nucleus and/or ii) the translocation of activated mediators from the cytoplasm to the nucleus. Eventually, stimuli surpassing a certain threshold can instruct transcriptional changes and regulate the gene expression pattern of a given cell.
Figure 3.
Signal propagation to the nucleus. Biochemical and mechanical stimuli are transferred to the nucleus and become effective through i) direct physical interactions of the membrane, the cytoskeleton and the nucleus and/or ii) the translocation of activated mediators from the cytoplasm to the nucleus. Eventually, stimuli surpassing a certain threshold can instruct transcriptional changes and regulate the gene expression pattern of a given cell.

Figure 4.
Hemodynamics and stages of cardiogenesis in zebrafish and mammals. In both zebrafish and mammals, shear forces that occur from the oscillatory/retrograde blood flow and are driven by cardiac contractility and heart looping, direct valvulogenesis. Hence, cardiac valves are formed following the initiation of heart function. Valve cells originated from endocardial cells forming cardiac cushions. Valve Endothelial Cells (VECs) and Valve Intersitial Cells (VICs) can be identified in both systems. In zebrafish, embryonic cushions give rise to cardiac leaflets that invaginate and generate cardiac valves. In contrast, in mammals, endocardial cells undergo an endothelial-to-mesenchymal transition, delaminate generate cardiac valves upon invasion into the cardiac jelly. The outer layer of epicardial cells and the cardiac jelly is omitted in all schemes for figure clarity.
Figure 4.
Hemodynamics and stages of cardiogenesis in zebrafish and mammals. In both zebrafish and mammals, shear forces that occur from the oscillatory/retrograde blood flow and are driven by cardiac contractility and heart looping, direct valvulogenesis. Hence, cardiac valves are formed following the initiation of heart function. Valve cells originated from endocardial cells forming cardiac cushions. Valve Endothelial Cells (VECs) and Valve Intersitial Cells (VICs) can be identified in both systems. In zebrafish, embryonic cushions give rise to cardiac leaflets that invaginate and generate cardiac valves. In contrast, in mammals, endocardial cells undergo an endothelial-to-mesenchymal transition, delaminate generate cardiac valves upon invasion into the cardiac jelly. The outer layer of epicardial cells and the cardiac jelly is omitted in all schemes for figure clarity.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsata, V.; Beis, D. In Full Force. Mechanotransduction and Morphogenesis during Homeostasis and Tissue Regeneration. J. Cardiovasc. Dev. Dis. 2020, 7, 40. https://doi.org/10.3390/jcdd7040040
AMA Style
Tsata V, Beis D. In Full Force. Mechanotransduction and Morphogenesis during Homeostasis and Tissue Regeneration. Journal of Cardiovascular Development and Disease. 2020; 7(4):40. https://doi.org/10.3390/jcdd7040040
Chicago/Turabian StyleTsata, Vasiliki, and Dimitris Beis. 2020. "In Full Force. Mechanotransduction and Morphogenesis during Homeostasis and Tissue Regeneration" Journal of Cardiovascular Development and Disease 7, no. 4: 40. https://doi.org/10.3390/jcdd7040040
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.