
Ramomarthamyces octomerus sp. nov. and Insights into the Evolution and Diversification of Ramomarthamyces (Ascomycota, Leotiomycetes, Marthamycetales)

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens, Cultures, and Sequences Used in This Study
2.2. Additional Specimens Examined
2.3. Purchased Cultures Sequenced
2.4. Morphological Analysis
2.5. Culture Methods
2.6. Structures Measured
2.7. Apothecium Terminology
2.8. DNA Extraction, PCR Amplification and Sequencing
2.9. Handling Sequence Data
2.10. Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analyses

3.2. Morphological Analyses
4. Taxonomy
5. Discussion
5.1. Phylogenetic Placement
5.2. On the Placement of Our New Species
5.3. Notes on Life History
5.3.1. Trophic Mode
5.3.2. Endophytic States
5.3.3. Mitosporic States
5.4. Ecological Description of Collection Sites
5.5. Morphology
5.5.1. Amyloid Ascus Pores
5.5.2. Hymenial Gel
5.5.3. Paraphyses
5.5.4. Ascospores
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baral, H.O.; Lantz, H.; Hustad, V.; Minter, D.W. Marthamycetaceae. In Nomenclatural Novelties; Index Fungorum No. 225; Baral, H.O., Ed.; 2015; ISSN 2049-2375. http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp. [This treatment of Marthamycetaceae is adapted with a few minor changes in Baral, H.O. 2016. In Frey, W. (ed.), Syllabus of Plant Families (13th edn). [Adolf Engler’s Syllabus der Pflanzenfamilien]. Part 1/2. Ascomycota. Stuttgart: Borntraeger Science Publishers. https://www.schweizerbart.de/publications/detail/isbn/9783443010898/Syllabus_of_Plant_Families_Part_1_2_A]. ISSN 2049-2375.
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Magurno, F. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Bellemère, A. Contribution a l’étude du dévelopment de l’apothécie chez les discomycètes inoperculés I. Bull. Trimest. Soc. Mycol. 1967, 83, 393–640, [treatment of Propolis farinosa (as Propolis faginea)]. [Google Scholar]
- Fries, E.M. Systema Mycologicum; Lund: Berling, Germany, 1822; Volume 2, Part 1. [Google Scholar]
- Corda, A.K.J. Icones Fungorum Hucusque Cognitorum; J.G. Calve: Prague, Czech Republic, 1838; Volume 2. [Google Scholar]
- von Butin, H. Morphologische und taxonomische Untersuchungen an Naemacyclus niveus (Pers. ex Fr.) Fuck. ex Sacc. und verwandten Arten. Eur. J. Forest Pathol. 1973, 3, 146–163. [Google Scholar] [CrossRef]
- Sherwood, M.A. Taxonomic studies in the Phacidiales: Stictis maritima and the genus Lasiostictis. Mycotaxon 1974, 1, 41–44. [Google Scholar]
- DiCosmo, F. Lasiostictis reassessed. Can. J. Bot. 1979, 57, 1838–1840. [Google Scholar] [CrossRef]
- Sherwood, M.A. Taxonomic studies in the Phacidiales: Propolis and Propolomyces. Mycotaxon 1977, 5, 320–330. [Google Scholar]
- DiCosmo, F.; Peredo, H.; Minter, D.W. Cyclaneusma gen. nov., Naemacyclus and Lasiostictis, a nomenclatural problem resolved. Eur. J. For. Pathol. 1983, 13, 206–212. [Google Scholar] [CrossRef]
- DiCosmo, F.; Nag Raj, T.R.; Kendrick, W.B. A revision of Phacidiaceae and related anamorphs. Mycotaxon 1984, 21, 1–234. [Google Scholar]
- Minter, D.W. Some Members of the Rhytismataceae (Ascomycetes) on Conifer Needles from Central and North America. In Recent Research on Conifer Needle Diseases, Conference Proceedings, Gulfport, Mississippi, October 14–18 1984; Peterson, G.W., Ed.; USDA Forest Service General Technical Report GTR-WO 50; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1986; pp. 71–106. https://www.biodiversitylibrary.org/item/171832#page/1/mode/1up.
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2005; 660p, [Cyclaneusma needle-cast]. [Google Scholar]
- Baral, H.O. Key to Marthamycetaceae (Propoloid Discomycetes). 2015. https://in-vivo-veritas.de/keys/key-to-marthamycetaceae-propoloid-discomycetes.
- Johnston, P.R. Rhytismataceae in New Zealand 1. Some foliicolous species of Coccomyces de Notaris and Propolis (Fries) Corda. N. Z. J. Bot. 1986, 24, 89–124. [Google Scholar] [CrossRef]
- Johnston, P.R. Rhytismatales of Australia: The genus Marthamyces. Aust. Syst. Bot. 2006, 19, 135–146. [Google Scholar] [CrossRef]
- Hosoya, T.; Hosaka, K.; Saito, Y.; Degawa, Y.; Suzuki, R. Naemacyclus culmigenus, a newly reported potential pathogen to Miscanthus sinensis, new to Japan. Mycoscience 2013, 54, 433–437. [Google Scholar] [CrossRef]
- Chlebická, M. Propolis rhodoleuca (Leotiomycetes, inc. sed.) compared with P. farinosa, P. occulta sp. nov. and P. strobilina. Nova Hedwig. 2014, 98, 491–506. [Google Scholar] [CrossRef]
- Minter, D.W. Propolis farinosa. I. M. I. Descript. Fungi Bact.; No. 218: Sheet 2176; CABI International: Wallingford, UK, 2019. [Google Scholar] [CrossRef]
- Johnston, P.R.; Park, D. New species of Marthamyces and Ramomarthamyces gen. nov. from New Zealand and the Cook Islands. Mycotaxon 2019, 134, 489–516. [Google Scholar] [CrossRef]
- Sherwood, M.A. Convergent evolution in discomycetes from bark and wood. Bot. J. Linn. Soc. 1981, 82, 15–34. [Google Scholar] [CrossRef]
- Baral, H.O.; Weber, E.; Marson, G. Monograph of Orbiliomycetes (Ascomycota) Based on Vital Taxonomy. Part I; National Museum of Natural History: Luxembourg, 2020; 1752p, https://www.mnhn.lu/science/monograph-of-orbiliomycetes/?lang=en.
- Lantz, H.; Johnston, P.R.; Park, D.; Minter, D.W. Molecular phylogeny reveals a core clade of Rhytismatales. Mycologia 2011, 103, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Hustad, V.P.; Miller, A.N. Phylogenetic placement of four genera within the Leotiomycetes (Ascomycota). N. Am. Fungi 2011, 6, 1–13. [Google Scholar] [CrossRef]
- Fries, E.M. Summa Vegetabilium Scandinaviae (Sectio Posterior); Typographia Academica: Uppsala, Sweden, 1849. [Google Scholar]
- Minter, D.W. Propolis and Marthamyces gen. nov. (Rhytismatales). Mycotaxon 2003, 87, 43–52. [Google Scholar]
- Baral, H.O. Vital versus herbarium taxonomy: Morphological differences between living and dead cells of ascomycetes, and their taxonomic implications. Mycotaxon 1992, 44, 333–390. [Google Scholar]
- van Brummelen, J. A World-Monograph of the Genera Ascobolus and Saccobolus (Ascomycetes, Pezizales); Rijksherbarium: Leiden, The Netherlands, 1967; Persoonia Supplement Volume 1. [Google Scholar]
- Karakehian, J.M.; Quijada, L.; Pfister, D.H.; Tocci, G.E.; Miller, A.N. Methods for observing, culturing, and studying living ascospores. Asian J. Mycol. 2021, 4, 1–18. [Google Scholar] [CrossRef]
- Sherwood, M.A. The ostropalean fungi. Mycotaxon 1977, 5, 1–277. [Google Scholar]
- Letrouit-Galinou, M.-A. The apothecia of the discolichens. Bryologist 1968, 71, 297–327. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes: Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Samuels, G.J. Molecular systematics of the Hypocreales: A teleomorph gene phylogeny and the status of their anamorphs. Can. J. Bot. 1995, 73 (Suppl. 1), S816–S823. [Google Scholar]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Molec. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucl. Acids Res. 2015, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Improved accuracy of multiple ncRNA alignment by incorporating structural information into a MAFFT-based framework. BMC Bioinform. 2008, 9, 212. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT, a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acid Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Abadi, S.; Azouri, D.; Mayrose, I.; Pupko, T. Model selection may not be a mandatory step for phylogeny reconstruction. Nat. Commun. 2019, 10, 934. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Quijada, L.; Baral, H.-O.; Jaen-Molina, R.; Weiss, M.; Caujapé-Castells, J.; Beltrán-Tejera, E. Phylogenetic and morphological circumscription of the Orbilia aurantiorubra group. Phytotaxa 2014, 175, 1–18. [Google Scholar] [CrossRef]
- Baral, H.O. Lugol’s solution/IKI versus Melzer’s reagent: Hemiamyloidity, a universal feature of the ascus wall. Mycotaxon 1987, 29, 399–450. [Google Scholar]
- von Schweinitz, L.D. Synopsis Fungorum in America Boreali Media Degentium; The Society: Washington, DC, USA, 1832; Secundum observations Ludovici Davidis de Schweinitz; Communicated to the American Philosophical Society, Philadelphia, 15 April 1831. Trans. Amer. Philos. Soc. 4 (New Series): 141–316. 1 pl. (pl. XIX) [Also issued with the same numbering in the completed volume of the Transactions of the American Philosophical Society, title page dated 1834]. [Google Scholar]
- Desmazières, J.B.H.J. Plantes cryptogames du Nord de la France, ed. 2, sér. 2. Fasc. 15–16, nos. 701–800. [Exsiccati]. 1860. [Google Scholar]
- Rodway, L. Botanical notes. Pap. Proc. R. Soc. Tasman. 1918, 105–110. [Google Scholar]
- Karakehian, J.M.; Quijada, L.; Friebes, G.; Tanney, J.B.; Pfister, D.H. Placement of Triblidiaceae in Rhytismatales and comments on unique ascospore morphologies in Leotiomycetes (Fungi, Ascomycota). MycoKeys 2019, 54, 99–133. [Google Scholar] [CrossRef] [PubMed]
- Araújo-Magalhães, G.R.; Maciel, M.H.C.; da Silva, L.F.; Agamez-Montalvo, G.S.; da Silva, I.R.; Bezerra, J.D.P.; Souza-Motta, C.M.; Moreira, K.A. Fungal endophytes from leaves of Mandevilla catimbauensis (Apocynaceae): Diversity and potential for L-asparaginase production. Braz. J. Microbiol. 2021, 52, 1431–1441. [Google Scholar] [CrossRef]
- Gibson, E.; Zimmerman, N.B. Urban biogeography of fungal endophytes across San Francisco. PeerJ 2023, 11, e15454. [Google Scholar] [CrossRef]
- von Petrak, F. Über die Gattungen Eriosporopsis n. gen. und Eriospora Berk. et Br. Sydowia 1947, 1, 94–97. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Palacios, J.M.; Árelavol, J.R.; Balguerías, E.; Barone, R.; Delgado, J.D.; de Nascimiento, L.; Elias, R.B.; Fernández-Lugo, S.; Méndez, J.; Naranjo-Cigala, A.; et al. La Laurisilva. Canarias, Madeira y Azores; Macaronesia Editorial: Santa Cruz de Tenerife, Spain, 2017; 417p. [Google Scholar]
- Müller, E.R.; Hütter, R. Quelques discomycètes méditerranéens. Rev. Mycol. 1962, 27, 69–75. [Google Scholar]
- Kohn, L.M.; Korf, R.T. Variation in ascomycete iodine reactions: KOH pretreatment explored. Mycotaxon 1975, 3, 165–172. [Google Scholar]
- Galán, R. Estudos micologicos en el parque natural de Monfragüe (Extremadura, España) V. Leotiales (=Helotiales auct.); Ascomycotina. Crypt. Mycol. 1991, 12, 257–291. [Google Scholar]







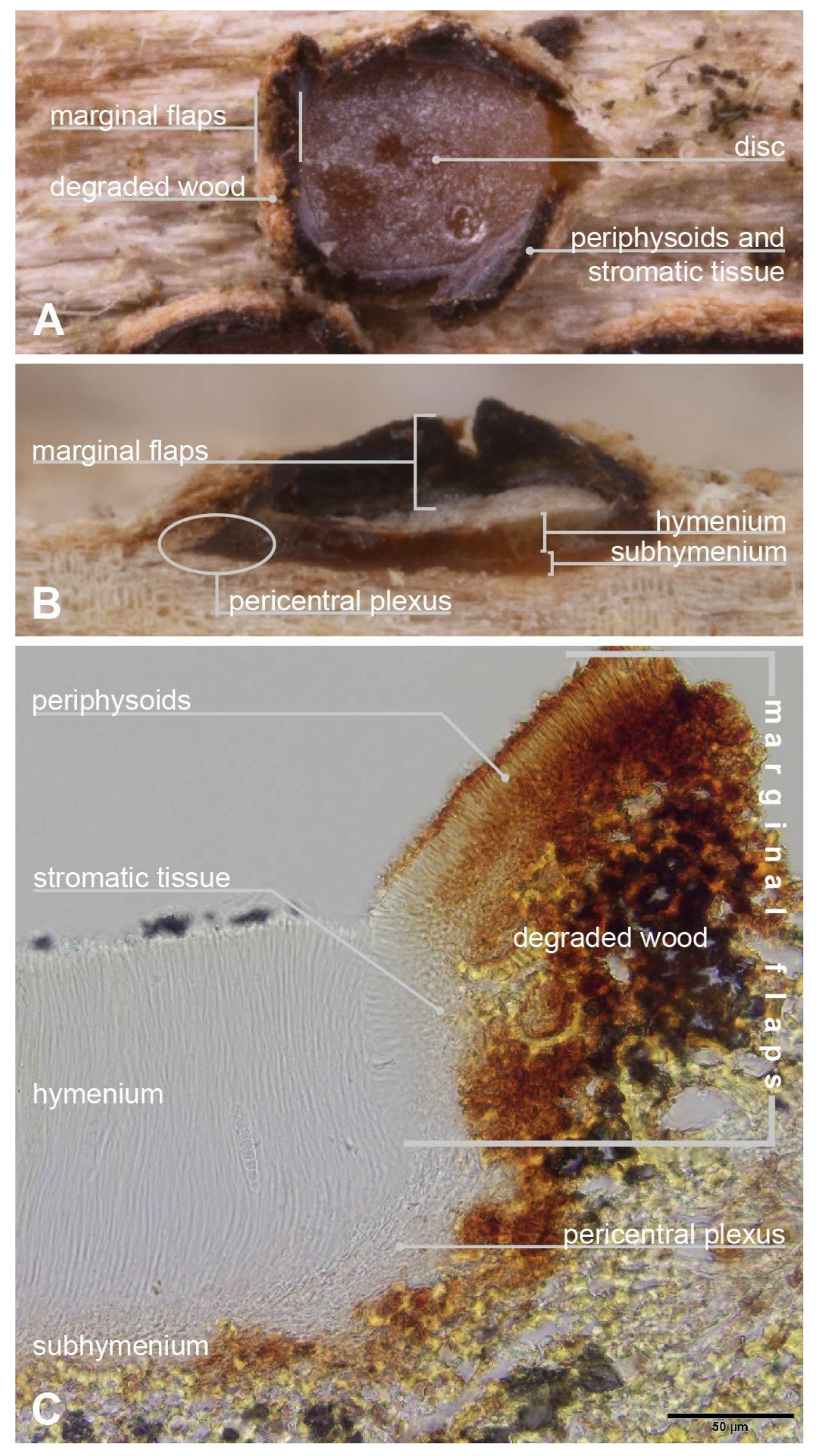{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
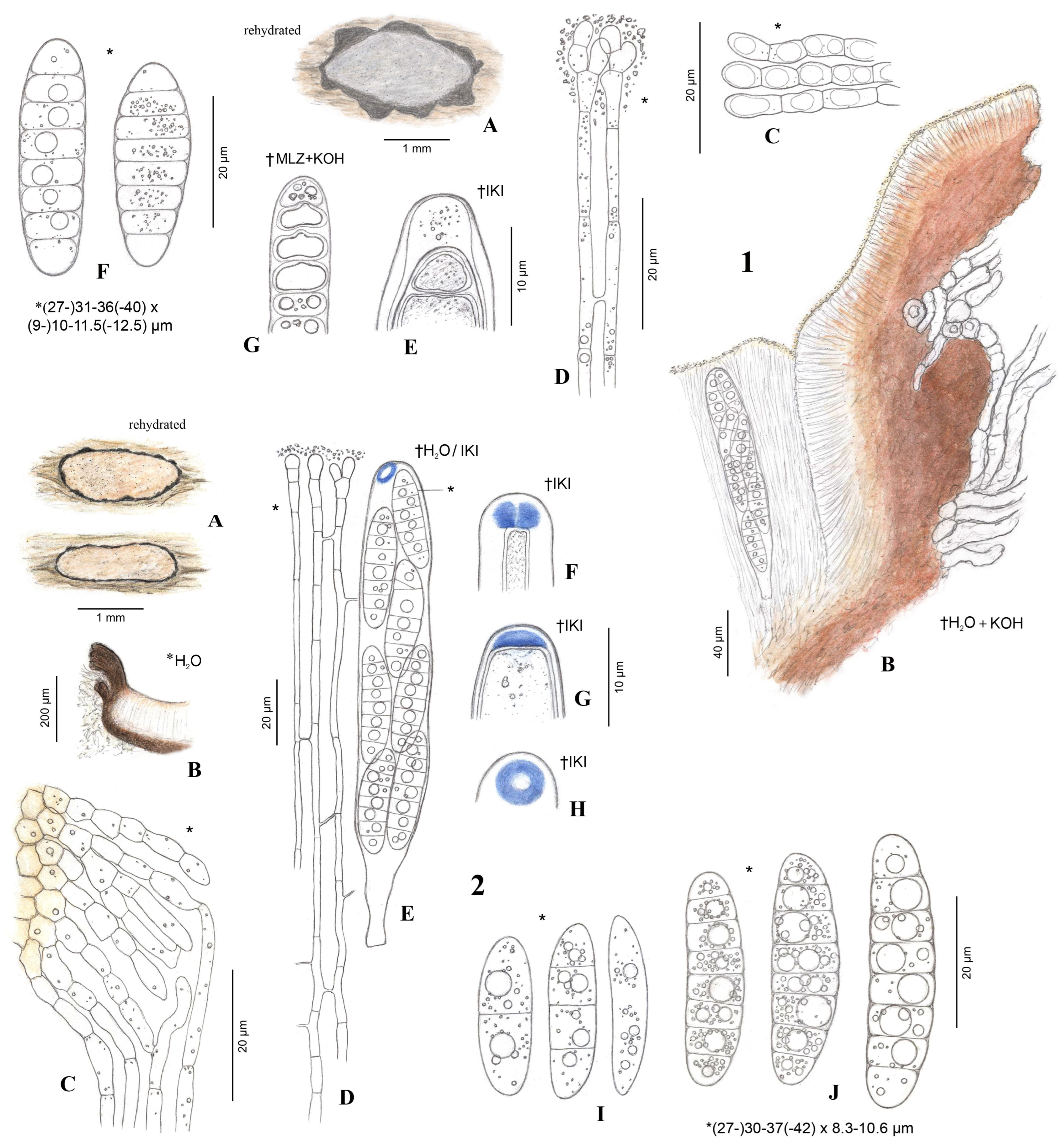{kind=link}
| Number | Name | Herbarium No./Barcode | Culture | SSU | ITS | LSU | mtSSU | Plant Associate | Country |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Bulgaria inquinans | NA | CBS 118.31 | AJ224362 | NA | DQ470960 | NA | NA | NA |
| 2 | Chaetomella oblonga | BPI 843553 | CBS 110.78 | AY487084 | AY487082 | AY487083 | NA | NA | NA |
| 3 | Coccomyces dentatus | OSC 100021 | NA | AY544701 | DQ491499 | AY544657 | AY544736 | NA | NA |
| 4 | Coccomyces proteae | CBS H-20681 | CBS 111703 | NA | JN712451 | JN712515 | NA | NA | NA |
| 5 | Cyclaneusma minus | NA | CBS 496.73 | FJ176812 | NR_153910 | FJ176868 | FJ190629 | Pinus radiata | Chile |
| 6 | Cyclaneusma minus | PDD 64257 | ICMP 17358 | KJ606669 | KJ606680 | KJ606674 | MH698452 | Pinus radiata | New Zealand |
| 7 | Cyclaneusma niveum | NA | CBS 495.73 | PP576077 | PP576096 | PP576086 | PP554500 | Pinus nigra | Germany |
| 8 | Cyclaneusma niveum | ILLS 00122400 | CBS 149209 | PP576070 | PP576089 | PP576079 | PP554493 | Pinus sylvestris | Germany |
| 9 | Leotia lubrica | OSC 100001 | NA | NG_013133 | DQ491484 | AY544644 | AY544746 | NA | NA |
| 10 | Ramomarthamyces octomerus | ILLS 00122394 | CBS 150560 | PP576071 | PP576090 | PP576080 | PP554494 | Laurus novocanariensis | Spain, Canary Islands |
| 11 | Ramomarthamyces octomerus | ILLS 00122395 | CBS 150559 | PP576072 | PP576091 | PP576081 | PP554495 | Olea europaea | Portugal |
| 12 | Lophodermium pinastri | NA | CBS 323.50 | AF106014 | NA | AY004334 | AF431957 | NA | NA |
| 13 | Marthamyces cf emarginatus | PDD 81847 | ICMP 22621 | MK681770 | MK599212 | MK599204 | MK598752 | Eucalyptus sp. | Australia |
| 14 | Marthamyces culmigenus | TNS F-41728 | NA | NA | AB745435 | AB745437 | AB745436 | Miscanthus sinensis | Japan |
| 15 | Marthamyces desmoschoeni | PDD 61761 | ICMP 17350 | KJ606670 | KJ606679 | KJ606673 | OM201128 | Desmoschoenus spiralis | New Zealand |
| 16 | Marthamyces emarginatus | PDD 81846 | ICMP 22854 | MK681769 | MH921869 | MK599203 | MK598751 | Eucalyptus sp. | Australia |
| 17 | Marthamyces harakeke | PDD 108765 | NA | MK681771 | MK599213 | MK599205 | MK598753 | Phormium tenax | New Zealand |
| 18 | Marthamyces maccormackii | PDD 112256 | ICMP 15829 | OM201123 | MK599217 | OM201133 | OM201130 | Metrosideros collina | Cook Islands |
| 19 | Marthamyces metrosideri | PDD 82920 | ICMP 17398 | MH682226 | MH682227 | HM140558 | HM143831.2 | Metrosideros robusta | New Zealand |
| 20 | Marthamyces quadrifidus | PDD 43971 | ICMP 18329 | MK681772 | MK599214 | HM140559 | HM143832 | Weinmannia racemosa | New Zealand |
| 21 | Marthamyces renga | PDD 112257 | ICMP 15830 | OM201124 | MK599221 | OM201134 | OM201127 | Metrosideros collina | Cook Islands |
| 22 | Microglossum rufum | OSC 100641 | NA | DQ471033 | NA | DQ470981 | NA | NA | NA |
| 23 | Naemacyclus fimbriatus | NA | CBS 289.61 | PP576078 | PP576097 | PP576087 | PP554501 | NA | Switzerland |
| 24 | Naemacyclus fimbriatus | CBS H-20006 | CBS 122316 | NA | PP576098 | PP576088 | PP554502 | Pinus pinea | Spain |
| 25 | Naemacyclus fimbriatus | ILLS 00122402 | CBS 149212 | PP576073 | PP576092 | PP576082 | PP554496 | Pinus banksiana | USA, Maine |
| 26 | Naemacyclus fimbriatus | ILLS 00122403 | CBS 149213 | PP576074 | PP576093 | PP576083 | PP554497 | Pinus rigida | USA, Rhode Island |
| 27 | Naemacyclus fimbriatus | ILLS 00122404 | CBS 149214 | PP576075 | PP576094 | PP576084 | PP554498 | Pinus banksiana | USA, Maine |
| 28 | Phacidium pseudophacidioides | NA | CBS 590.69 | NA | KJ663853 | KJ663894 | NA | NA | NA |
| 29 | Pilidium acerinum | BPI 843555 | CBS 736.68 | AY487093 | NA | AY487092 | NA | NA | NA |
| 30 | Potebniamyces pyri | NA | NA | DQ470997 | DQ491510 | DQ470949 | NA | NA | NA |
| 31 | Propolis farinosa | PDD 62678 | ICMP 17354 | MH682223 | MH682229 | HM140562 | HM143834 | Myrsine chathamica | New Zealand |
| 32 | Propolis farinosa | ILLS 00122401 | NA | PP576076 | PP576095 | PP576085 | PP554499 | Fagus sylvatica | Austria |
| 33 | Ramomarthamyces barbatus | PDD 81891 | ICMP 22853 | MK681773 | MH921868 | MK599206 | MK598749.2 | Eucalyptus sp. | Australia |
| 34 | Ramomarthamyces dracophylli | PDD 44691 | ICMP 17381 | NA | MH682228 | NA | MH698450 | Dracophyllum sinclairii | New Zealand |
| 35 | Ramomarthamyces gilvus | PDD 81857 | ICMP 22855 | OM201125 | MH921870 | OM201132 | OM201129 | Caustis sp. | Australia |
| 36 | Ramomarthamyces tuku | PDD 62161 | ICMP 22562 | OM201126 | MK599226 | MK599208 | MK598750 | Juncus sp. | New Zealand |
| 37 | Triblidiopsis pinastri | NA | CBS 445.71 | DQ471035 | NA | DQ470983 | AF431963 | NA | NA |
| 38 | Xeropilidium dennisii | TU 104524 | NA | KX090859 | NA | KX090807 | NA | NA | NA |
| Spain, Canary Islands, La Gomera, Parque Nacional de Garajonay | Europe | |||||
|---|---|---|---|---|---|---|
| H.B. 6986 (Chorros de Epina) | ILLS 00122394, TFC Mic. 25521, RN-14041504, RN-24012001 (Raso de la Bruma) | RN-23102901 (Los Baranquillos) | TFC Mic. 10068 (Agua de Los Llanos) | H.B. 6690 (Croatia, Korčula) | ILLS 00122395 (Portugal, Algarve) | |
| Date | 28 April 2001 | 18 March 2023, 15 April 2014, 20 January 2024 | 29 October 2023 | 28 April 2001 | 8 June 2000 | 22 February2023 |
| Plant associate | Laurus novocanariensis | Laurus novocanariensis | Laurus novocanariensis | Laurus novocanariensis | Olea europaea | Olea europaea |
| Elevation | 1000 | 1070–1090 | 1000 | 1000 | 45 | 200 |
| Apothecium diameter | 1.8–2.5 × 1.2–1.5 | 0.7–2 × 0.6–1.3 | 1.3–2.2 × 0.5–1 | 0.9–1.6 × 0.5–1 | ||
| Ascospores | * 27–40 × 9–12.5 † 24–37 × 8.5–12 | * (31.5–)37–43(–45) × 9.5–11.2 † 27.3–43.3 × 7.5–10.5 | † 33–44.5 × 10–13 | * 27–42 × 8.3–10.6 † (19–)23.5–36.5 × (7–)8–9.5 | * 28.3–52 × 7.4–10.7 † 26.6–42.5 × 7.5–10.5 | |
| Ascospores, number of septa | 7(–9) | (3–)7(–8) | (6–)7(–9) | (3–6–)7 | 3–7 | |
| Asci | † 125–177 × 17–25 | † 136–177 × 17–23 | † 93–140 × 12–21 | † 127–167 × 15–21 | ||
| Ascus apical wall thickness | † 0.2 | † 0.4–0.5 | † ~0.4–0.5 | † 3.3 → 2–2.2 | † 4–4.5 → 1.8–2.6 | |
| Ascus apex in MLZ | Inamyloid | Inamyloid | Amyloid (pale to strong bright blue 3) | Inamyloid or amyloid (light to bright blue 3) | ||
| Ascus apex in IKI | Inamyloid | Inamyloid | Inamyloid? | Inamyloid | Amyloid (strong blue and pale, dingy red 4) | Amyloid (strong dingy to bright blue 5) |
| Ascus apex in KOH pretreat + MLZ | Inamyloid | Inamyloid or weakly amyloid 1 | Amyloid (strong, bright blue) | Amyloid (strong, bright blue) | ||
| Ascus apex in KOH pretreat + IKI | Inamyloid | Amyloid? 2 | Inamyloid? | Inamyloid | Amyloid (strong, bright blue) | Amyloid (strong, bright blue) |
| Paraphyses apical cell | * 3.5–6 × 2.5–3.2 | † 3.5–6 × 1.3–2.4 | * 4–7.5 × 3.5–4.5 | † 4–7.5 × 1.4–2 | ||
| Periphysoids | * 25–40 × 3–4 | * 25–33 × 2.5–3 † 22–36 × 1.8–3 | * 22–25 × 2.5–3.5 | † 17–24 × 2.1 | ||
| Periphysoids, terminal cell | * 5–9 × 3–4 | * 4.5–7 × 2.5–3 | * 5.5–12 × 2.5–3.5 | † 5.5–12 × 2.1 | ||
| Stromatic tissue thickness | 20–52 | 6–23 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karakehian, J.M.; Quijada, L.; Miller, A.N.; Krieglsteiner, L.; Baral, H.-O. Ramomarthamyces octomerus sp. nov. and Insights into the Evolution and Diversification of Ramomarthamyces (Ascomycota, Leotiomycetes, Marthamycetales). J. Fungi 2024, 10, 301. https://doi.org/10.3390/jof10050301
Karakehian JM, Quijada L, Miller AN, Krieglsteiner L, Baral H-O. Ramomarthamyces octomerus sp. nov. and Insights into the Evolution and Diversification of Ramomarthamyces (Ascomycota, Leotiomycetes, Marthamycetales). Journal of Fungi. 2024; 10(5):301. https://doi.org/10.3390/jof10050301
Chicago/Turabian StyleKarakehian, Jason M., Luis Quijada, Andrew N. Miller, Lothar Krieglsteiner, and Hans-Otto Baral. 2024. "Ramomarthamyces octomerus sp. nov. and Insights into the Evolution and Diversification of Ramomarthamyces (Ascomycota, Leotiomycetes, Marthamycetales)" Journal of Fungi 10, no. 5: 301. https://doi.org/10.3390/jof10050301