
Silencing of Sporothrix schenckii GP70 Reveals Its Contribution to Fungal Adhesion, Virulence, and the Host–Fungus Interaction
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing Conditions
2.2. Molecular Techniques
2.3. 3-Carboxy-cis,cis-Muconate Cyclase Activity Assays
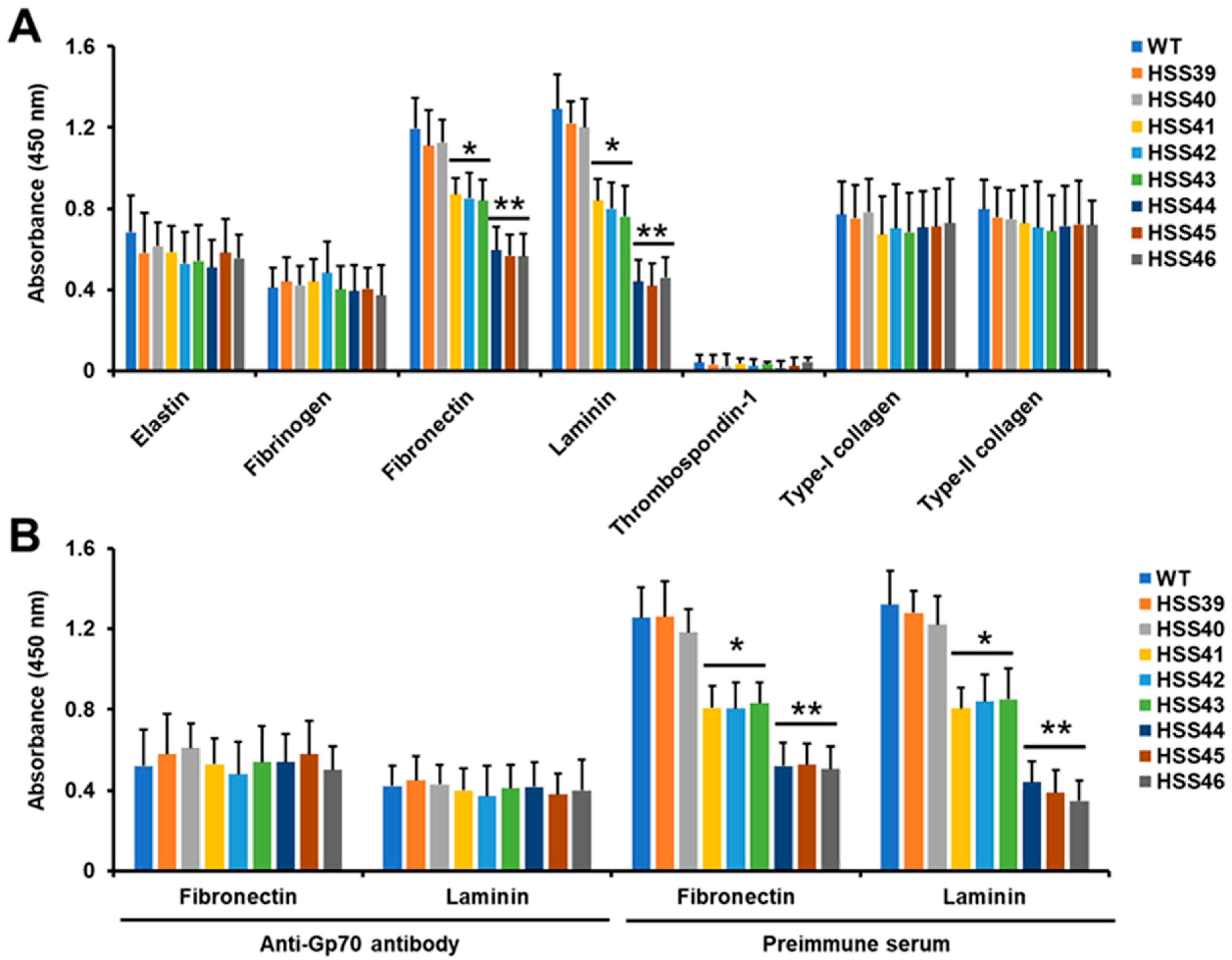
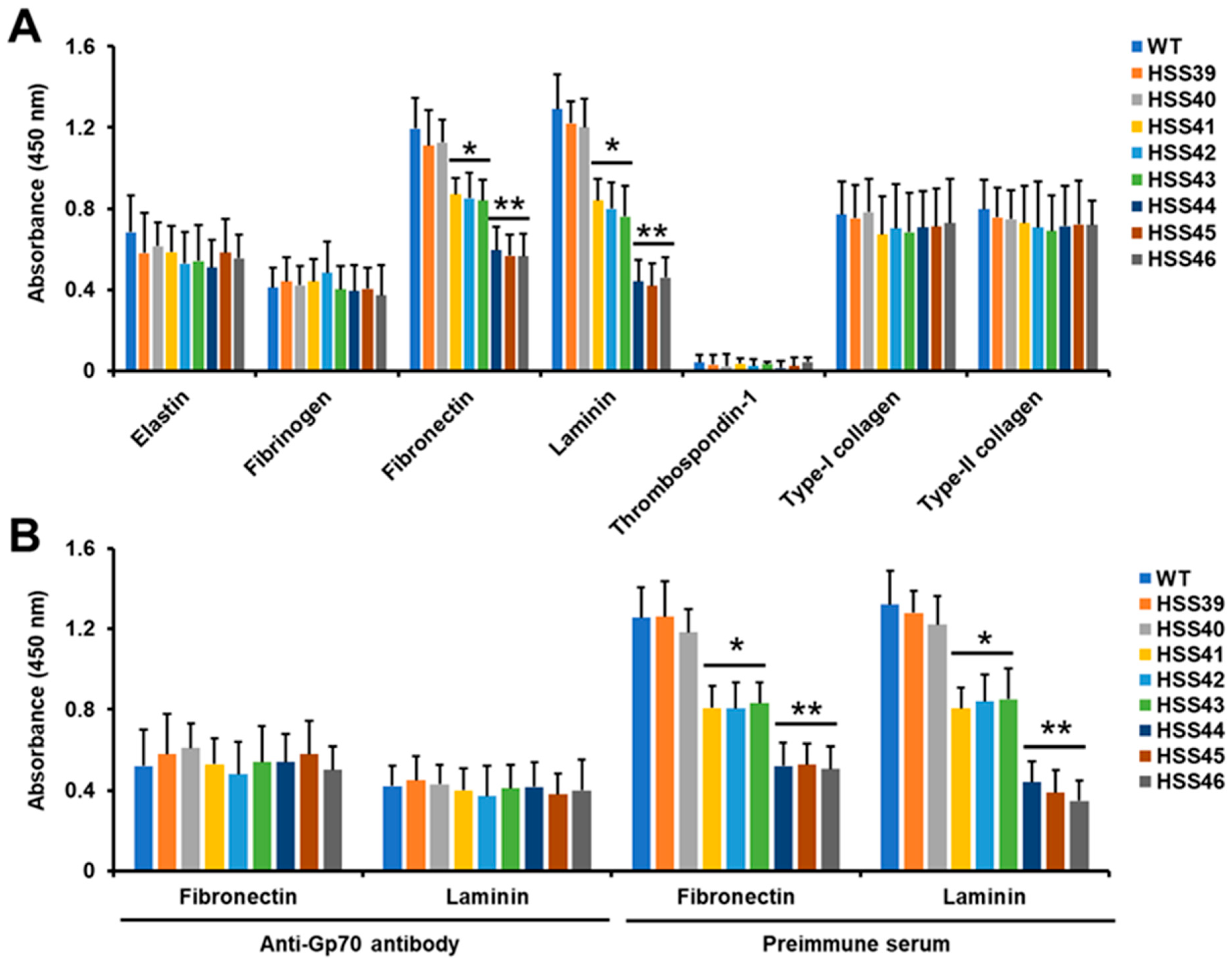
2.4. Adhesion Assays
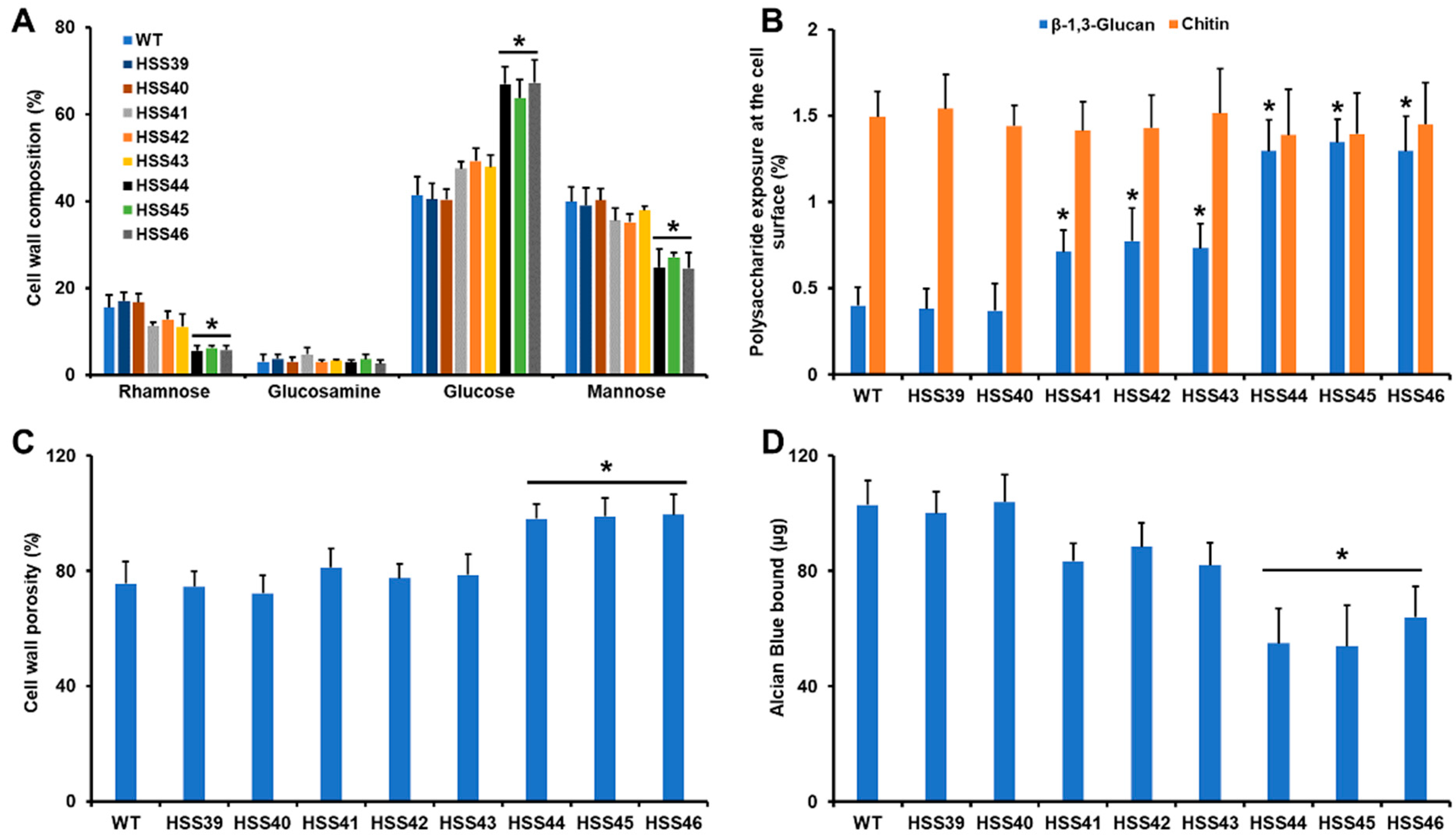
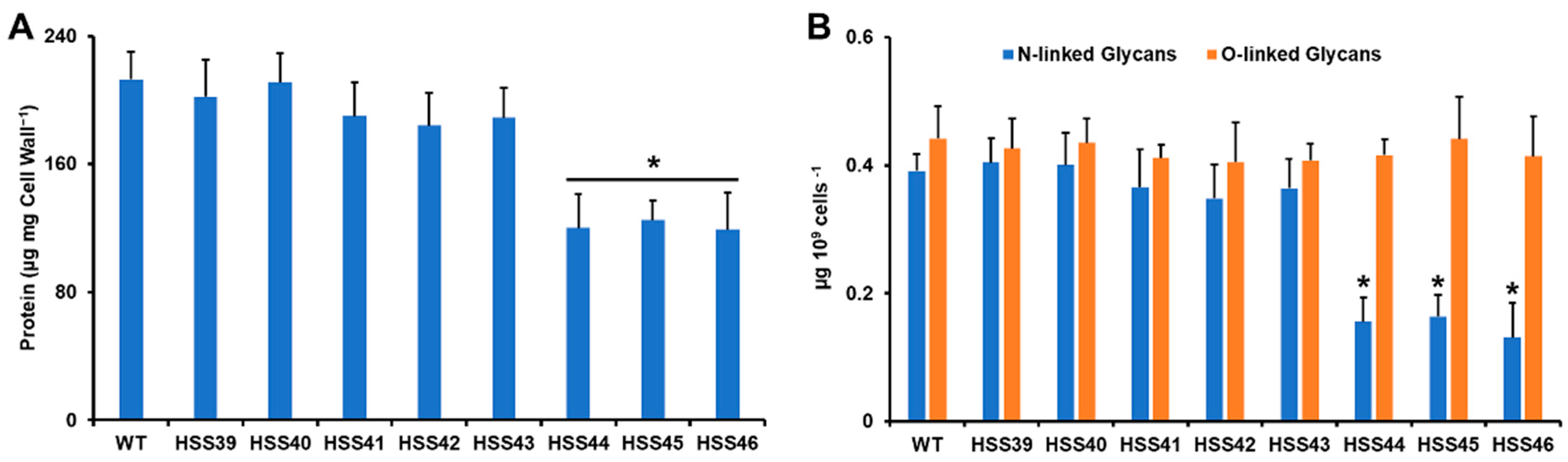
2.5. Cell Wall Analysis
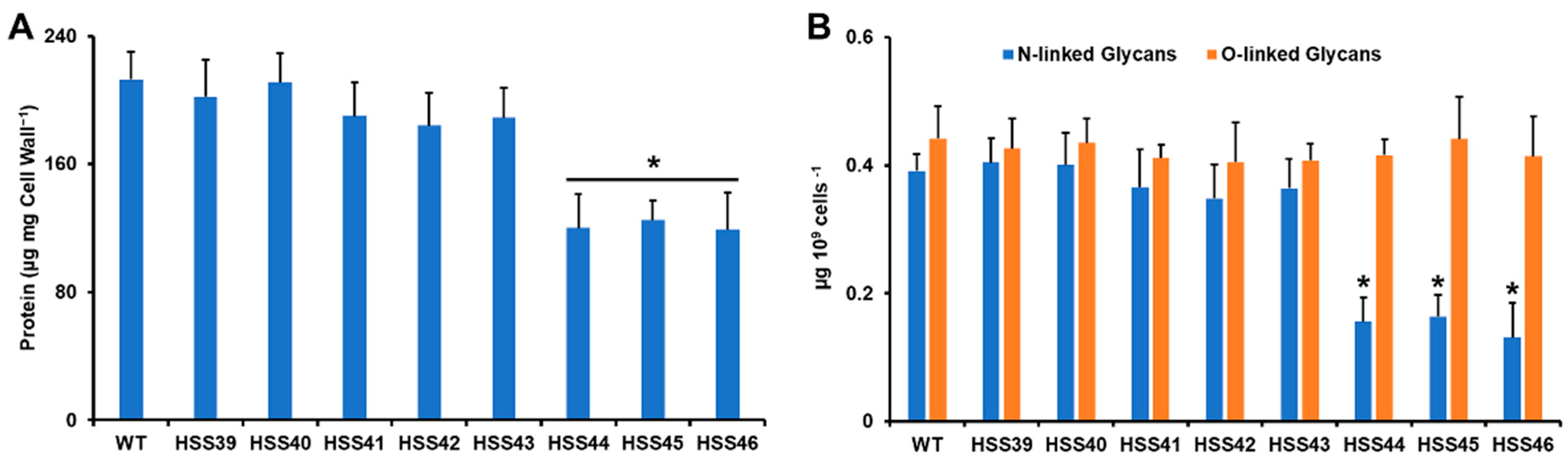
2.6. Quantification of Cell Wall N-Linked and O-Linked Glycans
2.7. Ethics Statement
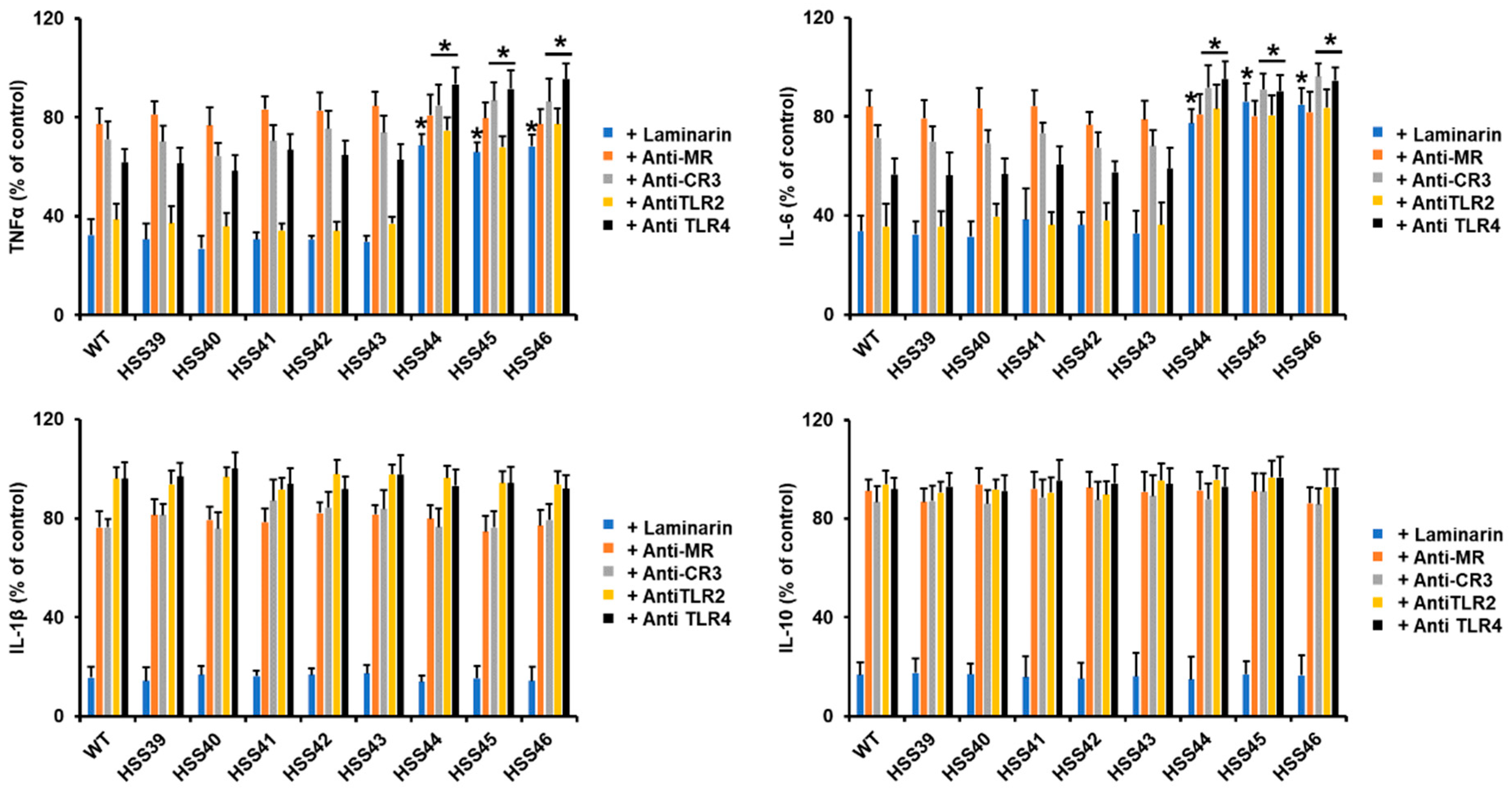
2.8. Analysis of the Fungus–Human Mononuclear Cell Interaction
2.9. Analysis of Phagocytosis by Human Granulocytes
2.10. Quantification of Neutrophils Extracellular Traps
2.11. Virulence Assays in Galleria mellonella
2.12. Statistical Analysis
3. Results
3.1. Silencing of Sporothrix schenckii GP70
3.2. Silencing of Sporothrix schenckii GP70 Affected the 3-Carboxy-cis,cis-Muconate Cyclase Activity and Cell Adhesion
3.3. Silencing of Sporothrix schenckii GP70 Affected the Cell Wall Composition and Organization
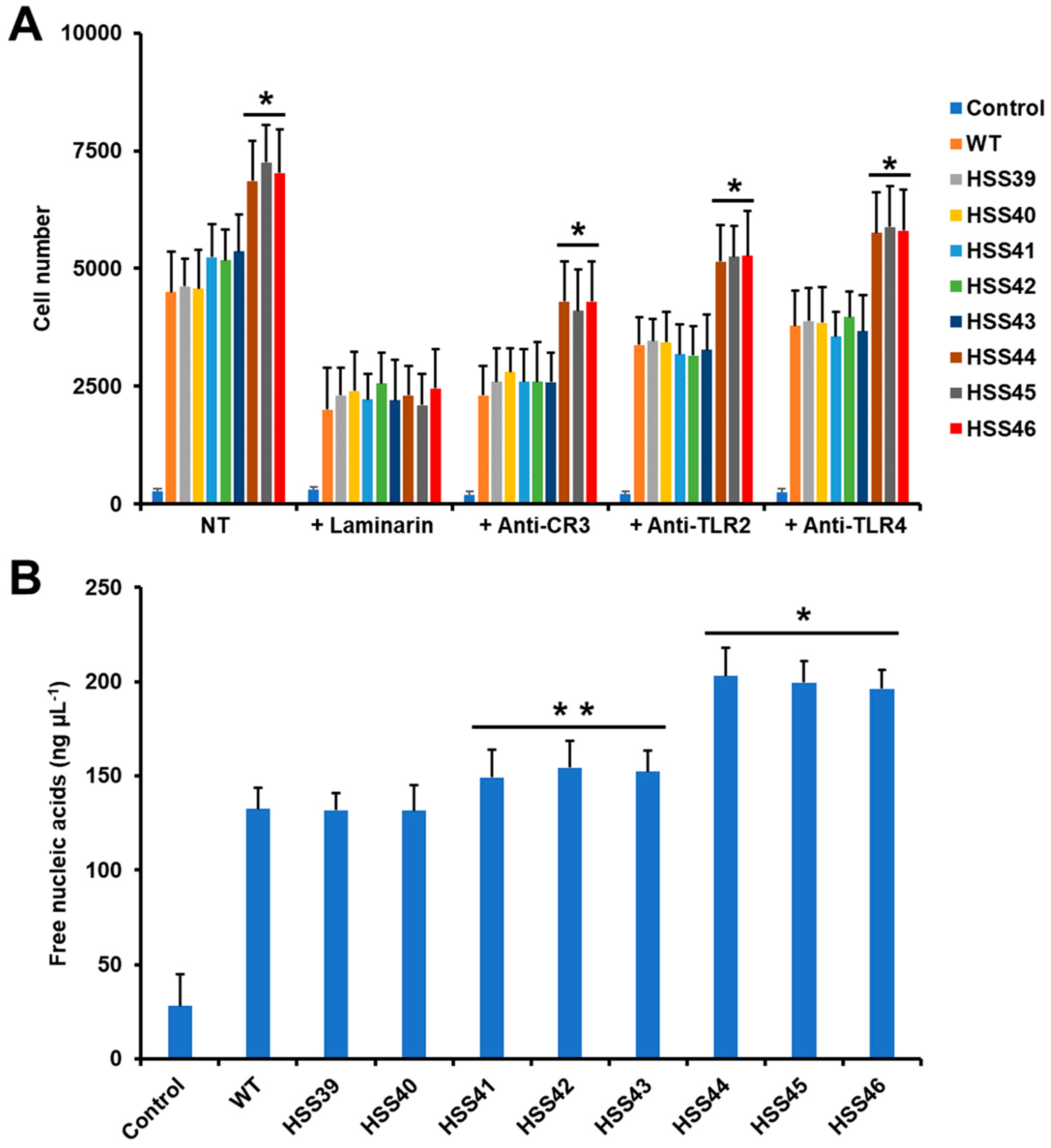
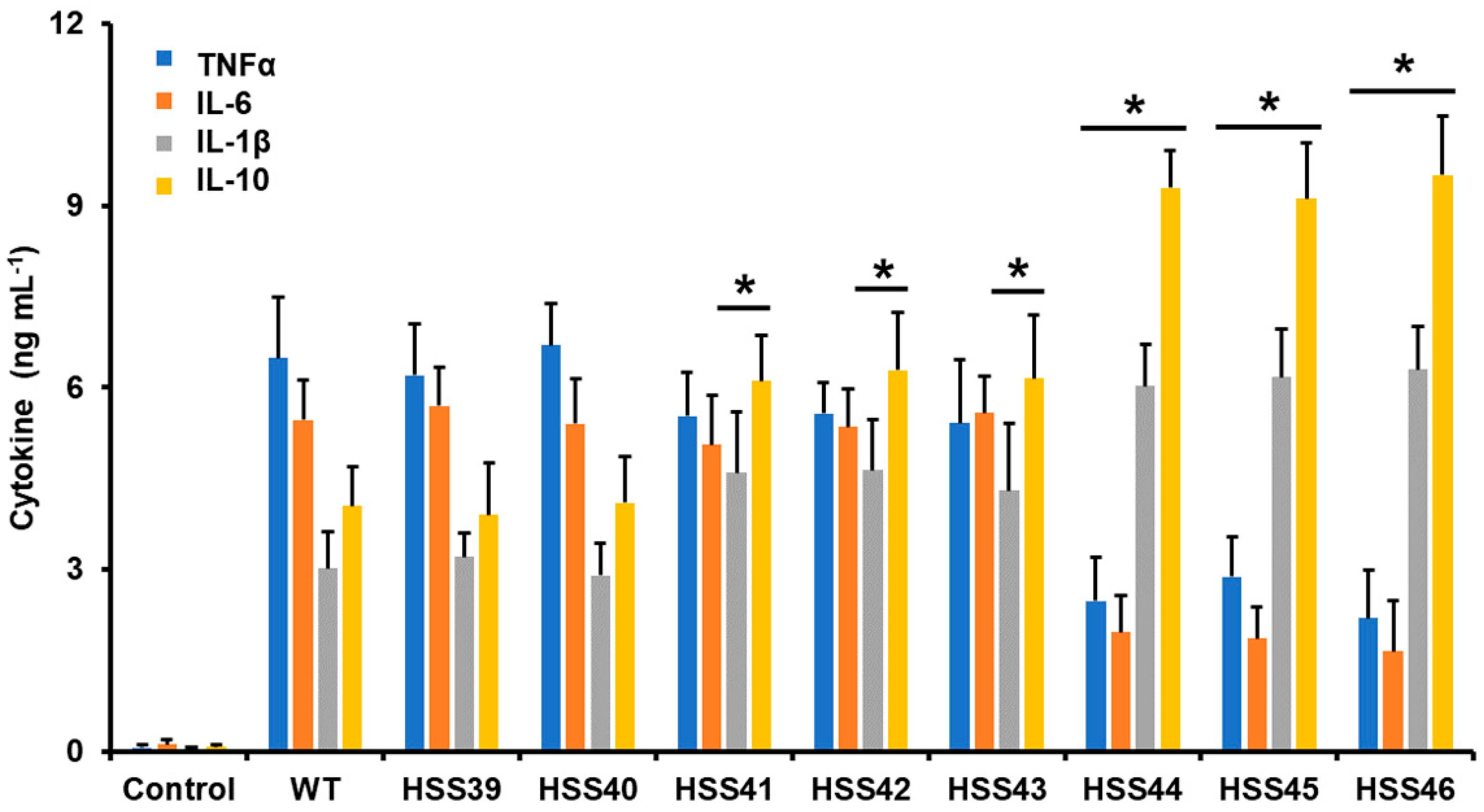
3.4. The GP70 Silencing Affected the Human Innate Immune Cell–Sporothrix schenckii Interaction
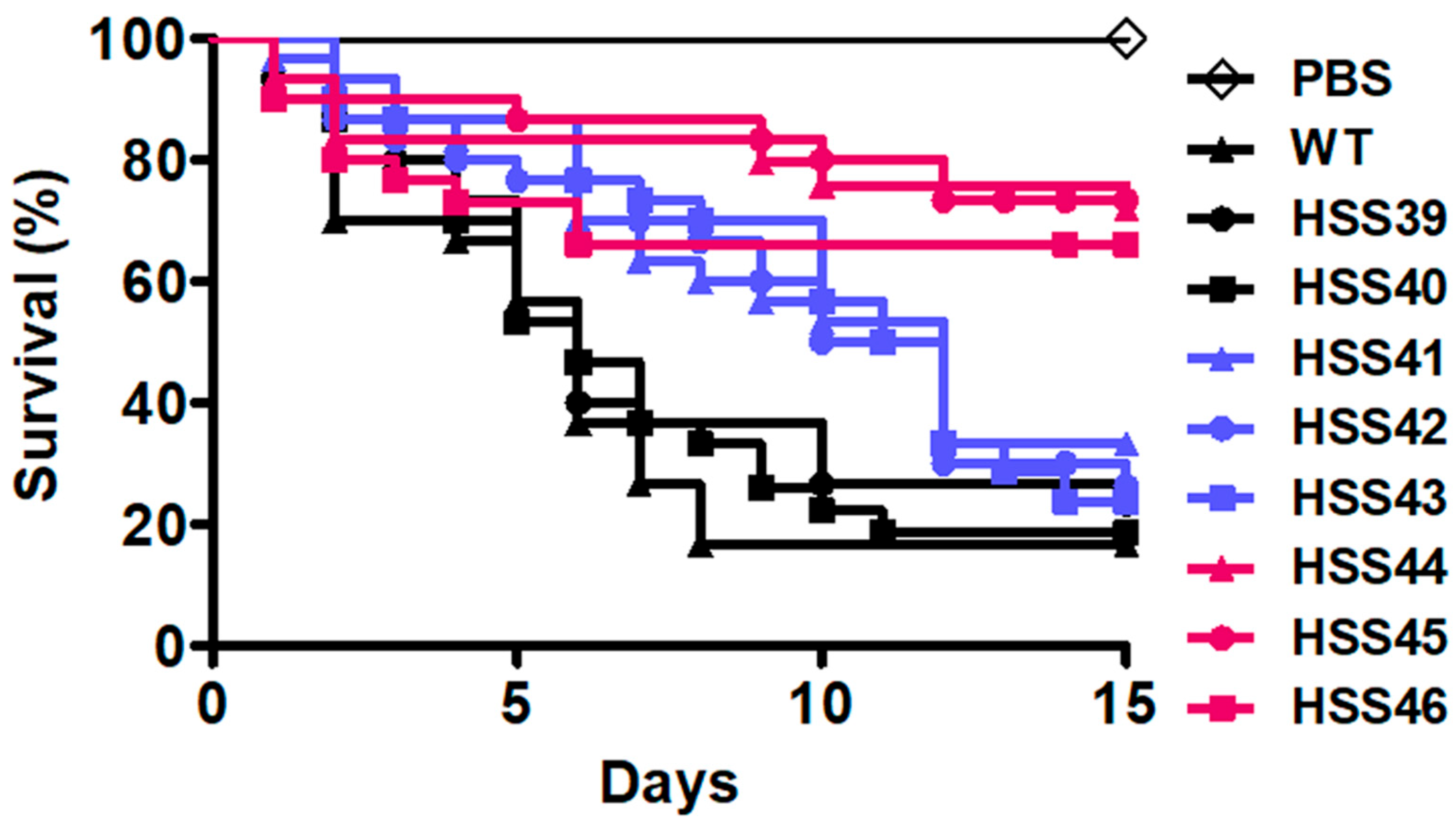
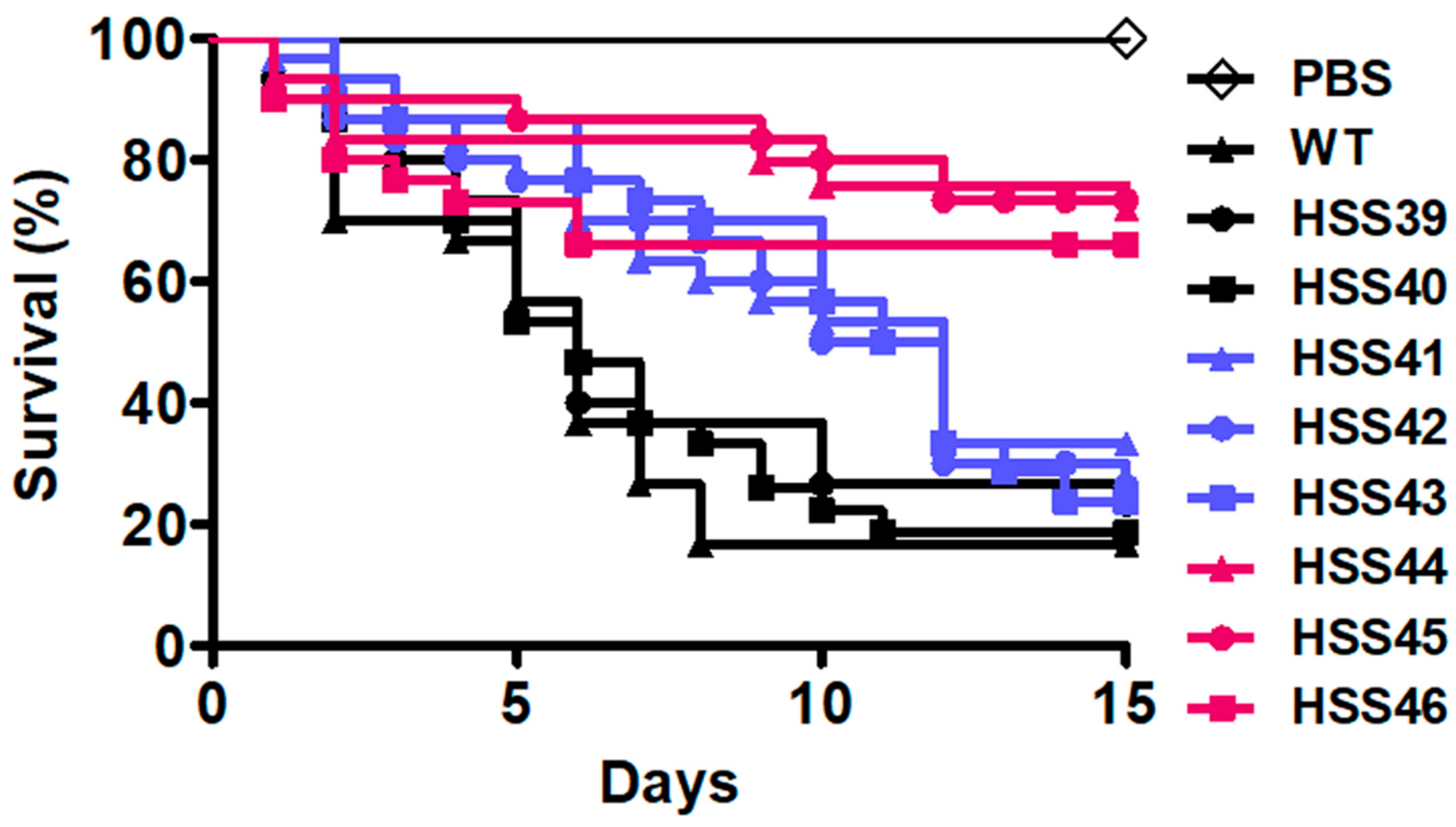
3.5. Virulence Attenuation in the Sporothrix schenckii GP70-Silenced Mutant Strains
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Beer, Z.W.; Duong, T.A.; Wingfield, M.J. The divorce of Sporothrix and Ophiostoma: Solution to a problematic relationship. Stud. Mycol. 2016, 83, 165–191. [Google Scholar] [CrossRef] [PubMed]
- de Beer, Z.W.; Procter, M.; Wingfield, M.J.; Marincowitz, S.; Duong, T.A. Generic boundaries in the Ophiostomatales reconsidered and revised. Stud. Mycol. 2022, 101, 57–120. [Google Scholar] [CrossRef] [PubMed]
- Gremião, I.D.F.; Martins da Silva da Rocha, E.; Montenegro, H.; Carneiro, A.J.B.; Xavier, M.O.; de Farias, M.R.; Monti, F.; Mansho, W.; de Macedo Assunção Pereira, R.H.; Pereira, S.A.; et al. Guideline for the management of feline sporotrichosis caused by Sporothrix brasiliensis and literature revision. Braz. J. Microbiol. 2021, 52, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Baes Pereira, S.; Dos Reis Gomes, A.; Bressan Waller, S.; Batista Xavier, J.R.; Messias Rodrigues, A.; Kutscher Ripoll, M.; Ferreira, M.R.A.; Rochedo Conceição, F.; Osório de Faria, R.; Pascoti Bruhn, F.R. Sporotrichosis in dogs: Epidemiological and clinical-therapeutic profile and the emergence of itraconazole-resistant isolates. Med. Mycol. 2022, 60, myac089. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef]
- Bonifaz, A.; Vázquez-González, D. Diagnosis and treatment of lymphocutaneous sporotrichosis: What are the options? Curr. Fungal Infect. Rep. 2013, 7, 252–259. [Google Scholar] [CrossRef]
- Vásquez-del-Mercado, E.; Arenas, R.; Padilla-Desgarenes, C. Sporotrichosis. Clin. Dermatol. 2012, 30, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Host-pathogen interactions: Redefining the basic concepts of virulence and pathogenicity. Infect. Immun. 1999, 67, 3703–3713. [Google Scholar] [CrossRef]
- Staniszewska, M. Virulence factors in Candida species. Curr. Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; de Oliveira, L.C.; Oliveira, M.M.E.; Gutierrez-Galhardo, M.C.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Phenotypic characteristics associated with virulence of clinical isolates from the Sporothrix complex. BioMed Res. Int. 2015, 2015, 212308. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 28513415. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Bernal, G.; Barbosa-Sabanero, G.; Shibayama, M.; Perez-Torres, A.; Tsutsumi, V.; Sabanero, M. Cell wall glycoproteins participate in the adhesion of Sporothrix schenckii to epithelial cells. Mycopathologia 2011, 171, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Lima, O.C.; Figueiredo, C.C.; Pereira, B.A.; Coelho, M.G.; Morandi, V.; Lopes-Bezerra, L.M. Adhesion of the human pathogen Sporothrix schenckii to several extracellular matrix proteins. Braz. J. Med. Biol. Res. 1999, 32, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Lima, O.C.; Figueiredo, C.C.; Previato, J.O.; Mendonça-Previato, L.; Morandi, V.; Lopes Bezerra, L.M. Involvement of fungal cell wall components in adhesion of Sporothrix schenckii to human fibronectin. Infect. Immun. 2001, 69, 6874–6880. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, P.A.C.; de Castro, R.A.; Nascimento, R.C.; Tronchin, G.; Pérez Torres, A.; Lazéra, M.; de Almeida, S.R.; Bouchara, J.-P.; Loureiro y Penha, C.V.; Lopes-Bezerra, L.M. Cell surface expression of adhesins for fibronectin correlates with virulence in Sporothrix schenckii. Microbiology 2009, 155, 3730–3738. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Baca, E.; Toriello, C.; Pérez-Torres, A.; Sabanero-López, M.; Villagómez-Castro, J.C.; López-Romero, E. Isolation and some properties of a glycoprotein of 70 kDa (Gp70) from the cell wall of Sporothrix schenckii involved in fungal adherence to dermal extracellular matrix. Med. Mycol. 2009, 47, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Kubitschek-Barreira, P.H.; Fernandes, G.F.; de Almeida, S.R.; Lopes-Bezerra, L.M.; de Camargo, Z.P. Immunoproteomic analysis reveals a convergent humoral response signature in the Sporothrix schenckii complex. J. Proteom. 2015, 115, 8–22. [Google Scholar] [CrossRef]
- Martínez-Álvarez, J.A.; García-Carnero, L.C.; Kubitschek-Barreira, P.H.; Lozoya-Pérez, N.E.; Belmonte-Vázquez, J.L.; de Almeida, J.R.; Gómez-Infante, A.d.J.; Curty, N.; Villagómez-Castro, J.C.; Peña-Cabrera, E.; et al. Analysis of some immunogenic properties of the recombinant Sporothrix schenckii Gp70 expressed in Escherichia coli. Future Microbiol. 2019, 14, 397–410. [Google Scholar] [CrossRef]
- Castro, R.A.; Kubitschek-Barreira, P.H.; Teixeira, P.A.C.; Sanches, G.F.; Teixeira, M.M.; Quintella, L.P.; Almeida, S.R.; Costa, R.O.; Camargo, Z.P.; Felipe, M.S.S.; et al. Differences in cell morphometry, cell wall topography and Gp70 expression correlate with the virulence of Sporothrix brasiliensis clinical isolates. PLoS ONE 2013, 8, e75656. [Google Scholar] [CrossRef]
- de Almeida, J.R.; Santiago, K.L.; Kaihami, G.H.; Maranhão, A.Q.; de Macedo Brígido, M.; de Almeida, S.R. The efficacy of humanized antibody against the Sporothrix antigen, gp70, in promoting phagocytosis and reducing disease burden. Front. Microbiol. 2017, 8, 345. [Google Scholar] [CrossRef]
- de Almeida, J.R.; Kaihami, G.H.; Jannuzzi, G.P.; de Almeida, S.R. Therapeutic vaccine using a monoclonal antibody against a 70-kDa glycoprotein in mice infected with highly virulent Sporothrix schenckii and Sporothrix brasiliensis. Med. Mycol. 2015, 53, 42–50. [Google Scholar] [CrossRef]
- de Almeida, J.R.F.; Jannuzzi, G.P.; Kaihami, G.H.; Breda, L.C.D.; Ferreira, K.S.; de Almeida, S.R. An immunoproteomic approach revealing peptides from Sporothrix brasiliensis that induce a cellular immune response in subcutaneous sporotrichosis. Sci. Rep. 2018, 8, 4192. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Jiang, R.; Wang, Y.; Zhu, M.; Zhang, X.; Dong, S.; Shi, H.; Wang, L. Recombinant phage elicits protective immune response against systemic S. globosa infection in mouse model. Sci. Rep. 2017, 7, 42024. [Google Scholar] [CrossRef]
- Teixeira, M.M.; de Almeida, L.G.; Kubitschek-Barreira, P.; Alves, F.L.; Kioshima, E.S.; Abadio, A.K.; Fernandes, L.; Derengowski, L.S.; Ferreira, K.S.; Souza, R.C.; et al. Comparative genomics of the major fungal agents of human and animal sporotrichosis: Sporothrix schenckii and Sporothrix brasiliensis. BMC Genom. 2014, 15, 943. [Google Scholar] [CrossRef] [PubMed]
- Tamez-Castrellón, A.K.; van der Beek, S.L.; López-Ramírez, L.A.; Martínez-Duncker, I.; Lozoya-Pérez, N.E.; van Sorge, N.M.; Mora-Montes, H.M. Disruption of protein rhamnosylation affects the Sporothrix schenckii-host interaction. Cell Surf. 2021, 7, 100058. [Google Scholar] [CrossRef]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Stone, M.; Schlagnhaufer, C.; Romaine, C.P. A fruiting body tissue method for efficient Agrobacterium-mediated transformation of Agaricus bisporus. Appl. Environ. Microbiol. 2000, 66, 4510–4513. [Google Scholar] [CrossRef]
- Nakayashiki, H.; Hanada, S.; Nguyen, B.Q.; Kadotani, N.; Tosa, Y.; Mayama, S. RNA silencing as a tool for exploring gene function in ascomycete fungi. Fungal Genet. Biol. 2005, 42, 275–283. [Google Scholar] [CrossRef]
- Trujillo-Esquivel, E.; Martínez-Álvarez, J.A.; Clavijo-Giraldo, D.M.; Hernández, N.V.; Flores-Martínez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. The Sporothrix schenckii gene encoding for the ribosomal protein L6 has constitutive and stable expression and works as an endogenous control in gene expression analysis. Front. Microbiol. 2017, 8, 1676. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, D.R.; Cain, R.B. Metabolism of aromatic compounds by fungi. Kinetic properties and mechanism of 3-carboxy-cis,cis-muconate cyclase from Aspergillus niger. Eur. J. Biochem. 1975, 56, 193–204. [Google Scholar] [CrossRef] [PubMed]
- García-Carnero, L.C.; Salinas-Marín, R.; Lozoya-Pérez, N.E.; Wrobel, K.; Wrobel, K.; Martínez-Duncker, I.; Niño-Vega, G.A.; Mora-Montes, H.M. The Heat shock protein 60 and Pap1 participate in the Sporothrix schenckii-host interaction. J. Fungi 2021, 7, 960. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Diaz-Jimenez, D.F.; Lopez-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.; Odds, F.C.; et al. Endoplasmic reticulum alpha-glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Marakalala, M.J.; Vautier, S.; Potrykus, J.; Walker, L.A.; Shepardson, K.M.; Hopke, A.; Mora-Montes, H.M.; Kerrigan, A.; Netea, M.G.; Murray, G.I.; et al. Differential adaptation of Candida albicans in vivo modulates immune recognition by dectin-1. PLoS Pathog. 2013, 9, e1003315. [Google Scholar] [CrossRef]
- De Nobel, J.G.; Klis, F.M.; Munnik, T.; Priem, J.; Van Den Ende, H. An assay of relative cell wall porosity in Saccharomyces cerevisiae, Kluyveromyces lactis and Schizosaccharomyces pombe. Yeast 1990, 6, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Netea, M.G.; Ferwerda, G.; Lenardon, M.D.; Brown, G.D.; Mistry, A.R.; Kullberg, B.J.; O’Callaghan, C.A.; Sheth, C.C.; Odds, F.C.; et al. Recognition and blocking of innate immunity cells by Candida albicans chitin. Infect. Immun. 2011, 79, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.M.; Tsoni, S.V.; Willment, J.A.; Williams, D.L.; Taylor, P.R.; Gordon, S.; Dennehy, K.; Brown, G.D. Soluble Dectin-1 as a tool to detect beta-glucans. J. Immunol. Methods 2006, 314, 164–169. [Google Scholar] [CrossRef]
- Hobson, R.P.; Munro, C.A.; Bates, S.; MacCallum, D.M.; Cutler, J.E.; Heinsbroek, S.E.; Brown, G.D.; Odds, F.C.; Gow, N.A. Loss of cell wall mannosylphosphate in Candida albicans does not influence macrophage recognition. J. Biol. Chem. 2004, 279, 39628–39635. [Google Scholar] [CrossRef]
- Endres, S.; Ghorbani, R.; Lonnemann, G.; van der Meer, J.W.; Dinarello, C.A. Measurement of immunoreactive interleukin-1 beta from human mononuclear cells: Optimization of recovery, intrasubject consistency, and comparison with interleukin-1 alpha and tumor necrosis factor. Clin. Immunol. Immunopathol. 1988, 49, 424–438. [Google Scholar] [CrossRef]
- Schwartz, S.N.; Medoff, G.; Kobayashi, G.S.; Kwan, C.N.; Schlessinger, D. Antifungal properties of polymyxin B and its potentiation of tetracycline as an antifungal agent. Antimicrob. Agents Chemother. 1972, 2, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Neves, G.W.P.; Wong, S.S.W.; Aimanianda, V.; Simenel, C.; Guijarro, J.I.; Walls, C.; Willment, J.A.; Gow, N.A.R.; Munro, C.A.; Brown, G.D.; et al. Complement-mediated differential immune response of human macrophages to Sporothrix Species through interaction with their cell wall peptidorhamnomannans. Front. Immunol. 2021, 12, 749074. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.E.; Hu, J.C.; Athanasiou, K.A. Ammonium-chloride-potassium lysing buffer treatment of fully differentiated cells increases cell purity and resulting neotissue functional properties. Tissue Eng. Part C Methods 2016, 22, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Chávez, M.J.; Franco, B.; Clavijo-Giraldo, D.M.; Hernández, N.V.; Estrada-Mata, E.; Mora-Montes, H.M. Role of protein phosphomannosylation in the Candida tropicalis–macrophage interaction. FEMS Yeast Res. 2018, 18, foy053. [Google Scholar] [CrossRef] [PubMed]
- White, P.C.; Chicca, I.J.; Ling, M.R.; Wright, H.J.; Cooper, P.R.; Milward, M.R.; Chapple, I.L. Characterization, quantification, and visualization of neutrophil extracellular traps. Methods Mol. Biol. 2017, 1537, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Clavijo-Giraldo, D.M.; Matinez-Alvarez, J.A.; Lopes-Bezerra, L.M.; Ponce-Noyola, P.; Franco, B.; Almeida, R.S.; Mora-Montes, H.M. Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis virulence in Galleria mellonella. J. Microbiol. Methods 2016, 122, 73–77. [Google Scholar] [CrossRef] [PubMed]
- García-Carnero, L.C.; Clavijo-Giraldo, D.M.; Gómez-Gaviria, M.; Lozoya-Pérez, N.E.; Tamez-Castrellón, A.K.; López-Ramírez, L.A.; Mora-Montes, H.M. Early virulence predictors during the Candida species-Galleria mellonella Interaction. J. Fungi 2020, 6, 152. [Google Scholar] [CrossRef] [PubMed]
- López-Ramírez, L.A.; Martínez-Duncker, I.; Márquez-Márquez, A.; Vargas-Macías, A.P.; Mora-Montes, H.M. Silencing of ROT2, the encoding gene of the endoplasmic reticulum glucosidase II, affects the cell wall and the Sporothrix schenckii-host interaction. J. Fungi 2022, 8, 1220. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.H.; Ramírez-Prado, J.H.; Neves, G.W.P.; Torrado, E.; Sampaio, P.; Felipe, M.S.S.; Vasconcelos, A.T.; Goldman, G.H.; Carvalho, A.; Cunha, C.; et al. Ploidy determination in the pathogenic fungus Sporothrix spp. Front. Microbiol. 2019, 10, 284. [Google Scholar] [CrossRef]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef]
- Barros, M.B.d.L.; de Almeida Paes, R.; Schubach, A.O. Sporothrix schenckii and sporotrichosis. Clin. Microbiol. Rev. 2011, 24, 633–654. [Google Scholar] [CrossRef] [PubMed]
- García-Carnero, L.C.; Martínez-Duncker, I.; Gómez-Gaviria, M.; Mora-Montes, H.M. Differential recognition of clinically relevant Sporothrix species by human mononuclear cells. J. Fungi 2023, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gaviria, M.; Martínez-Duncker, I.; García-Carnero, L.C.; Mora-Montes, H.M. Differential recognition of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa by human monocyte-derived macrophages and dendritic cells. Infect. Drug Resist. 2023, 16, 4817–4834. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. Human neutrophils ≠ murine neutrophils: Does it matter? Immunol. Rev. 2023, 314, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Lozoya-Pérez, N.E.; Clavijo-Giraldo, D.M.; Martínez-Duncker, I.; García-Carnero, L.C.; López-Ramírez, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Influences of the culturing media in the virulence and cell wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. J. Fungi 2020, 6, 323. [Google Scholar] [CrossRef] [PubMed]
- Romero-Martinez, R.; Wheeler, M.; Guerrero-Plata, A.; Rico, G.; Torres-Guerrero, H. Biosynthesis and functions of melanin in Sporothrix schenckii. Infect. Immun. 2000, 68, 3696–3703. [Google Scholar] [CrossRef] [PubMed]
- Hatinguais, R.; Leaves, I.; Brown, G.D.; Brown, A.J.P.; Brock, M.; Peres da Silva, R. CRISPR-based tools for targeted genetic manipulation in pathogenic Sporothrix species. Microbiol. Spectr. 2023, 11, e0507822. [Google Scholar] [CrossRef] [PubMed]
- Madrid, I.M.; Xavier, M.O.; Mattei, A.S.; Fernandes, C.G.; Guim, T.N.; Santin, R.; Schuch, L.F.; Nobre Mde, O.; Araújo Meireles, M.C. Role of melanin in the pathogenesis of cutaneous sporotrichosis. Microbes Infect. 2010, 12, 162–165. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; Frases, S.; Araújo Gde, S.; de Oliveira, M.M.; Gerfen, G.J.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Biosynthesis and functions of a melanoid pigment produced by species of the Sporothrix complex in the presence of L-tyrosine. Appl. Environ. Microbiol. 2012, 78, 8623–8630. [Google Scholar] [CrossRef]
- Giosa, D.; Felice, M.R.; Giuffrè, L.; Aiese Cigliano, R.; Paytuví-Gallart, A.; Lo Passo, C.; Barresi, C.; D’Alessandro, E.; Huang, H.; Criseo, G.; et al. Transcriptome-wide expression profiling of Sporothrix schenckii yeast and mycelial forms and the establishment of the Sporothrix Genome DataBase. Microb. Genom. 2020, 6, mgen000445. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, J.F.; González-Ibarra, J.; Macías-Segoviano, J.I.; Cuéllar-Cruz, M.; Álvarez-Vargas, A.; Cano-Canchola, C.; López-Romero, E. Congo Red affects the growth, morphology and activity of glucosamine-6-phosphate synthase in the human pathogenic fungus Sporothrix schenckii. Arch. Microbiol. 2019, 201, 135–141. [Google Scholar] [CrossRef]
- Kennedy, A.D.; Willment, J.A.; Dorward, D.W.; Williams, D.L.; Brown, G.D.; DeLeo, F.R. Dectin-1 promotes fungicidal activity of human neutrophils. Eur. J. Immunol. 2007, 37, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Galván-Hernández, A.K.; Gómez-Gaviria, M.; Martínez-Duncker, I.; Martínez-Álvarez, J.A.; Mora-Montes, H.M. Differential recognition of clinically relevant Sporothrix species by human granulocytes. J. Fungi 2023, 9, 986. [Google Scholar] [CrossRef]
- Villalobos-Duno, H.L.; Barreto, L.A.; Alvarez-Aular, Á.; Mora-Montes, H.M.; Lozoya-Pérez, N.E.; Franco, B.; Lopes-Bezerra, L.M.; Niño-Vega, G.A. Comparison of cell wall polysaccharide composition and structure between strains of Sporothrix schenckii and Sporothrix brasiliensis. Front. Microbiol. 2021, 12, 726958. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Bezerra, L.M.; Walker, L.A.; Niño-Vega, G.; Mora-Montes, H.M.; Neves, G.W.P.; Villalobos-Duno, H.; Barreto, L.; Garcia, K.; Franco, B.; Martínez-Álvarez, J.A.; et al. Cell walls of the dimorphic fungal pathogens Sporothrix schenckii and Sporothrix brasiliensis exhibit bilaminate structures and sloughing of extensive and intact layers. PLoS Negl. Trop. Dis. 2018, 12, e0006169. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relative GP70 Expression (%) a | GP70 Copy Number a | Gp70 Enzyme Activity b |
|---|---|---|---|
| Wild-type | 100.0 ± 0.5 | 1.0 ± 0.1 | 21.2 × 10−2 ± 6.7 × 10−2 |
| HSS39 | 98.2 ± 1.4 | 1.2 ± 0.3 | 19.8 × 10−2 ± 5.2 × 10−2 |
| HSS40 | 99.4 ± 1.2 | 1.4 ± 0.3 | 18.0 × 10−2 ± 8.4 × 10−2 |
| HSS41 | 46.7 ± 1.2 * | 2.9 ± 0.3 * | 7.2 × 10−2 ± 3.4 × 10−2 * |
| HSS42 | 38.6 ± 2.4 * | 2.9 ± 0.4 * | 6.4 × 10−2 ± 4.7 × 10−2 * |
| HSS43 | 37.2 ± 3.4 * | 3.1 ± 0.3 * | 7.7 × 10−2 ± 2.5 × 10−2 * |
| HSS44 | 1.8 ± 0.7 * | 2.8 ± 0.2 * | No detected |
| HSS45 | 12.1 ± 1.6 * | 3.1 ± 0.3 * | 2.4 × 10−2 ± 1.6 × 10−2 * |
| HSS46 | 9.1 ± 0.7 * | 3.1 ± 0.4 * | No detected |
| Strain | Colony-Forming Units (×105) a | Cytotoxicity (%) b | Hemocytes (×106) mL−1 | Melanin c | Phenoloxidase d |
|---|---|---|---|---|---|
| PBS e | 0.0 ± 0.0 | 10.5 ± 3.3 | 3.6 ± 0.4 | 1.1 ± 0.5 | 0.4 ± 0.2 |
| WT f | 3.2 ± 0.8 | 92.3 ± 7.7 | 7.9 ± 0.2 | 5.2 ± 0.4 | 3.6 ± 0.6 |
| HSS39 | 3.1 ± 0.7 | 87.9 ± 10.5 | 7.8 ± 0.6 | 5.2 ± 0.6 | 3.3 ± 0.5 |
| HSS40 | 3.4 ± 0.6 | 90.5 ± 8.4 | 8.2 ± 0.4 | 5.4 ± 0.3 | 3.7 ± 0.2 |
| HSS41 | 2.9 ± 0.7 | 60.3 ± 8.9 *,† | 5.4 ± 0.3 *,† | 3.2 ± 0.5 *,† | 2.4 ± 0.3 *,† |
| HSS42 | 3.1 ± 0.3 | 54.5 ± 10.4 *,† | 5.3 ± 0.5 *,† | 3.5 ± 0.7 *,† | 2.1 ± 0.6 *,† |
| HSS43 | 3.3 ± 0.7 | 61.1 ± 9.9 *,† | 5.0 ± 0.3 *,† | 3.4 ± 0.5 *,† | 2.3 ± 0.4 *,† |
| HSS44 | 3.2 ± 0.8 | 20.2 ± 9.5 * | 3.9 ± 0.4 * | 1.7 ± 0.2 * | 1.0 ± 0.5 * |
| HSS45 | 3.4 ± 0.6 | 28.2 ± 8.6 * | 4.0 ± 0.6 * | 1.8 ± 0.6 * | 0.8 ± 0.2 * |
| HSS46 | 3.1 ± 0.4 | 22.1 ± 7.5 * | 3.8 ± 0.3 * | 1.6 ± 0.7 * | 1.2 ± 0.6 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Ramírez, L.A.; Martínez-Álvarez, J.A.; Martínez-Duncker, I.; Lozoya-Pérez, N.E.; Mora-Montes, H.M. Silencing of Sporothrix schenckii GP70 Reveals Its Contribution to Fungal Adhesion, Virulence, and the Host–Fungus Interaction. J. Fungi 2024, 10, 302. https://doi.org/10.3390/jof10050302
López-Ramírez LA, Martínez-Álvarez JA, Martínez-Duncker I, Lozoya-Pérez NE, Mora-Montes HM. Silencing of Sporothrix schenckii GP70 Reveals Its Contribution to Fungal Adhesion, Virulence, and the Host–Fungus Interaction. Journal of Fungi. 2024; 10(5):302. https://doi.org/10.3390/jof10050302
Chicago/Turabian StyleLópez-Ramírez, Luz A., José A. Martínez-Álvarez, Iván Martínez-Duncker, Nancy E. Lozoya-Pérez, and Héctor M. Mora-Montes. 2024. "Silencing of Sporothrix schenckii GP70 Reveals Its Contribution to Fungal Adhesion, Virulence, and the Host–Fungus Interaction" Journal of Fungi 10, no. 5: 302. https://doi.org/10.3390/jof10050302