
Four New Species and One New Record of Thelephora from China
by
 and
and
Mei-Zhi Tian
1,2,3,
Hai-Bin Xia
2,
Zheng-Lin Gao
4,
Chang-Yin Zhao
4,
Dan Ma
4,
Zhu-Liang Yang
1,3,* and
Yan-Chun Li
1,3,* 1
Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
2
College of Life Sciences, Shanxi Normal University, Xi’an 710100, China
3
Yunnan Key Laboratory for Fungal Diversity and Green Development, Kunming 650201, China
4
Amu Mountain Provincial Nature Reserve Management and Protection Bureau, Honghe 654400, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2024, 10(4), 300; https://doi.org/10.3390/jof10040300
Submission received: 8 March 2024
/
Revised: 10 April 2024
/
Accepted: 18 April 2024
/
Published: 22 April 2024
(This article belongs to the Special Issue Women in Mycology, 2nd Edition)
Abstract
:Species of the genus Thelephora (Thelephorales, Thelephoraceae) are ectomycorrhizal symbionts of coniferous and broad-leaved plants, and some of them are well-known edible mushrooms, making it an exceptionally important group ecologically and economically. However, the diversity of the species from China has not been fully elucidated. In this study, we conducted a phylogenetic analysis based on the internal transcribed spacer (ITS) regions, using Maximum Likelihood and Bayesian analyses, along with morphological observations of this genus. Four new species from China are proposed, viz., T. dactyliophora, T. lacunosa, T. petaloides, and T. pinnatifida. In addition, T. sikkimensis originally described from India is reported for the first time from China. Thelephora dactyliophora, T. pinnatifida, and T. sikkimensis are distributed in subtropical forests and mainly associated with plants of the families Fagaceae and Pinaceae. Thelephora lacunosa and T. petaloides are distributed in tropical to subtropical forests. Thelephora lacunosa is mainly associated with plants of the families Fagaceae and Pinaceae, while T. petaloides is mainly associated with plants of the family Fagaceae. Line drawings of microstructures, color pictures of fresh basidiomes, and detailed descriptions of these five species are provided.
1. Introduction
Thelephora Ehrh. ex Willd. was originally described by Ehrhart and validly published by Willdenow with T. terrestris Ehrh. as its type species [1,2]. The genus is characterized by the diverse shapes of basidiomes which are stereoid, imbricate, rosette, infundibuliform, coralloid, or resupinate; the sulcate, zonate, glabrous to strigose, somewhat radially rugulose or wrinkled abhymenial surfaces; the smooth, slightly rugose to warty hymenial surfaces; the monomitic, clamped hyphal systems; the verruculose or echinulate basidiospore ornamentations; and the presence or absence of cystidia [1,2,3,4,5,6]. Species of the genus Thelephora are associated with a variety of plants of the families Pinaceae, Casuarinaceae, Ericaceae, Fagaceae, and Betulaceae. The mycelia of these species around the plant roots can help plants obtain essential minerals and water from the soil and resist diseases and drought [7], making significant contributions to plant health and ecosystem stability. In addition, some species in the genus play an essential role in the edible and medicinal fungal industry; e.g., Thelephora aurantiotincta Corner and T. ganbajun M. Zang have anticancer and anti-allergic effects [8,9,10]. Meanwhile, T. ganbajun is also one of the most popular edible mushrooms in China and some East Asian countries [11,12,13,14].
According to our preliminary statistics, 64 species of Thelephora have been accepted (http://www.indexfungorum.org/Names/Names.asp, accessed date: 25 February 2024), and they are mainly distributed in the northern temperate and tropical regions from Europe [15], North America [16,17,18,19,20], and Asia [21,22]. In China, 24 species have been reported [12,13,14,23,24,25]. During our recent research on Thelephora across China, we encountered five impressive species that have not been reported from this country. These species can be easily recognized by their conspicuous colors in the field. Molecular phylogenetic analyses of this genus based on the nuclear ribosomal internal transcribed spacer (ITS) indicated that they represent five distinct species. Combined with morphological characteristics, five species including four new to science: T. dactyliophora Yan C. Li & Zhu L. Yang, T. lacunosa Yan C. Li & Zhu L. Yang, T. petaloides Yan C. Li & Zhu L. Yang, and T. pinnatifida Yan C. Li & Zhu L. Yang, and one new to China, viz., T. sikkimensis K. Das, Hembrom & Kuha, were studied and documented herein.
2. Materials and Methods
2.1. Specimen Collections
Nineteen specimens of Thelephora were collected from China, including seventeen from southwestern China (Yunnan Province) and two from eastern China (Zhejiang Province). Photographs of these fresh specimens were taken in the field. Macroscopic characteristics and field notes including date, location, and habitat were made for each specimen, and then these specimens were dried using a fungal dryer and deposited in the fungal herbarium of the Herbarium KUN (Kunming Institute of Botany, Chinese Academy of Sciences) for taxonomic studies. Small pieces of each specimen were also dried in silica gel for molecular studies.
2.2. Morphological Studies
Macroscopic characteristics were obtained from specimen records and photos captured in the field. Color codes followed Kornerup and Wanscher [26,27]. Microscopic characteristics that include the structure of the pileipellis and the morphology of context hyphae, subhymenium hyphae, basidia, cystidia, and basidiospores were observed under a ZEISS Axiostar Plus microscope (Carl Zeiss AG, Oberkochen, Germany). Tissues were sectioned and mounted in 10% KOH, Cotton Blue (test for cyanophily), and Melzer’s reagent (test for amyloidity and dextrinoidity) [13,25,28,29]. Microscopic structures of pileipellis, hymenium, and subhymenium were drawn freehand; basidiospores and hyphae were measured at 1000× g magnification. The observations of basidiospore ornamentations were obtained with a ZEISS Sigma 300 scanning electron microscope (SEM) (Carl Zeiss AG, Oberkochen, Germany). For the measurement of basidiospores, at least 20 basidiospores were measured for each specimen; the notation “basidiospores (n/m/p)” is used to mean n basidiospores measured from m basidiomes of p specimens. L denotes spore length (arithmetic average of all spores); W denotes spore width (arithmetic average of all spores); Q denotes variation in the length/width ratios of basidiospores in side view; Qm denotes the average Q of all basidiospores ± sample standard deviation [30,31,32]. Since the shape of basidiospore is mainly inferred by its length/width ratio, we have defined the terms of basidiospore shapes as follows: globose (length/width ratio = 1.01–1.05), subglobose to broadly ellipsoid (length/width ratio = 1.05–1.3), ellipsoid (length/width ratio = 1.3–1.6), and elongated (length/width ratio = 1.6–2). The abbreviation masl means meters above sea level [33].
2.3. DNA Extraction, PCR and DNA Sequencings
The extraction of genomic DNA from dried specimens was conducted using the CTAB method [34] and Ezup Column Fungi Genomic DNA Purification Kit (Shanghai Jinban Biotechnology Co., Ltd., Shanghai, China). PCR reactions contained 1 μL DNA solution (adjusted to approximately 20 ng), 1 µL of each primer, and 15 μL 2× Taq PCR Master Mix including Taq DNA Polymerase, MgCl2, and dNTP mix (Beijing Biomed Gene Technology Co., Ltd., Beijing, China). The final volume was adjusted to 30 μL with distilled sterile H2O. The amplification conditions were set as follows: denaturation at 95 °C for 4 min, 35 cycles of 30 s at 94 °C, 40 s at 53 °C, 1 min at 72 °C, a final extension of 8 min at 72 °C, and then coolant at 14 °C [35,36]. In this study, the internal transcribed spacer (ITS) regions were amplified with the primers ITS1F/ITS4 [30,34,37,38]. Some full lengths of ITS sequences that failed to be amplified were divided into two parts for amplification with primer pairs ITS1F/5.8S and 5.8SR/ITS4, respectively [39], and then sequences of these two parts were spliced together to obtain the full length of the ITS sequence. Sequences newly generated in this study were deposited in GenBank (Table 1).
2.4. Phylogenetic Analyses
DNA sequences were compiled with SeqMan (DNASTAR Lasergene 9). Sequences related to Thelephora downloaded from GenBank (https://www.ncbi.nlm.nih.gov/, accessed on 6 April 2024) and UNITE (https://unite.ut.ee/, accessed on 25 February 2024), and newly generated in this study were aligned with Mafft V7.490 [42] and manually adjusted with PhyDE when necessary. Then we performed phylogenetic analyses by Maximum Likelihood (ML) and Bayesian Inference (BI) approaches. The ML analysis was inferred with RaxmlGUI 2.0.10 [43] with model set as GTRGAMMAI for 1000 bootstrap replicates combined with an ML search. The BI analysis was conducted using MrBayes 3.2 [44] running 2,000,000 bootstrap replicates; the best-fit likelihood model employed for ITS dataset was HKY + I + G + F based on the Akaike Information Criterion (AIC) with ModelFinder [45,46]; the posterior probabilities (PP) were determined twice by running one cold and three heated chains in parallel mode, saving trees every 1000th generation. Other parameters were kept at their default settings. Runs were terminated once the average standard deviation of split frequencies went below 0.01. Genetic distances among ITS sequences were calculated using the Maximum Composite Likelihood model [47] in MEGA 11 [48].
3. Results
3.1. Phylogenetic Analyses
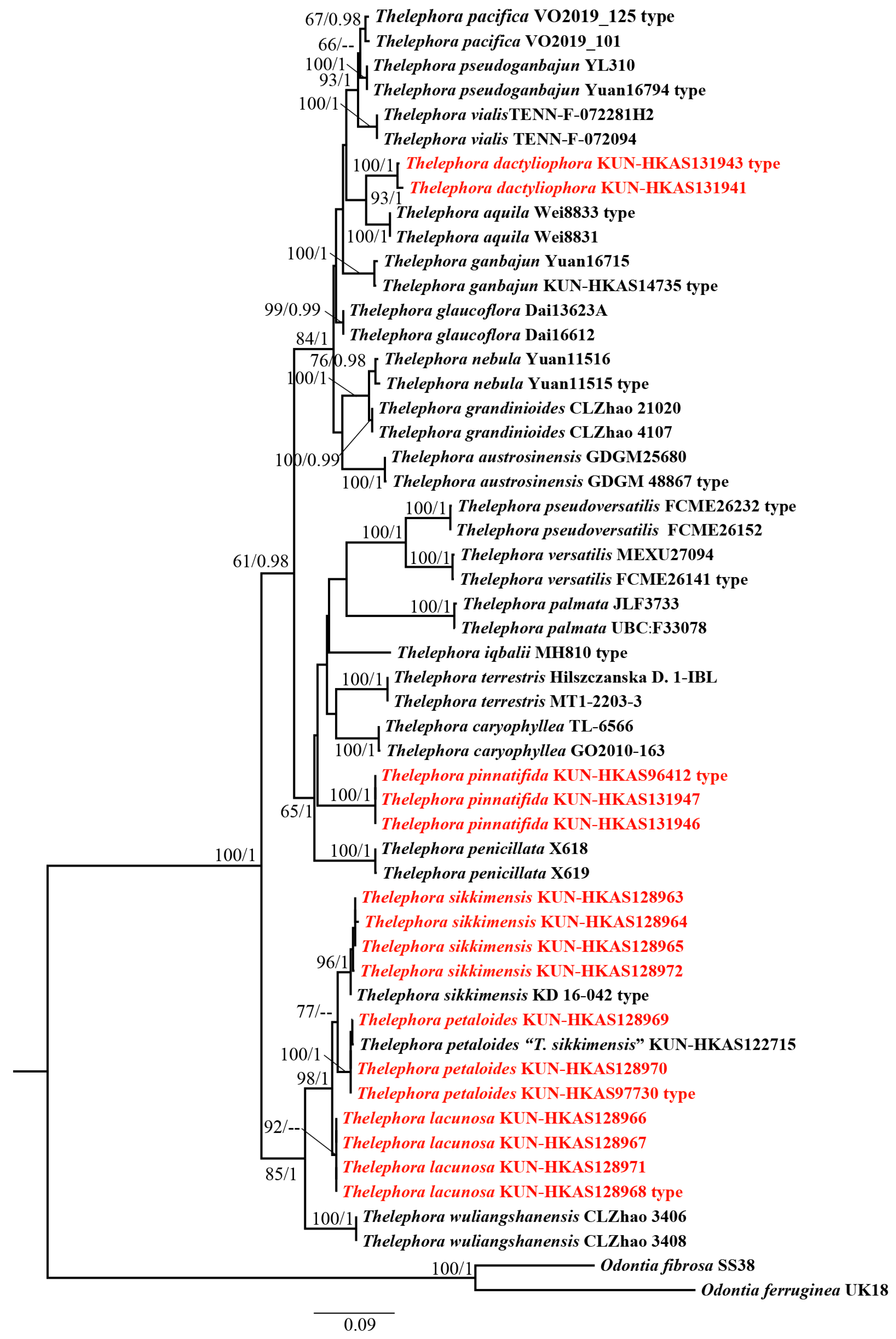
Phylogenetic analyses were conducted based on 53 ITS sequences of which 16 were newly generated in this study and 37 were downloaded from UNITE and GenBank (Table 1). Odontia fibrosa (Berk. & M.A. Curtis) Kõljalg and O. ferruginea Pers. were used as the outgroup taxa [14,18,22,47]. The alignment was submitted to TreeBASE (S30371). ML and BI approaches showed minimal differences in evaluation results; thus, only an ML tree was used for display (Figure 1). Our phylogenetic analyses indicated that T. sikkimensis, T. petaloides, and T. lacunosa cluster together with high support values (98/1). T. dactyliophora is sister to T. aquila S.R. Yang, Y.L. Wei & H.S. with moderate support values (93/1). Thelephora pinnatifida forms a separate branch with high support values (100/1).
3.2. Analyses of Genetic Distances
In this study, the Maximum Composite Likelihood model was used to calculate the genetic distances within and between species of Thelephora used in this study. As shown in Table S1, the genetic distances of the ITS sequences within species ranged from 0 to 0.9%, with an average distance of 0.2%; the genetic distances between species ranged from 1.2% to 22.3%, with an average of 12.8%.
3.3. Taxonomy
Thelephora dactyliophora Yan C. Li & Zhu L. Yang, sp. nov., Figure 2a–c, Figure 3a–c and Figure 4a,b.
MycoBank: 852625.
Etymology: dactyliophora refers to the finger-like branches of the basidiomes.
Diagnosis: Basidiomes coralloid; branch multiple ranks, finger-like to narrow flabelliform when young, spathulate to narrow petaloid when mature; abhymenial surface slightly rugulose, non-zonate, brownish gray to gray; hymenial surface concolorous with abhymenial surface, rugulose and non-zonate. Basidiospores nodulose to verrucose. This species is similar to T. palmata (Scop.) Fr. in its coralloid basidiomes, but differs in its slightly rugulose brownish gray to gray hymenial surface and relatively small basidiospores.
Type: China, Zhejiang Province: Lishui City, Jingning County, altitude 759 masl, 22 June 2021, Liu-Kun Jia 1577 (KUN-HKAS131943, GenBank Acc. No. OR940523).
Description: Basidiomes 35–65 mm high, 30–40 mm wide, gregarious to caespitose, humid and leathery when fresh, corky to hard and brittle when drying; branches arising from a shared stipe or center and arranged in coralloid shape; branch multiple ranks, flat, finger-like to narrow flabelliform when young, spathulate to narrow petaloid when mature; margin thin, about 0.1 mm thick, deeply lacerate. Abhymenial surface slightly rugulose, non-zonate, brownish gray to gray (6F2–6F1), but chalky white to orange-white (1A1–5A2) at margin. Hymenial surface concolorous with abhymenial surface, rugulose and non-zonate. Context 0.1–0.5 mm thick, relatively thin at margin and thick towards base, white (1A1), orange-white to gray-white (5A2–2B1), odor mild when fresh, sweet odor when drying. Stipe 10–20 × 2–5 mm, irregularly cylindrical to flatted or broadened at base, surface smooth to slightly rugose, brownish gray (6F2).
Basidia 45–72 × 6–9 µm, slightly clavate, hyaline, thin-walled, clamped at base, with four sterigmata; sterigmata 3.5–5.5 µm long and 1.5–2 µm wide at base, yellowish in Melzer’s reagent. Basidiospores (60/3/2) (4.5–) 5–7.5 × 4–6.5 µm (not including ornamentations), L = 6 µm, W = 5 µm, Q = 1–1.6, Qm = 1.2 ± 0.15, subglobose to subellipsoid in side view, broadly ellipsoid to ellipsoid to irregularly lobed in ventral view, slightly thick-walled; surface nodulose to verrucose, ornamentation usually isolated, sometimes in groups from two to three, echinulis 0.3–0.5 µm high, subconical but apex somewhat obtuse; yellowish brown in 10% KOH, yellow to yellow-brown in Melzer’s reagent, non-amyloid, non-dextrinoid, cyanophilic.
Monomitic hyphal system. Pileipellis: generative hyphae 3–5.5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, moderately branched, frequently branched near clamp connections, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Context: generative hyphae 3.5–4.5 (–5.5) µm wide, hyaline, subparallel to loosely interwoven, thin-walled, moderately branched, frequently branched near clamp connections, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Subhymenium: generative hyphae 3–5 µm wide, hyaline, subparallel to slightly divergent, thin-walled, frequently branched near clamp connections, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic.
Habitat: In subtropical evergreen broad-leaved forests dominated by plants of Fagaceae and Betulaceae, or in mixed forests dominated by plants of Fagaceae and Pinaceae.
Distribution: Currently known from Zhejiang Province (eastern China).
Additional specimens examined: China, Zhejiang Province: Lishui, Jingning County, Wangdongyang Forest Wetland National Park, 1321 masl, 11 September 2021, Geng-Shen Wang 1795 (KUN-HKAS131941); the same location, 15 July 2022, Peng-Cheng Yuan 698 (KUN-HKAS131940).
Notes: Thelephora dactyliophora is phylogenetically related to T. aquila. These two species share similar sizes of basidiomes (50 × 40 mm in T. aquila vs. 35–65 × 30–40 mm in T. dactyliophora) and similar sizes of basidiospores (5−7.3 × 4−6.5 µm in T. aquila vs. 4.5–7.5 × 4–6.5 µm in T. dactyliophora). However, T. aquila is easily distinguished from T. dactyliophora by its flabelliform to applanate–lobate branches of basidiomes and its black and zonate abhymenial surface which is somewhat radially rugulose or wrinkled [25]. The coralloid basidiomes of T. dactyliophora are very similar to those of T. palmata. However, T. palmata is clearly different from T. dactyliophora in its velvet brownish purple to black or purple-brown branches and relatively large basidiospores (7.4–9.3 × 8–11.1 µm in T. palmata vs. 4.5–7.5 × 4–6.5 µm in T. dactyliophora) [20,49].
MycoBank: 852626.
Etymology: lacunosa refers to the lacunary abhymenial surface.
Diagnosis: Basidiomes imbricate, rosette to flabelliform; branches flabelliform, petaloid to applanate–lobate; abhymenial surface lacunary, sulcate, irregularly ridged, non-zonate, and warty, brown to grayish brown or grayish orange at center then gradually becoming paler towards margin; hymenial surface imperceptibly rugose and warty, brownish orange to gray or purple-gray at center and becoming pale orange towards margin. Basidiospores nodulose to verrucose. This species is morphologically similar to T. ganbajun in the imbricate to rosette basidiomes, but it differs in its lacunary, irregularly ridged, non-zonate, brown to grayish brown or grayish orange abhymenial surface and brownish red stipe.
Type: China, Yunnan Province: Honghe, Jiache Town, 2140 masl, 22 September 2022, Yan-Chun Li 3913 (KUN-HKAS128968, GenBank Acc. No. OR512335).
Description: Basidiomes 67–80 mm high, 80–150 mm wide, solitary, gregarious to caespitose, humid and leathery when fresh, corky to hard and brittle when drying; branches arising from a shared stipe or center and arranged in imbricate, rosette to flabelliform; branches flat, flabelliform, petaloid to applanate–lobate; margin 0.5–3 mm thick, nearly entire, rarely lacerate, imperceptibly wavy. Abhymenial surface lacunary, sulcate, irregularly ridged, non-zonate, and warty, brown to grayish brown (6E5–7E3) or grayish orange (6B3) at center then gradually becoming paler towards margin, but white to pale orange-gray (5A1–5B2) at margin. Hymenial surface imperceptibly rugose and warty, brownish orange to gray (6B4–7D2) or purple-gray (14E2) at center and becoming pale orange (6A3) towards margin, but orange-white (5A2) at margin. Context 0.5–3.5 mm thick, yellowish white to gray-white (2A2–2B1), odor mild when fresh, yeast powder flavor when drying. Stipe 20–25 × 12–35 mm, subconical to broadened or flatted towards base, surface rugose, glabrous, brownish red to brownish yellow (7E8–5D8).
Basidia 72–100 × 8–12 µm, subclavate, hyaline, thin-walled, clamped at base, with four sterigmata; sterigmata 3–6 µm long and 1.5–3 µm wide at base, yellowish in Melzer’s reagent. Basidiospores (100/5/4) 4.5–6.5 × 4–5.5 µm (not including ornamentations), L = 5.5 µm, W = 5 µm, Q = 1–1.5, Qm = 1.1 ± 0.11, subglobose to subellipsoid in side view, broadly ellipsoid to ellipsoid to irregularly lobed in ventral view, slightly thick-walled, pale yellowish brown in 10% KOH, yellow-brown to brown in Melzer’s reagent, non-amyloid, non-dextrinoid, cyanophilic; surface nodulose to verrucose, ornamentation usually isolated, sometimes in groups from two to three, echinulis 0.4–0.5 µm high, subconical but somewhat obtuse at apex.
Monomitic hyphal system. Pileipellis: generative hyphae 5–6 µm wide, hyaline to yellowish, subparallel to loosely interwoven, thin-walled, frequently branched near clamp connections, commonly with simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Context: generative hyphae 3–5.5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, frequently branched near the clamp connections, with a few simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Subhymenium: generative hyphae 2.5–4.5 µm wide, hyaline, subparallel to slightly divergent, thin-walled, moderately branched, with clamp connections and a few simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic.
Habitat: In subtropical evergreen broad-leaved forests dominated by plants of Fagaceae, or in subtropical mixed forests dominated by plants of Fagaceae and Pinaceae.
Distribution: Currently known from Yunnan Province (southwestern China).
Additional specimens examined: China, Yunnan Province: Honghe, Jiache Town, Sanjian Mountains, 2140 masl, 22 September 2022, Yan-Chun Li 3971 (KUN-HKAS128971); Puer, Taiyanghe Nature Reserve, 1400 masl, 2 July 2022, Yan-Chun Li 3307 (KUN-HKAS128966); the same date and location, Yan-Chun Li 3311 (KUN-HKAS128967).
Notes: In morphology, T. lacunosa shares similar features with T. sikkimensis: the flabelliform to petaloid branches of basidiomes with thin and imperceptibly wavy margin; the warty and non-zonate hymenial surface; and the imbricate, rosette basidiomes [22]. However, T. lacunosa differs from T. sikkimensis clearly in the lacunary, non-zonate, and grayish brown to grayish orange abhymenial surface and the subconical stipe. The yeast powder flavor when drying, the imbricate to rosette basidiomes, and the flabelliform branches of basidiomes make T. ganbajun similar to T. lacunosa, but T. ganbajun has an inconspicuously gray to black abhymenial surface and a brown to yellowish brown stipe which is cylindrical to flatted or broadened [25]. Thelephora lacunosa and T. pseudoganbajun S.R. Yang & H.S. Yuan share some common features including the imperceptibly wavy margin of the branches and the imbricate to rosette basidiomes [25]. However, T. pseudoganbajun differs from T. lacunosa in its brown to yellowish brown and somewhat radially rugulose or wrinkled abhymenial surface which is obscurely zonate, radially rugulose or longitudinally wrinkled hymenial surface, relatively small basidia (45–65 × 6–10 µm in T. pseudoganbajun vs. 72–100 × 8–12 µm in T. lacunosa), and occasionally isotypical clamp connections symmetrically growing on both sides of the hyphae [25]. Thelephora pacifica is similar to T. lacunosa due to its grayish brown abhymenial surface and similar size of basidiospores (4.3–7.3 × 3.6–6 µm in T. pacifica vs. 4.5–6.5 × 4–5.5 µm in T. lacunose), but T. pacifica differs from T. lacunosa in its pale olive-yellow margin of basidiomes, dark grayish brown stipe, and relatively small basidia (19.5–30 × 4.7–6.3 µm in T. pacifica vs. 72–100 × 8–12 µm in T. lacunosa) [41].
MycoBank: 852627.
Etymology: petaloides refers to the petaloid basidiomes.
Diagnosis: Basidiomes imbricate to umbrella; branches flabelliform to applanate–lobate; abhymenial surface sulcate, somewhat irregularly ridged, glabrous, zonate, orange-yellow to grayish orange or pale orange elsewhere; hymenial surface imperceptibly rugose, warty, zonate near margin, bright orange elsewhere. Basidiospores nodulose to verrucose. This species is similar to T. sikkimensis in its sulcate abhymenial surface and warty hymenial surface but differs in its orange-gray and densely warty hymenial surface and visibly wavy margin of basidiomes.
Type: China, Yunnan Province: Puer City, Lancang County, Huimin Town, 1410 masl, 26 September 2016, Jian-Wei Liu 530828MF-226 (KUN-HKAS97730, GenBank Acc. No. OR512334).
Description: Basidiomes (40–) 80–100 mm high, 55–140 (–220) mm wide, solitary to concrescent, humid and leathery when fresh, corky to hard and brittle when drying; branches arising from a shared stipe or center and arranged in imbricate to umbrella shape; branches flat, flabelliform, applanate–lobate; margin 1–2 mm thick, nearly entire, rarely lacerate, visibly wavy. Abhymenial surface sulcate, somewhat irregularly ridged, glabrous, zonate, brownish gray (6E2) at center, orange-yellow to grayish orange (6A8–6B3) or pale orange (5A2) elsewhere, but chalky white (1A1) at margin. Hymenial surface imperceptibly rugose, warty, zonate near margin, orange-gray (5B2) at center, becoming bright orange (5A4) elsewhere, but chalky white (1A1) at margin. Context 1–4 mm thick, relatively thin at margin and thick towards center, white to yellowish white (1A1–2A2), odor mild when fresh, yeast powder flavor when drying. Stipe 10–40 × 9–13 mm, irregularly cylindrical to flatted or broadened at base, surface rugose, glabrous, brownish gray (5E2).
Basidia 53–92 × 9–12 µm, subclavate, pale yellow to pale yellowish brown, thin-walled, clamped at base, with four sterigmata; sterigmata 3–5 µm long and 2.5–3 µm wide at base, yellowish in Melzer’s reagent. Basidiospores (100/5/5) 4.5–7.5 × 3.5–6.5 µm (not including ornamentations), L = 6 µm, W = 5.2 µm, Q = 1–1.71, Qm = 1.16 ± 0.14, subglobose to subellipsoid or sometimes elongated in side view, globose to ellipsoid to irregularly lobed in ventral view; surface nodulose to verrucose, slightly thick-walled; ornamentation in groups from one to two, echinulis 0.5–0.7 µm high, subconical but apex somewhat obtuse; pale yellowish brown in 10% KOH, yellow-brown to brownish in Melzer’s reagent, non-amyloid, non-dextrinoid, cyanophilic.
Monomitic hyphal system. Pileipellis: generative hyphae 4–5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, moderately branched, with clamp connections and a few simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Context: generative hyphae 4–5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, moderately branched, with clamp connections and a few simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Subhymenium: generative hyphae 2–4 µm wide, pale yellow to pale yellowish brown, subparallel to slightly divergent, thin-walled, frequently branched near clamp connections, with a few simple septa, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic.
Habitat: In subtropical forests dominated by plants of Fagaceae.
Distribution: Currently known from Yunnan Province (southwestern China).
Additional specimens examined: China, Yunnan Province: Honghe, Jiache Town, Sanjian Mountains, 2140 masl, 22 September 2022, Yan-Chun Li 3918 (KUN-HKAS128969); the same date and location, Yan-Chun Li 3960 (KUN-HKAS128970); Baoshan, Longling County, 1700 masl, 12 September 2002, Zhu L. Yang 3581 (KUN-HKAS42087); Nujiang, Lushui County, Pianma Town, 1850 masl, 1 April 2001, Fu-Qiang Yu 696 (KUN-HKAS38935).
Notes: In our phylogenetic analyses, T. petaloides, T. sikkimensis, T. lacunosa, and T. wuliangshanensis clustered together with high support values (85/1) (Figure 1). However, T. wuliangshanensis is characterized by its infundibuliform basidiomes, the buff to salmon abhymenial surface with radially black striations, and the umber to coffee and nearly glabrous hymenial surface [14]. Morphologically, T. petaloides and T. lacunosa share similar characteristics in the flat, flabelliform branches of basidiomes and warty hymenial surface. However, T. lacunosa can be distinguished by its imperceptibly wavy margin of basidiomes; lacunary, sulcate, irregularly ridged, non-zonate, and warty abhymenial surface which is brown, grayish brown to grayish orange but white to pale orange-gray at the margin; and brownish orange hymenial surface with orange-white tint at the margin. In addition, the sulcate abhymenial surface, the flat and flabelliform branches of basidiomes, the white context, and the warty hymenial surface make T. petaloides similar to T. sikkimensis. However, T. sikkimensis is obviously distinguished from T. petaloides in its relatively small basidiomes (45–67 × 50–70 mm in T. sikkimensis vs. 40–100 × 55–220 mm in T. petaloides); grayish black to purplish gray or grayish green abhymenial surface which is chalky white to orange-white at margin; smooth to irregularly plicate hymenium surface which is pale orange to purplish gray or grayish green, but orange-white to almost white at the margin; and rugose and warty stipe which is pale orange or sometimes purplish gray.
MycoBank: 852628.
Etymology: pinnatifida refers to the pinnatifid margin of branches of the basidiomes.
Diagnosis: Basidiomes clavarioid; branch clavate to pinnatifid or ramiform with tips needle-like; surface sulcate, non-zonate, visibly ribbed, brownish orange to brown. Basidiospores nodulose to verrucose. This species is similar to T. iqbalii Khalid & Hanif in its clavarioid basidiomes and needle-like tips of branches but differs in its brownish orange to brown basidiomes with chalky white to orange-white margin and relatively long basidia.
Type: China, Yunnan Province: Kunming, Chongming County, Dian Yuan Town, Liangwang Mountain, Jiaochangba, 2622 masl, 23 July 2016, Xiang-Hua Wang 3823 (KUN-HKAS96412, GenBank Acc. No. OR940526).
Description: Basidiomes 35–45 mm high, 28–85 mm wide, gregarious to caespitose, humid and leathery when fresh, corky when drying; branches arising from a shared stipe or center and arranged in clavarioid shape, sometimes sub-resupinate to resupinate; branch clavate to pinnatifid or ramiform with tips needle-like, surface sulcate, non-zonate, visibly ribbed, brownish orange to brown (5C5–7E8), but chalky white to orange-white (1A1–5A2) at margin; margin 0.1–0.5 mm thick, deeply lacerate, velvety. Context 0.1–2 mm thick, relatively thin at margin and thick towards center, yellowish white to brown (2A2–7E8), odor mild when fresh, yeast powder flavor when drying. Stipe (0–) 10–15 × (0–) 3–10 mm, irregularly cylindrical, surface rugose, brown (7E7).
Hymenium predominantly amphigenous. Basidia 47–90 × 10–11.5 µm, nearly clavate, pale yellow, thin-walled, clamped at base, with four sterigmata; sterigmata 5–8 (–10) µm long and 1.5–3 (–4) µm wide at base, yellowish in Melzer’s reagent. Basidiospores (80/4/4) (5.5–) 6–9.5 × 5–8.5 µm (not including ornamentations), L = 7.6 µm, W = 6.4 µm, Q = 1–1.7, Qm = 1.19 ± 0.15, subglobose to subellipsoid or sometimes elongated in side view, globose to broadly ellipsoid or ellipsoid to irregularly lobed in ventral view, slightly thick-walled; surface verrucose to subechinate, ornamentation usually isolated, sometimes in groups of two or more, echinulis 0.3–0.5 (–0.6) µm high, subconical but apex somewhat obtuse; pale yellowish brown in 10% KOH, yellow-brown to brown in Melzer’s reagent, non-amyloid, non-dextrinoid, cyanophilic.
Monomitic hyphal system. Hair-like appendages of pointed tips: generative hyphae 3.5–5 (–5.5) µm wide, subparallel to loosely interwoven, thin-walled, often branched near clamp connections, hyaline in 10% KOH, yellow to pale yellow in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Context: generative hyphae 4.5–6.5 µm wide, pale yellow, subparallel to loosely interwoven, thin-walled, moderately branched, frequently branched often near clamp connections, septate absent, yellowish in Melzer’s reagent, non-amyloid, non-dextrinoid, acyanophilic. Subhymenium: generative hyphae 4–5.5 µm wide, pale yellow, subparallel to slightly divergent, thin-walled, moderately branched, frequently branched often near clamp connections, septate absent, yellowish in Melzer’s reagent, with acyanophilic reaction in Cotton Blue.
Habitat: In subtropical broad-leaved forests dominated by plants of Fagaceae, or in coniferous forests dominated by plants of Pinaceae.
Distribution: Currently known from Yunnan Province (southwestern China).
Additional specimens examined: China, Yunnan Province: Kunming, Chongming County, Dianyuan Town, Liangwang Mountain, Jiaochangba, 2622 masl, 23 July 2016, Xiang-Hua Wang 3823 (KUN-HKAS96412); Kunming, Kunming Botanical Garden, 1977 masl, 15 August 2023, Mei-Zhi Tian 394 (KUN-HKAS131946) and Mei-Zhi Tian 395 (KUN-HKAS131947); Dali, Jianchuan County, Diannan Town, Fada Village, 2200 masl, 10 August 2023, Mei-Zhi Tian 392 (KUN-HKAS131944) and Mei-Zhi Tian 393 (KUN-HKAS131945).
Notes: Thelephora pinnatifida can be confused with T. iqbalii, T. penicillata (Pers.) Fr., T. pseudoversatilis Ram.-Lóp. & Villegas, and T. versatilis Ramírez-López & M. Villegas due to their clavarioid basidiomes [4,18,41]. However, they cluster into five independent lineages and represent five distinct species in our phylogenetic analyses. Furthermore, T. iqbalii differs from T. pinnatifida in its grayish brown basidiomes with pinkish white margin of branches and relatively short basidia (40–55 × 6–10 µm in T. iqbalii vs. 47–90 × 10–11.5 µm in T. pinnatifida) [41]. Thelephora penicillata differs from T. pinnatifida in its fuscous purple basidiomes and much higher echinulis on the surface of basidiospores (echinulis up to 1.5 µm high in T. penicillata vs. 0.3–0.6 µm high in T. pinnatifida) [4]. Thelephora pseudoversatilis differs from T. pinnatifida in its much higher echinulis on the surface of basidiospores (echinulis 1–2 µm high in T. pseudoversatilis vs. 0.3–0.6 µm high in T. pinnatifida). Thelephora versatilis differs from T. pinnatifida in its clavate branches without pinnatifid margin and much higher echinulis on the surface of basidiospores (echinulis 0.5–1.5 high in T. versatilis vs. 0.3–0.6 µm high in T. pinnatifida).
Description: Basidiomes 45–67 mm high, 50–70 mm wide, gregarious to caespitose, humid and leathery when fresh, corky to hard and brittle when drying, branches arising from a shared stipe or center and arranged in imbricate to rosette shape; branches effused to reflexed, flabelliform to petaloid, margin thin about 1 mm thick, nearly entire, rarely lacerate, imperceptibly wavy. Abhymenial surface sulcate, indistinctly zonate, radially rugulose or wrinkled, grayish black (18F1) at center, purplish gray to grayish green (13E2–25E5) elsewhere, but chalky white to orange-white (1A1–5A2) at margin. Hymenial surface rugose, warty, gradually from smooth to irregularly plicate near the margin, non-zonate, pale orange (5A4) when young, purplish gray (13E2) to grayish green (24C1) when mature, but orange-white to almost white (5A2–1A1) at margin. Context 1–2 mm thick, relatively thin at margin and thick towards center, white, odor mild when fresh, yeast powder flavor when drying. Stipe 12–14 × 2–9 mm, irregularly cylindrical, surface rugose and warty, pale orange (5A3), sometimes purplish gray (13E2).
Basidia 72–100 × 7.5–10 µm, subclavate to subcylindrical, hyaline, thin-walled, clamped at base, with four sterigmata; sterigmata 4–6 µm long and 2–3 µm wide at base, yellowish in Melzer’s reagent. Basidiospores (80/4/4) 4.5–6 (–6.5) × 4–5.5 µm (not including ornamentations), L = 5.6 µm, W = 5 µm, Q = 1–1.38, Qm = 1.13 ± 0.09, subglobose to subellipsoid in side view, globose to broadly ellipsoid or sometimes ellipsoid to irregularly lobed in ventral view, slightly thick-walled; surface nodulose to verrucose, ornamentation in groups from one to two, echinulis 0.3–0.7 µm high, subconical but apex somewhat obtuse; pale brown in 10% KOH, yellowish brown to brownish in Melzer’s reagent, non-amyloid, non-dextrinoid, cyanophilic.
Monomitic hyphal system. Pileipellis: generative hyphae 3–5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, moderately branched, with clamp connections and a few simple septa, yellowish in Melzer’s reagent, with acyanophilic reaction in Cotton Blue. Context: generative hyphae 2.5–5.5 µm wide, hyaline, subparallel to loosely interwoven, thin-walled, frequently branched near clamp connections, with a few simple septa, yellowish in Melzer’s reagent, with acyanophilic reaction in Cotton Blue. Subhymenium: generative hyphae 2–4 µm wide, hyaline, subparallel to slightly divergent, thin-walled, frequently branched near clamp connections, with a few simple septa, yellowish in Melzer’s reagent, with acyanophilic reaction in Cotton Blue.
Habitat: In subtropical mixed forests dominated by plants of Fagaceae and Pinaceae.
Distribution: Currently known from southwestern China and northwestern India.
Additional specimens examined: China, Yunnan Province: Baoshan, Mangkuan Town, Baihualing Village, 2100 masl, 7 August 2022, Mei-Zhi Tian 157 (KUN-HKAS128964); the same date and location, Mei-Zhi Tian 164 (KUN-HKAS128972) and Mei-Zhi Tian 173 (KUN-HKAS128965); Tengchong County, Tiantai Mountains, 2000 masl, 25 July 2022, Mei-Zhi Tian 051 (KUN-HKAS128963); Tengchong County, Zhonghe Town, Gaotian Village, 2040 masl, 13 August 2010, Yan-Jia Hao 252 (KUN-HKAS69236).
Notes: Thelephora sikkimensis was originally described from India and characterized by its imbricate to rosette-shaped basidiomes with branches arising from a shared stipe or center, pale orange to grayish green and non-zonate, warty hymenial surface, and rugose to warty and pale orange stipe [22].
Thelephora aurantiotincta has a warty and pale orange hymenial surface, subclavate to subcylindrical basidia, and irregularly lobed basidiospores. Such traits are very similar to T. sikkimensis, but T. aurantiotincta can be easily distinguished from T. sikkimensis by its non-zonate pale orange abhymenial surface [4,50]. The imbricate to rosette basidiomes, the zonate abhymenial surface, and the presence of cystidia of T. ganbajun make it similar to T. sikkimensis. However, T. ganbajun is different from T. sikkimensis by its gray to dark-brown abhymenial surface, gray to black hymenial surface, brown to yellowish brown stipe without warty ornamentation, and relatively short basidia (25–55 µm long in T. ganbajun vs. 72–100 µm long in T. sikkimensis). Thelephora sikkimensis is reported here for the first time from China.
Phylogenetically, T. sikkimensis, T. petaloides, and T. lacunosa clustered together with high support values. However, T. petaloides can be distinguished from T. sikkimensis by its big basidiomes (40–100 × 55–220 mm in T. petaloides vs. 45–67 × 50–70 mm in T. sikkimensis); nearly glabrous abhymenial surface which is brownish gray at the center becoming orange-yellow to grayish orange or pale orange elsewhere but chalky white at the margin; rugose and warty hymenium which is orange-gray at the center, bright orange elsewhere, and chalky white at the margin; rugose and glabrous brownish gray stipe; and a relatively thick context which is up to 4 mm thick. Thelephora lacunosa has medium-sized basidiomes (67–80 × 80–150 mm); lacunary, irregularly ridged, non-zonate, and warty abhymenial surface which is brown, grayish brown to grayish orange at the center, much paler towards the margin, and white to pale orange-gray at the margin; rugose and warty hymenium surface, brownish orange to gray or purple-gray at the center, pale orange towards the margin, and orange-white at the margin; and subconical stipe which is broadened or flatted at the base with a brownish red to brownish yellow surface.
3.4. Discussion
In this study, five species, T. sikkimensis new to China and T. petaloides, T. lacunosa, T. dactyliophora, and T. pinnatifida new to science, are reported based on a combination of phylogenetic analyses, genetic differences, and morphological features. In our phylogenetic analyses, T. lacunosa, T. petaloides, and T. sikkimensis clustered together with high support values, but the relationships among the three taxa are not well solved by the ML and Bayesian analyses. In addition, the diagnostic morphological characters among them are often subtle. It seems that these species might have started diverging from each other relatively recently. Genetic distance measurements have been used frequently as a tool to investigate species boundaries and to identify cryptic diversity in fungi [51,52,53]. Based on the Maximum Composite Likelihood model, the genetic variations between the three closely related species T. lacunosa/T. sikkimensis, T. petaloides/T. sikkimensis, and T. lacunosa/T. petaloides are 1.9–3%, 2.7–3.4%, and 2.1–3%, respectively, which are much higher than the within-species genetic variation (ranging from 0 to 0.9%). Morphologically, all of these five species have nodulose to verrucose ornamentations on the surface of basidiospores. But T. sikkimensis, T. petaloides, and T. lacunosa have imbricate basidiomes, while T. dactyliophora and T. pinnatifida have clavarioid basidiomes. It is evident that the shapes of basidiomes and the morphology of the hymenial surface are important characteristics distinguishing species of Thelephora. However, for the precise identification of Thelephora species, the size of basidiomes, basidia, and basidiospores; the shapes and size of the stipe; the color and zonation of abhymenial and hymenial surfaces; and the thickness of the context are also important characteristics. Detailed morphology comparisons among these five species are shown in Table 2. The combination of phylogenetic analyses, genetic differences, and morphological features can enhance the reliability and objectivity of species delimitation [54].
To date, 285 species of this genus have been reported and described (Index Fungorum: http://www.indexfungorum.org/Names/Names.asp, accessed on 6 April 2024), of which 29 (including the 5 species reported in this study) have been reported from China. China is one of the most biodiverse countries in the world. Like other groups of fungi, there are likely to be many Thelephora species that have not been scientifically described or remain to be discovered [55]. Further studies are needed to enrich the species diversity of this genus.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/jof10040300/s1: Table S1: Estimates of evolutionary divergence among sequences of Thelephora.
Author Contributions
Conceptualization, Y.-C.L., Z.-L.Y., H.-B.X. and M.-Z.T.; Field Sampling, Y.-C.L., M.-Z.T., Z.-L.G., C.-Y.Z. and D.M.; Molecular Experiments and Data Analysis, M.-Z.T.; Morphological Observation, Y.-C.L. and M.-Z.T.; Writing—Original Draft Preparation, M.-Z.T.; Writing—Review and Editing, Y.-C.L., Z.-L.Y. and H.-B.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ten Thousand Talents Program of Yunnan (YNWR-QNBJ-2018-125), the National Natural Science Foundation of China (U2202205), and the Yunnan Xingdian Talents Support Plan—Science and Technology Leading Talents Program (202305AB350004).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study (https://www.ncbi.nlm.nih.gov/, accessed on 6 April 2024; https://unite.ut.ee, accessed on 25 February 2024).
Acknowledgments
The authors appreciate the anonymous reviewers for their valuable comments and suggestions. We express our gratitude to Jian-Wei Liu (Kunming Institute of Botany, Chinese Academy of Sciences) for providing related specimens. We appreciate Zhi-jia Gu (Kunming Institute of Botany, Chinese Academy of Sciences) for assisting the observation of basidiospores under a scanning electron microscope (SEM).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Willdenow, C.L. Florae Berolinensis Prodromus: Secundum Systema Linneanum Ab Illustr. Viro Ac Eq. C. P. Thunbergio Emendatum Conscriptus (Classic Reprint); Impensis Wilhelmi Viewegii Berolini: Berlin, Germany, 1787; pp. 1–476. [Google Scholar]
- Ehrhart, J.F. 1785–1795: Plantae Cryptogamae Linn., Quas in Locis Earum Natalibus Collegit et Exsiccavit Fridericus Ehrhart; Hannover: Lower Saxony, Germany, 2020; pp. 1–320. [Google Scholar]
- Cunningham, G.H. The Thelephoraceae of Australia and New Zealand. N. Z. Dep. Sci. Ind. Res. Bull. 1963, 145, 228–239. [Google Scholar]
- Corner, E.J.H. A monograph of Thelephora (Basidiomycetes). Nova Hedwig. Beih. 1968, 27, 1–110. [Google Scholar]
- Corner, E.J.H. Futher notes on cantharelloid fungi and Thelephora. Nova Hedwig. 1976, 27, 325–342. [Google Scholar]
- Stalpers, J.A. The Aphyllophoraceous fungi I. Keys to the species of the Thelephorales. Stud. Mycol. 1993, 35, 1–168. [Google Scholar]
- Hilszczanska, D.; Sierota, Z. Persistence of ectomycorrhizas by Thelephora terrestris on outplanted Scots pine seedlings. Acta Mycol. 2006, 41, 313–318. [Google Scholar] [CrossRef]
- Xu, D.P.; Zheng, J.; Zhou, Y.; Li, Y.; Li, S.; Li, H.B. Extraction of natural antioxidants from the Thelephora ganbajun mushroom by an ultrasound-assisted extraction technique and evaluation of antiproliferative activity of the extract against human cancer cells. Int. J. Mol. Sci. 2016, 17, 1664. [Google Scholar] [CrossRef]
- Hu, L.; Liu, J.K. P-terphenyls from the Basidiomycete Thelephora aurantiotincta. Z. Für Naturforschung C 2003, 58, 452–454. [Google Scholar]
- Hu, L.; Liu, J.K. Two novel phenylacetoxylated p-terphenyls from Thelephora ganbajun Zang. Z. Für Naturforschung C 2001, 56, 983–987. [Google Scholar]
- Zang, M. Some new and noteworthy higher fungi from eastern himalayas. Acta Bot. Yunn. 1987, 9, 81–88. [Google Scholar]
- Qin, W.M. Thelephora caryophyllea—A corticioid fungus new to China. Guizhou Sci. 2009, 27, 19–21. [Google Scholar]
- Li, T.; Li, T.H.; Song, B.; Hosen, I. Thelephora austrosinensis (Thelephoraceae), a new species close to T. ganbajun from southern China. Phytotaxa 2020, 471, 208–220. [Google Scholar] [CrossRef]
- Liu, X.F.; Tibpromma, S.; Xu, J.C.; Kumla, J.; Karunarathna, S.C.; Zhao, C.L. Taxonomy and phylogeny reveal two new potential edible ectomycorrhizal mushrooms of Thelephora from east Asia. Diversity 2021, 13, 646. [Google Scholar] [CrossRef]
- Chacón, S.; Tapia, F.; Jarvio, D. Four interesting aphyllophoroid species in the tropical northern region of Veracruz, Mexico. Mycotaxon 2018, 133, 153–163. [Google Scholar] [CrossRef]
- Ramírez-López, I.; Villegas-Ríos, M.; Cano-Santana, Z. Phenotypic plasticity of the basidiomata of Thelephora sp. (Thelephoraceae) in tropical forest habitats. Rev. Biol. Trop. 2013, 61, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-López, I.; Villegas-Rios, M.; Salas-Lizana, R.; Garibay-Orijel, R.; Alvarez-Manjarrez, J. Thelephora versatilis and Thelephora pseudoversatilis: Two new cryptic species with polymorphic basidiomes inhabiting tropical deciduous and sub-perennial forests of the Mexican Pacific coast. Mycologia 2015, 107, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A.; Angelini, C.; Losi, C.; Ercole, E. Thelephora dominicana (Basidiomycota, Thelephorales), a new species from the Dominican Republic, and preliminary notes on Thelephoroid genera. Phytotaxa 2016, 265, 27–38. [Google Scholar] [CrossRef]
- Lentz, P.L. The genus Thelephora in Iowa. Proc. Iowa Acad. Sci. 1942, 49, 175–184. [Google Scholar]
- Coker, W.C. Notes on the Thelephoraceae of North Carolina. J. Elisha Mitchell Sci. Soc. 1921, 36, 146–196. [Google Scholar]
- Roberts, P.J.; Spooner, B.M. Cantharelloid, clavarioid and thelephoroid fungi from Brunei Darussalam. Kew Bull. 2000, 55, 843–851. [Google Scholar] [CrossRef]
- Das, K.; Manoj, E.; Ghosh, A.; Parihar, A.; Kuhar, F. Thelephora sikkimensis sp. nov. (Thelephoraceae) from the eastern himalayas (India). Nova Hedwig. 2018, 107, 337–347. [Google Scholar] [CrossRef]
- Sha, T.; Xu, J.P.; Palanichamy, M.G.; Zhang, H.B.; Li, T.; Zhao, Z.W.; Zhang, Y.P. Genetic diversity of the endemic gourmet mushroom Thelephora ganbajun from south-western China. Microbiology 2008, 154, 3460–3468. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Yang, S.R.; Wei, Y.L.; Yuan, H.S. Molecular phylogeny and morphology reveal four new species of Thelephora (Thelephorales, Basidiomycota) from subtropical China, closely related to T. ganbajun. Front. Microbiol. 2023, 14, 1109924. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Pham, T.H.G. New species of Entoloma subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from central Vietnam. J. Fungi 2023, 9, 621. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon Der Farben—1440 Farbnuancen Und 600 Farbnamen; Muster-schmidt: Göttingen, Germany, 1981; pp. 1–241. [Google Scholar]
- Niemel, T.; Dai, Y.C.; Kinnunen, J.; Schigel, D.S. New and in north Europe rare polypore species (Basidiomycota) with annual, monomitic basidiocarps. Karstenia 2004, 44, 67–77. [Google Scholar] [CrossRef]
- Cao, T.; Hu, Y.P.; Yu, J.R.; Wei, T.Z.; Yuan, H.S. A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China—Science Direct. Stud. Mycol. 2021, 99, 100121. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Pérez, A.; Guzmán-Dávalos, L.; Ramírez-Cruz, V.; Villalobos-Arámbula, A.R.; Ruiz-Sanchez, E.; Ramírez-Guillén, F. New species of bioluminescent Mycena sect. Calodontes (Agaricales, Mycenaceae) from Mexico. J. Fungi 2023, 9, 902. [Google Scholar] [CrossRef] [PubMed]
- Song, H.B.; Bau, T. Conocybe section Pilosellae in China: Reconciliation of taxonomy and phylogeny reveals seven new species and a new record. J. Fungi 2023, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.M.; Liu, H.G.; Gates, G.M.; Wu, F.; Dai, Y.C.; Cooper, J.A. Phylogeny and diversity of the genus Pseudohydnum (Auriculariales, Basidiomycota). J. Fungi 2022, 8, 658. [Google Scholar] [CrossRef]
- Largent, D.L.; Johnson, D.; Watling, R. How to Identify Mushrooms to Genus III: Microscopic Features; Mad River Press: Eureka, America, 1977; pp. 1–148. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes–application to the identification of mycorrhizae and rusts. Mol. Ecol. 2010, 2, 113–118. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L.; Tolgor, B. Phylogenetic and biogeographic relationships of chroogomphus species as inferred from molecular and morphological data. Fungal Divers. 2009, 38, 85–104. [Google Scholar]
- Li, Y.C.; Feng, B.; Yang, Z.L. Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 2011, 49, 125–143. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Three new species of Fusicolla (Hypocreales) from China. J. Fungi 2023, 9, 572. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Karunarathna, S.C.; Xu, R.; Lu, Y.; Suwannarach, N.; Mapook, A.; Bao, D.; Xu, J.; Tibpromma, S. Three new species, two new records and four new collections of Tubeufiaceae from Thailand and China. J. Fungi 2022, 8, 206. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Hibbett, D. Resuspending Freeze-Dried Oligonucleotides. Available online: https://www2.clarku.edu/faculty/dhibbett/Protocols_Folder/Primers/Primers.htm (accessed on 1 September 2022).
- Villarruel-Ordaz, J.L.; Maldonado-Bonilla, L.D.; Sánchez-Espinoza, A.C.; Garibay-Orijel, R.; Álvarez-Manjarrez, J. Thelephora pacifica (Basidiomycota: Thelephorales), a new species of the tropical forests in the Mexican pacific coast. Phytotaxa 2024, 634, 204–214. [Google Scholar] [CrossRef]
- Khalid, A.N.; Hanif, M. Thelephora iqbalii sp. nov. from the himalayan moist temperate forests of Pakistan. Mycotaxon 2017, 132, 943–950. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.V.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovli, I.; Zou, H.; Zhang, J.; Wen, X.L.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Fries, E.M. Systema Mycologicum: Sistens Fungorum Ordines, Genera et Species, huc Usque Cognitas, quas ad Normam Methodi Naturalis Determinavit; Lundae, Ex Officina Berlingiana: Lund, Swedish, 1821; Volume 1, p. 432. [Google Scholar]
- Weldon, A.L.; Ovrebo, C.L. Notes on tropical and warm temperate Basidiomycetes. Mycotaxon 1989, 34, 601–614. [Google Scholar]
- Seifert, K.A.; Samson, R.A.; deWaard, J.R.; Houbraken, J.; Levesque, C.A.; Moncalvo, J.M.; Louis-Seize, G.; Hebert, P.D.N. Prospects for fungus identification using CO1 DNA barcodes, with Penicillium as a test case. Proc. Nati. Acad. Sci. USA 2007, 104, 3901–3906. [Google Scholar] [CrossRef] [PubMed]
- Simon, U.K.; Weiss, M. Intragenomic variation of fungal ribosomal genes is higher than previously thought. Mol. Biol. Evol. 2008, 25, 2251–2254. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.; Douhan, G.W.; Rizzo, D.M. Intra-specific and intra-sporocarp ITS variation of ectomycorrhizal fungi as assessed by rDNA sequencing of sporocarps and pooled ectomycorrhizal roots from a Quercus woodland. Mycorrhiza 2007, 18, 15–22. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Hao, Y.J.; Guo, T.; Yang, Z.L.; Cai, Q. Species diversity of Amanita section Vaginatae in eastern China, with a description of four new species. J. Fungi 2023, 9, 862. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L. The boletes of China: Tylopilus s.l.; Springer: Singapore, 2021; pp. 1–418. [Google Scholar]
Figure 1.
Phylogenetic relationships between species of Thelelphora inferred from ITS dataset using the Maximum Likelihood and Bayesian inference approaches (ML topology is shown). ML bootstrap support (≥50) and Bayesian posterior probability (≥0.95) are shown at the branches (BS/PP). Sequences newly generated in this study are highlighted in red. Vouchers are indicated after the species names. Odontia fibrosa and O. ferruginea were used as the outgroup taxa.
Figure 1.
Phylogenetic relationships between species of Thelelphora inferred from ITS dataset using the Maximum Likelihood and Bayesian inference approaches (ML topology is shown). ML bootstrap support (≥50) and Bayesian posterior probability (≥0.95) are shown at the branches (BS/PP). Sequences newly generated in this study are highlighted in red. Vouchers are indicated after the species names. Odontia fibrosa and O. ferruginea were used as the outgroup taxa.

Figure 2.
Fresh basidiomes. (a–c) Thelephora dactyliophora ((a,b) from KUN-HKAS131943, type; (c) from KUN-HKAS131941); (d,e) Thelephora lacunosa (KUN-HKAS128968, type); (f,g) Thelephora petaloides ((f) from KUN-HKAS97730, type; (g) from KUN-HKAS128969); (h,i) Thelephora pinnatifida ((h) from KUN-HKAS96412, type; (i) from KUN-HKAS131946); (j–l) Thelephora sikkimensis ((j,k) from KUN-HKAS128965, type; (l) from KUN-HKAS69236). Bars = 20 mm.
Figure 2.
Fresh basidiomes. (a–c) Thelephora dactyliophora ((a,b) from KUN-HKAS131943, type; (c) from KUN-HKAS131941); (d,e) Thelephora lacunosa (KUN-HKAS128968, type); (f,g) Thelephora petaloides ((f) from KUN-HKAS97730, type; (g) from KUN-HKAS128969); (h,i) Thelephora pinnatifida ((h) from KUN-HKAS96412, type; (i) from KUN-HKAS131946); (j–l) Thelephora sikkimensis ((j,k) from KUN-HKAS128965, type; (l) from KUN-HKAS69236). Bars = 20 mm.

Figure 3.
Basidiospores under SEM. (a–c) Thelephora dactyliophora (KUN-HKAS131943, type); (d–f) Thelephora lacunosa (KUN-HKAS128968, type); (g–i) Thelephora petaloides (KUN-HKAS97730, type); (j–l) Thelephora pinnatifida (KUN-HKAS96412, type); (m–o) Thelephora sikkimensis (KUN-HKAS128965, type). Bars = 2 µm.
Figure 3.
Basidiospores under SEM. (a–c) Thelephora dactyliophora (KUN-HKAS131943, type); (d–f) Thelephora lacunosa (KUN-HKAS128968, type); (g–i) Thelephora petaloides (KUN-HKAS97730, type); (j–l) Thelephora pinnatifida (KUN-HKAS96412, type); (m–o) Thelephora sikkimensis (KUN-HKAS128965, type). Bars = 2 µm.

Figure 4.
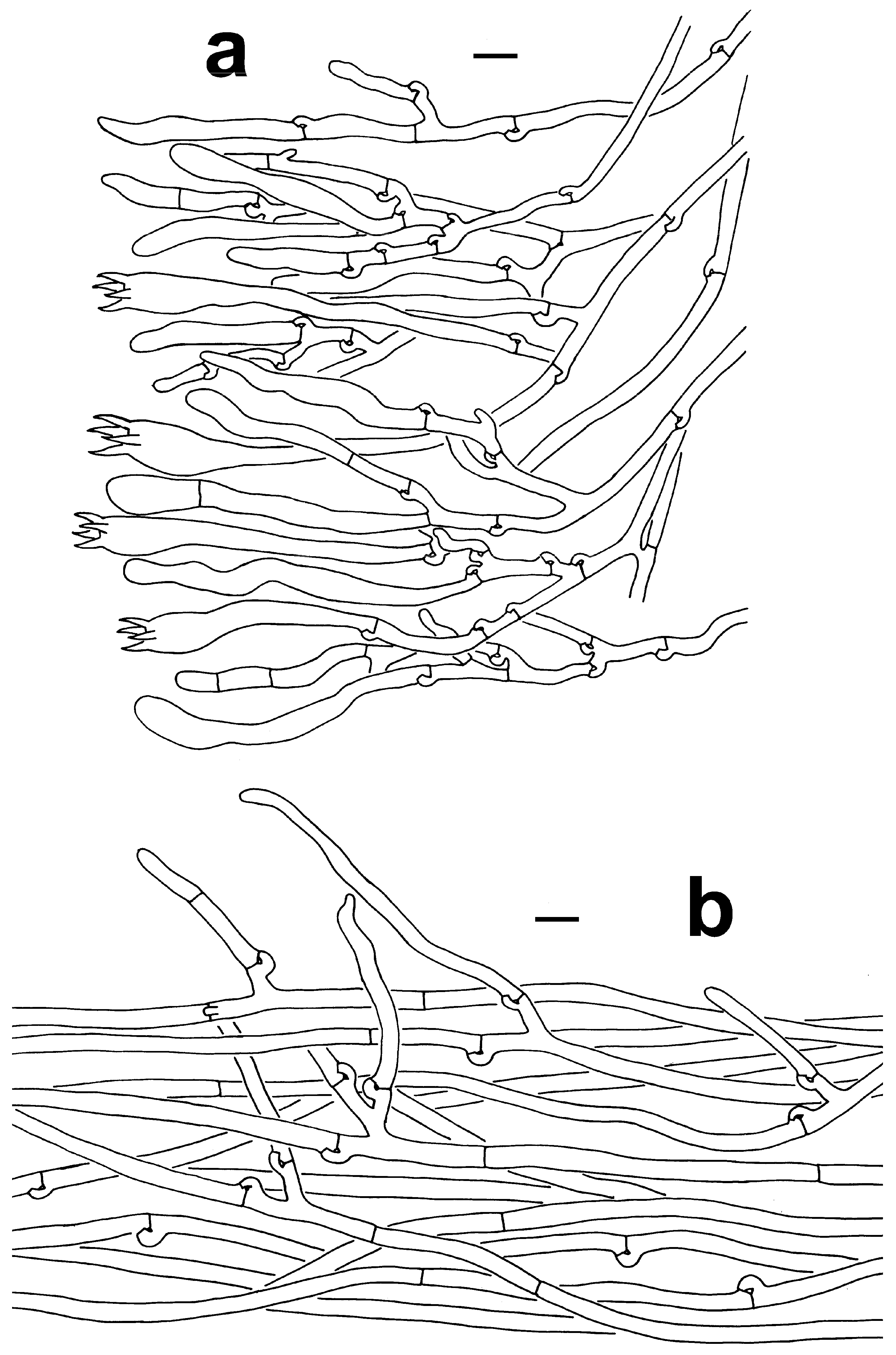
Microscopic features of T. dactyliophora (KUN-HKAS131943, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm.
Figure 4.
Microscopic features of T. dactyliophora (KUN-HKAS131943, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm.

Figure 5.
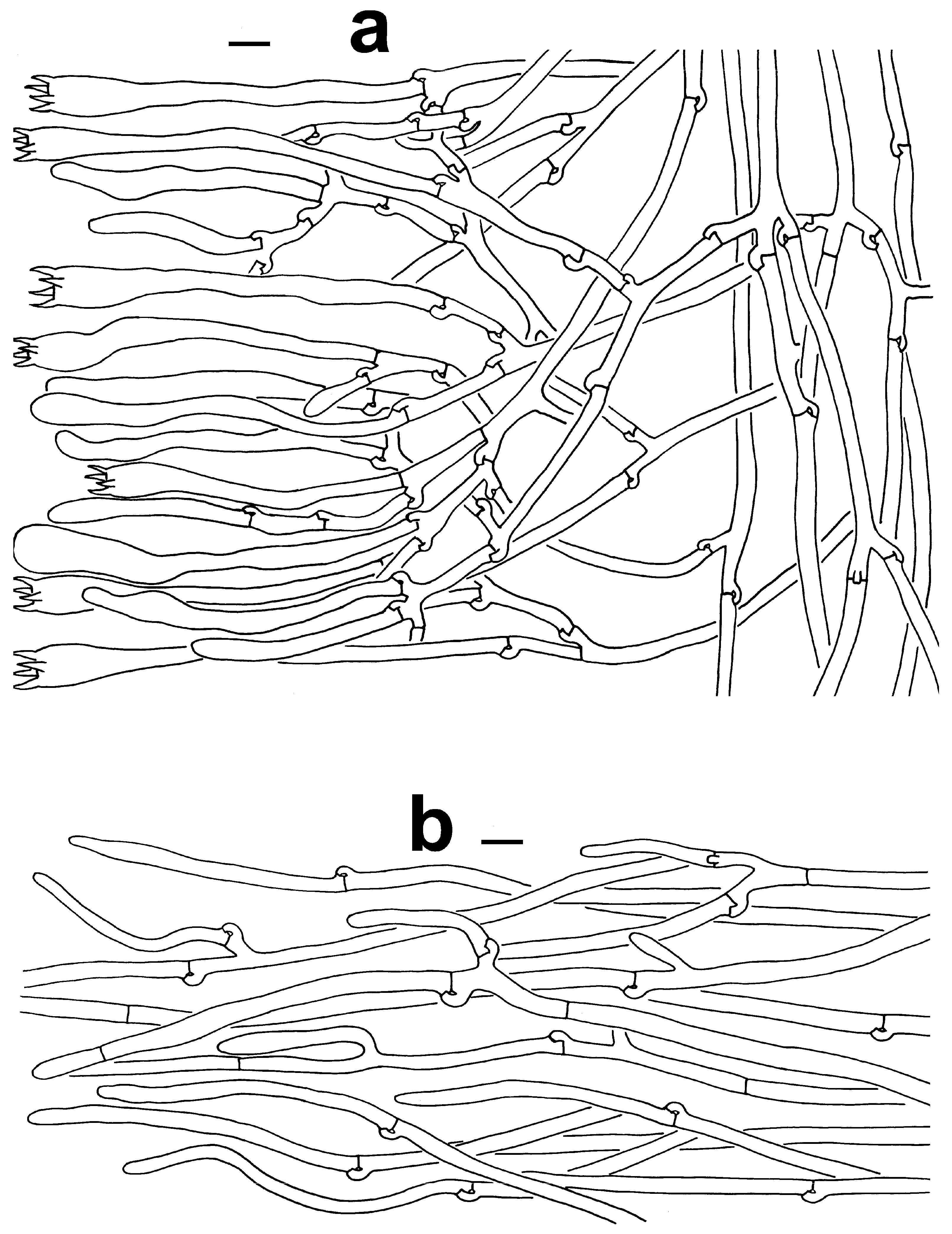
Microscopic features of T. lacunosa (KUN-HKAS128968, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm. Bars = 2 µm.
Figure 5.
Microscopic features of T. lacunosa (KUN-HKAS128968, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm. Bars = 2 µm.

Figure 6.
Microscopic features of T. petaloides (KUN-HKAS97730, type). (a) Hymenium and subhymenium (KUN-HKAS97730, type). (b) Pileipellis. Bars = 10 µm.
Figure 6.
Microscopic features of T. petaloides (KUN-HKAS97730, type). (a) Hymenium and subhymenium (KUN-HKAS97730, type). (b) Pileipellis. Bars = 10 µm.

Figure 7.
Microscopic features of T. pinnatifida (KUN-HKAS96412, type). (a) Hymenium and subhymenium. (b) Hair-like appendages of pointed tips. Bars = 10 µm.
Figure 7.
Microscopic features of T. pinnatifida (KUN-HKAS96412, type). (a) Hymenium and subhymenium. (b) Hair-like appendages of pointed tips. Bars = 10 µm.

Figure 8.
Microscopic features of T. sikkimensis (KUN-HKAS128965, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm.
Figure 8.
Microscopic features of T. sikkimensis (KUN-HKAS128965, type). (a) Hymenium and subhymenium. (b) Pileipellis. Bars = 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Species, vouchers, GenBank/UNITE accessions, and localities of specimens used in this study. Sequences newly generated in this study are shown in bold.
Table 1.
Species, vouchers, GenBank/UNITE accessions, and localities of specimens used in this study. Sequences newly generated in this study are shown in bold.
| Taxa | Voucher | GenBank/UNITE No. | Locality | Reference |
|---|---|---|---|---|
| ITS | ||||
| Thelephora lacunosa | KUN-HKAS128966 | OR512336 | China | This study |
| T. lacunosa | KUN-HKAS128967 | OR512337 | China | This study |
| T. lacunosa | KUN-HKAS128971 | OR512338 | China | This study |
| T. lacunosa | KUN-HKAS128968 | OR512335 | China | This study |
| T. petaloides (labeled as T. sikkimensis) | KUN-HKAS122715 | ON794445 | China | GenBank |
| T. petaloides | KUN-HKAS128969 | OR512332 | China | This study |
| T. petaloides | KUN-HKAS128970 | OR512333 | China | This study |
| T. petaloides | KUN-HKAS97730 | OR512334 | China | This study |
| T. sikkimensis | KUN-HKAS128964 | OR512329 | China | This study |
| T. sikkimensis | KUN-HKAS128965 | OR512331 | China | This study |
| T. sikkimensis | KUN-HKAS128972 | OR512330 | China | This study |
| T. sikkimensis | KUN-HKAS128963 | OR512328 | China | This study |
| T. sikkimensis | KD 16-042 | MF684018 | India | [22] |
| T. wuliangshanensis | CLZhao 3406 | MZ400677 | China | [14] |
| T. wuliangshanensis | CLZhao 3408 | MZ400678 | China | [14] |
| T. vialis | TENN-F-072094 | MN121022 | USA | [25] |
| T. vialis | TENN-F-072281H2 | MN121029 | USA | [25] |
| T. pseudoganbajun | YL3-10 | KY245255 | China | [25] |
| T. pseudoganbajun | Yuan16794 | OP793768 | China | [25] |
| T. ganbajun | KUN-HKAS14735 | KY245240 | China | GenBank |
| T. ganbajun | Yuan16715 | OP793760 | China | [25] |
| T. dactyliophora | KUN-HKAS131941 | OR940521 | China | This study |
| T. dactyliophora | KUN-HKAS131943 | OR940523 | China | This study |
| T. pacifica | VO2019_125 | OR548196 | Mexico | [40] |
| T. pacifica | VO2019_101 | OR548195 | Mexico | [40] |
| T. penicillata | X619 | OL469899 | Prague | GenBank |
| T. penicillata | X618 | OL469898 | Prague | GenBank |
| T. pinnatifida | KUN-HKAS131946 | OR940524 | China | This study |
| T. pinnatifida | KUN-HKAS131947 | OR940525 | China | This study |
| T. pinnatifida | KUN-HKAS96412 | OR940526 | China | This study |
| T. iqbalii | MH810 | JX241471 | Pakistan | [41] |
| T. glaucoflora | Dai13623A | OP793751 | China | [25] |
| T. glaucoflora | Dai16612 | OP793754 | China | [25] |
| T. grandinioides | CLZhao 4107 | MZ400675 | China | [14] |
| T. grandinioides | CLZhao 21020 | MZ400676 | China | [14] |
| T. nebula | Yuan11516 | OP793746 | China | [25] |
| T. nebula | Yuan11515 | OP793745 | China | [25] |
| T. austrosinensis | GDGM48867 | MF593265 | China | [13] |
| T. austrosinensis | GDGM25680 | MF593261 | China | [13] |
| T. aquila | Wei8831 | OP793743 | China | [25] |
| T. aquila | Wei8833 | OP793744 | China | [25] |
| T. palmata | JMP0085 | EU819443 | USA | GenBank |
| T. palmata | iNAT:30911260 | MZ276274 | USA | GenBank |
| T. palmata | UBC: F33078 | MF908479 | Canada | GenBank |
| T. palmata | JLF3733 | MK847520 | USA | GenBank |
| T. pseudoversatilis | FCME26152 | KJ462486 | Mexico | [17] |
| T. pseudoversatilis | FCME26232 | JX075890 | Mexico | [17] |
| T. versatilis | MEXU27094 | KC595628 | Mexico | [17] |
| T. versatilis | FCME26141 | NR154492 | Mexico | [17] |
| T. terrestris | Hilszczanska D. 1-IBL | FJ532478 | Poland | GenBank |
| T. terrestris | MT1-2203-3 | EU427323 | Finland | GenBank |
| Odontia fibrosa | SS38 | MH310788 | Sweden | [14] |
| O. ferruginea | UK18 | UDB000285 | Estonia | [14,18,22] |
Table 2.
Comparisons among species of Thelephora in this study.
| Structures | T. sikkimensis | T. petaloides | T. lacunosa | T. dactyliophora | T. pinnatifida |
|---|---|---|---|---|---|
| Basidiome | Imbricate to rosette, small, 45–67 × 50–70 mm, with flabelliform to petaloid branches | Imbricate to umbrella, large, 40–100 × 55–220 mm, with flabelliform, applanate-lobate branches | Imbricate, rosette to flabelliform, medium-sized, 67–80 × 80–150 mm, with flabelliform, petaloid to applanate-lobate branches | Coralloid, small, 35–65 × 30–40 mm, with spathulate to narrow petaloid branches | Clavarioid, small, 35–45 × 28–85 mm, with clavate to pinnatifid or ramiform branches with tips needle-like |
| Abhymenium | Sulcate, indistinctly zonate, radially rugulose or wrinkled; grayish black at center, purplish gray to grayish green elsewhere, but chalky white to orange-white at margin | Sulcate to irregularly ridged, zonate, glabrous; brownish gray at center, orange-yellow to grayish orange or pale orange elsewhere, but chalky white at margin | Lacunal, sulcate, irregularly ridged, non-zonate, and warty; brown, grayish brown to grayish orange at center, much paler towards margin, but white to pale orange-gray at margin | Slightly rugulose, non-zonate; grayish yellow at center, brownish gray to gray, but chalky white to orange-white at margin | Without abhymenium |
| Hymenium | Imperceptibly rugose, warty, non-zonate, gradually from smooth to irregularly plicate near margin; pale orange to purplish gray or grayish green, but orange-white to almost white at margin | Imperceptibly rugose, warty, zonate near margin; orange-gray at center, bright orange elsewhere, but chalky white at margin | Imperceptibly rugose to warty, non-zonate; brownish orange to gray or purple-gray at center, pale orange towards margin, but orange-white at margin | Rugulose, non-zonate; brownish gray to gray, but chalky white to orange-white at margin | Predominantly amphigenous, sulcate, non-zonate; visibly ribbed; brownish orange to brown, but chalky white to orange-white at margin |
| Stipe | Irregularly cylindrical, 12–14 × 2–9 mm; surface rugose to warty, pale orange, sometimes purplish gray | Irregularly cylindrical to flatted or broadened at base, 10–40 × 9–13 mm; surface rugose, glabrous, brownish gray | Subconical to broadened or flatted towards base, 20–25 × 12–35 mm; surface rugose, glabrous, brownish red to brownish yellow | Irregularly cylindrical to flatted or broadened at base, 10–20 × 2–5 mm; surface smooth to slightly rugose, brownish gray | Irregularly cylindrical, 0–15 × 0–10 mm; surface rugose, brown |
| Context | 1–2 mm thick | 1–4 mm thick | 0.5–3.5 mm thick | 0.1–0.5 mm thick | 0.1–2 mm thick |
| Basidia | 72–100 × 7.5–10 µm | 53–92 × 9–12 µm | 72–100 × 8–12 µm | 45–72 × 6–9 µm | 47–90 × 10–11.5 µm |
| Basidiospore | 4.5–6.5 × 4–5.5 µm | 4.5–7.5 × 3.5–6.5 µm | 4.5–6.5 × 4–5.5 µm | 4–8.5 × 4–6.5 µm | 5.5–9.5 × 5–8.5 µm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tian, M.-Z.; Xia, H.-B.; Gao, Z.-L.; Zhao, C.-Y.; Ma, D.; Yang, Z.-L.; Li, Y.-C. Four New Species and One New Record of Thelephora from China. J. Fungi 2024, 10, 300. https://doi.org/10.3390/jof10040300
AMA Style
Tian M-Z, Xia H-B, Gao Z-L, Zhao C-Y, Ma D, Yang Z-L, Li Y-C. Four New Species and One New Record of Thelephora from China. Journal of Fungi. 2024; 10(4):300. https://doi.org/10.3390/jof10040300
Chicago/Turabian StyleTian, Mei-Zhi, Hai-Bin Xia, Zheng-Lin Gao, Chang-Yin Zhao, Dan Ma, Zhu-Liang Yang, and Yan-Chun Li. 2024. "Four New Species and One New Record of Thelephora from China" Journal of Fungi 10, no. 4: 300. https://doi.org/10.3390/jof10040300
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.