Two-Stage Aeration Fermentation Strategy to Improve Bioethanol Production by Scheffersomyces stipitis



CICECO—Aveiro Institute of Materials, Chemistry Department, University of Aveiro, Campus Universitário de Santiago, 3810-193 Aveiro, Portugal
*
Author to whom correspondence should be addressed.
Fermentation 2018, 4(4), 97; https://doi.org/10.3390/fermentation4040097
Submission received: 28 October 2018
/
Revised: 15 November 2018
/
Accepted: 16 November 2018
/
Published: 20 November 2018
(This article belongs to the Special Issue Microbial Production of Added-value Products from Renewable Resources)
Abstract
:Hardwood spent sulfite liquor (HSSL) is a by-product from pulp industry with a high concentration of pentose sugars, besides some hexoses suitable for bioethanol production by Scheffersomyces stipitis. The establishment of optimal aeration process conditions that results in specific microaerophilic conditions required by S. stipitis is the main challenge for ethanol production. The present study aimed to improve the ethanol production from HSSL by S. stipitis through a two-stage aeration fermentation. Experiments with controlled dissolved oxygen tension (DOT) in the first stage and oxygen restriction in the second stage were carried out. The best results were obtained with DOT control at 50% in the first stage, where the increase of oxygen availability provided faster growth and higher biomass yield, and no oxygen supply with an agitation rate of 250 rpm, in the second stage allowed a successful induction of ethanol production. Fermentation using 60% of HSSL (v/v) as substrate for S. stipitis provided a maximum specific growth rate of 0.07 h−1, an ethanol productivity of 0.04 g L h−1 and an ethanol yield of 0.39 g g−1, respectively. This work showed a successful two-stage aeration strategy as a promising aeration alternative for bioethanol production from HSSL by S. stipitis.
1. Introduction
In 2016, according to the International Energy Agency (IEA), 81.1% of all energy produced worldwide came from fossil fuels, with the transport sector accounting for 64.5% of all oil consumed [1]. However, owing to the increasing demand for energy, the more and more expensive extraction of fossil fuels, and the high amount of greenhouse gases emitted by their utilization, finding alternative energy sources became urgent. Biofuels are an alternative to fossil fuels with great potential, as they are produced from renewable raw materials and allow the CO2 released from their combustion to be recycled through photosynthetic pathways [2].
Bioethanol is the most widely used biofuel in the transportation sector worldwide, being commonly blended with gasoline in proportions from 5–85% in bioethanol [3]. Nevertheless, first-generation bioethanol, produced from food feedstocks, has raised the food prices in international markets, triggering serious social problems. Therefore, strategies that allow the commercial viability of the second-generation bioethanol, produced from lignocellulosic biomass, are needed [2].
Hardwood spent sulfite liquor (HSSL) is a by-product from the acidic sulfite pulping process of wood in the pulping industry [4]. HSSL is produced in large quantities, being generated around 8 m3 of HSSL per ton of pulp produced [5]. During the hard acidic sulfite pulping process wood delignification takes place for subsequent celluloses recovery, as pulps, and simultaneously the majority of hemicelluloses is hydrolyzed to monosaccharides, remaining soluble in the HSSL. The high content in monomeric sugars makes HSSL an advantageous feedstock since it can be directly fermented to ethanol avoiding the usual difficult and expensive step of polysaccharides hydrolysis required by most of the lignocellulosic biomass. Therefore, HSSL has a great potential as raw material for the production of second-generation bioethanol, due its high content in fermentable sugars and by applying this strategy costs associated to its elimination can be reduced. An efficient production of bioethanol from HSSL can be a way to valorize this by-product increasing the sustainability of the pulping industry through the implementation of a biorefinery in pulping mil industrial plants [4].
Pentoses, mainly xylose, are the largest fraction of sugars in HSSL with minor amounts of hexoses [6]. Among the pentose-fermenting yeasts, Scheffersomyces stipitis, formerly classified as Pichia stipitis [7], is one of the microorganisms with the highest native capacity to ferment xylose [8]. Its metabolism starts with the pentose phosphate pathway and proceeds with the glycolytic pathway. Then, S. stipitis uses aerobic energetic pathways for respiration and cell biosynthesis or alternatively uses anaerobic fermentative pathways producing ethanol and less energy. The utilization of xylose must be carefully regulated in order to optimize bioethanol fermentation over growth. Several studies were already attempted with this yeast, showing that S. stipitis can lead to successful bioethanol production from HSSL [4]. Nevertheless, bioethanol production from HSSL by S. stipitis still presents some challenges that need to be overcome in order to enable commercial production. One of the main challenges is the low tolerance of S. stipitis towards microbial inhibitors found in HSSL [4,9], namely lignosulfonates, acetic acid and low molecular weight phenolic compounds [6]. Among the pretreatments already tested, a biological detoxification using filamentous fungus was successful applied to HSSL. Paecilomyces variotii was able to consume some inhibitors with residual sugars consumption. The remaining sugars were later used for bioethanol production by S stipitis [10]. However, a different strategy, the development of HSSL-adapted strains by evolutionary engineering, can mitigate the adverse effect of the inhibitors of HSSL [11,12]. Pereira et al. [11] obtained a stable clone, S. stipitis C4, from a population of S. stipitis adapted to a medium composed of 60% of HSSL in a continuous stirred tank reactor (CSTR) able to produce bioethanol with a good efficiency. Nonetheless, the establishment of optimal aeration conditions is still the main obstacle to the success of S. stipitis in bioethanol production, since this yeast requires a tight control of oxygen availability during ethanol production [12,13]. The availability of oxygen should be enough for cell growth and maintenance but must be limited in order to prevent excessive cell growth [14,15] and reassimilation of the produced ethanol [16]. The best aeration conditions for ethanol production by S. stipitis were investigated by several authors [12,13,17,18]. The effect of volumetric oxygen transfer coefficient (KLa), oxygen transfer rate (OTR), and dissolved oxygen tension (DOT) on ethanol production by S. stipitis was evaluated. Concerning KLa, the values providing the best balance between productivity and yield were described in the range between 2.3 and 5.9 h−1 [19,20]. Regarding DOT, values below 1% of saturation were found to enhance ethanol production by S. stipitis [21]. An optimum OTR value of 2.0 mmolO2 L−1 h−1 was found for HSSL fermentation by an adapted strain of S. stipitis [12].
The present study aimed to improve the ethanol production from HSSL by S. stipitis C4 through a two-stage aeration strategy in order to regulate the culture behavior by promoting aerobic metabolism to attain enough cell concentration and then by promoting microaerophilic metabolism to obtain significant ethanol production. Firstly, the effect of DOT on the growth kinetics of S. stipitis C4 was characterized. Then, a two-stage aeration fermentation was studied using synthetic medium. During the first stage, the yeast growth should be maximized by aeration, while during the second stage, oxygen availability conditions were changed in order to enhance bioethanol production. Finally, the performance of S. stipitis C4 in the fermentation of 60% HSSL/40% synthetic medium (v/v) was assessed by applying the aeration conditions studied that allowed the most efficient bioethanol production in the previous assays. To our knowledge, this is the first time that a two-stage aeration strategy was applied to bioethanol production by S. stipitis using HSSL. The results of this work are expected to contribute to the establishment of an industrial HSSL-based biorefinery.
2. Materials and Methods
2.1. HSSL Supply and Pre-Treatment
HSSL from magnesium-based acidic sulfite pulping of Eucalyptus globulus was supplied by Caima-Indústria de Celulose, ALTRI (Constância, Portugal). Pre-evaporated HSSL was collected from an inlet evaporator from a set of multiple-effect evaporators to avoid the presence of free SO2. To remove part of the lignosulfonates and other recalcitrant compounds, HSSL was submitted to a preliminary pretreatment [10]. The pretreatment started with a pH adjustment to 7.0 with 6 M KOH, followed by aeration with compressed air (2880 L of compressed air per liter of HSSL in 6 h). Then, the liquor was centrifuged for 1 h at 5000 rpm. The precipitated colloids were filtered using a 1 µm glass microfiber filter. Finally, the pre-treated HSSL was stored at 4 °C.
2.2. Microorganism
The strain S. stipitis C4 was obtained in a previous work of evolutionary engineering. In this work it was selected from an adapted population of S. stipitis NRRL Y-7124 to HSSL increasing concentrations as described by Pereira et al. [11]. The strain S. stipitis C4 was grown at 28.0 ± 0.5 °C and maintained at 4 °C on YMxyl solid medium (Yeast Mold solid medium supplemented with xylose).
2.3. Culture Media
YMxyl solid medium was prepared with the following composition: 3.0 g L−1 yeast extract; 3.0 g L−1 malt extract; 10.0 g L−1 glucose; 40.0 g L−1 xylose; and 2.0% (m/v) agar. The pH of YMxyl solid medium was adjusted to 5.5.
Synthetic medium was prepared in order to have the same sugars concentration as HSSL. It was prepared with the following composition: 5.0 g L−1 glucose; 40.0 g L−1 xylose; 2.5 g L−1 yeast extract; 2.0 g L−1 (NH4)2HPO4; 1.0 g L−1 (NH4)2SO4; and 0.5 g L−1 MgSO4∙7H2O. Synthetic medium was buffered at pH 5.5 with 25 mM potassium hydrogen phthalate.
2.4. Pre-Inocula and Inocula
Pre-inocula were prepared by transferring a colony of S. stipitis C4 from maintenance YMxyl solid medium to a 100 mL Erlenmeyer flask containing 40 mL of synthetic medium and was incubated at 180 rpm for 24 h at 28.0 ± 0.5 °C in orbital shaker (B. Braun® Certomat® S, Melsugen, Germany). After this period, the inoculum was prepared by transferring the adequate volume of pre-inoculum to a 500 mL Erlenmeyer flask with fresh synthetic medium in order to obtain an initial optical density of 0.4 and a final volume of 200 mL. The inoculum was incubated for 24 h as described for pre-inocula.
2.5. Assays
Fermentations were carried out in batch in a 5 L BIOSTAT Aplus bioreactor (Sartorius Stedim Biotech®, Gottingen, Germany) with a working volume of 3 L and automatic control by micro-DCU software (Sartorius Stedim Systems, Gottingen, Germany), and data acquisition (MFCS/DA 3.0 Sartorius Stedim Systems). The temperature was controlled at 28 °C and the pH was controlled at 5.5 by the addition of 5 M KOH and 1 M H2SO4. DOT was measured by a OxyFerm® FDA 325 electrode (Hamilton®, Reno, NV, USA), and automatically controlled. Stirring was achieved by a six-blade Rushton turbine, 3 cm above from the ring disperser, and by four baffles. When necessary, antifoam (Silicone 426 R, 10%, v/v) was added. Several assays of single-stage aeration and two-stage aerations were performed. In every assay, the reactor was inoculated by transferring the required volume of inoculum to obtain an initial optical density of 0.4. Single-stage aeration and two-stage aeration experiments were made with synthetic medium, the second stage started when an optical density of 14 was reached. In the two-stage aeration experiment with 60% HSSL/40% synthetic medium (v/v), the second stage started when an optical density of 20 was reached. The conditions applied in every assay are described in Table 1. In experiments with fixed DOT, the aeration flow and the stirring rate were controlled by the bioreactor.
Samples were collected throughout the operational time for biomass quantification and further analysis. Samples were centrifuged at 13,000 rpm for 10 min, the pellet discarded, and the pH of the supernatant measured before storage under −16 °C for later determination of glucose, xylose, acetic acid, xylitol, ethanol, and magnesium concentrations. Five milliliter samples were collected at the end of each assay for the determination of cell dry weight.
2.6. Analytical Methods
Biomass was monitored by measuring the optical density at 620 nm (Shimadzu® UVmini-1240®, Kyoto, Japan). Subsequently, the optical density was converted into dry weight concentration through the appropriated calibration curve.
Cell dry weight was determined at the end of each assay by filtration of 5.00 mL of sample using 0.45 µm pore diameter membranes (Whatman® ME 25/21 ST, Maidstone, UK). After filtration, the filter was dried at 105 °C until constant weight.
Glucose, xylose, acetic acid, xylitol, and ethanol concentrations were determined by high-performance liquid chromatography (HPLC). For this, all samples were centrifuged and filtered off with centrifuge tube 0.20-μm filters CoStar Spin-X (Corning, NY, USA) at 8000 rpm for 20 min, properly diluted and acidified with 4 % (v/v) H2SO4 0.25 M before the analysis 650 μL of each sample. Samples were then injected (Auto-sampler HITACHI L-2200, Hitachi, Ltd., Chiyoda, Japan) in an ion exchange column Aminex HPX-87H (Bio-Rad, Hercules, CA, USA) at 40 °C (Oven Gecko-2000, CIL Cluzeau, Sainte-Foy-la-Grande, France), and analyzed by a refractive index detector (HITACHI L-2490, Hitachi, Ltd., Chiyoda, Japan). The eluent 0.01 N H2SO4 was pumped at a flow rate of 0.6 mL/min (HITACHI L-2130 pump, Hitachi, Ltd., Chiyoda, Japan). Standard calibration curves were obtained by injecting standards for all analyzed compounds. Magnesium (Mg2+) concentration was determined by inductively coupled plasma optical emission spectroscopy (ICP-OES-Horiba Jobin Yvon Activa, Kyoto, Japan). Prior to ICP-OES analysis, 1.0 mL of sample was acidified by adding HNO3 until a concentration of 2% (v/v). Finally, the acidified sample was injected into the Horiba Jobin Yvon Activa M ICP-OES system.
2.7. Determination of Volumetric Oxygen Transfer Coefficient (KLa)
KLa was determined by the static method of gassing out in the absence of microorganisms [22]. After switching off the aeration, the fermentative medium was sparged with nitrogen for oxygen stripping, the air flow was restarted and the DOT values were recorded over time. The KLa value was calculated by the integrated form of Equation (1) that results from a simplification of the equation of mass balance to oxygen in a bioreactor in batch mode [23], where the value of KLa corresponds to the to the slope of the resulting straight line representing the ln () versus time t (h):
where (mol m−3) corresponds to the saturated dissolved oxygen concentration and to the value of dissolved oxygen concentration.
2.8. Calculations
The specific growth rate, µ (h−1), was determined during the exponential phase of cell growth by the slope of the straight line of logarithm of biomass concentration (g L−1) along time (h). The maximum specific growth rate, µMAX (h−1), and saturation constant for oxygen, (%), were determined by non-linear regression through fitting Monod kinetic Equation (2) to experimental data of µ (h−1) versus DOT (%), using GraphPad Prism® 5 software (GraphPad, La Jolla, CA, USA).
Glucose volumetric consumption rate, rglucose (g L−1 h−1), xylose volumetric consumption rate, rxylose (g L−1 h−1), acetic acid volumetric consumption rate, racetic acid (g L−1 h−1), ethanol volumetric production rate, rethanol (g L−1 h−1), and xylitol volumetric production rate, rxylitol (g L−1 h−1), were calculated using the Equation (3):
where c represents the concentration of glucose, xylose, acetic acid, ethanol, or xylitol. The growth yield, Ybiomass/substrate (g g−1), ethanol yield, Yethanol/substrate (g g−1), and xylitol yield, Yxylitol/xylose (g g−1), were calculated, respectively, by the Equations (4)–(6):
For Ybiomass/substrate, in synthetic medium, substrate was considered the sum of glucose and xylose, while in 60% HSSL/40% synthetic medium (v/v), substrate corresponded to glucose, xylose, and acetic acid. In turn, for Yethanol/substrate, in both synthetic medium and 60% HSSL/40% synthetic medium (v/v), only glucose and xylose were considered as substrate. Lastly, considering a theoretical ethanol yield of 0.51 g g−1, the conversion efficiency (%) was calculated by the Equation (7):
3. Results and Discussion
3.1. Single-Stage Aeration Experiments
S. stipitis growth was studied for DOT values of 1.0%; 2.5%; 10%; 25%; and 50%. None of the DOT values studied resulted in ethanol production (Supplementary Materials, Figure S1). This observation is in accordance with the findings of du Preez et al. [21] that observed the best ethanol yield with a DOT value of 0%, and the best ethanol productivity with DOT controlled at 0.7%. In fact, several studies with S. stipitis without DOT control report, despite the supply of oxygen, the DOT rapidly reaching the value of 0% after the start of fermentation, due to oxygen consumption by the growing cells of S. stipitis [13,19,24,25,26]. The presence of oxygen allowed S. stipitis C4 to reach values of specific growth rate that increased with DOT (Supplementary Materials, Figure S2). In the DOT 50% experiment a µ of 0.44 h−1 was obtained, which was close to the µMAX value. Thus, the DOT value of 50% was high enough to obtain a µ close to the highest specific growth rate that can be achieved under the culture conditions studied.
Although DOT control did not allow ethanol production, µ values close to the maximum specific growth rate were achieved when culture growth was not limited by oxygen. Thereby, the DOT control at 50% is a promising strategy for S. stipitis C4 growth stimulus in HSSL. Thus, the best strategy for ethanol production from HSSL may be through stage with oxygen restriction, in order to promote ethanol production, preceded by a first stage with high DOT, for growth stimulus to maximize the amount of biomass. In the first stage the culture is expected to grow in the best conditions and after attaining a high biomass concentration, the limitation of oxygen will result on the change of metabolism by S. stipitis to convert sugars to bioethanol.
3.2. Two-Stage Aeration Experiments in Synthetic Medium
In the two-stage aeration experiments, DOT was controlled at 50% in the first stage, since the experimental results showed to be the best strategy to stimulate S. stipis growth. In the second stage, three different aeration strategies were tested: one without air supply (0 mLAir min−1 and 250 rpm); another with air supply at a fixed flow rate (50 mLAir min−1 and 150 rpm); and a third one, also with air supply, but with variable air flow rate and stirring speed in order to control the DOT at 1%. The experiments were carried out in synthetic medium and an aeration stage change was performed when the optical density reached about 14.
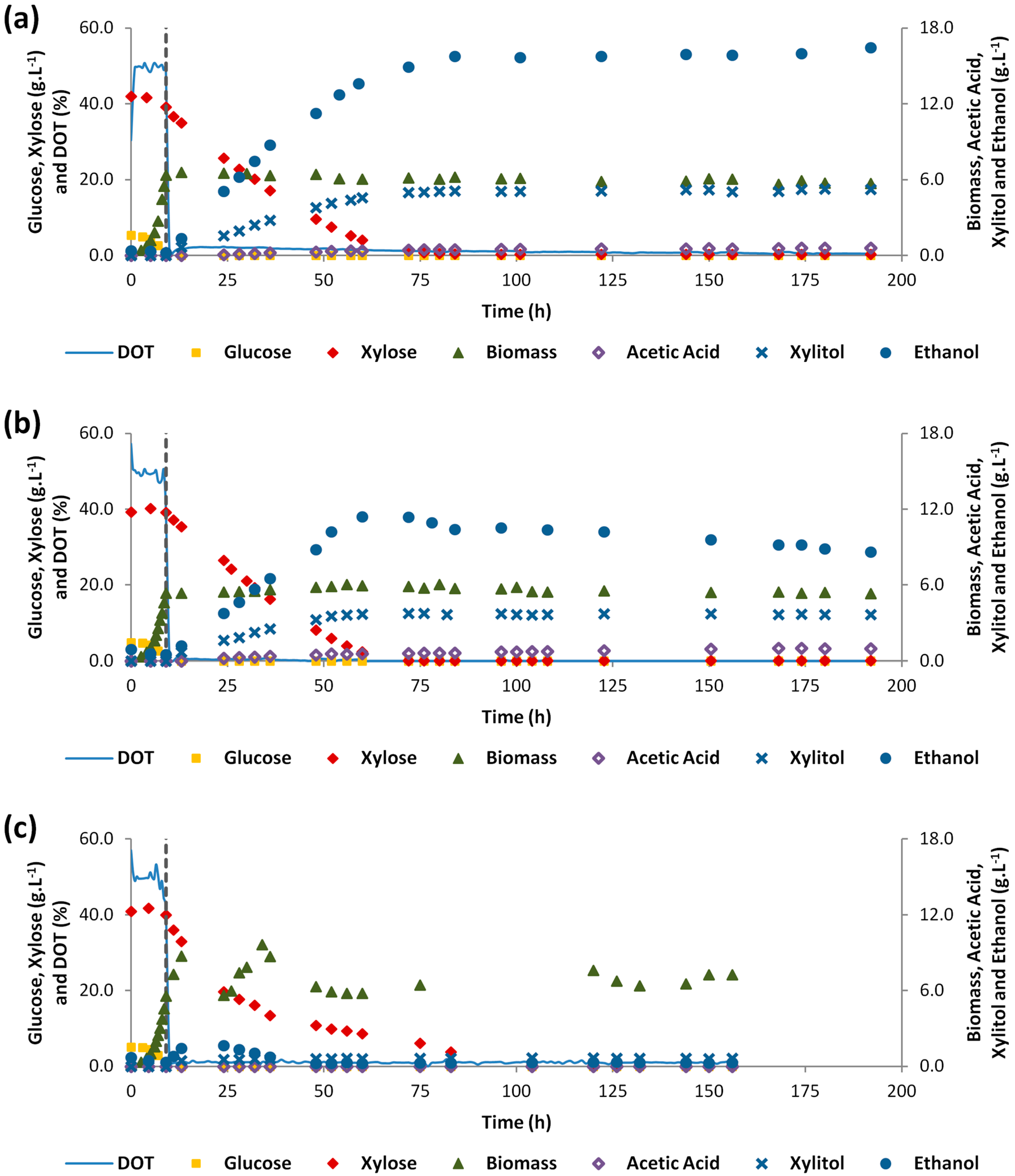
As expected, S. stipitis C4 presented a similar performance in the first aeration stage of the three experiments (Figure 1, Table 2). The experimental results showed that changing from the first aeration stage with oxygen availability, to a second aeration stage, with oxygen limiting conditions, allowed to induce ethanol production and to suppress S. stipitis C4 growth. The exception was the experiment with DOT controlled at 1%, since the amount of biomass almost doubled and low amount of ethanol was produced in the second stage, 1.67 g L−1. The maximum ethanol concentration of the second aeration stage, 15.8 g L−1, and the maximum ethanol yield, 0.40 g g−1, were obtained with 0 mLAir min−1 and 250 rpm imposed in the second stage. The same value of ethanol volumetric production rate, 0.21 g L−1 h−1 was obtained in experiments with 0 mLAir min−1 and 250 rpm and 50 mLAir min−1 and 150 rpm, respectively (Table 2).
The higher Yethanol/substrate in the second aeration stage of the 0 mLAir min−1 and 250 rpm experiment can be explained by the fact that, unlike the 50 mLAir min−1 and 150 rpm experiment, ethanol reassimilation by S. stipitis C4 was not verified, as evidenced by the ethanol plateau obtained after xylose exhaustion in the experiment 0 mLAir min−1 and 250 rpm (Figure 1b). The ethanol reassimilation in the DOT 50%—50 mLAir min−1 and 150 rpm experiment probably resulted from the higher oxygen availability in the second aeration stage of this experiment, since an air flow rate of 50 mL min−1 was supplied. Skoog et al. [16] also observed an increase on the ethanol specific consumption rate increased from 0.13 to 0.17 g g−1 h−1 by S. stipitis after changing from microaerophilic conditions (100 mLAir min−1 and 550 rpm) to aerobic conditions (1000 mLAir min−1 and 1000 rpm), in a continuous stirred tank reactor (CSTR) fed with xylose and ethanol. Thus, the best aeration strategy for the second stage was to operate without air supply, i.e., with 0 mLAir min−1 and 250 rpm, because, in this way, ethanol reassimilation was eliminated and the ethanol yield was maximized.
In the DOT 50%–-50 mLAir min−1 and 150 rpm experiment, the oxygen transfer to fermentation medium during the second aeration stage was performed with a volumetric oxygen transfer coefficient (KLa) of 2.1 h−1, which is a value close to the KLa values reported in literature as optimal for ethanol production by S. stipitis, 2.3–4.9 h−1 [14,20]. However, these literature values were obtained in fermentations with just a single aeration stage, in which a compromise between ethanol and biomass production was required in order to ensure a balance between ethanol yield and productivity [14]. Thus, as fermentations with two aeration stages allowed for a higher cell density in the second stage, the KLa value of this stage could be lower since the objective of the second aeration stage is maximizing the ethanol yield. This finding was confirmed in the experiment with DOT 50%—0 mLAir min−1 and 250 rpm, when the best ethanol results were obtained without any air supply.
In the experiments DOT 50%—0 mLAir min−1 and 250 rpm (Figure 1a) and DOT 50%—50 mLAir min−1 and 150 rpm (Figure 1b) xylitol accumulation was observed during the second aeration stage. Thus, in these two experiments, the maximum xylitol concentration was respectively 5.10 and 3.72 g L−1, the Yxylitol/xylose in the second aeration stage was respectively 0.13 and 0.10 g g−1, and the rxylitol in the second aeration stage was respectively 0.068 and 0.073 g L−1 h−1 (Table 2). In turn, in the experiment DOT 50%—DOT 1% (Figure 1c) xylitol accumulation was not observed. This can be a result from differences in oxygen availability during the second stage. Oxygen availability allowed balancing the ratio between NADH and NAD+, which probably resulted in lower xylitol accumulation as observed by Bellido et al. that observed the reduction of xylitol accumulation with increasing values of KLa [20].
Table 3 shows the global kinetic and stoichiometric parameters of the two-stage aeration experiments in synthetic medium considering both stages. As expected, the best results were obtained in the DOT 50%—0 mLAir min−1 and 250 rpm experiment, in which a maximum ethanol concentration of 15.8 g L−1, a rethanol of 0.18 g L−1 h−1, and a Yethanol/substrate of 0.33 g g−1 were obtained, which corresponded to a conversion efficiency of 64.1%. Furlan et al. [17] also tested the application of a two-stage aeration strategy to ethanol production by S. stipitis from 50 g L−1 of xylose. These authors applied a KLa of 4.8 h−1 (128 mLAir min−1 and 250 rpm) in the first aeration stage and 0 mLAir min−1 and 250 rpm in the second aeration stage, and changed the stage after 60 h of fermentation [17]. Thus, through this aeration strategy, Furlan et al. [17] obtained a Yethanol/substrate of 0.38 g g−1 and a rethanol of 0.09 g L−1 h−1. These authors obtained a slightly higher Yethanol/substrate than that of the DOT 50%—0 mLAir min−1 and 250 rpm experiment (Table 3), however the rethanol obtained by these authors was half of that obtained in the DOT 50%—0 mLAir min−1 and 250 rpm experiment (Table 3), which demonstrated the control of DOT at 50% in the first aeration stage allowed a considerable ethanol productivity improvement in a two-stage aeration strategy.
3.3. Two-Stage Aeration Experiment with 60% of HSSL
The potential of the two-stage aeration strategy in the fermentation of HSSL was assessed by performing an experiment using a medium containing 60% of HSSL and 40% of synthetic medium (v/v) (Figure 2). The experiment was performed by applying the aeration conditions that led to the most efficient bioethanol production in synthetic medium, i.e., DOT 50%—0 mLAir min−1 and 250 rpm. In this experiment, a maximum ethanol concentration of 12.2 g L−1 was obtained, corresponding to a rethanol of respectively 0.03 g L−1 h−1, and a Yethanol/substrate of 0.38 g g−1. These values were slightly lower than those obtained under the same conditions in synthetic medium. This probably resulted from the presence of microbial inhibitors in HSSL. In turn, a conversion efficiency of 74.4% was obtained, which was higher than the correspondent in synthetic medium, 64.1%. The results of the present study showed that the two-stage aeration strategy was more efficient in both growth stimulus and ethanol production when using a medium containing 60% of HSSL.
The effect of inhibitory compounds present in HSSL was visible in other features of the experiment. Contrary to what was observed in the experiment with synthetic medium, in which almost no lag phase was observed (Figure 1a), in this experiment with 60% HSSL (v/v) a lag phase of about 61 h was observed (Figure 2). Another consequence of the inhibitory effect was the lower value of µ of the experiment with 60% HSSL, around 6 times lower (Table 2). In fact, Xavier et al. verified the decreasing of the µ of S. stipitis with the increasing of HSSL percentage [27]. The inhibition of S. stipitis C4 growth by HSSL can be attributed to lignosulfonates, low molecular weight phenolic compounds, and acetic acid, which are the main inhibitors present in HSSL [6,27,28]. These compounds can inhibit growth by various mechanisms, while phenolic compounds are responsible for loss of cell membrane integrity [29] and for amino acid synthesis inhibition [30], acetic acid is responsible for cytoplasm acidification [29,31]. Furthermore, several authors have further shown that when combined, the toxicity of these inhibitors is higher than that resulting from the sum of the toxicities of each one [32,33,34]. In the experiment with 60% HSSL, along with glucose, acetic acid was also assimilated, which was completely consumed during the first aeration stage. Pereira et al. [11] also observed acetic acid consumption by S. stipitis C4 in a medium containing 60% HSSL (v/v). In the present work, the use of acetic acid as a carbon source resulted in the consumption of only 5.7% of the sugars during the first aeration stage, which corresponded to a third of the sugars consumed in the experiment in synthetic medium (Table 2). Thus, the use of acetic acid as a carbon source for S. stipitis C4 growth allowed a higher availability of sugars for ethanol production during the second aeration stage. In the experiment in synthetic medium the aeration stage change occurred when an optical density at 620 nm of 14 was reached, whereas in the experiment in 60% HSSL the aeration stage change occurred when an optical density at 620 nm of 20 was reached since HSSL has some turbidity associated. By other side a strong and enough biomass was needed in order to face 60% of HSSL and its inhibitors. Nonetheless, the amount of biomass at the end of the first aeration stage of the two experiments was similar 6.38 g L−1 in synthetic medium and 6.47 g L−1 with 60% HSSL (Table 2). As happened with synthetic medium, with 60% HSSL the change from the first aeration stage, with DOT control at 50%, to the second aeration stage, with 0 mLAir min−1 and 250 rpm, allowed for the induction of ethanol production and for the repression of S. stipitis C4 growth. Nevertheless, unlike the experiment in synthetic medium, in the end of experiment with 60% HSSL the ethanol production slowed down and still 12.0 g L−1 of xylose were remaining in the medium. These results demonstrate that besides inhibition of growth during the first stage there was also inhibition of ethanol production in the second stage. A maximum ethanol concentration of 12.2 g L−1 was obtained, corresponding to values of Yethanol/substrate and rethanol of 0.39 g g−1 and 0.04 g L−1 h−1, respectively (Table 2). Despite the similarity between the values of Yethanol/substrate, the rethanol obtained in the experiment with 60% HSSL was considerably lower than the value obtained in the corresponding experiment in synthetic medium (Table 2). These results showed that during the second aeration stage of the experiment with 60% HSSL, although no acetic acid was present, there were probably other inhibitors, namely lignosulfonates and low molecular weight phenolic compounds.
In the experiment with 60% HSSL during the second aeration stage, xylitol was produced reaching a maximum concentration of 2.66 g L−1 with a Yxylitol/xylose of 0.03 g g−1 (Table 2). The yield was 4.3 fold than in the assays using synthetic medium under the same conditions (Table 2). Mahler et al. showed that xylitol production by S. stipitis is associated with magnesium limitation in addition to oxygen limitation [35,36]. Mg2+ limitation inhibits the alcohol dehydrogenase (ADH), which leads to the increase of intracellular NADH, stimulating the xylitol accumulation [35]. In the experiment in synthetic medium with DOT 50%—0 mLAir min−1 and 250 rpm Mg2+ concentration at the aeration stage change was 26 mg L−1. In turn, in the experiment with 60% HSSL under the same conditions Mg2+ concentration was 3670 mg L−1. These results confirm the findings of Mahler et al. [35] and the relation between Mg2+ limitation and xylitol accumulation. HSSL coming from a magnesium-based acidic sulfite pulping process is a renewable raw material with a high Mg2+ concentration, which can be an advantage of using HSSL as feedstock since less xylitol is produced deviating less carbon from ethanol production.
Pereira et al. [11], applying a single-stage aeration strategy with 240 mLAir min−1 and 240 rpm and using, such as in the present study, S. stipitis C4, obtained a rethanol of 0.05 g L−1 h−1, and a Yethanol/substrate of 0.16 g g−1 in the fermentation of 60% HSSL (v/v). However, the higher rethanol obtained may be associated with a lower inhibitor content of the HSSL used which was previously detoxified with fungus Paecilomyces variotti [11]. Nonetheless, the Yethanol/substrate obtained by Pereira et al. [11] was 2.4 times lower than the Yethanol/substrate obtained in the present study for the two-stage aeration experiment with 60% HSSL. The lower Yethanol/substrate obtained by these authors resulted, probably, from excessive growth and ethanol reassimilation. Thus, the lower efficiency of S. stipitis C4 in the conversion of sugars to ethanol in the work developed by Pereira et al. [11] confirms the great potential of the two-stage aeration strategy in relation to the strategy with just a single aeration stage in the fermentation of HSSL.
4. Conclusions
The use of a two-stage aeration fermentation strategy in synthetic medium, with DOT controlled in the first stage and the oxygen restriction in the second stage, allowed for an efficient conversion of sugars to ethanol. In the first aeration stage, the increase of oxygen availability provided faster growth and higher biomass yield, which allowed the reduction in both stage duration and percentage of sugars consumed. In the second aeration stage, the oxygen restriction allowed to successfully induce the ethanol production. Furthermore, it was verified that rethanol of the second aeration stage increased with the increased oxygen availability in the first aeration stage. Moreover, the restricted oxygen availability imposed in the second aeration stage allowed not only minimized growth but also eliminated ethanol reassimilation. The two-stage aeration strategy was crucial for S. stipitis C4 growth in 60% HSSL, since the yeast tolerance to HSSL inhibitors was increased by the high oxygen availability of the first aeration stage. The two-stage aeration strategy, allowing growth stimulus and high ethanol yield, appears as a promising operational aeration alternative for the second-generation bioethanol production from HSSL. Future research should be conducted in order to evaluate the industrial integration of bioethanol production with this strategy of two aeration stages by S. stipitis C4 in order to ensure the viability of the second-generation bioethanol production from HSSL.
Supplementary Materials
The following are available online at https://www.mdpi.com/2311-5637/4/4/97/s1: Figure S1: Single-stage aeration experiments in synthetic medium: (a) DOT 1%; (b) DOT 2.5%; (c) DOT 10%; (d) DOT 25%; (e) DOT 50%; Figure S2: Fitting of Monod kinetic equation to experimental data of µmax (h−1) versus DOT (%) by non-linear regression.
Author Contributions
T.M.H. planned the study, performed the C4 batch reactor tests and wrote the manuscript. S.R.P. assisted the batch reactor tests. L.S.S. designed and supervised the experiments and revised the manuscript and A.M.R.B.X. conceived the original aeration idea, supervised experiments and revised the manuscript.
Funding
This work was developed within the scope of the project CICECO-Aveiro Institute of Materials, POCI-01-0145-FEDER-007679 (FCT Ref. UID/CTM/50011/2013), financed by national funds through the FCT/MEC and when appropriate co-financed by FEDER under the PT2020 Partner-ship Agreement.
Acknowledgments
Authors acknowledge Eng. A. Prates from CAIMA, Indústria de Celulose, ALTRI, Constância, Portugal for HSSL and Dmitry Evtyugin for technical advices.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IEA. Key World Energy Statistics; OECD IEA: Paris, France, 2018. [Google Scholar]
- Escobar, J.C.; Lora, E.S.; Venturini, O.J.; Yáñez, E.E.; Castillo, E.F.; Almazan, O. Biofuels: Environment, technology and food security. Renew. Sustain. Energy Rev. 2009, 13, 1275–1287. [Google Scholar] [CrossRef]
- Balat, M. Production of bioethanol from lignocellulosic materials via the biochemical pathway: A review. Energy Convers. Manag. 2011, 52, 858–875. [Google Scholar] [CrossRef]
- Pereira, S.R.; Portugal-Nunes, D.J.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Advances in ethanol production from hardwood spent sulphite liquors. Process Biochem. 2013, 48, 272–282. [Google Scholar] [CrossRef]
- Lawford, H.G.; Rousseau, J.D. Production of ethanol from pulp mill hardwood and softwood spent sulfite liquors by genetically engineered E. coli. Appl. Biochem. Biotechnol. 1993, 39–40, 667–685. [Google Scholar] [CrossRef]
- Marques, A.P.; Evtuguin, D.V.; Magina, S.; Amado, F.M.L.; Prates, A. Chemical composition of spent liquors from acidic magnesium-based sulphite pulping of Eucalyptus globulus. J. Wood Chem. Technol. 2009, 29, 322–336. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Suzuki, M. Phylogenetic analysis of ascomycete yeasts that form coenzyme q-9 and the proposal of the new genera Babjeviella, Meyerozyma, Millerozyma, Priceomyces, and Scheffersomyces. Mycoscience 2010, 51, 2–14. [Google Scholar] [CrossRef]
- Delgenes, J.P.; Moletta, R.; Navarro, J.M. The effect of aeration on d-xylose fermentation by Pachysolen tannophilus, Pichia stipitis, Kluyveromyces marxianus and Candida shehatae. Biotechnol. Lett. 1986, 8, 897–900. [Google Scholar] [CrossRef]
- Limtong, S.; Sumpradit, T.; Kitpreechavanich, V.; Tuntirungkij, M.; Seki, T.; Yoshida, T. Effect of acetic acid on growth and ethanol fermentation of xylose fermenting yeast and Saccharomyces cerevisiae. Kasetsart J. (Nat. Sci.) 2000, 34, 64–73. [Google Scholar]
- Pereira, S.R.; Ivanuša, T.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Biological treatment of eucalypt spent sulphite liquors: A way to boost the production of second generation bioethanol. Bioresour. Technol. 2012, 103, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Sànchez i Nogué, V.; Frazão, C.J.R.; Serafim, L.S.; Gorwa-Grauslund, M.F.; Xavier, A.M.R.B. Adaptation of scheffersomyces stipitis to hardwood spent sulfite liquor by evolutionary engineering. Biotechnol. Biofuels 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nigam, J.N. Ethanol production from hardwood spent sulfite liquor using an adapted strain of pichia stipitis. J. Industrial Microbiol. Biotechnol. 2001, 26, 145–150. [Google Scholar] [CrossRef]
- Delgenes, J.P.; Moletta, R.; Navarro, J.M. Fermentation of d-xylose, d-glucose, l-arabinose mixture by pichia stipitis: Effect of the oxygen transfer rate on fermentation performance. Biotechnol. Bioeng. 1989, 34, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C.; Teixeira, J.A. Fermentation medium and oxygen transfer conditions that maximize the xylose conversion to ethanol by Pichia stipitis. Renew. Energy 2012, 37, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Skoog, K.; Hahn-Hagerdal, B. Effect of oxygenation on xylose fermentation by Pichia stipitis. Appl. Environ. Microbiol. 1990, 56, 3389–3394. [Google Scholar] [PubMed]
- Skoog, K.; Hahn-Hagerdal, B.; Degn, H.; Jacobsen, J.P.; Jacobsen, H.S. Ethanol reassimilation and ethanol tolerance in Pichia stipitis cbs 6054 as studied by 13c nuclear magnetic resonance spectroscopy. Appl. Environ. Microbiol. 1992, 58, 2552–2558. [Google Scholar] [PubMed]
- Furlan, S.A.; Bouilloud, P.; De Castro, H.F. Influence of oxygen on ethanol and xylitol production by xylose fermenting yeasts. Process Biochem. 1994, 29, 657–662. [Google Scholar] [CrossRef]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C. The influence of initial xylose concentration, agitation, and aeration on ethanol production by Pichia stipitis from rice straw hemicellulosic hydrolysate. Appl. Biochem. Biotechnol. 2010, 162, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, M.; Tohma, T.; Itaya, T.; Fujii, M. Ethanol production from a mixture of glucose and xylose by co-culture of pichia stipitis and a respiratory-deficient mutant of Saccharormyces cerevisiae. J. Ferment. Bioeng. 1997, 83, 364–370. [Google Scholar] [CrossRef]
- Bellido, C.; González-Benito, G.; Coca, M.; Lucas, S.; García-Cubero, M.T. Influence of aeration on bioethanol production from ozonized wheat straw hydrolysates using Pichia stipitis. Bioresour. Technol. 2013, 133, 51–58. [Google Scholar] [CrossRef] [PubMed]
- du Preez, J.C.; van Driessel, B.; Prior, B.A. D-xylose fermentation by Candida shehatae and Pichia stipitis at low dissolved oxygen levels in fed-batch cultures. Biotechnol. Lett. 1989, 11, 131–136. [Google Scholar] [CrossRef]
- Wise, W.S. The measurement of the aeration of culture media. J. Gen. Microbiol. 1951, 5, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Schaepe, S.; Kuprijanov, A.; Sieblist, C.; Jenzsch, M.; Simutis, R.; Lübbert, A. kLa of stirred tank bioreactors revisited. J. Biotechnol. 2013, 168, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Laplace, J.M.; Delgenes, J.P.; Moletta, R.; Navarro, J.M. Alcoholic fermentation of glucose and xylose by Pichia stipitis, Candida shehatae, Saccharomyces cerevisiae and Zymomonas mobilis: Oxygen requirement as a key factor. Appl. Microbiol. Biotechnol. 1991, 36, 158–162. [Google Scholar] [CrossRef]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C.; Teixeira, J.A. Ethanol production from xylose by pichia stipitis nrrl y-7124 in a stirred tank bioreactor. Braz. J. Chem. Eng. 2011, 28, 151–156. [Google Scholar] [CrossRef]
- Su, Y.K.; Willis, L.B.; Jeffries, T.W. Effects of aeration on growth, ethanol and polyol accumulation by Spathaspora passalidarum nrrl y-27907 and Scheffersomyces stipitis nrrl y-7124. Biotechnol.Bioeng. 2015, 112, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.M.R.B.; Correia, M.F.; Pereira, S.R.; Evtuguin, D.V. Second-generation bioethanol from eucalypt sulphite spent liquor. Bioresour. Technol. 2010, 101, 2755–2761. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, D.L.A.; Silva, C.M.; Xavier, A.M.R.B.; Evtuguin, D.V. Fractionation of sulphite spent liquor for biochemical processing using ion exchange resins. J. Biotechnol. 2012, 162, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. Ii: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Ejim, L.; Mirza, I.A.; Capone, C.; Nazi, I.; Jenkins, S.; Chee, G.L.; Berghuis, A.M.; Wright, G.D. New phenolic inhibitors of yeast homoserine dehydrogenase. Bioorg. Med. Chem. 2004, 12, 3825–3830. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Modig, T.; Petersson, A.; Hähn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.F. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Díaz, M.J.; Ruiz, E.; Romero, I.; Cara, C.; Moya, M.; Castro, E. Inhibition of pichia stipitis fermentation of hydrolysates from olive tree cuttings. World J. Microbiol. Biotechnol. 2009, 25, 891–899. [Google Scholar] [CrossRef]
- Nigam, J.N. Ethanol production from wheat straw hemicellulose hydrolysate by Pichia stipitis. J. Biotechnol. 2001, 87, 17–27. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, J.; Zhu, Y.; Zhang, L.; Yong, Q.; Xu, Y.; Li, X.; Yu, S. Cause analysis of the effects of acid-catalyzed steam-exploded corn stover prehydrolyzate on ethanol fermentation by Pichia stipitis cbs 5776. Bioprocess Biosyst. Eng. 2014, 37, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Mahler, G.; Nudel, C. Effect of magnesium ions on fermentative and respirative functions in pichia stipitis under oxygen-restricted growth. Microbiol. Res. 2000, 155, 31–35. [Google Scholar] [CrossRef]
- Mahler, G.F.; Guebel, D.V. Influence of magnesium concentration on growth, ethanol and xylitol production by Pichia stipitis nrrl y-7124. Biotechnol. Lett. 1994, 16, 407–412. [Google Scholar] [CrossRef]
Figure 1.
Two-stage aeration experiments in synthetic medium: (a) Dissolved oxygen tension (DOT) 50%—0 mLAir min−1 and 250 rpm; (b) DOT 50%—50 mLAir min−1 and 150 rpm; (c) DOT 50%—DOT 1%. The dashed vertical line indicates the aeration stage change.
Figure 1.
Two-stage aeration experiments in synthetic medium: (a) Dissolved oxygen tension (DOT) 50%—0 mLAir min−1 and 250 rpm; (b) DOT 50%—50 mLAir min−1 and 150 rpm; (c) DOT 50%—DOT 1%. The dashed vertical line indicates the aeration stage change.

Figure 2.
Two-stage aeration experiment (DOT 50%—0 mLAir min−1 and 250 rpm) with 60% HSSL/40% synthetic medium—The dashed vertical line indicates the aeration stage change.
Figure 2.
Two-stage aeration experiment (DOT 50%—0 mLAir min−1 and 250 rpm) with 60% HSSL/40% synthetic medium—The dashed vertical line indicates the aeration stage change.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Aeration conditions of all experiments carried out in the present study.
| Fermentation Medium | Single-Stage Aeration Experiments | Two-Stage Aeration Experiments | |
|---|---|---|---|
| 1st Stage | 2nd Stage | ||
| Synthetic medium | DOT 1% | DOT 50% | 0 mLAir min−1 and 250 rpm |
| DOT 2.5% | DOT 50% | 50 mLAir min−1 and 150 rpm | |
| DOT 10% | DOT 50% | DOT 1% | |
| DOT 25% | |||
| DOT 50% | |||
| 60% HSSL/40% synthetic medium (v/v) | - | DOT 50% | 0 mLAir min−1 and 250 rpm |
Table 2.
Kinetic and stoichiometric parameters for two-stage aeration experiments.
| Medium | Synthetic Medium | 60% HSSL | ||||||
|---|---|---|---|---|---|---|---|---|
| Experiment | DOT 50%—0 mLAir min−1 and 250 rpm | DOT 50%—50 mLAir min−1 and 150 rpm | DOT 50%—DOT 1% | DOT 50%—0 mLAir min−1 and 250 rpm | ||||
| Stage | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd |
| µmax (h−1) | 0.43 | - | 0.44 | - | 0.43 | - | 0.07 | - |
| [Biomass]max (g L−1) | 6.38 | 6.58 | 5.36 | 6.06 | 5.57 | 10.2 | 6.47 | - |
| [Ethanol]max (g L−1) | - | 15.8 | - | 11.4 | - | 1.67 | - | 12.2 |
| [Xylitol]max (g L−1) | - | 5.10 | - | 3.72 | - | 0.698 | - | 2.66 |
| rglucose (g L−1 h−1) | 0.58 | - | 0.51 | - | 0.53 | - | 0.03 | - |
| rxylose (g L−1 h−1) | 0.31 | 0.52 | - | 0.72 | - | 0.42 | - | 0.11 |
| racetic acid (g L−1 h−1) | - | - | - | - | - | - | 0.08 | - |
| Consumed sugars (%) | 16.9 | 82.2 | 10.4 | 84.2 | 12.6 | 86.4 | 5.7 | 66.8 |
| rethanol (g L−1 h−1) | - | 0.21 | - | 0.21 | - | 0.09 | - | 0.04 |
| rxylitol (g L−1 h−1) | - | 0.068 | - | 0.073 | - | 0.038 | - | 0.004 |
| Ybiomass/substrate (g g−1) | 0.78 | - | 1.13 | 0.02 | 0.93 | - | 0.65 | - |
| Yethanol/substrate (g g−1) | - | 0.40 | - | 0.29 | - | 0.06 | - | 0.39 |
| Yxylitol/xylose (g g−1) | - | 0.13 | - | 0.10 | - | 0.03 | - | 0.03 |
Table 3.
Global kinetic and stoichiometric parameters of the two-stage aeration experiments in synthetic medium.
Table 3.
Global kinetic and stoichiometric parameters of the two-stage aeration experiments in synthetic medium.
| Experiment | DOT 50%—0 mLAir min−1 and 250 rpm | DOT 50%—50 mLAir min−1 and 150 rpm | DOT 50%—DOT 1% |
|---|---|---|---|
| [Ethanol]max (g L−1) | 15.8 | 11.4 | 1.67 |
| rethanol (g L−1 h−1) | 0.18 | 0.18 | 0.04 |
| Yethanol/substrate (g g−1) | 0.33 | 0.25 | 0.04 |
| Conversion Efficiency (%) | 64.1 | 49.4 | 7.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Henriques, T.M.; Pereira, S.R.; Serafim, L.S.; Xavier, A.M.R.B. Two-Stage Aeration Fermentation Strategy to Improve Bioethanol Production by Scheffersomyces stipitis. Fermentation 2018, 4, 97. https://doi.org/10.3390/fermentation4040097
AMA Style
Henriques TM, Pereira SR, Serafim LS, Xavier AMRB. Two-Stage Aeration Fermentation Strategy to Improve Bioethanol Production by Scheffersomyces stipitis. Fermentation. 2018; 4(4):97. https://doi.org/10.3390/fermentation4040097
Chicago/Turabian StyleHenriques, Tiago M., Susana R. Pereira, Luísa S. Serafim, and Ana M. R. B. Xavier. 2018. "Two-Stage Aeration Fermentation Strategy to Improve Bioethanol Production by Scheffersomyces stipitis" Fermentation 4, no. 4: 97. https://doi.org/10.3390/fermentation4040097
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.