
Co-Production of Isobutanol and Ethanol from Prairie Grain Starch Using Engineered Saccharomyces cerevisiae

Department of Biochemistry, University of Alberta, Edmonton, AB 26G 2H7, Canada
*
Author to whom correspondence should be addressed.
Fermentation 2021, 7(3), 150; https://doi.org/10.3390/fermentation7030150
Submission received: 9 July 2021
/
Revised: 5 August 2021
/
Accepted: 9 August 2021
/
Published: 13 August 2021
(This article belongs to the Section Industrial Fermentation)
Abstract
:Isobutanol is an important and valuable platform chemical and an appealing biofuel that is compatible with contemporary combustion engines and existing fuel distribution infrastructure. The present study aimed to compare the potential of triticale, wheat and barley starch as feedstock for isobutanol production using an engineered strain of Saccharomyces cerevisiae. A simultaneous saccharification and fermentation (SSF) approach showed that all three starches were viable feedstock for co-production of isobutanol and ethanol and could produce titres similar to that produced using purified sugar as feedstock. A fed-batch process using triticale starch yielded 0.006 g isobutanol and 0.28 g ethanol/g starch. Additionally, it is demonstrated that Fusarium graminearum infected grain starch contaminated with mycotoxin can be used as an effective feedstock for isobutanol and ethanol co-production. These findings demonstrate the potential for triticale as a purpose grown energy crop and show that mycotoxin-contaminated grain starch can be used as feedstock for isobutanol biosynthesis, thus adding value to a grain that would otherwise be of limited use.
1. Introduction
A growing desire for energy security coupled with concerns over climate change have driven interest in the production of fuels and chemicals from renewable feedstock. Ethanol is currently the most widely used biofuel and all indications are that demand for it will continue to grow [1]. Despite its advantages and relative ease of production ethanol has some limitations in the degree to which it can be blended with petroleum fuels. Additionally, its miscibility with water means that it is not fully compatible with existing fuel storage and distribution infrastructure. Other candidate biofuels include butanol and its isomer, isobutanol. These four carbon alcohols possess greater energy density than ethanol and are immiscible with water [2]. Butanol and isobutanol are compatible with existing fuel distribution infrastructure and they can be used directly in contemporary combustion engines or blended with gasoline in any proportion [3]. Most butanol and isobutanol are currently synthesized from petrochemicals, however, Clostridium acetobutylicum will naturally produce 1-butanol and Saccharomyces cerevisiae, bacteria and cyanobacteria have been engineered to produce these alcohols [4,5,6,7,8]. Isobutanol production by E. coli and S. cerevisiae has used glucose as feedstock but microbial strains have been engineered to achieve isobutanol synthesis from xylose, cellulose, methanol, cheese whey and acetate [9,10,11,12,13].
Isobutanol has value both for use as a platform chemical and as a candidate biofuel. Cradle-to-grave life cycle analysis demonstrates that isobutanol produced by microbial conversion of sugar yields a 47% reduction in greenhouse gas emissions (GHG) and a 26% reduction in fossil energy consumption (FEC) when compared with isobutanol synthesized by petrochemical processes [14]. These considerations, which include reduced process energy, could make the bio-based product attractive economically, particularly if produced within the context of a biorefinery where co-products could add value to the process [15].
In North America, corn is the most extensively used feedstock for ethanol production and an industrial process has been developed using corn as feedstock for isobutanol production [16,17]. As the biofuel industry expands there will be an increased need for other low-cost feedstock to meet the rising product demand. Cereal grains such as wheat and barley have high starch content and have also been used for ethanol production [18]. An important economic consideration is that cereal grains can grow robustly in environments where corn does not owing to reduced time to maturity, temperature tolerance and reduced water requirements [19,20]. Barley (Hordeum vulgare) has a long history as a feedstock for ethanol production and has a well-developed industry supporting its processing and use in the production of malt beverages as well as fuel ethanol [21]. Wheat has also been explored as a feedstock for bioethanol production [22]. Triticale (triticosecale) is a hybrid grain produced by a cross between wheat and rye. This grain has a similar starch content to wheat and has been found to be suitable for bioethanol production [23]. Triticale-derived starch is particularly attractive as a purpose grown feedstock since it is not used extensively as a human food crop and has a greater environmental range than either wheat or barley [19].
Starch is a polymer not naturally fermented by S. cerevisiae as the organism does not secrete enzymes that can cleave the α1-4 glycosidic bonds between glucose moieties. Liquefaction and hydrolysis of starch to glucose can be achieved either through high temperature jet cooking or through an enzymatic process employing raw starch hydrolyzing enzymes. Jet cooking is effective but has some drawbacks in cost associated with the energy required for the process [24]. An enzymatic process, simultaneous saccharification and fermentation (SSF), combines starch, raw starch enzymes, and a microorganism to simultaneously convert biomass to digestible sugars and the desired end product [25,26]. SSF of biomass feedstock is an approach with numerous potential advantages. (1) It allows the derivation of glucose from starch feedstock with less energy than required when the starch is subjected to high temperature hydrolysis. (2) The process avoids osmotic shock to the fermenting organism caused by high initial glucose concentrations. (3) It limits the potential for microbial contamination since the inoculated yeast rapidly metabolizes the available glucose. (4) It maintains the equilibrium of enzymatic reactions in favor of starch degradation and end product formation, thus avoiding inhibition of the enzymes by the product glucose. SSF has been applied to ethanol fermentation from starch, and cellulosic feedstock and can be applied to fermentative production of other products [27]. Additionally, a fed-batch SSF approach has been applied to increase titre and yield in ethanol fermentations [28,29]. In this study we investigated the potential of employing SSF and an engineered strain of S. cerevisae to co-produce isobutanol and ethanol from grain starch. We compared the effectiveness of triticale, wheat and barley starch as feedstock. This procedure was successful with starch sourced from a variety of prairie grains and with starch derived from grain contaminated with Fusarium graminearum mycotoxins indicating that this process might be a route to add value to an otherwise low value material.
2. Materials and Methods
2.1. Yeast Strain
The Saccharomyces cerevisiae strain used in this study, DSYB60 was constructed in the W303 genetic background [30]. The strain was derived from DSYB37 [7]. This strain overexpresses the endogenous ILV2, ILV3, ILV5 genes, a feedback resistant variant of ILV6 (ILV6fr), the ketoacid decarboxylase gene kdcA from lactococcus lactis regulated by a PGK1 promoter and modified to encode a mitochondrial import signal peptide, an E. coli yqhD gene that was modified by addition of a mitochondrial import signal peptide. These genes were integrated as a single array at the LEU2 locus. An extra copy of the NADH kinase POS5 was integrated at the TRP1 locus. Complete details of the gene array used were previously described [7]. An extra copy of the ILV2 gene regulated by an FBA1 promoter was integrated at the HIS3 locus. The strain was further modified by transformation with DNA fragments encoding functional ADE2, and URA3 genes to generate a prototrophic production strain. Yeast transformation was achieved using the lithium acetate/polyethylene glycol procedure [31]. Transformants were isolated from agar plates prepared with the appropriate selective medium [32].
2.2. Grain Samples
The grains used in this study were developed by Agriculture Canada and have the designation AC. The AC Ultima Triticale, AC Metcalfe Barley and AC Sadash Wheat are all commercially available grain varieties. All were from grain stocks grown in 2011. Samples of commercially available Excel barley were obtained from the Agriculture Canada Brandon Research and Development Centre. These samples were obtained from a controlled experiment comparing Excel barley growth and productivity when experimentally contaminated with F. graminerium in a controlled environment. The uninfected control barley cultures were used as the “clean” barley sample. The grains were milled using a Retsch mill (model ZM 100, Retsch Gmbh and Co., Haan, Germany) with a 0.5 mm sieve. The resulting starches were stored in air-tight plastic bags at 4 °C until used.
2.3. Starch Hydrolysis Enzymes
All enzymes used in this study are commercially available and were produced by and obtained from Genecor International (Palo Alto, CA, USA) and were generously shared with us by Dr. David Bressler, University of Alberta. OPTIMASH™ TBG is a thermostable β-glucanase added to hydrolyse β-glucan polymers and reduce viscosity of mash preparations. FERMGEN is a fungal acid protease derived from a modified strain of Trichoderma reesei. The protease treatment aids with the release of starch as well as providing amino acids and peptides to support yeast growth. GC 626 is an acid stable α-amylase derived from a genetically modified strain of Trichoderma reesei. Stargen 002 is a cocktail of Aspergillus kawachi α-amylase and a glucoamylase from Trichoderma reesei that hydrolyze starch into glucose monomers.
2.4. Starch Feedstock Preparation
A mash from each starch sample was prepared as a 35% solids mixture with water. The mash was heated to 55 °C and the pH was adjusted to 4.0 by addition of 2 M HCl. Each mash was treated with an enzyme cocktail consisting of OPTIMASH™ TBG (160 µL/kg starch) Fermgen (1880 µL/kg starch), and GC626 (120 µL/kg starch). The mash was incubated at 55 °C for 1 h with gentle agitation. The samples were sterilized by the addition of diethylpyrocarbonate to 0.025%. After overnight incubation at 4 °C, the mashes were heated to 55 °C and Stargen 002 was added to 2.8 mL/kg starch. Subsequently the mashes were supplemented with urea and diluted to 10 mM of urea and 8% starch prior to transfer to sterile fermentation flasks. In experiments to investigate nitrogen source, the mashes were supplemented with urea, ammonium sulfate or ammonium chloride.
2.5. Fermentation Reactions
Fermentation trials were conducted in 500 mL shake flasks. The 120 mL mashes were inoculated with 1.2 × 106 CFU/mL of the yeast DSYB60 and incubated at 30 °C with mild agitation (140 rpm). Samples of the fermentation broth were collected at 0, 24, 48, and 72 h for analysis of glucose concentration, xylose concentration, cell number, isobutanol concentration, ethanol concentration and in some cases Deoxynivalenol (DON) concentration. Fed-batch fermentation trials were similarly performed in 500 mL shake flasks with an initial volume of 120 mL of starch mash. After 48 h of fermentation the fed-batch culture was supplemented with an additional 1.6 g of starch and fermentation was allowed to proceed for an additional 24 h before terminating the reactions and measuring the final products.
2.6. Mycological Analysis
The presence of fungal contamination of grain samples was determined by a colony counting procedure based on an established protocol [33]. We milled 10 g of each grain sample and added it to 90 mL of sterile 0.1% peptone. The mixture was incubated with mild agitation (100 rpm) on a rotary shaker for 10 min. A series of 10-fold dilutions of each sample were prepared and 100 µL aliquots of each dilution were plated in triplicate on YEPD (10 g/L yeast extract, 20 g/L bactopeptone, 10 mg/L adenine hemisulfate, 10 mg/L tryptophan, 20 g/L glucose, 15 g/L agar) agar plates supplemented with 100 mg/L chloramphenicol. Plates were incubated at 30 °C in the dark for 6 days and then colonies were counted. Only plates displaying between 5 and 150 colonies were included in the analysis.
2.7. Analytical Methods
Glucose and xylose concentrations were determined during the fermentation time-courses using a glucose oxidase/peroxidase assay and xylose mutarotase/β-xylose dehydrogenase assay, respectively. Both assays were performed as directed by the manufacturer (Megazyme Ltd, Lansing, MI, USA). Isobutanol and ethanol concentrations were determined by gas chromatography (GC) as previously described [7] with slight modifications. 1-pentanol was used as an internal standard with an Agilent Technologies (Santa Clara, CA, USA) 7890A GC System and 7693 Autosampler using a Restek Stabilwax-DA column (30 m by 0.53 mm inner diameter, 0.5-μm film thickness), a 1 μL injection in split mode (20:1 split ratio), an injector temperature of 170 °C, an FID temperature of 190 °C, and helium carrier gas in constant pressure mode (7.5 lb/in2). The oven program started at 35 °C, held for 3 min, followed by 20 °C/min to 190 °C with a final hold of 1 min. An aliquot of 800 μL of the supernatant sample and 200 μL of a 0.5% 1-pentanol internal standard solution were added to 1.5 mL GC vials and thoroughly mixed. For standards, 200 μL of pure isobutanol, and 200 µL ethanol and 1-pentanol (internal standard) were used. Water was used as blank. The GC response factors were determined and used with an internal standard correction to measure the concentration of the alcohol in the samples. A two-tailed t-test was applied to the data to determine whether differences in product concentrations were statistically significant. A p value < 0.05 was considered to reflect a significant difference. Deoxynivalenol concentrations were determined using a colorimetric competitive–ELISA technique in a 96-well format (E-TO-E011, Elabscience Biotechnology Inc., Houston, Texas, USA). Sample quantitation was achieved by comparison of diluted samples with a series of analytical standards ranging from 0 ng/Ml–810 ng/mL. Colour development of standards and samples was measured at an optical density of 450 nm using an automated multi-plate reader (Spectramax M3, Molecular Devices, San Jose, CA, USA).
Cell viability and density were determined on culture samples taken at 0, 24, 48, and 72 h time points by performing serial 10-fold dilutions and plating samples from each time point on to YEPD agar plates in triplicate. Agar plates were incubated at 30 °C and 3 days after plating the colonies were counted.
3. Results
3.1. SSF Treatment of Grain Starch
We investigated the potential of the prairie grains triticale, barley and wheat as feedstock for the production of isobutanol. The initial treatment of the starch mashes yielded available glucose with slightly more being liberated from the triticale sample 148 ± 6.4 mg/100 mL than from the wheat 107 ± 14.2 mg/100 mL or barley 129.65 ± 8.4 mg/100 mL, despite the same amounts of starch in the mash and identical enzymatic treatments (Table 1). The differences between glucose released from triticale and wheat was statistically significant p = 0.01, as was the difference between triticale and barley p = 0.029. These differences may result from variations in the content of arabinoxylan and β-glucan in the grain samples however we did not measure these directly. The concentration of xylose in the mash was relatively low, and this reflects the fact that the enzyme preparations used were not enriched in xylanases. The initially accumulated glucose was depleted from all fermentation reactions within 24 h and was maintained at a stable low concentration of 3–7 mg/100 mL over the subsequent fermentation period (Table 1). In contrast to the glucose, the pentose sugars were not depleted during the fermentation (Table 1). This was expected since the strain used to perform the fermentation has limited ability to ferment pentose sugars.
Over the fermentation time-course the viable cell biomass as measured by the number of colony forming units (CFU/mL) increased to a maximum at 72 h (Figure 1A). The cell numbers increased at similar rates in the triticale, wheat and barley fermentations with no significant difference between the cultures at 72 h. Sample analysis was performed in triplicate but some variation was observed in the sampling, possibly due to the presence of solids in the fermentation reactions. It is notable that only a minor increase in cell numbers occurred between 24 and 72 h suggesting that some nutrients required for growth may have become limiting by that time.
3.2. Isobutanol and Ethanol Production from Starch
Isobutanol was detected in the cultures within 24 h. At this early time point the isobutanol produced in the triticale fermentation (248.65 ± 4.92 mg/L) was significantly greater than in wheat (115.89 ± 13.34 mg/L) p < 0.001, or barley (156.12 ± 27.21 mg/L) p = 0.001 (Figure 1B). Isobutanol continued to accumulate and by 48 h the isobutanol produced in triticale (401.4 ±29.8 mg/L) remained significantly greater than that produced in wheat (288.1 ± 17.5 mg/L) p < 0.005 or barley (310.81 ± 28.1 mg/L) (Figure 1B). The greater concentration of isobutanol produced in triticale was consistent with increased glucose availability (Figure 1B). There was no significant difference observed between the concentrations of isobutanol achieved in the triticale (368.73 ± 9.08 mg/L), wheat (323.07 ± 53.29 mg/L) or barley (331.67 ± 41.8 mg/L) mashes at 72 h (Figure 1B). There was no increase in isobutanol accumulation in triticale at 72 h but isobutanol did continue to accumulate in the wheat and barley cultures so that it approached the isobutanol concentration achieved in triticale. This lack of further product accumulation in the triticale past 48 h is consistent with depletion of the available carbon and may also reflect evaporative loss of the product as the fermentation vessels were not sealed. The mashes all had some remaining glucose so carbon depletion may not have been the only limiting factor. Isobutanol is toxic to yeast and could in principle limit cell growth and further production of isobutanol. However, S. cerevisiae growth is not significantly impacted at concentrations of isobutanol less than 10 g/L [34]. The concentrations achieved here (~400 mg/L) are below the level expected to kill the cells.
S. cerevisiae is a natural ethanol producer and we observed that ethanol accumulated in the fermentation reactions with our engineered strain. Ethanol was detectable at 24 h and its abundance mirrored the accumulation of isobutanol and available glucose. Fermentations with triticale displayed a titre of 15.01 ± 0.2 g/L ethanol, which was significantly greater than wheat 11.68 ± 0.08 g/L or barley 13.1 ± 0.09 g/L at 48 h (Figure 1B). The accumulation of ethanol continued through 48 and 72 h and as was the case for isobutanol peak ethanol accumulation was achieved at 48 h in the triticale fermentation 20.52 ± 0.25 g/L and 72 h for wheat and barley 19.12 ± 0.15 g/L and 20.07 ± 0.22 g/L respectively (Figure 1B).
3.3. Isobutanol and Ethanol Production Using SSF in a Fed-Batch Fermentation
We tested the possibility of using a fed-batch strategy to increase isobutanol production from starch. This strategy has been successfully applied to ethanol fermentation from raw starch [29]. A mash was prepared from triticale starch using our standard protocol and 120 mL aliquots were transferred to sterile fermentation vessels prior to inoculation with yeast DSYB60 at a density of 1.6 × 106 CFU/mL to initiate the fermentation. At 48 h an aliquot of the original mash corresponding to 1.6 g of starch was spiked into the 120 mL fermentation reaction to determine if the increased starch feedstock would increase isobutanol production. Isobutanol accumulation could be detected at 24 h and titres approached 400 mg/L by 48 h consistent with previous batch fermentation trials (Figure 2B). No further isobutanol accumulation could be detected in the batch fermentation vessels with a final concentration of 395 ± 17.4 mg/L isobutanol and 21.63 ± 2.8 g/L ethanol corresponding to yields of 0.005 g/g starch for isobutanol and 0.27 g/g starch for ethanol. In contrast, the addition of 1.6 g of starch resulted in a 14.8% increase in isobutanol titre from 48 h to 72 h. A final concentration of (552.72 ± 68.91 mg/L) isobutanol and 26.31 ± 2.5 g/L ethanol was achieved (Figure 2B). These increases in product titre correspond to an improvement in isobutanol yield to 0.006 g/g starch and ethanol yield 0.28 g/g starch. The biomass yield based on CFU did not display a fully proportional increase (2.7 × 107 compared to 2.1 × 107 CFU/mL) implying that some condition in the fermentation was limiting or inhibitory for biomass accumulation or that the combination of accumulating isobutanol and ethanol inhibited growth.
3.4. Influence of the Nitrogen Source
Our initial fermentation trials employed urea as a source of nitrogen to promote cell growth. We wished to investigate the use of other nitrogen sources to determine if these would significantly influence the yield and titre of isobutanol and ethanol products of starch fermentation. We compared urea with ammonium sulfate and ammonium chloride at equimolar concentrations of nitrogen in a triticale starch fermentation. These are all considered to be good sources of assimilable nitrogen for S. cerevisiae [35]. To remain consistent with previous experiments a starch mash was prepared and diluted to 80 g/L solids prior to addition of the nitrogen source and inoculation with the engineered yeast strain. After 48 h of fermentation, the titres of isobutanol and ethanol in all cultures with added nitrogen were significantly greater (p < 0.001) than in cultures with no added nitrogen source (Table 2). A comparison of the effects of added urea, (NH4)2SO4, or NH4Cl demonstrated that isobutanol titres were similar at nearly 400 mg/L and there was no statistically significant difference between the tested conditions (Table 2). Final ethanol titres were all near 19 g/L and there was no significant difference between added sources of nitrogen. Increasing the concentration of nitrogen in the mashes did not yield any increase in product alcohol accumulation indicating that nitrogen availability was not limiting in the process (Table 2). Based on these observations, any of the tested nitrogen sources would be effective.
3.5. Fermentation of Mycotoxin Contaminated Grains
Starch is an effective feedstock for production of ethanol and isobutanol but is also a food crop creating concern around the use of food for producing fuel and chemicals. However, this strategy has the potential to be applied to starch from grains or other sources that have been damaged or are otherwise undesirable for human or animal consumption. Fusarium graminearum is a fungus that infects grain crops causing Fusarium Head Blight (FHB) and leading to significant economic losses [36]. In addition to the damage done to the grain and reduction in starch yields the fungus secretes deoxynivalenol (DON), zearalenone and other trichothecene microtoxins that cause vomiting and make the grain unsuitable for human or animal consumption [37,38]. We investigated the possibility that F. graminearum damaged barley could be used as a feedstock for isobutanol biosynthesis.
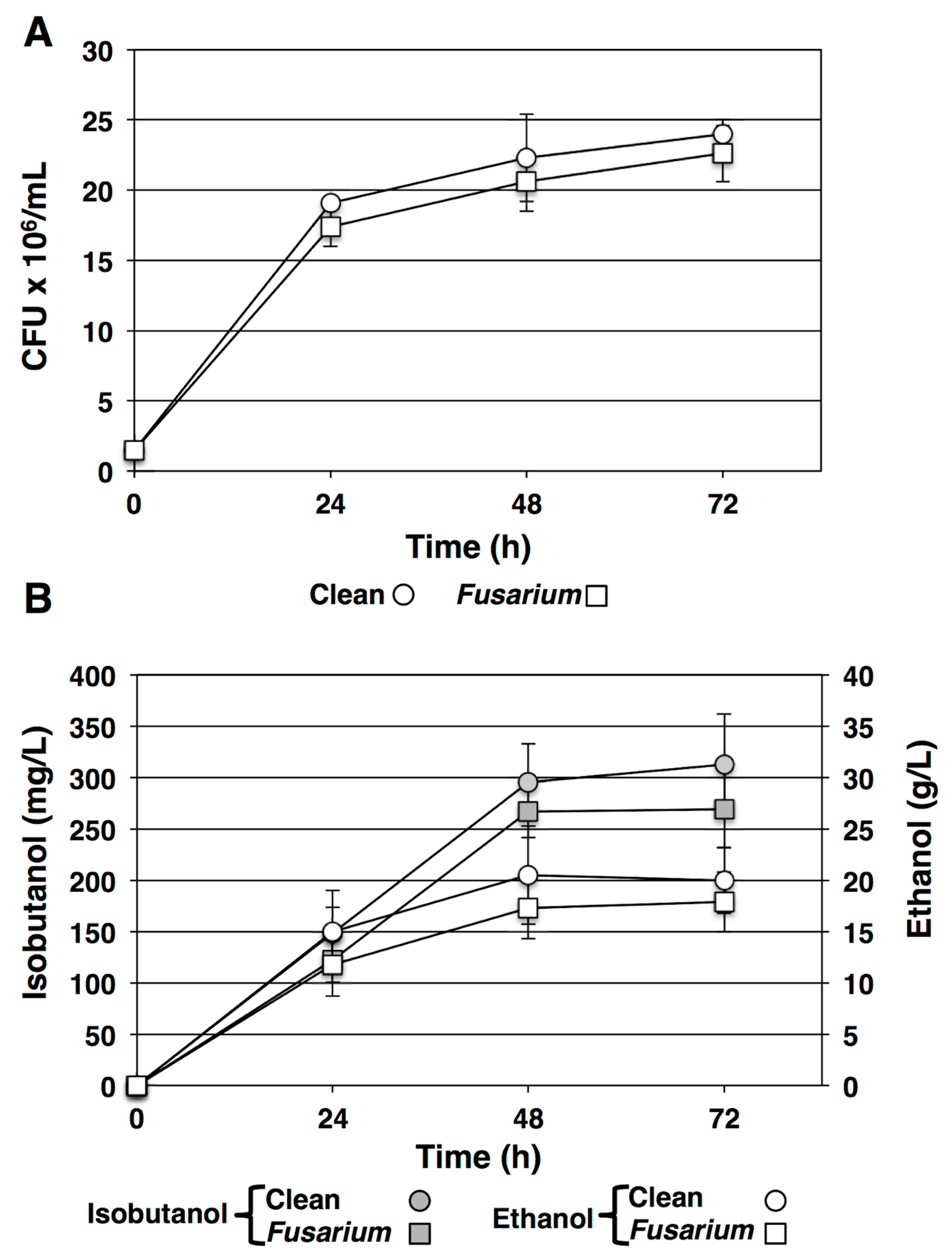
Starch from milled uninfected barley (clean) or F. graminearum infected barley (Fusarium) was prepared and subjected to mycological analysis. The clean barley sample presented 676.70 ± 12.2 CFU/g of grain and the F. graminearum infected sample presented 82,180.33 ± 901.1 CFU/g of grain. Starch samples of clean and F. graminearum infected grains were subjected to the SSF preparation protocol prior to initiating fermentation. The mash prepared from fungus-infected grain was contaminated with deoxynivalenol at a concentration greater than would be recommended for human or animal consumption (150.4 ± 12.5 µg/100 mL) and is substantially greater than the “clean” barley sample (0.08 ± 0.01 µg/100 mL) (Table 3). The glucose yield from the F. graminearum contaminated grain was lower than from the clean grain but the difference was not statistically significant (Table 3). Following inoculation of the mashes with engineered S. cerevisiae DSYB60 the cell biomass accumulated in a near parallel fashion in both the clean and the deoxynivalenol contaminated samples (Figure 3A). This is consistent with reports that S. cerevisiae is resistant to greater than 150 mg/L deoxynivalenol [39,40]. Analysis of the fermentation reactions by gas chromatography revealed that isobutanol was synthesized from both clean (312.72 ± 48.9 mg/L) and contaminated (269.24 ± 37.4 mg/L) feedstock and there was no significant difference between titre or yield of isobutanol produced (Figure 3B). Ethanol was also produced in the fermentation reactions and similar to the case with isobutanol there was no significant difference in the titre or yield of ethanol produced from deoxynivalenol contaminated grain (17.5 ± 3.1 g/L) relative to the clean grain samples (20.5 ± 4.6 g/L) (Figure 3B). Assay of the fermentation broth at 72 h showed a small but not statistically significant reduction in DON concentration (137.3 ± 13.2) consistent with previous observations that S. cerevisiae has a limited ability to metabolize or otherwise detoxify DON [41].
4. Discussion
The results of this study demonstrate that isobutanol can be produced from grain starch using an SSF procedure and that a fed-batch protocol can potentially increase both yield and titre of the products. The yields of isobutanol achieved with batch SSF of triticale starch, 401 mg/L, (8.35 mg/L/h) were comparable to the yields obtained from glucose when the cells are cultured in minimal medium [7]. However, the conversion rate achieved here was only 0.005 g/g starch based on the weight of input feedstock and the theoretical maximum yield of isobutanol from glucose is 0.41 g/g indicating that a substantial amount of the assimilated carbon was not effectively directed toward isobutanol production. Clearly a substantial amount of the input carbon was directed toward the synthesis of ethanol and the batch fermentation process of triticale yielded 0.29 g isobutanol + ethanol/g starch. Higher product titres can be achieved through a fed-batch process and it is likely that higher gravity starting cultures may produce higher titres. These data demonstrate the feasibility of a process for co-producing isobutanol and ethanol from prairie grain starch. Ethanol fermentation is already a cost-effective process and the addition of a higher value co-product can potentially improve the economics further. A benefit of using a co-production strategy is the potential to create a series of strains able to produce varying amounts of each product so that the process could be adapted to market demand for each. Yields of each product can be improved by both strain engineering and by increased product tolerance. Greater yields may also be possible when the yeast is engineered to effectively assimilate pentose sugars, which make up 1–2% of the sugars in the present in the mash. Some progress has been made on improving the conversion of xylose to isobutanol in S. cerevisiae but to date yields remain low and there has been no effective demonstration of the conversion of pentose sugars from a mixed carbon feedstock [9,42,43]. This may also open the possibility of using cellulosic and hemicellulosic materials as feedstock.
Isobutanol is naturally produced by S. cerevisiae in very small quantities mostly by deamination of branched chain amino acids [44]. Most efforts to improve isobutanol production in this organism have focused on increasing flux through the branched chain amino acid biosynthesis pathway [45,46]. The strain used in this study has been engineered to overproduce enzymes in the valine biosynthesis pathway and overproduce a heterologous ketoacid decarboxylase and alcohol dehydrogenase. Similar strategies by others have resulted in similar isobutanol yields 0.004–0.016 g/g glucose [8,47,48]. Higher titres and yields of isobutanol (1.6 g/L, 0.016 g/g glucose and 8.49 g/L, 0.053 g/g glucose) have been reported in strains engineered to increase isobutanol synthesis at the expense of ethanol [45,49].
S. cerevisiae is a natural ethanol producer and disruption of the ethanol production process is inhibitory to its growth when using glucose as a carbon source [50]. Efforts have been made to divert carbon flux away from ethanol and toward isobutanol. Deletion of genes encoding pyruvate decarboxylases PDC1 PDC5 in an isobutanol producing strain reduced both isobutanol and ethanol biosynthesis while deletion of either PDC1 or PDC5 individually resulted in little decrease in ethanol production or increase in isobutanol production [7,46]. Deletion of the ADH1 gene encoding the major alcohol dehydrogenase reduced ethanol production but triggered a compensatory increase in glycerol synthesis yielding little increase in isobutanol [46].
Some success has been achieved by deletions of adh1 gat1 to limit both ethanol and glycerol synthesis resulting in isobutanol titres of 560 mg/L with yield of 0.013 g/g glucose [46]. Zhao et.al. 2020 reported production of titres of 8.49 g/L (yield 0.053 g/g) of isobutanol from glucose through the use of an optogenic strategy to regulate PDC1 expression [49]. This approach appeared effective but required some special apparatus that might limit the scale of its application. Strains lacking PDC1, PDC5 and PDC6 function that can grow on glucose without producing ethanol have been generated with a compensatory mutation in the MTH1 gene [50]. An isobutanol-producing strain lacking the PDC1, PDC5 and PDC6 has been independently isolated through adaptive laboratory evolution but the compensatory mutations allowing that strain to effectively utilize glucose as a carbon source have not been reported [51]. It is likely that a fully optimized isobutanol producing strain would achieve higher product yields from grain starch making prairie grains an economically viable feedstock for the process.
In this study triticale, wheat and barley proved to be effective as feedstock for the co-production of isobutanol and ethanol and give similar yields of the final products. Triticale may hold an advantage as feedstock for biofuel or chemical production, it is high yielding, hardy, drought tolerant and capable of growth on marginal lands that will not support wheat or barley [52]. Triticale also requires less nitrogen fertilizer than wheat [53]. This is an important consideration in both cost of production and the fact that nitrogen fertilizer accounts for 70% of the carbon footprint of grain production [53,54]. Triticale is not extensively used as a human food, rather its largest market is for use as animal feed [55]. Thus, triticale may be an excellent candidate feedstock for biofuel or chemical production.
Many cereal grain crops including wheat, barley and triticale are susceptible to F. graminearum and other Fusarium strains that damage the grain starch and secrete DON, zearalenone and other mycotoxins [37]. The advent of global warming has allowed an expansion of the range for these fungal diseases with a subsequent increase in economic losses in grain production [56]. FDA guidance on DON contamination of wheat products for human and animal consumption recommend contamination of 10 mg/L or less [57]. Low-value grain damaged by fungal infection or other pathogens is an economic loss for grain producers. Development of a process that would allow for fuel or chemical production from this material would provide economic benefits and would fit within the parameters of existing biorefineries that utilize grain to generate flour and ethanol as well as co-products [58,59]. Additionally, it is important that any process for fuel or chemical production from a purpose grown feedstock be tolerant to environmental influences affecting that feedstock.
From the perspective of process optimization a significant cost component for starch fermentation remains the enzymes that are required for conversion of polymer starch into fermentable sugars. While the SSF process reduces enzyme cost a yeast strain that could synthesize and secrete amylases could in principle replace the exogenously added enzymes for a further reduction in cost. The application of amylase secreting yeasts for ethanol production from starch has been attempted but to date that has not been successfully implemented [60]. Product toxicity also remains a challenge for isobutanol and butanol synthesis as these alcohols are more toxic to microbial cells than ethanol. A variety of strategies for in situ product removal have been tested and a low-pressure evaporation process has been demonstrated commercially [17,61]. These processes can be applied to the recovery of both ethanol and isobutanol from a co-production process.
5. Conclusions
Engineered S. cerevisiae can effectively produce isobutanol from triticale, wheat and barley starches that have been subjected to enzymatic hydrolysis with raw starch enzymes. The yields of isobutanol produced from starch are similar to the yields produced from purified sugar. The application of a fed-batch strategy may be effective in increasing titre and yield of product isobutanol. Additionally, it is demonstrated that mycotoxin-contaminated grain serves as an effective feedstock for isobutanol and ethanol co-production.
Author Contributions
Conceptualization, D.T.S.; methodology, D.T.S., E.U., X.L.; validation, X.L., E.U. and D.T.S.; formal analysis, D.T.S.; investigation, X.L., E.U.; resources, D.T.S.; writing—original draft preparation, review and editing, D.T.S.; visualization, X.L.; supervision, D.T.S.; project administration, D.T.S.; funding acquisition, D.T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by operating grants from The Network Centre of Excellence BioFuelNET and Alberta Innovates BioSolutions through the Biorefining Conversions Network.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank David Bressler and Archana Paraschar (University of Alberta) for supplying enzymes and starch samples, Ana Badea and James Tucker (Barley Breeding and Genetics, Brandon Research and Development Centre, Agriculture and Agri-Food Canada) for providing samples of barley grain and F. graminearum contaminated barley grain and Jingui Lan and the University of Alberta Faculty of Medicine and Dentistry Lipidomics Core, RRID:SCR_019176, which receives financial support from the Faculty of Medicine & Dentistry, Canada Foundation for Innovation (CFI) and Natural Sciences and Engineering Research Council of Canada (NSERC) for assistance with gas chromatography analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- U.S. Energy Information Administration. Short-Term Energy Outlook; US EIA: Washington, DC, USA, 2014.
- Christensen, E.; Yanowitz, J.; Ratcliff, M.; McCormick, R.L. Renewable oxygenate blending effects on gasoline properties. Energy Fuels 2011, 25, 4723–4733. [Google Scholar] [CrossRef]
- Dürre, P. Biobutanol: An attractive biofuel. Biotechnol. J. 2007, 2, 1525–1534. [Google Scholar] [CrossRef]
- Jones, D.T.; Woods, D.R. Acetone-butanol fermentation revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–99. [Google Scholar] [CrossRef]
- Varman, A.M.; Xiao, Y.; Pakrasi, H.B.; Tang, Y.J. Metabolic engineering of Synechocystis sp. strain PCC 6803 for isobutanol production. Appl. Environ. Microbiol. 2013, 79, 908–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofuonye, E.J.; Kutin, K.; Stuart, D.T. Engineering Saccharomyces cerevisiae fermentative pathways for the production of isobutanol. Biofuels 2013, 4, 185–201. [Google Scholar] [CrossRef]
- Chen, X.; Nielsen, K.F.; Borodina, I.; Kielland-Brandt, M.C.; Karhumaa, K. Increased isobutanol production in Saccharomyces cerevisiae by overexpression of genes in valine metabolism. Biotechnol. Biofuels 2011, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Lane, S.; Zhang, Y.; Yun, E.J.; Ziolkowski, L.; Zhang, G.; Jin, Y.S.; Avalos, J.L. Xylose assimilation enhances the production of isobutanol in engineered Saccharomyces cerevisiae. Biotechnol. Bioeng. 2020, 117, 372–381. [Google Scholar] [CrossRef]
- Higashide, W.; Yongchao, L.; Yang, Y.; Liao, J.C. Metabolic engineering of Clostridium cellulolyticum for production of isobutanol from cellulose. Appl. Environ. Microbiol. 2011, 77, 2727–2733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.X.; Zhang, M.; Zhang, C.T.; Zhang, H.; Mo, X.H.; Xing, X.H.; Yang, S. Metabolomic analysis improves bioconversion of methanol to isobutanol in Methylorubrum extorquens AM1. Biotechnol. J. 2021, 16, e2000413. [Google Scholar] [CrossRef]
- Novak, K.; Baar, J.; Freitag, P.; Pflügl, S. Metabolic engineering of Escherichia coli W for isobutanol production on chemically defined medium and cheese whey as alternative raw material. J. Ind. Microbiol. Biotechnol. 2020, 47, 1117–1132. [Google Scholar] [CrossRef] [PubMed]
- Song, H.S.; Seo, H.M.; Jeon, J.M.; Moon, Y.M.; Hong, J.W.; Hong, Y.G.; Bhatia, S.K.; Ahn, J.; Lee, H.; Kim, W.; et al. Enhanced isobutanol production from acetate by combinatorial overexpression of acetyl-CoA synthetase and anaplerotic enzymes in engineered Escherichia coli. Biotechnol. Bioeng. 2018, 115, 1971–1978. [Google Scholar] [CrossRef]
- Adom, F.; Dunn, J.B.; Han, J.; Sather, N. Life-cycle fossil energy consumption and greenhouse gas emissions of bioderived chemicals and their conventional counterparts. Environ. Sci. Technol. 2014, 48, 14624–14631. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, W.; Zhang, A.; Shao, W. Biorefinery: The production of isobutanol from biomass feedstocks. Appl. Sci. 2020, 10, 8222. [Google Scholar] [CrossRef]
- Cheng, J.J.; Timilsina, G.R. Status and barriers of advanced biofuel technologies: A review. Renew. Energy 2011, 36, 3541–3549. [Google Scholar] [CrossRef]
- Ryan, C. GEVO White Paper: An Overview of GEVO’s Biobased Isobutanol Production Process; Gevo: Englewood, CO, USA, 2019. [Google Scholar]
- Weightman, R.M.; Kindred, D.R.; Clarke, S. Cereals for Bioethanol: Quantifying the Alcohol Yield of UK Hard Wheats and the Grain Yields and N Requirements of Triticale in the Second Cereal Position; HGCA Agriculture and Horticulture Development Board: Warwickshire, UK, 2011. [Google Scholar]
- Fioj, R.; Heidari, B.; Dadkhodaie, A. Investigation of triticale and wheat perfromance under dry land conditions on the basis of variations in agronomic and morphological traits. J. Adv. Biol. Biotech. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Ayalew, H.; Kumssa, T.T.; Butler, T.J.; Ma, X.F. Triticale improvement for forage and cover crop uses in the southern great plains of the United States. Front. Plant Sci. 2018, 9, 1130. [Google Scholar] [CrossRef] [Green Version]
- Puligundla, P.; Smogrovicova, D.; Obulam, V.S.; Ko, S. Very high gravity (VHG) ethanolic brewing and fermentation: A research update. J. Ind. Microbiol. Biotechnol. 2011, 38, 1133–1144. [Google Scholar] [CrossRef]
- Patni, N.; Pillai, S.G.; Dwivedi, A.H. Wheat as a promising substitute of corn for bioethanol production. Procedia Eng. 2013, 51, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Davis-Knight, H.R.; Weightman, R.M. The Potential of Triticale As a Low Input Cereal for Bioethanol Production; HGCA Agriculture and Horticulture Development Board: Warwikshire, UK, 2008. [Google Scholar]
- Montalbo-Lomboy, M.; Khanal, S.K.; van Leeuwen, J.; Raj Raman, D.; Grewell, D. Simultaneous saccharification and fermentation and economic evaluation of ultrasonic and jet cooking pretreatment of corn slurry. Biotechnol. Prog. 2011, 27, 1561–1569. [Google Scholar] [CrossRef]
- Olofsson, K.; Bertilsson, M.; Liden, G. A short review on SSF—An interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol. Biofuels 2008, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Cheng, J.Y. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Tang, Y.; Zhu, L.; Zhang, W.; Shang, X.; Jiang, J. Integrated process of starch ethanol and cellulosic lactic acid for ethanol and lactic acid production. Appl. Microbiol. Biotechnol. 2013, 97, 1923–1932. [Google Scholar] [CrossRef]
- Mendes, C.V.T.; dos Santos Rocha, J.M.; de Menezes, F.F.; da Graça Videira Sousa Carvalho, M. Batch and fed-batch simultaneous saccharification and fermentation of primary sludge from pulp and paper mills. Environ. Technol. 2017, 38, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Szymanowska, D.; Wlodzimierz, G. Fed-batch simultaneous saccharification and ethanol fermentation of native corn starch. Acta Sci. Pol. Technol. Aliment. 2009, 8, 5–16. [Google Scholar]
- Thomas, B.J.; Rothstein, R. Elevated recombination rates in transcriptionally active DNA. Cell 1989, 56, 619–630. [Google Scholar] [CrossRef]
- Gietz, D.R.; Woods, R.A. Transformation of Yeast by Lithium Acetate/Single-Stranded Carrier DNA/Polyethylene Glycol Method. In Methods in Enzymology; Guthrie, C., Fink, G.R., Eds.; Academic Press: Cambridge, MA, USA, 2002; pp. 87–96. [Google Scholar]
- Sherman, F. Getting Started with Yeast. In Methods in Enzymology; Guthrie, C., Fink, G.R., Eds.; Academic Press: Cambridge, MA, USA, 2002; pp. 3–41. [Google Scholar]
- Rabie, C.J.; Lübben, A.; Marais, G.J.; Jansen van Vuuren, H. Enumeration of fungi in barley. Int. J. Food Microbiol. 1997, 35, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ramos, D.; van den Broek, M.; van Maris, A.J.; Pronk, J.T.; Daran, J.M. Genome-scale analyses of butanol tolerance in Saccharomyces cerevisiae reveal an essential role of protein degradation. Biotechnol. Biofuels 2013, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Seguinot, P.; Sanchez, I.; Ortiz-Julien, A.; Heras, J.M.; Querol, A.; Camarasa, C.; Guillamón, J.M. Nitrogen sources preferences of non-Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions. Food Microbiol. 2020, 85, 103287. [Google Scholar] [CrossRef]
- Nganje, W.E.; Kaitibie, S.; Wilson, W.W.; Leistritz, F.L.; Bangsund, D.A. Economic Impacts of Fusarium Head Blight in Wheat and Barley: 1993–2001; Agribusiness and Applied Economics Rep. No. 528; North Dakota State University: Grand Forks, ND, USA, 2004. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Kugler, K.G.; Jandric, Z.; Beyer, R.; Klopf, E.; Glaser, W.; Lemmens, M.; Shams, M.; Mayer, K.; Adam, G.; Schüller, C. Ribosome quality control is a central protection mechanism for yeast exposed to deoxynivalenol and trichothecin. BMC Genom. 2016, 17, 417. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Iwahashi, Y. Low toxicity of deoxynivalenol-3-glucoside in microbial cells. Toxins 2015, 7, 187–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böswald, C.; Engelhardt, G.; Vogel, H.; Wallnöfer, P.R. Metabolism of the Fusarium mycotoxins zearalenone and deoxynivalenol by yeast strains of technological relevance. Nat. Toxins 1995, 3, 138–144. [Google Scholar] [CrossRef]
- Brat, D.; Boles, E. Isobutanol production from D-xylose by recombinant Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Promdonkoy, P.; Mhuantong, W.; Champreda, V.; Tanapongpipat, S.; Runguphan, W. Improvement in D-xylose utilization and isobutanol production in S. cerevisiae by adaptive laboratory evolution and rational engineering. J. Ind. Microbiol. Biotechnol. 2020, 47, 497–510. [Google Scholar] [CrossRef]
- Dickinson, J.R.; Harrison, S.J.; Hewlins, M.J. An investigation of the metabolism of valine to isobutyl alcohol in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 25751–25756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, F.; Ishii, J.; Kondo, T.; Ida, K.; Tezuka, H.; Kondo, A. Increased isobutanol production in Saccharomyces cerevisiae by eliminating competing pathways and resolving cofactor imbalance. Microb. Cell Fact. 2013, 12, 119. [Google Scholar] [CrossRef] [Green Version]
- Wess, J.; Brinek, M.; Boles, E. Improving isobutanol production with the yeast Saccharomyces cerevisiae by successively blocking competing metabolic pathways as well as ethanol and glycerol formation. Biotechnol. Biofuels 2019, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Brat, D.; Weber, C.; Lorenzen, W.; Bode, H.B.; Boles, E. Cytosolic re-localization and optimization of valine synthesis and catabolism enables inseased isobutanol production with the yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2012, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-H.; Seo, S.-O.; Bae, Y.-H.; Nan, H.; Jin, Y.-S.; Seo, J.-H. Isobutanol production in engineered Saccharomyces cerevisiae by overexpression of 2-ketoisovalerate decarboxylase and valine biosynthetic enzymes. Bioprocess Biosyst. Eng. 2012, 35, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.M.; Zhang, Y.; Mehl, J.; Park, H.; Lalwani, M.A.; Toettcher, J.E.; Avalos, J.L. Optogenetic regulation of engineered cellular metabolism for microbial chemical production. Nature 2018, 555, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Oud, B.; Flores, C.-L.; Gancedo, C.; Zhang, X.; Trueheart, J.; Daran, J.-M.; Pronk, J.T.; van Maris, A.J.A. An internal deletion in MTH1 enables growth on glucose of pyruvate-decarboxylase negative, non-fermentative Saccharomyces cerevisiae. Microb. Cell Factories 2012, 11, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, R.M.R.; Gunawardena, U.; Urano, J.; Maeinhold, P.; Aristidou, A.A.; Dundon, C.A.; Smith, C. Yeast Organism Producing Isobutanol at High Yield. U.S. Patent US8017375B2, 13 September 2011. [Google Scholar]
- Oettler, G. The fortune of a botanical curiosity—Triticale: Past, present and future. J. Agric. Sci. 2005, 143, 329–346. [Google Scholar] [CrossRef]
- Graham, R.; Geytenbeek, P.; Radcliffe, B. Response of triticale, wheat, rye and barley to nitrogen fertilizer. Aust. J. Exp. Agric. 1983, 23, 73–79. [Google Scholar] [CrossRef]
- Brock, P.; Madden, P.; Schwenke, G.; Herridge, D. Greenhouse gas emissions profile for 1 tonne of wheat produced in Central Zone (East) New South Wales: A life cycle assessment approach. Crop Pasture Sci. 2012, 63, 319–329. [Google Scholar] [CrossRef]
- Myer, R.; Lozano del Rio, A.J. Triticale as Animal Feed. In Triticale Improvement and Production; Mergoun, M., Gomez-Macpherson, H., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; pp. 49–58. [Google Scholar]
- Khan, M.K.; Pandey, A.; Athar, T.; Choudhary, S.; Deval, R.; Gezgin, S.; Hamurcu, M.; Topal, A.; Atmaca, E.; Santos, P.A.; et al. Fusarium head blight in wheat: Contemporary status and molecular approaches. 3 Biotech 2020, 10, 172. [Google Scholar] [CrossRef]
- U.S. FDA. Guidance for Industry and FDA: Advisory Levels for Deoxynivalenol (DON) in Finished Wheat Products for Human Consumption and Grains and Grain By-Products Used for Animal Feed. 2010. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-fda-advisory-levels-deoxynivalenol-don-finished-wheat-products-human (accessed on 19 February 2021).
- Elmekawy, A.; Diels, L.; De Wever, H.; Pant, D. Valorization of cereal based biorefinery byproducts: Reality and expectations. Environ. Sci. Technol. 2013, 47, 9014–9027. [Google Scholar] [CrossRef]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Eksteen, J.M.; Van Rensburg, P.; Cordero Otero, R.R.; Pretorius, I.S. Starch fermentation by recombinant Saccharomyces cerevisiae strains expressing the alpha-amylase and glucoamylase genes from Lipomyces kononenkoae and Saccharomycopsis fibuligera. Biotechnol. Bioeng. 2003, 84, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Li, Z.; Jia, C.; Zhang, W.; Zhang, Y.; Yi, C.; Xie, S. Recent advances on bio-based isobutanol separation. Energy Convers. Manag. X 2021, 10, 100059. [Google Scholar]
Figure 1.
SSF of triticale (squares), wheat (triangles) and barley (circles) mashes. (A). Viable yeast cells in each mash at each time point determined by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L (filled symbols) and ethanol concentration in g/L (open symbols) was measured in the triticale (squares), wheat (triangles) and barley (circles) cultures. Values displayed are the means of triplicate measures. Error bars indicate standard deviations.
Figure 1.
SSF of triticale (squares), wheat (triangles) and barley (circles) mashes. (A). Viable yeast cells in each mash at each time point determined by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L (filled symbols) and ethanol concentration in g/L (open symbols) was measured in the triticale (squares), wheat (triangles) and barley (circles) cultures. Values displayed are the means of triplicate measures. Error bars indicate standard deviations.
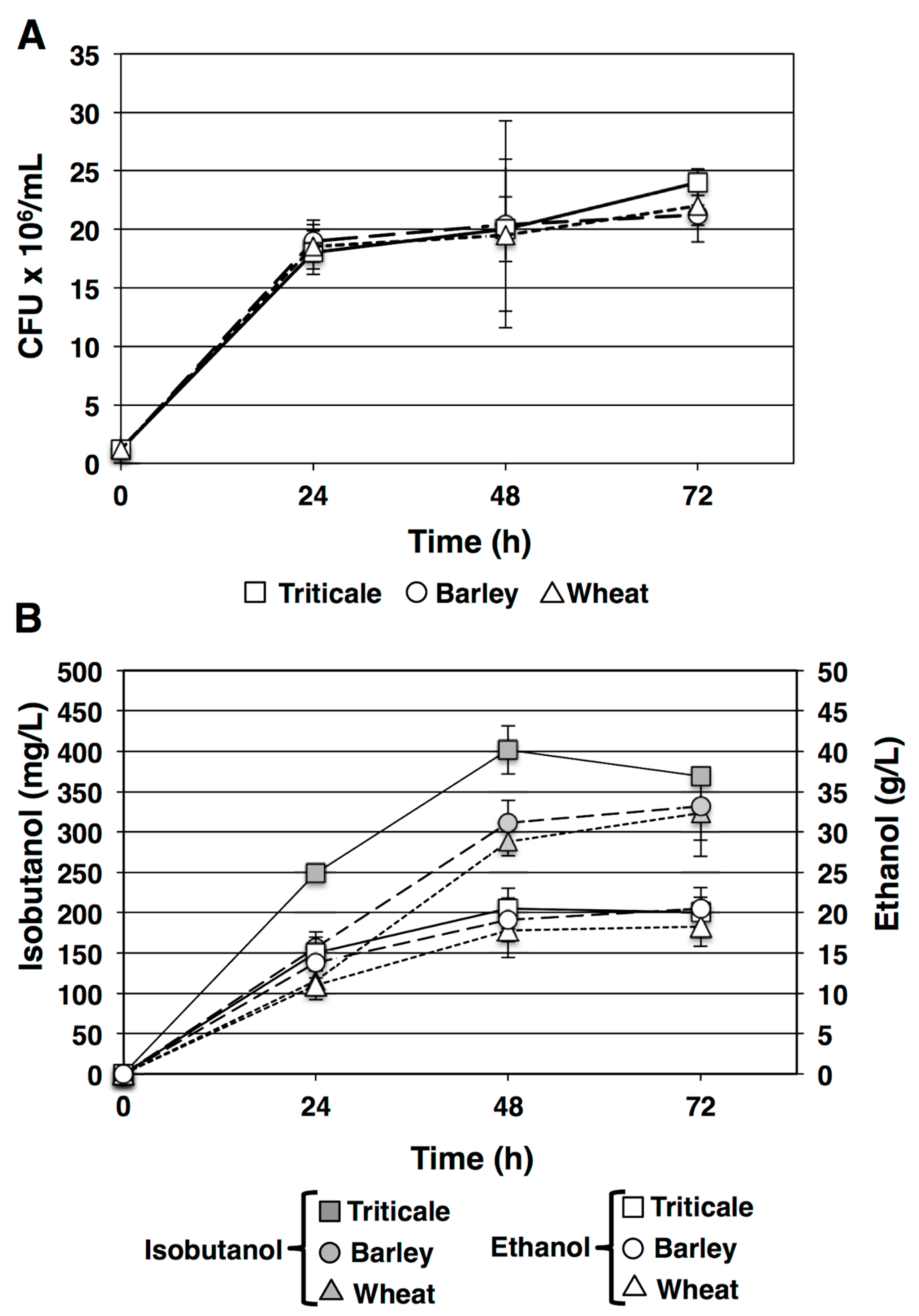
Figure 2.
SSF in batch (open symbols) and fed-batch (filled symbols) fermentations of triticale starch. (A) Viable yeast cell counts at each time point determined by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L in fermentation reactions (square symbols), ethanol concentration in g/L in the fermentation reactions (circles), batch fermentation values are shown in the open symbols and fed-batch in filled symbols. The vertical arrow indicates the time of the starch feed addition. Values displayed are the means of triplicate cultures. Error bars indicate standard deviations.
Figure 2.
SSF in batch (open symbols) and fed-batch (filled symbols) fermentations of triticale starch. (A) Viable yeast cell counts at each time point determined by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L in fermentation reactions (square symbols), ethanol concentration in g/L in the fermentation reactions (circles), batch fermentation values are shown in the open symbols and fed-batch in filled symbols. The vertical arrow indicates the time of the starch feed addition. Values displayed are the means of triplicate cultures. Error bars indicate standard deviations.

Figure 3.
SSF of uninfected (Clean, circles) and F. graminearum infected (Fusarium, squares) barley. (A). Viable yeast cell counts at each time point from fermentation of clean barley starch feedstock (circles) or barley starch prepared from barley experimentally infected with F. graminearum (squares) by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L (filled symbols) and ethanol concentration in g/L (open symbols) was measured in fermentation reactions fed with clean barley starch (circles) or F. graminearum-infected barley starch (squares). Values displayed are the means of triplicate measures. Error bars indicate standard deviations.
Figure 3.
SSF of uninfected (Clean, circles) and F. graminearum infected (Fusarium, squares) barley. (A). Viable yeast cell counts at each time point from fermentation of clean barley starch feedstock (circles) or barley starch prepared from barley experimentally infected with F. graminearum (squares) by CFU. Data shown are mean values from triplicate platings. Error bars indicate standard deviation. (B) Isobutanol concentration in mg/L (filled symbols) and ethanol concentration in g/L (open symbols) was measured in fermentation reactions fed with clean barley starch (circles) or F. graminearum-infected barley starch (squares). Values displayed are the means of triplicate measures. Error bars indicate standard deviations.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Sugar concentrations in starch mashes mg/100 mL.
| Grain | Sugar | 0 h | 24 h | 48 h | 72 h |
|---|---|---|---|---|---|
| Triticale | Glucose | 147.99 ± 6.42 | 7.11 ± 0.49 | 6.38 ± 0.22 | 6.81 ± 0.83 |
| Xylose | 1.9 ± 0.01 | 2.4 ± 0.03 | 2.7 ± 0.01 | 2.8 ± 0.1 | |
| Wheat | Glucose | 107.66 ± 14.23 | 3.49 ± 0.17 | 3.11 ± 0.22 | 6.02 ± 1.62 |
| Xylose | 2.4 ± 0.06 | 2.7 ± 0.1 | 2.9 ± 0.03 | 2.9 ± 0.08 | |
| Barley | Glucose | 129.65 ± 8.4 | 5.82 ± 1.2 | 4.21 ± 0.3 | 5.91 ± 1.5 |
| Xylose | 3.1 ± 0.1 | 3.5 ± 0.03 | 3.3 ± 0.08 | 3.3 ± 0.2 |
Table 2.
Influence of nitrogen source on isobutanol and ethanol synthesis a.
| Nitrogen Source | Isobutanol mg/L b | Ethanol g/L b |
|---|---|---|
| None added | 62.31 ± 18.11 | 4.83 ± 0.37 |
| 10 mM Urea | 398.01 ± 25.06 | 20.33 ± 0.08 |
| 20 mM Urea | 397.82 ± 12.46 | 18.34 ± 0.11 |
| 10 mM (NH4)2SO4 | 396.83 ± 25.77 | 19.38 ± 0.10 |
| 20 mM (NH4)2SO4 | 389.12 ± 17.87 | ND |
| 20 mM NH4Cl | 390.57 ± 34.80 | 18.77 ± 0.18 |
| 40 mM NH4Cl | 387.82 ± 27.89 | 17.34 ± 0.41 |
Values reported are mean concentrations ±SD derived from three replicate 120 mL cultures. a All mashes were prepared for SSF procedure with 80 g/L starch. b Alcohol titres were determined 48 h following inoculation with the yeast. ND—Not determined.
Table 3.
Initial characteristics of F. graminearum contaminated barley mashes.
| Glucose (mg/100 mL) | Xylose (mg/100 mL) | DON (µg/100 mL) | |
|---|---|---|---|
| Barley (Fusarium) | 123 ± 7.9 | 2.9 ± 0.4 | 150.4 ± 12.5 |
| Barley (clean) | 138 ± 6.1 | 4.3 ± 0.6 | 0.08 ± 0.01 |
Values for sugars are mean concentrations from triplicate cultures ±SD. DON values determined by competitive ELISA are mean values from triplicate cultures ±SD.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, X.; Unaegbunam, E.; Stuart, D.T. Co-Production of Isobutanol and Ethanol from Prairie Grain Starch Using Engineered Saccharomyces cerevisiae. Fermentation 2021, 7, 150. https://doi.org/10.3390/fermentation7030150
AMA Style
Liu X, Unaegbunam E, Stuart DT. Co-Production of Isobutanol and Ethanol from Prairie Grain Starch Using Engineered Saccharomyces cerevisiae. Fermentation. 2021; 7(3):150. https://doi.org/10.3390/fermentation7030150
Chicago/Turabian StyleLiu, Xiaodong, Ebele Unaegbunam, and David T. Stuart. 2021. "Co-Production of Isobutanol and Ethanol from Prairie Grain Starch Using Engineered Saccharomyces cerevisiae" Fermentation 7, no. 3: 150. https://doi.org/10.3390/fermentation7030150
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.