
Dairy-Based Probiotic-Fermented Functional Foods: An Update on Their Health-Promoting Properties



1
Animal Biochemistry Division, Indian Council of Agricultural Research-National Dairy Research Institute (ICAR-NDRI), Karnal 132001, India
2
Centre for Healthy Brain Ageing, School of Psychiatry, University of New South Wales, Sydney, NSW 2052, Australia
3
Mark Wainwright Analytical Centre, Bioanalytical Mass Spectrometry Facility, University of New South Wales, Sydney, NSW 2052, Australia
4
Cell Biology and Proteomics Lab, Animal Biotechnology Center, ICAR-NDRI, Karnal 132001, India
5
Division of Proteomics of Stem Cells and Cancer, German Cancer Research Center (DKFZ), 69120 Heidelberg, Germany
*
Author to whom correspondence should be addressed.
Fermentation 2022, 8(9), 425; https://doi.org/10.3390/fermentation8090425
Submission received: 19 July 2022
/
Revised: 14 August 2022
/
Accepted: 22 August 2022
/
Published: 27 August 2022
(This article belongs to the Special Issue Trends in the Development and Use of Fermented Dairy Products)
Abstract
:Numerous studies have shown a link between the consumption of fermented dairy foods and improved health outcomes. Since the early 2000s, especially probiotic-based fermented functional foods, have had a revival in popularity, mostly as a consequence of claims made about their health benefits. Among them, fermented dairy foods have been associated with obesity prevention and in other conditions such as chronic diarrhea, hypersensitivity, irritable bowel syndrome, Helicobacter pylori infection, lactose intolerance, and gastroenteritis which all are intimately linked with an unhealthy way of life. A malfunctioning inflammatory response may affect the intestinal epithelial barrier’s ability to function by interfering with the normal metabolic processes. In this regard, several studies have shown that fermented dairy probiotics products improve human health by stimulating the growth of good bacteria in the gut at the same time increasing the production of metabolic byproducts. The fermented functional food matrix around probiotic bacteria plays an important role in the survival of these strains by buffering and protecting them from intestinal conditions such as low pH, bile acids, and other harsh conditions. On average, cultured dairy products included higher concentrations of lactic acid bacteria, with some products having as much as 109/mL or g. The focus of this review is on fermented dairy foods and associated probiotic products and their mechanisms of action, including their impact on microbiota and regulation of the immune system. First, we discussed whey and whey-based fermented products, as well as the organisms associated with them. Followed by the role of probiotics, fermented-product-mediated modulation of dendritic cells, natural killer cells, neutrophils, cytokines, immunoglobulins, and reinforcement of gut barrier functions through tight junction. In turn, providing the ample evidence that supports their benefits for gastrointestinal health and related disorders.
1. Introduction
Milk and dairy products are known to contain bioactive compounds that are needed for many biochemical and physiological processes [1]. Because of this reason, they are an important part of a healthy diet. Fermented dairy functional foods, when consumed in enough proportions as part of a balanced diet on a regular basis, provide health benefits. In addition, this functional food also provides the body with critical nutrients such as vitamins and minerals [1]. Since fermented dairy functional foods are one of the fastest-growing parts of the food industry, it is important to know what they are. In addition to providing nutrients and energy, functional foods have a positive effect on one or more specific biological functions, either by improving a particular physiological response or by lowering the risk of disease, mostly by protecting the gut epithelial layer and maintaining the balance for gut immune homeostasis [2]. The epithelial cells in the gut serve as primary barrier surfaces that separate hosts from their external environment and have a substantial influence on the formation and function of the mucosal immune system.
Dietary components have the utmost influence on the intestinal environment due to their ability to modulate the intestinal barrier and perform various biochemical processes such as antimicrobial, antihypertensive, antioxidant, and anticancer [3]. Many studies have been undertaken to investigate the role of dietary components of fermented functional foods in intestinal permeability to identify potential correlations between diet, permeability, obesity, and inflammatory bowel disease and other diseases. Additionally, fermented dairy products are ideal for delivering probiotic bacteria to the human gut and provide the optimal environment for their growth [4]. Probiotics are described as “live microorganisms that bestow a health benefit on the host when provided in sufficient doses” [5,6]. Lactic acid bacteria (LAB) such as Lactobacillus, Streptococcus, Lactococcus, Bifidobacterium, and Leuconostoc are most common probiotic bacterial strains in fermented dairy products, either as starter cultures or naturally existing components of the raw material. Fermentation preserves and enhances the viability and production of microorganisms while retaining their probiotic characteristics [7,8]. The molecular components of probiotics associated with fermented foods influence disease-regulating mechanisms either as prophylactics or therapeutically, and such foods are often known as nutraceuticals, foodiceuticals, functional foods, or medifoods [9]. Recently, extensive research has been conducted on the effect of fermented functional products on human health, including epidemiological, observational, and clinical investigations. This review focuses on the importance of dietary fermented functional foods and their role in maintaining the homeostasis of gut microbiota and the associated immune system.
2. Dairy-Based Functional Food: Its Composition and Application in Human Health
Milk is the richest source of nutrients coupled with bioactive functional characteristics which contributes to the growth and nourishment of infants. In milk, whey accounts for around 85–95 percent of milk’s volume and maintains 55% of milk nutrients. Of these, lactose (4.5–5 percent w/v), soluble proteins (0.6–0.8 percent w/v), lipids (0.4–0.5 percent w/v), and mineral salts (8–10 percent w/v of dry extract) are the most prevalent nutrients. Whey contains more than half of total milk solids, lactose, and water-soluble vitamins as well as whey protein (20% of total protein), which is rich in essential amino acids such as lysine, isoleucine, threonine, and tryptophan [10]. Beta-lactoglobulin (BLG) (58%) and alpha-lactalbumin (13%), together with trace levels of immunoglobulin, serum albumin, and protease peptone, are the predominant whey proteins. It includes a variety of anti-microbial peptides and other compounds that have a beneficial effect on a number of diseases [11,12]. Most whey salts are composed of sodium chloride and potassium chloride (>50%), and calcium salts (mainly phosphate). In addition to these, whey contains lactic (0.5% w/v) and citric acids, non-protein nitrogen molecules such as urea and uric acid, and B group vitamins.
Studies also suggested that the fermentation of whey produces protein hydrolysates that are more absorbable than complete proteins, improve gastrointestinal health, reduce allergies/intolerance-related symptoms, and have immune-boosting effects [13]. Using mass-spectrometer-based high throughput analysis, it was shown that the milk whey contains 6210 unique proteins that are engaged in a wide variety of biological activities [14]. In addition to a high number of milk proteins, it also known to contain omega-3 fatty acids, phytosterols, isoflavins, conjugated linoleic acid, minerals, and vitamins, and so has a significant impact on the creation of functional foods. In vitro and in vivo studies have observed that fermented dairy products assisted in the maintenance of normal mucosal homeostasis and provide protection against various metabolic and pathogen-mediated diseases through their anti-oxidative, anti-microbial, anti-fungal, anti-inflammatory, anti-diabetic, and anti-atherosclerotic properties (Table 1).
Fermented whey is not only a source of nutrients; it also plays a role in the regulation of several physiological activities in the body [43]. This is particularly true for the intestinal tract, which is constantly in contact with dietary antigens. Numerous health benefits linked with fermented whey or whey protein ingestion have been well documented, including decreased inflammatory gene expression, protection against diarrhea caused by DSS (sodium trimethylsilylpropanesulfonate) [44], and significant influence in gut microbiota modification. These positive benefits may also be mediated by whey protein extract, which promotes mucosal innate immunity in childhood [45].
2.1. Other Dairy-Based Fermented Products
Another important product arises from the dairy industry is yogurt. The yogurt itself is not a probiotic food, but it could have non-probiotic bacteria that grew from milk fermentation. However, it can also be made better by adding probiotic bacteria. Probiotic yogurt is a combined mixture of probiotic (L. rhamnosus GG) and prebiotic (dietary fiber), and its consumption aids in the improvement of inflammatory bowel syndrome (IBS) and the maintenance of normal fecal microflora in patients with decreased putrefactive bacteria Clostridium difficile and E. coli. Recent research found that fermented probiotic yogurt improved IBS symptoms such as constipation, stomach pain, and frequency of bowel movements within six weeks of intake [46]. Another common fermented dairy product that has the ability to assist the human digestive system is cheese. It has a higher pH, a lower titratable acidity, a greater buffering capacity, a more solid consistency, a larger fat content, increased nutritional availability, and a lower oxygen concentration than yogurt. These characteristics serve to safeguard probiotic microorganisms throughout storage and transit through the gastrointestinal system [47,48]. Kefir can be produced using dairy culture as well as koumiss. In the industrial production of kefir, dairy culture is used more often than kefir grain. In addition, koumiss and kefir are almost identical when both are made using yeast and lactic acid fermentation [49,50]. Recently, Li et al. discovered that koumiss resolves thymus and spleen atrophies, increases the number of leukocytes, lymphocytes, and CD4+/CD8+ ratio of peripheral blood lymphocytes in an immunosuppressed rat model. In addition, it was able to efficiently enhance the structure of the small intestine mucosa, indicating that koumiss might serve as a therapeutic agent with a beneficial impact on intestinal immune function in immunocompromised conditions [51]. A study by Tiss et al. (2020) showed that under kefir fermented drink, in vivo and in vitro conditions showed a decrease in intestinal and pancreatic lipase activity, leading to a reduction in total cholesterol and LDL-cholesterol, and an increase in HDL-cholesterol rates, as well as body weight loss [52] (Table 2).
2.2. Positive Influence of Fermented Whey-Based Products
In addition to essential nutrients, fermented whey includes a wide range of natural bioactive chemicals such as lactalbumin, lactoferrin, lactoperoxidase, lysozyme, EGF, and TGF [58]. A milk-derived whey protein concentrate rich in TGF has been developed by the food industry to stabilize epithelial barrier function and protect the intestinal barrier from inflammatory damage in infants and children [59]. TGF-1 boosted the activity of the claudin-4 promoter and protected HT-29/B6 cells against IFN-γ-induced barrier damage in a dose-dependent manner [60]. Furthermore, fermented whey supplementation protects intestinal integrity by decreasing the production of pro-inflammatory cytokines in the intestinal tract and increasing the expression of TJ proteins such as occludin and ZO-1 in LPS-treated pigs.
The anti-inflammatory properties of goat whey were investigated in a mouse model of colitis induced by 2,4-dinitrobenzenesulfonic acid (DNBS). Goat whey alleviated intestinal inflammatory symptoms, by lowering the disease activity index, colonic weight/length, and leukocyte infiltration. Additionally, goat whey boosted the expression of mucins, occludin proteins, and suppressors of cytokine signaling [61]. Bovine WPE (whey protein extract) inhibited apoptosis in human blood neutrophils and had a dose-dependent priming impact on these cells, enhancing their adhesion, chemotaxis, phagocytosis, oxidative burst, and degranulation capacities in response to further stimulation. Not only did WPE stimulate neutrophil chemotaxis, but it also raised the surface expression of CD11b, a mediator of neutrophil adherence to endothelial cells [62].
On the other hand, the bioactive forms of proteins and growth factors found in low-heat-treated whey protein concentrate (WPC) promote cell proliferation and immune response enhancement. Lactoferrin and TGF-2 were retained at a higher concentration level in low-heat-treated WPC compared to regular WPC in in vitro conditions [63]. Glycated Whey Protein Concentrate (G-WPC) generated by a non-enzymatic Maillard process exhibited no cytotoxic effect on RAW264.7 macrophage cells but elevated the production of several cytokines (i.e., TNF-α, IL-1, and IL-6 mRNA) that were shown to promote phagocytosis [64]. Interleukin-2 receptor surface expression and interferon production may also be inhibited when modified WPC is added to T and B cell cultures [65]. The NF-kB and MAPK pathways in neutrophils treated with whey protein extract generated more IL-1Ra than IL-1 [66]. Low-heat-treated WPC from acid whey substantially boosted cytokine response and proliferation in intestinal epithelial cells (IEC) as compared to low-heat-treated sweet whey [67]. WPC’s bioactive peptides promoted innate immunity in children by largely elevating CD8+ IEL (intestinal intraepithelial lymphocytes) and NK cells (natural killer cells) in the mucosal location [43]. Goat whey inhibited the production of many proinflammatory markers such TNF-α, IL-1, and IL-6 in the colonic tissue of mice with DNBS-induced colitis, as shown by [61]. Moreover, Rusu et al. found that WPE induced a time-dependent phosphorylation of inhibitory kappa B (IκB), the inhibitor of NF-B in neutrophils, which released IkB from NF-κB and activated NF-κB, thereby activating the immune response [62].
Recent research by Gupta et al. [68] showed that the fermented probiotic L. rhamnosus MTCC 5897 whey exhibited anti-inflammatory properties against E. coli-induced inflammation in intestinal epithelial cells (Caco-2) [68]. Similarly, Kaur et al. found that L. rhamnosus MTCC 5897 fermented whey had an intriguing effect on enhancing the intestinal epithelial barrier function in ulcerative colitis rats. Additionally, it may increase the RBC and lymphocyte counts in cases of anemia, which is critical for the body’s capacity to fight against infections [16]. In pregnancy, during the third trimester, the administration of a probiotic-fermented milk containing Streptococcus thermophilus, Lactobacillus bulgaricus, Lactobacillus acidophilus LA5, and Bifidobacterium animalis subsp. lactis BB12 has been associated with a decreased risk of maternal insulin resistance. In addition, yogurt intake has been associated with immunological advantages, such as a lower level of inflammatory markers in pregnant women [69] and has been shown to alter both humoral and cellular immunity [70].
2.3. Fermented Dairy Product as a Probiotic Vehicle
Fermented dairy products provide an ideal atmosphere for delivering probiotic bacteria to the human body (Figure 1). Probiotic fermentation combines the health benefits of fermented food with the benefits of good bacteria by increasing the amount of bioactive peptides that are released [71]. Additionally, because of the presence of high levels of lactic, butyric, citric, and acetic acids found in dairy products including cream, cheese, yogurt, acidophilus-bifidus milk, koumiss, kefir, and fermented drinks, these items have long been regarded the ideal matrix for probiotic microorganisms [72]. Rezac et al. observed a wide range of fermented foods and discovered that most of them had at least 105–7 LAB per milliliter or gram, but that this number varied greatly depending on sampling time and location. Bacteria at quantities of up to 109/mL or g were often found in cultured dairy products [8]. Understanding the distinguishing characteristics of these species is greatly aided by cutting-edge omics technologies [73]. As a result, it very important to understand the genomics and proteomics profile of lactobacillus probiotics bacteria in order to gain insight into their beneficial properties [74,75,76,77,78,79].
According to Swanson et al. [80], guidelines for the minimum viable count of each probiotic strain per gram or milliliter of probiotic products differ in terms of providing probiotic-organism-related health benefits [80]. To begin, fermented dairy products such as probiotic yogurt are often believed to contain living microorganisms because, unlike many other fermented foods, dairy products are usually not pasteurized or heated at the point of manufacture, ensuring microbial viability [81]. Additionally, bacterial growth and survival in dairy products have been extensively researched [82]. A recent study demonstrated that the culture bacteria did not decrease in quantity after frozen storage of low-fat yogurt including the probiotic strains Streptococcus thermophiles and L. acidophilus [83]. Yogurt’s composition, which includes milk proteins, fat, and lactose, among other components, has proven to be an ideal environment for microbes. Its efficiency is enhanced by its frozen state and the recommended pH of yogurt is from 5.5 to 6.5 to ensure the viability of the lactic cultures during storage. Additionally, the lowered acidity promotes client acceptance, especially among those who choose milder products [84]. Therefore, fermented dairy products are the most effective probiotic delivery system (Figure 1).
Similarly, fermented milk treated with lemon and orange fibers boosted L. acidophilus numbers and improved cold storage stability when compared to the control group [85]. Likewise, fermented dairy cheese has a great potential for introducing beneficial microorganisms into the human gut owing to its unique physicochemical properties as described earlier in the text. The valuable physiochemical qualities of cheese keep probiotic microorganisms safe while they are stored and move through the gastrointestinal system [47,86]. In this way, it is safe to report that fermentation permits the maintenance and optimization of microbial viability and production while preserving probiotic properties [87].
2.4. Health Benefits of Probiotic Functional Food
Fermented probiotics are live microbial dietary additives that provide several health advantages by assisting in the stability and maintenance of the gut microbiota’s composition and enhancing resistance to pathogen infection [15]. Probiotics are also considered as potential functional foods since they give much more health advantages than standard nutritional diets [88]. As public knowledge of the influence of nutrition on health rises, the market for probiotic functional foods increases swiftly and persistently [89,90]. Probiotics have been shown to be effective in treating a variety of diarrheal illnesses, modulating immunological function, preventing colon cancer, and alleviating other chronic gastrointestinal inflammatory disorders, as shown by a growing body of high-quality, scientific clinical research [91,92]. The potential effectiveness of probiotics is increasing the interest in determining the activities and therapeutic benefits in a wide variety of neurological diseases too [93] (Figure 2).
2.4.1. Obesity and Diabetes
Diabetes is a major public health problem worldwide. Several reports have suggested the anti-diabetic properties of probiotic food containing L. paracasei and L. rhamnosus [94]. LAB alone dramatically decreased hyperinsulinemia, hyperglycemia, and glucose intolerance, suggesting a viable preventative treatment option for metabolic syndrome. According to Alihosseini et al. [95], diabetic patients who consumed probiotic-fermented milk (kefir) had lower fasting blood glucose and HbA1C levels than those who received conventional fermented milk [95]. In other research, when administered orally to 8-week-old diabetic male rats, fermented milk containing the L. casei strain AP was as effective as the commonly used diabetes drug metformin [96]. Another study demonstrated that consumption of probiotic fermented milk containing L. rhamnosus (MTCC 5897), L. fermentum (MTCC 5898), and L. rhamnosus (MTCC 5957) had an anti-diabetic effect both individually and in combination by regulating glucose metabolism, inflammation status, and serum lipid profile in diabetic rats compared to the control group [97].
2.4.2. Lactose Intolerance
Even in nations where the majority of the population can digest lactose without issue, lactose intolerance may manifest as a medical disease. Symptoms comparable to celiac disease may develop in individuals who are lactose intolerant due to the toxicity of lactose and the damage it causes to the intestinal mucosa [98]. Considering the widespread occurrence of lactose intolerance, it is preferable that milk has as little lactose as possible [99]. Because fermented milk often has lower quantities of lactose than low fat milk, it may be tolerated by those who otherwise cannot consume lactose [100]. It has been hypothesized that lactase-containing bacteria present in fermented milk persist in the digestive tract and aid in the digestion of ingested lactose [101]. Drinking fermented milk products, especially yogurt and acidophilus, was advocated for the health of Swedish volunteers aged 18–50 [102]. When planning meals for those who are lactose intolerant, it is important to keep in mind that neither yogurt nor acidophilus milk were reported to cause any gastrointestinal distress among the responders [100]. Similarly, probiotic ingestion shortened the duration of diarrhea, reduced inflammatory responses, and fostered the growth of beneficial microbes in the digestive tract [103].
2.4.3. Allergies
Consumption of probiotic-fermented foods alleviates the symptoms of food allergies in vulnerable people. A meta-analysis evaluating the effects of supplementation with various lactobacilli combinations on atopic dermatitis in normal and allergic risk groups revealed beneficial effects [62]. Consumption of probiotics such as L. gasseri KS-13, B. bifidum G9-1, and B. longum MM-2 in placebo-controlled research showed an anti-allergic effect in young people suffering from seasonal allergies through immune response regulation [104]. Further, Liu et al. [105] demonstrated that administration of the probiotic B. infantis CGMCC313-2 inhibited allergen-induced IgE antibody secretion as well as Th2 cytokine secretion and further attenuated allergic inflammation in a mouse model of allergen-induced airway inflammation and lactoglobulin-induced food allergies [105]. Saliganti et al. found that L. rhamnosus (MTCC 5897), another probiotic, reduced ovalbumin-induced allergy reactions in weanling mice [105] (Figure 3).
2.4.4. Oxidative Stress
Bioactive peptides produced by probiotics are effective antioxidants that may reduce the effects of free radical damage. The probiotic B. animalis 01 improved anti-oxidative enzyme efficiency and reduced lipid oxidation in elderly mice by decreasing MDA, lipofuscin levels, and monoamine oxidase activity [106,107]. According to the findings of Wang et al., Bacillus amyloliquefaciens SC06 has an anti-oxidative effect on intestinal porcine epithelial cells via decreasing reactive oxygen species (ROS) and regulating the Nrf-2 and Keap-1 pathways [108]. To inhibit ascorbate auto-oxidation and the generation of amino butyric acid, Garcia et al. [69] isolated L. plantarum DM5 from fermented beverages and demonstrated its anti-oxidative activity by decreasing hydroxyl radicals [69]. Another study reported that consuming fermented goats’ milk over the course of 21 days boosted overall antioxidative activity, reduced peroxidized lipoproteins, oxidized LDL, 8-isoprostanes, and the glutathione redox ratio, and improved anti-atherogenicity in healthy human individuals [109].
2.4.5. Skin Repair
Probiotic L. paracasei NCC 2461 supplementation reduced skin irritation and enhanced skin barrier function in female groups during a two-month placebo-controlled study [110]. A recent study has shown that the cell wall components isolated from L. rhamnosus GG were shown to have immunomodulatory effects against UV radiation and to inhibit the formation of skin cancers [111]. In 2019, Khmaladze et al. [112] looked at how probiotic L. reuteri DSM and its cell lysate affected overall skin health. The researchers found that the anti-inflammatory and antibacterial characteristics of probiotic bacteria helped reduce photoaging and provided greater protection against UVR-B-mediated inflammatory pathways [112]. Additionally, oral administration of L. rhamnosus GG improved skin health by reducing water loss and preserving moisture [113].
2.4.6. Hypercholesterolemia
Cardiovascular diseases (CVDs) are the major cause of mortality worldwide and are continually growing, spreading, and becoming more frequent around the globe. By 2030, the WHO predicts that CVDs would account for up to 40% of all fatalities, impacting roughly 23.6 million people worldwide. Indeed, people with hypercholesterolemia have a threefold increased risk of heart attack compared to those with normal blood lipid levels [114]. To prevent hypercholesterolemia, there has been significant interest in dietary and pharmacological interventions that block cholesterol absorption [115]. Dairy-fermented products with LAB are expected to reduce circulating cholesterol levels, hence lessening the population’s risk of cardiovascular disease. These microbes must be bile resistant and capable of deconjugating and binding bile acids and cholesterol. If these parameters are satisfied, fermented dairy products may be beneficial for lowering elevated cholesterol levels and avoiding CVDs [116].
2.4.7. Gastrointestinal Disorders
Probiotic strains for treating inflammatory bowel illness are Lactobacillus, Bifidobacterium, and designer probiotics are most commonly used in supplemented fermented foods [117]. Probiotics promote gut health by restoring a healthy balance to the intestinal flora and decreasing the probability of colonization by pathogenic microorganisms [118]. Likewise, Dore et al. investigated the effect of administering a specific dose of L. acidophilus affected ulcerative colitis symptoms in colitis mice by altering the microbiota composition in the distal colon [119]. In functional dyspepsia patients, fermented milk containing the probiotic Bifidobacterium bifidum YIT 10347 improves stomach and lower abdominal symptoms associated with the formation of gastro-protective mucin [120]. According to Eales et al., frequent use of probiotic food with the specific B. lactis DN-173 010 strain may improve gastrointestinal well-being and digestive symptoms in adult women with mild digestive illnesses [121].
3. Probiotic-Fermented Products: Stimulation and Regulation of the Immune System
Inflammation is the immune system’s response to infections, damaged cells, and toxins, among other adverse stimuli. It is a common cause of several chronic diseases, including cardiovascular disease, diabetes, rheumatoid arthritis, inflammatory bowel disease, and cancer [122]. It is essential to consume foods that possess anti-inflammatory properties. Therefore, consuming fermented probiotic foods monitors the creation of beneficial anti-inflammatory compounds. In this regard, the notable very good example is the process of fermentation of food especially by some LAB strains which potentially results in the elimination of lactose and galactose from fermented milk, hence reducing the risk of lactose intolerance and galactose buildup. At the same time, the most important aspect to note is that lactose is converted into lactic acid, which, in addition to other bioactive components, has anti-inflammatory and immunomodulatory characteristics [123].
3.1. Probiotic-Fermented-Product-Mediated Modulation of DCs
It was reported in many studies that fermented probiotics have a positive effect on intestinal epithelial cells (IECs). IECs are essential in driving the development of tolerance by suppressing DC activation, which subsequently controls the suppressive function of T regulatory cells (Tregs). IECs have also been designated to stimulate the development of CD103+ DC. This population of tolerogenic DCs induces Foxp3+ Treg cells and is dependent on TGFβ (Figure 3 and Figure 4). Under the effects of probiotics and commensal microbiota, the activated DCs induce the appropriate immune response (e.g., naïve CD4 T cells to Treg cell differentiation), which generally inhibits Th-1, Th-2, and Th17-mediated inflammatory response [124]. In vitro models based on cell lines or individual immune cells are especially appealing. These models could also be useful to quantify the development of nonspecific and specific immune responses by monitoring parameters such as cytokine and mediator production by antigen-presenting cells and biological markers for distinct cell types. Through a TLR-2 signaling route, cell-free concentrated whey from B. breve C50-fermented milk (BbC50sn) generated a unique interleukin-10 (IL-10)-rich cytokine profile and extended DC survival. The anti-apoptotic BbC50sn signal outperforms lipopolysaccharide’s pro-apoptotic action on DCs [125]. In a recent study, Jeffrey et al. showed that milk fermented with L. rhamnosus R0011 enhanced regulatory cytokine release from LPS-challenged U937 and THP-1 macrophages, while simultaneously upregulating the production of IL-1β and expression of DC-SIGN and CD206, a profile characteristic of polarization into the immunoregulatory M2 macrophage phenotype [126]. L. salivarius Ls33 stimulated the CD103+ DCs and CD4+ Foxp3+ regulatory T cells’ development in an IL-10-dependent manner [127]. Purification of peptidoglycan (PGN) from L. salivarius Ls33 demonstrated that the protective effects of L. salivarius were NOD2-dependent but MyD88-independent. Another study has identified a probiotic mixture, IRT5 (Streptococcus thermophilus, L. reuteri, B. bifidium, L. acidophilus, and L. casei), which upregulates the generation of CD4+ Foxp3+ regulatory T cells (Tregs) from the CD4+ CD25− population [128]. Overall, the data indicate that fermented probiotic factors and their bacterial components may stimulate DC maturation. In addition, evidence suggests that fermented probiotics have a specialized function in regulating intestinal DCs.
3.2. Probiotic-Fermented-Product-Mediated Modulation of NK Cells
Natural killer cells (NKs) play a crucial role in the immune response and regulation of homeostasis. They can distinguish between normal to abnormal cells with altered or missing MHC class I molecules (Figure 4). As people become older, their immune functions begin to deteriorate. It has been claimed that probiotic supplementation may reduce or reverse these age-related alterations. To confirm, a systematic meta-analysis reported that short-term probiotic supplementation (duration ranged from 3 to 12 weeks) in the elderly adults improves the cellular immune response via polymorphonuclear cell phagocytic capacity or natural killer (NK) cell tumoricidal activity [129]. Recently, another study by Gui et al. showed that probiotic Lactobacillus interventions alone or in combination with Bifidobacterium enhances the natural killer (NK) cell function in healthy elderly individuals [130]. A study conducted in Japan by Shida et al. on healthy volunteers showed that regular consumption of L. casei Shirota-fermented milks orally for three weeks resulted in the improved NK cell activity [131]. It found to be very helpful for individuals with low NK cell activity and in turn help in the overall better health and reduction in illness, as measured in peripheral blood mononuclear cells (PBMC). In a separate study, the administration of L. casei Shirota (LcS) to patients whose colonic polyps had been surgically removed has shown a significant reduction in the recurrence of colorectal cancer. The results also showed the specific mechanisms of action of LcS on positively correlated NK cell activity via induction of IL-12 and TNFα production [132].
However, ingestion of a combination of eight strains of sonicated probiotic bacteria named sAJ2 (Streptococcus thermophiles, B. longum, B. breve, B. infantis, L. acidophilus, Lactobacillus plantarum, and L. casei) by patients demonstrated improvement in the NK function both in terms of IFN-γ secretion as well as NK-cell-mediated cytotoxicity after four weeks of consumption [133]. The relevance of bacteria-mediated enhancement of immune activation and restoration of immunological function in patients is shown by the increased activity of NK cells observed in both in vitro and in vivo investigations. In addition, researchers had shown earlier, in vivo, in the humanized-BLT mouse model that the consumption of AJ2 probiotic bacteria improved NK cell activity and linked with the reduction or disappearance of oral and pancreatic cancers [134]. The primary effect of LAB on NK cells is the elevation of their cytotoxic activity and, therefore, antitumor responses, as opposed to the rise in NK cell count. In accordance with this, a number of human investigations have shown that LcS supplementation increases NK cell function, with in vivo studies revealing that this is due to IL-12 [135]. This information suggests that certain LAB initiate NK interactions effectively, which improves the cytolytic capacity of NK cells in mucosal immunity.
3.3. Probiotic-Fermented-Product-Mediated Modulation of Neutrophils
Neutrophils are crucial for innate immunity and pathogen identification and response. They are short-lived myeloid cells that readily combat infection by means of oxidative-dependent generation (ROS), also known as oxidative burst, and phagocytic clearance; both of which are assisted by neutrophil extracellular traps (NETs). Probiotic bacteria on neutrophils are likely to target its effector functions: ROS production, phagocytosis, NET formation, hydrolytic enzyme activity, chemokine-mediated recruitment, and inflammatory cytokine secretion; thus, having a profound effect on neutrophil-mediated responses associated with acute infection and chronic infection/immunopathogenesis. L. rhamnosus GG has also been demonstrated to inhibit NET formation and, as a result [136], caused decreased ROS generation and avoidance of tissue death by chronic inflammation, with the reduction of ROS production indicating an inhibitory impact on NF-κB.
Additionally, it was shown that cell-wall extract of L. gasseri ATC33323 upregulated the expression of TNF-α, IL-1β, MIP-1α, and MCP-1 in a Sprague-Dawley rat model of sepsis [137]. L. brevis enriched with selenium nanoparticles was found to be capable of reducing liver metastasis in a metastatic mouse (BALB/c) breast cancer model. The study is of relevance to neutrophils since the immune responses are in the form of elevated IFN-γ and IL-17 cytokine levels and enhancement of NK cell activity [138]. Recently, Li et al. reported that oral administration of microbial metabolite butyrate markedly ameliorated mucosal inflammation in DSS-induced murine colitis through inhibition of neutrophil-associated immune responses [139]. Further, in vitro studies confirmed that butyrate suppressed neutrophil migration and formation of NETs from both CD and UC patients. B. animalis MB5 and L. rhamnosus GG strains have been shown to suppress enterotoxigenic E. coli K88 induction of the NFκB-dependent inflammatory mediators TNFα, IL-1β, IL-8, Gro-α, and ENAP-78, resulting in a corresponding inhibition of neutrophil transmigration [140].
3.4. Probiotic-Fermented-Product-Mediated Modulation of Cytokines and Immunoglobulin
Following the ingestion of fermented milks, several LAB strains have been shown to enhance both innate and adaptive immunity in animal models. Probiotic-fermented products are being studied for their immunomodulatory properties in a number of animal and human clinical studies. The effects of LAB depend on the strain, and many lactobacilli work on payer patches to improve IgA synthesis, phagocytosis, and anti-inflammatory and anti-allergic properties by stopping the production of cytokines and immunoglobulin E (IgE) [141]. In a mouse colon cancer model, yogurt intake reduces tumor development by a reduction in inflammatory response, an increase in IL-10-secreting cells, apoptosis, and a decrease in procarcinogen enzymes [142].
Yogurt administration to DMH-injected mice causes a reduction in IgG and CD8 T cells while increasing IgA and CD4 T lymphocyte numbers in the lamina propria of the large intestine. Given that IgA is regarded as a crucial barrier in colonic neoplasia, the rise in IgA-secreting cells in the bowel of mice given yogurt but not IgG cells aid to control the inflammatory response. On the other hand, bioactive peptides in kefir cause macrophage activation, phagocytosis, and the generation of nitric oxide (NO) [143]. In healthy young women, regular consumption of fermented milk rich in probiotic and prebiotic bacteria showed favorable benefits on the skin by reducing dryness and on the intestines by encouraging bowel movements and cutting down on phenol synthesis by gut bacteria [144].
In a mouse model, animals administered Malleable Protein Matrix (MPM), a novel fermented whey protein-based product, demonstrated a substantial reduction in ear inflammation with minimal side effects when compared to mice given hydrocortisone. Blood polymorphonuclear cells and the quantity of myeloperoxidase in the ear showed that the MPM seemed to prevent neutrophil extravasation into tissue [145]. According to Cristofori et al., feeding mice B. breve for 9 days before generating DSS colitis lowers inflammation by altering the makeup of T cells in the distal colon by boosting T-reg-related cytokines and decreasing Th1-related cytokines [146]. In mice, oral treatment of the probiotic L. casei CRL 431 exhibited a protective effect by raising IFN-α, IL-6, and IL-10 production in the lamina propria of the small intestine and lowering TNF production [147]. Similarly, the fermented whey dairy beverage therapy significantly improved animal survival in the mouse model (70%) and reduced pathogen translocation to the liver (2/10). It also resulted in fewer lesions in the animal’s histology and provided inflammatory protection against Salmonella Typhimurium infection [148]. Furthermore, Nipa vinegar’s active ingredients, particularly the polyphenols, were found to enhance the anti-inflammatory, hypolipidemic, and weight loss effects in obese mice when compared to synthetic acetic acid vinegar, demonstrating that bacterially fermented Nipa vinegar has a favorable impact on inflammation and lipid metabolism [149]. Hunsche et al. investigated the immunological response in BALB/c mice fed milk fermented by L. casei DN-114001, L. delbrueckii subsp. Bulgaricus, or S. thermophilus over a period of 98 days. According to the results, oral consumption of fermented milks increased the number of intestinal samples with IgA+ TNF-α, interferon IFN-γ, interleukin IL-12, and interleukin (IL-10) generating cells. In the lamina propria of the gut, CD4+ and CD8+ cell populations were also increased [150]. However, mucosal areas outside of the gut lacked any alterations. Additionally, it has been shown that adherent cells derived from Peyer’s patches and peritoneal macrophages both produce more cytokines in vivo when exposed to kefir solids (which include bacteria) and liquid supernatant [151].
3.5. TLR-Mediated Immune Response and Probiotic-Fermented Products
Toll-like receptors contribute to IEC homeostasis by governing appropriate host responses to the intestinal microbes. LAB are capable of triggering TLR-2/TLR-6 signaling pathways, 16 different LAB strains of food and human origins were characterized and tested for their interactions with specific TLRs [152]. L. plantarum-purified DNA also modulates the immune response of host cells by interacting with TLRs, as reported by Kim, whose studies show that L. plantarum-purified DNA inhibits LPS-induced TNF-α production in THP-1 cells. Furthermore, L. plantarum-purified DNA blunt the expression of TLR4, TLR2, and TLR9, which induce NF-κB activation through the LPS signaling pathway, leading to pro-inflammatory cytokine upregulation [153]. Recently, Jia et al. showed that probiotic L. salivarius AR809 could attenuate the inflammatory response produced by S. aureus by elevating autophagic protein level and blocking the TLR-mediated NF-κB signaling network [154]. Supernatant from B. breve C50 strain culture (BbC50sn) induced maturation and activation of human DCs, but only after fermentation of a milk whey-based medium. BbC50sn induced a specific interleukin-10 (IL-10)-rich cytokine profile and prolonged DC survival via a TLR2 pathway [155]. Probiotic L. fermentum (MTCC-5898) exerts immunosuppressive effects in the presence of E. coli by inhibiting pro-inflammatory cytokines release and enhancing the expression of anti-inflammatory cytokines via TLR-2/TLR-4 and the NF-κB signaling pathways [156].
In summary, probiotics are bacteria found in fermented products that regulate immune responses by interacting with dendritic cells (DCs), monocytes/macrophages, and lymphocytes, resulting in an increase in IgA response [157]. Fermented foods rich in probiotics stimulate the production of antimicrobial peptides by host cells, including bacteriocins and beta-defensins, which serve as an infection defense. They also generate lactic acid and acetic acid, both of which have potent antibiotic effects on a range of Gram-negative bacteria. Probiotic-fermented items stop the colonization of harmful bacteria by blocking the mucosal attachment site or denying the bacteria food. This behavior is referred to as competitive exclusion. The integrity of the epithelial barrier is therefore changed by components generated from dairy-fermented probiotic bacteria, and tight junction protein synthesis is promoted by TLR regulation. Thus, the mucosal layer, antimicrobial peptide, secretory IgA, TLRs, and cytokines make up the overall intestinal barrier defense, all of which are crucial for maintaining the equilibrium of the gut’s immune system [158]. In addition, the complete nature and breadth of the probiotics and TLR signaling were both covered in the recently published review paper [159].
4. Probiotic-Fermented-Product-Mediated Reinforcement of Gut Barrier Function
Probiotic fermentation combines the health benefits of microorganisms with those of fermented foods [160]. Consuming probiotic bacteria via fermented milk is also a magnificent way to restore balance to the gut flora and preserve the epithelial barrier function [161]. By encouraging the production and assembly of tight junction proteins, probiotics and released metabolites improve barrier function. They also guard against disruption of tight junctions brought on by inflammatory stimuli. Additionally, they cause a variety of cell signaling pathways to be activated, which strengthens tight junctions and the barrier function [162].
4.1. Modulation of Tight Junctional Barrier Integrity
The paracellular gap is sealed by a circumferential belt made of tight junctions, which prevents macromolecules from diffusing across the epithelium [162]. Recently, Kaur et al. showed that L. rhamnosus fermented whey was important in maintaining barrier integrity in a mouse model of DSS-induced colitis. Prior to the induction of colitis, ingestion of whey fermented with the probiotic L. rhamnosus (PFW) substantially decreased the disease activity index and enhanced the histology scores. The considerably diminished levels of pro-inflammatory markers (IL-4, TNF-α, CRP, and MPO activity) and the enhanced levels of the anti-inflammatory cytokine TGF-β with IgA in the intestine upon feeding PFW appeared to prevent inflammation on colitis induction (Figure 2). This protection is mediated by the pathogen recognition receptors TLR-2, which result in the upregulation of tight junctional genes (ZO-1, occludin, and claudin-1) and the localized distribution of junctional (claudin, occludin, and ZO-1) and cytoskeleton (actin) proteins, thereby enhancing immune homeostasis and intestinal barrier integrity. In addition, PFW ingestion resulted in lower serum concentrations of the FITC-dextran marker, which directly confirmed the host gut’s better health [16]. In a similar manner, after consuming probiotic-fermented milk products, certain microorganisms (such as B. longum) become a permanent part of the human intestinal microbiota, while others, such as L. casei, exert their effects transiently as they pass through by remodeling or influencing the community of microbes that are already present [163].
Additionally, it was found that the broth prepared by fermentation of L. acidophilus in the presence of enhanced extracts from E. sativa seeds has a high concentration of glucoerucin and significantly decreased CXCL-8 expression in Caco-2 cells after EHEC infection, as well as epithelial disruption caused by EHEC infection [164]. L. casei BL23 and milk cooperate synergistically to protect the integrity of the epithelial barrier against pro-inflammatory cytokines [165]. Likewise, ingestion of L. plantarum Lp91 in the form of fermented dairy products may enhance the intestinal barrier’s function in both normal and enteric infection circumstances. Modulation of critical regulatory receptors TLR-2 and TLR-4, as well as upregulation of tight junction genes and secretory components mucin-2, -defensin-2, and cathalicidin, indicated the mechanism of protection against infection [166]. The L. paracasei 01 fermented milk beverage decreased macromolecule permeability and improved intestinal epithelial integrity that has been damaged by dextran sodium sulfate. Additionally, fermented milk beverages containing live L. paracasei 01 enhanced intestinal epithelial cell (Caco-2) proliferation, reduced lipopolysaccharide/tumor necrosis factor (TNF-α)/interferon (IFN-γ)-induced Caco-2 cell death, and raised the production of the chemokine CCL-20 [167].
The administration of B. longum subsp. longum YS108R fermented milk was shown to alleviate DSS-induced colitis through its anti-inflammatory properties, by protecting mucosal barrier integrity, and maintaining gut microbiota homeostasis. It also significantly decreased the expression of IL-6 and IL-17A and maintained the tight junction proteins, while increasing the expression of mucin2 [38]. Jeffrey et al. evaluated the immunomodulatory activity of culture supernatants obtained from milk fermented by various LAB (L. helveticus, L. casei ssp. Casei 31, L. casei subsp. rhamnosus 4008, L. acidophilus 41, L. delbrueckii subsp. bulgaricus 1208, and Streptococcus). Additionally, the effect of fermented milk supernatant by L. helveticus R389 regulated calcineurin expression and epithelial barrier strengthening [126]. The findings suggested that the supernatant of fermented milks improved gut mucosal immunity by bolstering the epithelium and nonspecific barriers as well as gut function at infection sites. This is an important signal in the network that activates the gut immune system [168].
4.2. Modulation of Gut Permeability
The gastrointestinal epithelium offers a structural and immunological barrier against the broad spectrum of noxious and immunogenic substances present in the gut lumen. Compromised intestinal mucosal integrity and breakdown of gastrointestinal mucosal barrier function, the condition generally referred to as “Leaky Gut Syndrome” [169]. Lactobacillus helveticus ASCC 511 (LH511)-fermented milk enriched with citrulline enhances the intestinal epithelial barrier function and inflammatory response in IPEC-J2 cells caused by pathogenic E. coli. It restored the transepithelial electrical resistance (TEER) and regulated the expression and distribution of tight junction (TJ) proteins (zonula occluden-1 (ZO-1), occludin, and claudin-1), toll-like receptors (TLRs) (TLR2 and TLR4), and negative regulators of the TLR signaling pathway (A20 and IRAK-M) [170].
Consumption of L. plantarum Lp91 in the form of fermented dairy food could increase the functioning of the intestinal barrier in normal health as well as enteric infection conditions. Protection against infection was demonstrated by the modulation of key regulatory receptors TLR-2, TLR-4 and by upregulating the expression of the tight junction genes along with secretory components, mucin-2, β-defensin-2, and maintaining gut permeability [166]. L. paracasei PS23-fermented milk (PS23 FM) and its heat-killed counterpart (HK PS23 FM) could protect or reverse the increased epithelial permeability by strengthening the epithelial barrier function in vitro by increasing transepithelial electrical resistance (TEER). In vivo analysis of the regulation of intestinal physiology demonstrated that low-dose L. paracasei PS23-fermented leads to upregulation of short-chain fatty acids which ameliorated DSS-induced colitis. This anti-colitis effect may be exerted by deactivating the inflammatory cascade and strengthening the tight junctions that cause the decrease in gut leakage [171]. Milk fermented with Lactococcus lactis ssp. cremoris JFR1 resulted in increased transepithelial electrical resistance, which remained constant for the duration of infection (up to 3 h), illustrating a protective effect on Salmonella invasion of intestinal epithelial cell cultures [172].
Moreover, L. casei BL23 and milk work synergistically to prevent damage to epithelial barrier integrity reduced losses to transepithelial electrical resistance (TEER) induced by pro-inflammatory cytokines [173]. Pretreatment of Caco-2 cell monolayers with L. plantarum significantly attenuates phorbol ester-induced redistribution of ZO-1 and occludin from the intercellular junctions and the increase in permeability. The surface layer lattice, being the cell’s outermost structure, is typically regarded to be the first bacterial component to interface directly with the intestinal epithelium. SLPs may preserve the intestinal barrier by altering F-actin distribution and modifying tight junctional proteins at the mRNA and protein levels [174]. Previous research has identified that in combination with fermented and probiotics foods, diet containing fruits, vegetables, and whole grains are essential modulators of gut microbes due to their high fiber, phenolic chemical, and prebiotic content [175]. Additionally, diets that include fermentable fibers and resistant starches promote gut fermentation and SCFA (short chain fatty acid) synthesis. Increased SCFA synthesis, in turn, would improve epithelial protection by strengthening the barrier through increased TJ protein and TEER, as well as reduced permeability and bacterial translocation [176,177]. Therefore, probiotic-fermented foods are well positioned in the market because they have strong consumer acceptability. As a result, research into this kind of product is attractive since it shows long-term promise. However, well-designed and relevant clinical trials are needed to find out more about how probiotics affect the gut-immune system.
5. Conclusions
Fermented functional foods are those that may provide extra health benefits beyond the nutrients they already contain. The classification of dairy-fermented products as functional foods represents a large potential for the dairy industry. Studies have shown that fermented dairy foods have a beneficial impact on gastrointestinal health and have the ability to improve the immune system. These benefits have been known for a long time. The frequency of gut disorders has grown in recent years due to the Western lifestyle and the eating habits linked with commercial items, which promote gut permeability and antigen transcytosis and produce inflammation, ulceration, and apoptosis. Milk is rich in LAB and bifidobacteria, which makes it an excellent environment for the growth of probiotic bacteria. This is one of the reasons why milk is used to create functional meals. The consumption of fermented dairy products has the potential to improve the health of the body in a range of diseases, including diarrhea, obesity, high cholesterol, cardiovascular health, and others. Fermentation also increases the amount of helpful bioactive peptides, vitamins, and other elements generated by bacteria, which are all beneficial to the body’s overall health. Probioactives are bioactive compounds related with the food matrix and probiotic microorganisms that are present in foods. The administration of fermented dairy foods with or without probiotic bacteria is essential for human health by preventing or treating inflammatory diseases, as proven by a range of strains, according to preclinical and clinical studies that promote health. As a result, fermented dairy foods have the potential to evolve into a “functional food package” with extended shelf life and sensory qualities for the benefit of humankind as a whole.
Author Contributions
Conceptualization, S.A.A.; writing—original draft preparation, H.K.; writing—review and editing, G.K.; project administration and funding acquisition, S.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would also like to thank the Indian Council of Agriculture Research-National Dairy Research Institute (NDRI) Director, India, for providing the necessary facilities to carry out research work. All the figures are prepared with the help of purchased Bio render online tool.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Górska, W.H.; Rejman, K.L.W.; Czeczotko, M. Milk and dairy products and their nutritional contribution to the average polish diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef] [PubMed]
- Cena, H.; Calder, P.C. Defining a healthy diet: Evidence for the role of contemporary dietary patterns in health and disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [PubMed]
- Deledda, A.; Annunziata, G.; Tenore, G.C.; Palmas, V.; Manzin, A.; Velluzzi, F. Diet-derived antioxidants and their role in inflammation, obesity and gut microbiota modulation. Antioxidants 2021, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- García, B.M.; Moreno, F.J.; Alférez, M.J.; Díaz-Castro, J.; López, A.I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods. 2020, 72, 104059. [Google Scholar] [CrossRef]
- Savaiano, D.A.; Hutkins, R.W. Yogurt, cultured fermented milk, and health: A systematic review. Nutr. Rev. 2021, 79, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Singh, P.; Nataraj, B.H.; Kokkiligadda, A.; Naithani, H.; Ali, S.A.; Nagpal, R. Fostering next-generation probiotics in human gut by targeted dietary modulation: An emerging perspective. Int. Food Res. J. 2021, 150, 110716. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Auestad, N.; Layman, D.K. Dairy bioactive proteins and peptides: A narrative review. Nutr Rev. 2021, 79, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ali, S.A.; Singh, S.K.; Bhushan, V.; Mathur, M.; Jamwal, S.; Kumar, S. Antimicrobial peptides in farm animals: An updated review on its diversity, function, modes of action and therapeutic prospects. Vet. Sci. 2020, 7, 206. [Google Scholar] [CrossRef] [PubMed]
- Solanki, S.S.; Singh, P.; Kashyap, P.; Sansi, M.S.; Ali, S.A. Promising role of defensins peptides as therapeutics to combat against viral infection. Microb. Pathog. 2021, 155, 104930. [Google Scholar] [CrossRef] [PubMed]
- Fortea, M.; Albert, B.M.; Abril, G.M.; Ganda Mall, J.P.; Serra-Ruiz, X.; Henao, P.A.; Expósito, E.; González-Castro, A.M.; Guagnozzi, D.; Lobo, B.; et al. Present and future therapeutic approaches to barrier dysfunction. Front. Nutr. 2021, 8, 718093. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Ali, S.A.; Bathla, S.; Rawat, P.; Vohra, V.; Kumar, S.; Mohanty, A.K. High-resolution mass spectrometer–based ultra-deep profile of milk whey proteome in Indian Zebu (Sahiwal) cattle. Front. Nutr. 2020, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.M.; Kang, E.A.; Park, J.M.; Oh, J.Y.; Lee, D.Y.; Choi, S.H.; Hahm, K.B. Dietary intake of fermented kimchi prevented colitis-associated cancer. J. Clin. Biochem. Nutr. 2020, 20–77. [Google Scholar] [CrossRef]
- Kaur, H.; Gupta, T.; Kapila, S.; Kapila, R. Protective effects of potential probiotic Lactobacillus rhamnosus (MTCC-5897) fermented whey on reinforcement of intestinal epithelial barrier function in a colitis-induced murine model. Food. Funct. 2021, 12, 6102–6116. [Google Scholar] [CrossRef]
- Matsuoka, K.; Uemura, Y.; Kanai, T.; Kunisaki, R.; Suzuki, Y.; Yokoyama, K.; Yoshimura, N.; Hibi, T. Efficacy of Bifidobacterium breve fermented milk in maintaining remission of ulcerative colitis. Dig. Dis. Sci. 2018, 63, 1910–1919. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, R.K.; Malhotra, S.; Pothuraju, R.; Shandilya, U.K. Lactobacillus rhamnosus NCDC17 ameliorates type-2 diabetes by improving gut function, oxidative stress and inflammation in high-fat-diet fed and streptozotocintreated rats. Benef. Microbes 2017, 8, 243–255. [Google Scholar] [CrossRef]
- Rong, J.; Zheng, H.; Liu, M.; Hu, X.; Wang, T.; Zhang, X.; Jin, F.; Wang, L. Probiotic and anti-inflammatory attributes of an isolate Lactobacillus helveticus NS8 from Mongolian fermented koumiss. BMC Microbiol. 2015, 15, 196. [Google Scholar] [CrossRef]
- Beltrán-Barrientos, L.M.; González-Córdova, A.F.; Hernández-Mendoza, A.; Torres-Inguanzo, E.H.; Astiazarán-García, H.; Esparza-Romero, J.; Vallejo-Cordoba, B. Randomized double-blind controlled clinical trial of the blood pressure–lowering effect of fermented milk with Lactococcus lactis: A pilot study. Int. J. Dairy Sci. 2018, 101, 2819–2825. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, J.L.; Serrano-Amatriain, C.; Ledesma-Amaro, R.; Jiménez, A. Formation of folates by microorganisms: Towards the biotechnological production of this vitamin. Appl. Microbiol. Biotechnol. 2018, 102, 8613–8620. [Google Scholar] [CrossRef] [PubMed]
- Saubade, F.; Hemery, Y.M.; Rochette, I.; Guyot, J.P.; Humblot, C. Influence of fermentation and other processing steps on the folate content of a traditional African cereal-based fermented food. Int. J. Food Microbiol. 2018, 266, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Thumthanaruk, B.; Shetty, K. Changes in physico-chemical, astringency, volatile compounds and antioxidant activity of fresh and concentrated cashew apple juice fermented with Lactobacillus plantarum. J. Food Sci. Technol. 2018, 55, 3979–3990. [Google Scholar] [CrossRef] [PubMed]
- Tarvainen, M.; Fabritius, M.; Yang, B. Determination of vitamin K composition of fermented food. Food Chem. 2019, 275, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Merenstein, D.; Murphy, M.; Fokar, A.; Hernandez, R.K.; Park, H.; Nsouli, H.; Sanders, M.E.; Davis, B.A.; Niborski, V.; Tondu, F.; et al. Use of a fermented dairy probiotic drink containing Lactobacillus casei (DN-114 001) to decrease the rate of illness in kids: The DRINK study A patient-oriented, double-blind, cluster-randomized, placebo-controlled, clinical trial. Eur. J. Clin. Nutr. 2010, 64, 669–677. [Google Scholar] [CrossRef]
- Perna, A.; Intaglietta, I.; Simonetti, A.; Gambacorta, E. Donkey milk for manufacture of novel functional fermented beverages. J. Food Sci. 2015, 80, S1352–S1359. [Google Scholar] [CrossRef]
- Ho, S.W.; El-Nezami, H.; Shah, N.P. The protective effects of enriched citrulline fermented milk with Lactobacillus helveticus on the intestinal epithelium integrity against Escherichia coli infection. Sci. Rep. 2020, 10, 499. [Google Scholar] [CrossRef] [PubMed]
- Sivasankari, R.; Hemalatha, G.; Kanchana, S.; Umamaheswari, T. Standardization and evaluation of probiotic shrikhand. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 41–47. [Google Scholar] [CrossRef]
- Sevencan, N.O.; Isler, M.; Kapucuoglu, F.N.; Senol, A.; Kayhan, B.; Kiztanir, S.; Kockar, M.C. Dose-dependent effects of kefir on colitis induced by trinitrobenzene sulfonic acid in rats. Food Sci. Nutr. 2019, 7, 3110–3118. [Google Scholar] [CrossRef]
- Sánchez, E.; Nieto, J.C.; Vidal, S.; Santiago, A.; Martinez, X.; Sancho, F.J.; Sancho-Bru, P.; Mirelis, B.; Corominola, H.; Juárez, C.; et al. Fermented milk containing Lactobacillus paracasei subsp. paracasei CNCM I-1518 reduces bacterial translocation in rats treated with carbon tetrachloride. Sci. Rep. 2017, 7, 45712. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.; Ali, S.A.; Behare, P.; Kaul, G. Dietary intake of probiotic fermented milk benefits the gut and reproductive health in mice fed with an obesogenic diet. Food Funct. 2022, 13, 737–752. [Google Scholar] [CrossRef]
- Yadav, R.; Vij, R.; Kapila, S.; Khan, S.H.; Kumar, N.; Meena, S.; Kapila, R. Milk fermented with probiotic strains Lactobacillus rhamnosus MTCC: 5957 and Lactobacillus rhamnosus MTCC: 5897 ameliorates the diet-induced hypercholesterolemia in rats. Ann. Microbiol. 2019, 69, 483–494. [Google Scholar] [CrossRef]
- Companys, J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Solà, R.; Pedret, A.; Valls, R.M. Fermented dairy products, probiotic supplementation, and cardiometabolic diseases: A systematic review and meta-analysis. Adv. Nutr. 2020, 11, 834–863. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Chen, H.; Chen, H.; Ding, W. Effects of fermentation with Lactobacillus rhamnosus GG on product quality and fatty acids of goat milk yogurt. Int. J. Dairy Sci. 2016, 99, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Patrignani, F.; Siroli, L.; Parolin, C.; Serrazanetti, D.I.; Vitali, B.; Lanciotti, R. Use of Lactobacillus crispatus to produce a probiotic cheese as potential gender food for preventing gynaecological infections. PLoS ONE 2019, 14, 0208906. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.; Kim, W. Cream cheese-derived Lactococcus chungangensis CAU 28 modulates the gut microbiota and alleviates atopic dermatitis in BALB/c mice. Sci. Rep. 2019, 9, 446. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Hayes, J.J.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Protection of candidate probiotic lactobacilli by Cheddar cheese matrix during simulated gastrointestinal digestion. J. Funct. Foods 2022, 92, 105042. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Ross, R.P.; Stanton, C.; Zhang, H.; Zhao, J.; Chen, W. Bifidobacterium longum subsp. longum YS108R fermented milk alleviates DSS induced colitis via anti-inflammation, mucosal barrier maintenance and gut microbiota modulation. J. Funct. Foods 2020, 73, 104153. [Google Scholar] [CrossRef]
- Lee, K.Y.; Tsai, Y.C.; Wang, S.Y.; Chen, Y.P.; Chen, M.J. Coculture Strategy for Developing Lactobacillus paracasei PS23 Fermented Milk with Anti-Colitis Effect. Foods 2021, 10, 2337. [Google Scholar] [CrossRef]
- Sumi, K.; Osada, K.; Sakuda, M.; Ashida, K.; Nakazato, K. Fermented milk retains beneficial effects on skeletal muscle protein anabolism after processing by centrifugation and supernatant removal. Int. J. Dairy Sci. 2021, 104, 1336–1350. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Han, X.; Cen, S.; Duan, H.; Feng, S.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; et al. Beneficial effect of GABA-rich fermented milk on insomnia involving regulation of gut microbiota. Microbiol. Res. 2020, 233, 126409. [Google Scholar] [CrossRef]
- Tavakoli, M.; Najafi, M.B.H.; Mohebbi, M. Effect of the milk fat content and starter culture selection on proteolysis and antioxidant activity of probiotic yogurt. Heliyon 2019, 5, 01204. [Google Scholar] [CrossRef]
- Minj, S.; Anand, S. Whey proteins and its derivatives: Bioactivity, functionality, and current applications. Dairy 2020, 1, 233–258. [Google Scholar] [CrossRef]
- Wolber, F.M.; Broomfield, A.M.; Fray, L.; Cross, M.L.; Dey, D. Supplemental dietary whey protein concentrate reduces rotavirus-induced disease symptoms in suckling mice. J. Nutr. 2005, 135, 1470–1474. [Google Scholar] [CrossRef]
- Vasconcelos, Q.D.J.S.; Bachur, T.P.R.; Aragão, G.F. Whey protein supplementation and its potentially adverse effects on health: A systematic review. Appl. Physiol. Nutr. Metab. 2021, 46, 27–33. [Google Scholar] [CrossRef]
- Lorentz, A.; Müller, L. Probiotics in the Treatment of Inflammatory Bowel Disease in Adulthood: A Systematic Review. Gastrointestin. Liver Dis. 2022, 31, 74–84. [Google Scholar] [CrossRef]
- Karimi, R.; Mortazavian, A.M.; Amiri, R.A. Selective enumeration of probiotic microorganisms in cheese. Food Microbiol. 2012, 29, 1–9. [Google Scholar] [CrossRef]
- Januś, E.; Sablik, P.; Jakubowska, M.; Wróbel, K. Microbiological, sensory, and physicochemical quality of curd cheeses originating from direct sales. Vet. World 2021, 14, 3038. [Google Scholar] [CrossRef]
- Tang, H.; Ma, H.; Hou, Q.; Li, W.; Xu, H.; Liu, W.; Sun, Z.; Haobisi, H.; Menghe, B. Profiling of koumiss microbiota and organic acids and their effects on koumiss taste. BMC Microbiol. 2020, 20, 85. [Google Scholar] [CrossRef]
- Afzaal, M.; Saeed, F.; Anjum, F.; Waris, N.; Husaain, M.; Ikram, A.; Ateeq, H.; Muhammad Anjum, F.; Suleria, H. Nutritional and ethnomedicinal scenario of koumiss: A concurrent review. Food Sci. Nutr. 2021, 9, 6421–6428. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, C.; Xilin, T.; Ji, M.; Meng, X.; Zhao, Y.; Siqin, B.; Zhang, N.; Li, M. Effects of Koumiss on Intestinal Immune Modulation in Immunosuppressed Rats. Front. Nutr. 2022, 9, 765499. [Google Scholar] [CrossRef]
- Tiss, M.; Souiy, Z.; ben Abdeljelil, N.; Njima, M.; Achour, L.; Hamden, K. Fermented soy milk prepared using kefir grains prevents and ameliorates obesity, type 2 diabetes, hyperlipidemia and liver-kidney toxicities in HFFD-rats. J. Funct. Foods 2020, 67, 103869. [Google Scholar]
- Mohan, J.; Ali, S.A.; Suvartan, R.; Kapila, S.; Sharma, R.; Tomar, S.K.; Yadav, H. Bioavailability of biotransformed zinc enriched ahi in wistar rats. Int. J. Probiotics Prebiotics 2018, 13, 45. [Google Scholar]
- Gawande, K.; Kolhekar, M.; Kumari, M.; Kapila, S.; Sharma, P.; Ali, S.A.; Behare, P.V. Lactic acid bacteria based purified exopolysaccharide showed viscofying and hypercholesterolemic capabilites. Food Hydrocoll. 2021, 1, 100042. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Dardis, C.; Garrote, G.L.; Abraham, A.G. Health-promoting properties of Lacticaseibacillus paracasei: A focus on kefir isolates and exopolysaccharide-producing strains. Foods 2021, 10, 2239. [Google Scholar] [CrossRef]
- Perazza, L.R.; Daniel, N.; Dubois, M.J.; Pilon, G.; Varin, T.V.; Blais, M.; Martinez Gonzales, J.L.; Bouchard, M.; Asselin, C.; Lessard, M.; et al. Distinct effects of milk-derived and fermented dairy protein on gut microbiota and cardiometabolic markers in diet-induced obese mice. J. Nutr. 2020, 150, 2673–2686. [Google Scholar] [CrossRef]
- Utz, V.E.M.; Perdigón, G.; de LeBlanc, A.D.M. Oral administration of milk fermented by Lactobacillus casei CRL431 was able to decrease metastasis from breast cancer in a murine model by modulating immune response locally in the lungs. J. Funct. Food. 2019, 54, 263–270. [Google Scholar]
- Gupta, C.; Prakash, D. Therapeutic potential of milk whey. Beverages 2017, 3, 31. [Google Scholar] [CrossRef]
- Mehra, R.; Kumar, H.; Kumar, N.; Ranvir, S.; Jana, A.; Buttar, H.S.; Telessy, I.G.; Awuchi, C.G.; Okpala, C.O.R.; Korzeniowska, M. Whey proteins processing and emergent derivatives: An insight perspective from constituents, bioactivities, functionalities to therapeutic applications. J. Funct. Foods 2021, 87, 104760. [Google Scholar]
- Hering, N.A.; Andres, S.; Fromm, A.; van Tol, E.A.; Amasheh, M.; Mankertz, J.; Fromm, M.; Schulzke, J.D. Transforming growth factor-β, a whey protein component, strengthens the intestinal barrier by upregulating claudin-4 in HT-29/B6 cells. J. Nutr. 2011, 141, 783–789. [Google Scholar] [CrossRef]
- Araujo, D.F.; Guerra, G.C.; Pintado, M.M.E.; Sousa, Y.R.; Algieri, F.; Rodriguez-Nogales, A.; Araujo Jr, R.F.; Galvez, J.; Queiroga, R.D.C.R.; Rodriguez-Cabezas, M.E. Intestinal anti-inflammatory effects of goat whey on DNBS-induced colitis in mice. PLoS ONE 2017, 12, e0185382. [Google Scholar] [CrossRef] [PubMed]
- Rusu, E.; Enache, G.; Cursaru, R.; Alexescu, A.; Radu, R.; Onila, O.; Cavallioti, T.; Rusu, F.; Posea, M.; Jinga, M.; et al. Prebiotics and probiotics in atopic dermatitis. Exp. Ther. Med. 2019, 18, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Sangild, P.T.; Li, Y.; Bering, S.B.; Chatterton, D.E. Processing of whey modulates proliferative and immune functions in intestinal epithelial cells. Int. J. Dairy Sci. 2016, 99, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Chun, S.H.; Lee, H.A.; Lee, K.B.; Kim, S.H.; Park, K.Y.; Lee, K.W. Effects of glycated whey protein concentrate on pro-inflammatory cytokine expression and phagocytic activity in RAW264. 7 macrophages. Biol. Pharm. Bull. 2016, 39, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanifer, M.L.; Guo, C.; Doldan, P.; Boulant, S. Importance of type I and III interferons at respiratory and intestinal barrier surfaces. Front. Immunol. 2020, 11, 608645. [Google Scholar] [CrossRef]
- Rusu, D.; Drouin, R.; Pouliot, Y.; Gauthier, S.; Poubelle, P.E. A bovine whey protein extract stimulates human neutrophils to generate bioactive IL-1Ra through a NF-κB-and MAPK-dependent mechanism. J. Nutr. 2010, 140, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, J.E.; Anderson, R.C.; Bassett, S.A.; Lloyd-West, C.M.; Haggarty, N.W.; Roy, N.C. Influence of bovine whey protein concentrate and hydrolysate preparation methods on motility in the isolated rat distal colon. Nutrients 2016, 8, 809. [Google Scholar] [CrossRef]
- Gupta, T.; Kaur, H.; Kapila, S.; Kapila, R. Potential probiotic Lacticaseibacillus rhamnosus MTCC-5897 attenuates Escherichia coli induced inflammatory response in intestinal cells. Arch. Microbiol. 2021, 203, 5703–5713. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-promoting role of lactiplantibacillus plantarum isolated from fermented foods. Microorganisms 2021, 9, 349. [Google Scholar]
- Wu, D.; Lewis, E.D.; Pae, M.; Meydani, S.N. Nutritional modulation of immune function: Analysis of evidence, mechanisms, and clinical relevance. Front. Immunol. 2019, 9, 3160. [Google Scholar] [CrossRef]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Widyastuti, Y.; Febrisiantosa, A.; Tidona, F. Health-Promoting Properties of Lactobacilli in Fermented Dairy Products. Front. Microbiol. 2021, 12, 1189. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.M.; Ali, S.A.; Ceciliani, F.; Eckersall, P.D.; Hernandez-Castellano, L.E.; Han, R.; Zachut, M. Domestic animal proteomics in the 21st century: A global retrospective and viewpoint analysis. J. Proteom. 2021, 241, 104220. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Ali, S.A.; Kumar, S.; Mohanty, A.K.; Behare, P. Label-free quantitative proteomic analysis of Lactobacillus fermentum NCDC 400 during bile salt exposure. J. Proteom. 2017, 167, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Pragya, P.; Kaur, G.; Ali, S.A.; Bhatla, S.; Rawat, P.; Lule, V.; Behare, P. High-resolution mass spectrometry-based global proteomic analysis of probiotic strains Lactobacillus fermentum NCDC 400 and RS2. J. Proteom. 2017, 152, 121–130. [Google Scholar] [CrossRef]
- Ali, S.A.; Kumar, S.; Mohanty, A.K.; Behare, P. Draft genome sequence of Lactobacillus fermentum NCDC 400, isolated from a traditional Indian dairy product. Genome Announc. 2018, 6, e01492-17. [Google Scholar]
- Ali, S.A.; Singh, P.; Tomar, S.K.; Mohanty, A.K.; Behare, P. Proteomics fingerprints of systemic mechanisms of adaptation to bile in Lactobacillus fermentum. J. Proteom. 2020, 213, 103600. [Google Scholar] [CrossRef]
- Behare, P.V.; Ali, S.A.; McAuliffe, O. Draft genome sequences of Fructobacillus fructosus DPC 7238 and Leuconostoc mesenteroides DPC 7261, mannitol-producing organisms isolated from fructose-rich honeybee-resident flowers on an Irish farm. Microbiol. Resour. Announc. 2020, 9, e01297-20. [Google Scholar] [CrossRef]
- Ali, S.A.; Singh, A.K.; Tomar, S.K.; Behare, P. Genome Sequence of Lacticaseibacillus rhamnosus Strain NCDC610, Isolated from a Traditional Cereal-Based Fermented Milk Product (Raabadi). Microbiol. Resour. Announc. 2021, 10, e00672-21. [Google Scholar]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- Hickey, C.D.; Sheehan, J.J.; Wilkinson, M.G.; Auty, M.A. Growth and location of bacterial colonies within dairy foods using microscopy techniques: A review. Front. Microbiol. 2015, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Nyanzi, R.; Jooste, P.J.; Buys, E.M. Invited review: Probiotic yogurt quality criteria, regulatory framework, clinical evidence, and analytical aspects. Int. J. Dairy Sci. 2021, 104, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in food systems: Significance and emerging strategies towards improved viability and delivery of enhanced beneficial value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.; Cui, C.; Ruan, Z. Fermentation-enabled wellness foods: A fresh perspective. Food Sci. Hum. Wellness 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Mahmoudi, R.; Tajik, H.; Ehsani, A.; Zare, P. Physicochemical and hygienic effects of Lactobacillus acidophilus in Iranian white cheese. Vet. Res. Forum 2012, 3, 193. [Google Scholar]
- Wendel, U. Assessing Viability and Stress Tolerance of Probiotics-A Review. Front. Microbiol 2021, 12, 818468. [Google Scholar] [CrossRef]
- Damián, M.R.; Cortes-Perez, N.G.; Quintana, E.T.; Ortiz-Moreno, A.; Garfias Noguez, C.; Cruceño-Casarrubias, C.E.; Sánchez Pardo, M.E.; Bermúdez-Humarán, L.G. Functional Foods, Nutraceuticals and Probiotics: A Focus on Human Health. Microorganisms 2022, 10, 1065. [Google Scholar] [CrossRef]
- Panicker, A.S.; Ali, S.A.; Anand, S.; Panjagari, N.R.; Kumar, S.; Mohanty, A.K.; Behare, P.V. Evaluation of some in vitro probiotic properties of Lactobacillus fermentum Strains. J. Food Sci. Technol. 2018, 55, 2801–2807. [Google Scholar] [CrossRef]
- Koirala, S.; Anal, A.K. Probiotics-based foods and beverages as future foods and their overall safety and regulatory claims. J. Food Agric. Environ. 2021, 3, 100013. [Google Scholar] [CrossRef]
- Hendler, R.; Zhang, Y. Probiotics in the treatment of colorectal cancer. Medicines 2018, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tran, D.Q.; Rhoads, J.M. Probiotics in disease prevention and treatment. J. Clin. Pharmacol. 2018, 58, S164–S179. [Google Scholar] [CrossRef]
- Begum, P.S.; Madhavi, G.; Rajagopal, S.; Viswanath, B.; Razak, M.A.; Venkataratnamma, V. Probiotics as functional foods: Potential effects on human health and its impact on neurological diseases. Int. J. Nutr. Pharmacol. Neurol. Dis. 2017, 7, 23. [Google Scholar]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.M.; Mota, J.F. Gut microbiota, probiotics and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Sui, H.; Qi, C.; Li, Q.; Zhang, J.; Wu, S.F.; Mei, M.Z.; Lu, Y.Y.; Wan, Y.T.; Chang, H.; et al. Ursolic acid inhibits proliferation and reverses drug resistance of ovarian cancer stem cells by downregulating ABCG2 through suppressing the expression of hypoxia-inducible factor-1α in vitro. Oncol. Rep. 2016, 36, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Alihosseini, N.; Moahboob, S.A.; Farrin, N.; Mobasseri, M.; Taghizadeh, A.; Ostadrahimi, A.R. Effect of probiotic fermented milk (kefir) on serum level of insulin and homocysteine in type 2 diabetes patients. Acta. Endocrinol. 2017, 13, 431. [Google Scholar] [CrossRef] [PubMed]
- Widodo, W.; Harsita, P.A.; Sukarno, A.S.; Nurrochmad, A. Antidiabetic effect of milk fermented using intestinal probiotics. Food Sci. Nutr. 2019, 49, 1063–1074. [Google Scholar] [CrossRef]
- Yadav, R.; Dey, D.K.; Vij, R.; Meena, S.; Kapila, R.; Kapila, S. Evaluation of anti-diabetic attributes of Lactobacillus rhamnosus MTCC: 5957, Lactobacillus rhamnosus MTCC: 5897 and Lactobacillus fermentum MTCC: 5898 in streptozotocin induced diabetic rats. Microb. Pathog. 2018, 125, 454–462. [Google Scholar] [CrossRef]
- Misselwitz, B.; Butter, M.; Verbeke, K.; Fox, M.R. Update on lactose malabsorption and intolerance: Pathogenesis, diagnosis and clinical management. Gut 2019, 68, 2080–2091. [Google Scholar] [CrossRef] [Green Version]
- Szilagyi, A.; Ishayek, N. Lactose intolerance, dairy avoidance, and treatment options. Nutrients 2018, 10, 1994. [Google Scholar] [CrossRef]
- Facioni, M.S.; Raspini, B.; Pivari, F.; Dogliotti, E.; Cena, H. Nutritional management of lactose intolerance: The importance of diet and food labelling. J. Transl. Med. 2020, 18, 260. [Google Scholar] [CrossRef] [PubMed]
- Forsgård, R.A. Lactose digestion in humans: Intestinal lactase appears to be constitutive whereas the colonic microbiome is adaptable. Am. J. Clin. Nutr. 2019, 110, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, U.; Malida, R.; Panda, R.; Halder, T.; Roymahapatra, G. Variety of yogurt and its health aspects—A brief review. Int. J. Innov. Pract. Appl. Res. 2017, 7, 56–66. [Google Scholar]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Dennis-Wall, J.C.; Culpepper, T.; Nieves Jr, C.; Rowe, C.C.; Burns, A.M.; Rusch, C.T.; Federico, A.; Ukhanova, M.; Waugh, S.; Mai, V.; et al. Probiotics (Lactobacillus gasseri KS-13, Bifidobacterium bifidum G9-1, and Bifidobacterium longum MM-2) improve rhinoconjunctivitis-specific quality of life in individuals with seasonal allergies: A double-blind, placebo-controlled, randomized trial. Am. J. Clin. Nutr. 2017, 105, 758–767. [Google Scholar] [CrossRef]
- Liu, M.Y.; Yang, Z.Y.; Dai, W.K.; Huang, J.Q.; Li, Y.H.; Zhang, J.; Qiu, C.Z.; Wei, C.; Zhou, Q.; Sun, X.; et al. Protective effect of Bifidobacterium infantis CGMCC313-2 on ovalbumin-induced airway asthma and β-lactoglobulin-induced intestinal food allergy mouse models. World, J. Gastroenterol. 2017, 23, 2149. [Google Scholar] [CrossRef]
- Saliganti, V.; Kapila, R.; Sharma, R.; Kapila, S. Feeding probiotic Lactobacillus rhamnosus (MTCC 5897) fermented milk to suckling mothers alleviates ovalbumin-induced allergic sensitisation in mice offspring. Br. J. Nutr. 2015, 114, 1168–1179. [Google Scholar] [CrossRef]
- Li Wang, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-derived antioxidant polysaccharides and their pharmacological potential in neurodegenerative diseases. Nutrients 2017, 9, 778. [Google Scholar]
- Wang, Y.; Wu, Y.; Wang, Y.; Fu, A.; Gong, L.; Li, W.; Li, Y. Bacillus amyloliquefaciens SC06 alleviates the oxidative stress of IPEC-1 via modulating Nrf2/Keap1 signaling pathway and decreasing ROS production. Appl. Microbiol. Biotechnol. 2017, 101, 3015–3026. [Google Scholar] [CrossRef]
- Nayik, G.A.; Jagdale, Y.D.; Gaikwad, S.A.; Devkatte, A.N.; Dar, A.H.; Dezmirean, D.S.; Bobis, O.; Ranjha, M.M.A.; Ansari, M.J.; Hemeg, H.A.; et al. Recent Insights into Processing Approaches and Potential Health Benefits of Goat Milk and Its Products: A Review. Front. Nutr. 2021, 8, 789117. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The gut microbiome as a major regulator of the gut-skin axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, A.D.; Leoni, J.; Paz, M.L.; González Maglio, D.H. Lipoteichoic Acid from Lacticaseibacillus rhamnosus GG Modulates Dendritic Cells and T Cells in the Gut. Nutrients 2022, 14, 723. [Google Scholar] [CrossRef] [PubMed]
- Khmaladze, I.; Butler, É.; Fabre, S.; Gillbro, J.M. Lactobacillus reuteri DSM 17938—A comparative study on the effect of probiotics and lysates on human skin. Exp. Dermatol. 2019, 28, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Kober, M.M.; Bowe, W.P. The effect of probiotics on immune regulation, acne, and photoaging. Int. J. Women’s Dermatol. 2015, 1, 85–89. [Google Scholar] [CrossRef]
- Bianconi, V.; Banach, M.; Pirro, M.; Panel, I.L.E. Why patients with familial hypercholesterolemia are at high cardiovascular risk? Beyond LDL-C levels. Trends Cardiovasc Med. 2021, 31, 205–215. [Google Scholar] [CrossRef]
- Jesch, E.D.; Carr, T.P. Food ingredients that inhibit cholesterol absorption. Prev. Nutr. Food Sci. 2017, 22, 67. [Google Scholar]
- Machlik, M.L.; Hopstock, L.A.; Wilsgaard, T.; Hansson, P. Associations Between Intake of Fermented Dairy Products and Blood Lipid Concentrations Are Affected by Fat Content and Dairy Matrix–The Tromsø Study: Tromsø7. Front. Nutr. 2021, 8, 773468. [Google Scholar] [CrossRef]
- Jakubczyk, D.; Leszczyńska, K.; Górska, S. The effectiveness of probiotics in the treatment of inflammatory bowel disease (IBD)—A critical review. Nutrients 2020, 12, 1973. [Google Scholar] [CrossRef]
- Sartor, R.B. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: Antibiotics, probiotics, and prebiotics. Gastroenterology 2004, 126, 1620–1633. [Google Scholar] [CrossRef]
- Dore, M.P.; Bibbò, S.; Fresi, G.; Bassotti, G.; Pes, G.M. Side effects associated with probiotic use in adult patients with inflammatory bowel disease: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2019, 11, 2913. [Google Scholar] [CrossRef]
- Gomi, A.; Yamaji, K.; Watanabe, O.; Yoshioka, M.; Miyazaki, K.; Iwama, Y.; Urita, Y. Bifidobacterium bifidum YIT 10347 fermented milk exerts beneficial effects on gastrointestinal discomfort and symptoms in healthy adults: A double-blind, randomized, placebo-controlled study. Int. J. Dairy Sci. 2018, 101, 4830–4841. [Google Scholar] [CrossRef] [PubMed]
- Eales, J.; Gibson, P.; Whorwell, P.; Kellow, J.; Yellowlees, A.; Perry, R.H.; Edwards, M.; King, S.; Wood, H.; Glanville, J. Systematic review and meta-analysis: The effects of fermented milk with Bifidobacterium lactis CNCM I-2494 and lactic acid bacteria on gastrointestinal discomfort in the general adult population. Therap. Adv. Gastroenter. 2017, 10, 74–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Kaur, H.; Gupta, T.; Kapila, S.; Kapila, R. Role of fermented dairy foods in human health. Ind. J. Dairy Sci. 2020, 73, 97–110. [Google Scholar] [CrossRef]
- Gómez-Llorente, C.; Munoz, S.; Gil, A. Role of Toll-like receptors in the development of immunotolerance mediated by probiotics. Proc. Nutr. Soc. 2010, 69, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, L.S.; Lie-Andersen, O.; Obers, A.; Crowther, M.D.; Svane, I.M.; Hansen, M. Long-term exposure to inflammation induces differential cytokine patterns and apoptosis in dendritic cells. Front. Immunol. 2019, 10, 2702. [Google Scholar] [CrossRef]
- Jeffrey, M.P.; Taggart, H.J.; Strap, J.L.; Edun, G.; Green-Johnson, J.M. Milk fermented with Lactobacillus rhamnosus R0011 induces a regulatory cytokine profile in LPS-challenged U937 and THP-1 macrophages. Curr. Res. Nutr. Food Sci. 2020, 3, 51–58. [Google Scholar] [CrossRef]
- Fernandez, E.M.; Valenti, V.; Rockel, C.; Hermann, C.; Pot, B.; Boneca, I.G.; Grangette, C. Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by NOD2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 2011, 60, 1050–1059. [Google Scholar] [CrossRef]
- Kwon, H.K.; Lee, C.G.; So, J.S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+ Foxp3+ T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar] [CrossRef]
- Miller, L.E.; Lehtoranta, L.; Lehtinen, M.J. Short-term probiotic supplementation enhances cellular immune function in healthy elderly: Systematic review and meta-analysis of controlled studies. Nutr. Res. 2019, 64, 1–8. [Google Scholar] [CrossRef]
- Gui, Q.; Wang, A.; Zhao, X.; Huang, S.; Tan, Z.; Xiao, C.; Yang, Y. Effects of probiotic supplementation on natural killer cell function in healthy elderly individuals: A meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2020, 74, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Shida, K.; Sato, T.; Iizuka, R.; Hoshi, R.; Watanabe, O.; Igarashi, T.; Miyazaki, K.; Nanno, M.; Ishikawa, F. Daily intake of fermented milk with Lactobacillus casei strain Shirota reduces the incidence and duration of upper respiratory tract infections in healthy middle-aged office workers. Eur. J. Nutr. 2017, 56, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Rowland, I.; Tuohy, K.M.; Thomas, L.V.; Yaqoob, P. Selective effects of Lactobacillus casei Shirota on T cell activation, natural killer cell activity and cytokine production. Clin. Exp. Immunol. 2010, 161, 378–388. [Google Scholar] [CrossRef]
- Ko, M.W.; Kaur, K.; Safaei, T.; Chen, W.; Sutanto, C.; Wong, P.; Jewett, A. Defective Patient NK Function Is Reversed by AJ2 Probiotic Bacteria or Addition of Allogeneic Healthy Monocytes. Cells 2022, 11, 697. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Kozlowska, A.K.; Topchyan, P.; Ko, M.W.; Ohanian, N.; Chiang, J.; Cook, J.; Maung, P.O.; Park, S.H.; Cacalano, N.; et al. Probiotic-treated super-charged NK cells efficiently clear poorly differentiated pancreatic tumors in Hu-BLT mice. Cancers 2019, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Suzuki, T.; Shimada, S.I.; Shida, K.; Nanno, M.; Okumura, K. Interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei Shirota. Clin. Exp. Immunol. 2006, 146, 109–115. [Google Scholar] [CrossRef]
- Xu, X.; Hicks, C.; Li, Y.; Su, J.; Shiloach, J.; Kaufman, J.B.; Fitz, Y.; Eichacker, P.Q.; Cui, X. Purified cell wall from the probiotic bacterium Lactobacillus gasseri activates systemic inflammation and, at higher doses, produces lethality in a rat model. Crit. Care. 2014, 18, R140. [Google Scholar] [CrossRef]
- Vong, L.; Lorentz, R.J.; Assa, A.; Glogauer, M.; Sherman, P.M. Probiotic Lactobacillus rhamnosus inhibits the formation of neutrophil extracellular traps. J. Immunol. 2014, 192, 1870–1877. [Google Scholar] [CrossRef]
- Frank, K.; Paust, S. Dynamic natural killer cell and T cell responses to influenza infection. Front. Cell. Infect. Microbiol. 2020, 10, 425. [Google Scholar] [CrossRef]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota metabolite butyrate constrains neutrophil functions and ameliorates mucosal inflammation in inflammatory bowel disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef]
- Roselli, M.; Finamore, A.; Britti, M.S.; Mengheri, E. Probiotic bacteria Bifidobacterium animalis MB5 and Lactobacillus rhamnosus GG protect intestinal Caco-2 cells from the inflammation-associated response induced by enterotoxigenic Escherichia coli K88. Br. J. Nutr. 2006, 95, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory effects of probiotics during pathogenic infections with emphasis on immune regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef] [PubMed]
- De Moreno De Leblanc, A.; Perdigón, G. The application of probiotic fermented milks in cancer and intestinal inflammation. Proc. Nutr. Soc. 2010, 69, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.; Moridnia, A.; Mortazavi, D.; Salehi, M.; Bagheri, M.; Sheikhi, A. Kefir: A powerful probiotics with anticancer properties. Med. Oncol. 2017, 34, 183. [Google Scholar] [CrossRef]
- Mori, N.; Kano, M.; Masuoka, N.; Konno, T.; Suzuki, Y.; Miyazaki, K.; Ueki, Y. Effect of probiotic and prebiotic fermented milk on skin and intestinal conditions in healthy young female students. Biosci. Microbiota Food Health 2016, 35, 105–112. [Google Scholar] [CrossRef]
- Strzepa, A.; Gurski, C.J.; Dittel, L.J.; Szczepanik, M.; Pritchard Jr, K.A.; Dittel, B.N. Neutrophil-derived myeloperoxidase facilitates both the induction and elicitation phases of contact hypersensitivity. Front. Immunol. 2021, 11, 3552. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 178. [Google Scholar] [CrossRef]
- Galdeano, C.M.; Cazorla, S.I.; Dumit, J.M.L.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar]
- Cordeiro, M.A.; Souza, E.L.S.; Arantes, R.M.E.; Balthazar, C.F.; Guimarães, J.T.; Scudino, H.; Silva, H.L.A.; Rocha, R.S.; Freitas, M.Q.; Esmerino, E.A.; et al. Fermented whey dairy beverage offers protection against Salmonella enterica ssp. enterica serovar Typhimurium infection in mice. Int. J. Dairy Sci. 2019, 102, 6756–6765. [Google Scholar] [CrossRef]
- Beh, B.K.; Mohamad, N.E.; Yeap, S.K.; Ky, H.; Boo, S.Y.; Chua, J.Y.H.; Tan, S.W.; Ho, W.Y.; Sharifuddin, S.A.; Long, K.; et al. Anti-obesity and anti-inflammatory effects of synthetic acetic acid vinegar and Nipa vinegar on high-fat-diet-induced obese mice. Sci. Rep. 2017, 7, 6664. [Google Scholar] [CrossRef]
- Hunsche, C.; Cruces, J.; De la Fuente, M. Improvement of redox state and functions of immune cells as well as of behavioral response in aged mice after two-week supplementation of fermented milk with probiotics. Curr. Microbiol. 2019, 76, 1278–1289. [Google Scholar] [CrossRef] [PubMed]
- Karaffová, V.; Mudroňová, D.; Mad’ar, M.; Hrčková, G.; Faixová, D.; Gancarčíková, S.; Ševčíková, Z.; Nemcová, R. Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms 2021, 9, 831. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Identification of TLR2/TLR6 signalling lactic acid bacteria for supporting immune regulation. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hee, K.C.; Geun, K.H.; Yun, K.J.; Ra, K.N.; Jun, J.B.; Hye, J.J. Probiotic genomic DNA reduces the production of pro-inflammatory cytokine tumor necrosis factor-alpha. FEMS Microbiol Lett. 2012, 328, 13–19. [Google Scholar]
- Jia, G.; Liu, X.; Che, N.; Xia, Y.; Wang, G.; Xiong, Z.; Zhang, H.; Ai, L. Human-origin Lactobacillus salivarius AR809 protects against immunosuppression in S. aureus-induced pharyngitis via Akt-mediated NF-κB and autophagy signaling pathways. Food Funct. 2020, 11, 270–284. [Google Scholar] [CrossRef]
- Hoarau, C.; Lagaraine, C.; Martin, L.; Velge-Roussel, F.; Lebranchu, Y. Supernatant of Bifidobacterium breve induces dendritic cell maturation, activation, and survival through a Toll-like receptor 2 pathway. J. Allergy Clin. Immunol. 2006, 117, 696–702. [Google Scholar] [CrossRef]
- Gupta, T.; Kaur, H.; Kapila, S.; Kapila, R. Lactobacillus fermentum (MTCC-5898) alleviates Escherichia coli-induced inflammatory responses in intestinal epithelial cells by modulating immune genes and NF-κB signalling. J. Appl Microbiol. 2021, 131, 3008–3017. [Google Scholar] [CrossRef]
- Javanshir, N.; Hosseini, G.N.G.; Sadeghi, M.; Esmaeili, R.; Satarikia, F.; Ahmadian, G.; Allahyari, N. Evaluation of the Function of Probiotics, Emphasizing the Role of their Binding to the Intestinal Epithelium in the Stability and their Effects on the Immune System. Biol. Proced. Online. 2021, 23, 23. [Google Scholar]
- McGuckin, M.A.; Lindén, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Kaur, H.; Ali, S.A. Probiotics and Gut Microbiota: Mechanistic Insights into Gut Immune Homeostasis through TLR Pathway Regulation. Food Funct. 2022, 13, 7423–7447. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Thrapeutic Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef]
- Krishna Rao, R.; Samak, G. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Bonvicini, F.; Pagnotta, E.; Punzo, A.; Calabria, D.; Simoni, P.; Mirasoli, M.; Passerini, N.; Bertoni, S.; Ugolini, L.; Lazzeri, L.; et al. Effect of Lactobacillus acidophilus fermented broths enriched with Eruca sativa seed extracts on intestinal barrier and inflammation in a co-culture system of an enterohemorrhagic Escherichia coli and human intestinal cells. Nutrients 2020, 12, 3064. [Google Scholar] [CrossRef]
- Zhai, Z.; Torres-Fuentes, C.; Heeney, D.D.; Marco, M.L. Synergy between probiotic Lactobacillus casei and milk to maintain barrier integrity of intestinal epithelial cells. J. Agric. Food Chem. 2019, 67, 1955–1962. [Google Scholar] [CrossRef]
- Rokana, N.; Singh, R.; Mallappa, R.H.; Batish, V.K.; Grover, S. Modulation of intestinal barrier function to ameliorate Salmonella infection in mice by oral administration of fermented milks produced with Lactobacillus plantarum MTCC 5690–a probiotic strain of Indian gut origin. J. Med. Microbiol. 2016, 65, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Hsu, C.A.; Hung, W.T.; Chen, M.J. Effects of Lactobacillus paracasei 01 fermented milk beverage on protection of intestinal epithelial cell in vitro. J. Sci. Food Agric. 2016, 96, 2154–2160. [Google Scholar] [CrossRef]
- Vinderola, G.; Matar, C.; Perdigón, G. Milk fermented by Lactobacillus helveticus R389 and its non-bacterial fraction confer enhanced protection against Salmonella enteritidis serovar Typhimurium infection in mice. Immunobiology 2007, 212, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Usuda, H.; Okamoto, T.; Wada, K. Leaky gut: Effect of dietary fiber and fats on microbiome and intestinal barrier. Int. J. Mol. Sci. 2007, 22, 7613. [Google Scholar] [CrossRef]
- Bruno, C.; Paparo, L.; Pisapia, L.; Romano, A.; Cortese, M.; Punzo, E.; Canani, R. Protective effects of the postbiotic deriving from cow’s milk fermentation with L. paracasei CBA L74 against Rotavirus infection in human enterocytes. Sci. Rep. 2022, 12, 16268. [Google Scholar]
- Zhu, Y.; Xu, Y.; Wang, X.; Rao, L.; Yan, X.; Gao, R.; Shen, T.; Zhou, Y.; Kong, C.; Zhou, L. Probiotic Cocktail Alleviates Intestinal Inflammation Through Improving Gut Microbiota and Metabolites in Colitis Mice. Front. Cell. Infect. Microbiol. 2022, 760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Corredig, M.; Morales-Rayas, R.; Hassan, A.; Griffiths, M.W.; LaPointe, G. Effect of fermented milk from Lactococcus lactis ssp. cremoris strain JFR1 on Salmonella invasion of intestinal epithelial cells. Int. J. Dairy Sci. 2019, 102, 6802–6819. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Han, D.; Wang, Y.; Wang, Q.; Yan, X.; Wang, S.; Liu, X.; Song, W.; Ma, Y. Lactobacillus casei protects intestinal mucosa from damage in chicks caused by Salmonella pullorum via regulating immunity and the Wnt signaling pathway and maintaining the abundance of gut microbiota. Poult. Sci. 2021, 100, 101283. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Factories 2020, 19, 23. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of intestinal barrier function by microbial metabolites. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 1463–1482. [Google Scholar] [CrossRef]
Figure 1.
Fermented dairy-based product health benefits.

Figure 2.
The illustration shows the components necessary for scavenging free radicals and supplying antioxidants that are present in fermented foods. These antioxidants neutralize free radicals in a number of different ways. Conceptual diagrams of the important cellular stress and anti-oxidative stress signaling pathways are provided.
Figure 2.
The illustration shows the components necessary for scavenging free radicals and supplying antioxidants that are present in fermented foods. These antioxidants neutralize free radicals in a number of different ways. Conceptual diagrams of the important cellular stress and anti-oxidative stress signaling pathways are provided.

Figure 3.
The figure illustrates that fermentation-enhanced bacteria control the microbiota in the gut and maintain intestinal balance.
Figure 3.
The figure illustrates that fermentation-enhanced bacteria control the microbiota in the gut and maintain intestinal balance.

Figure 4.
Healthy probiotic bacteria derived from fermented foods are responsible for keeping infections at bay and maintaining the immune system’s equilibrium.
Figure 4.
Healthy probiotic bacteria derived from fermented foods are responsible for keeping infections at bay and maintaining the immune system’s equilibrium.


{kind=link}
{kind=link}
{kind=link}
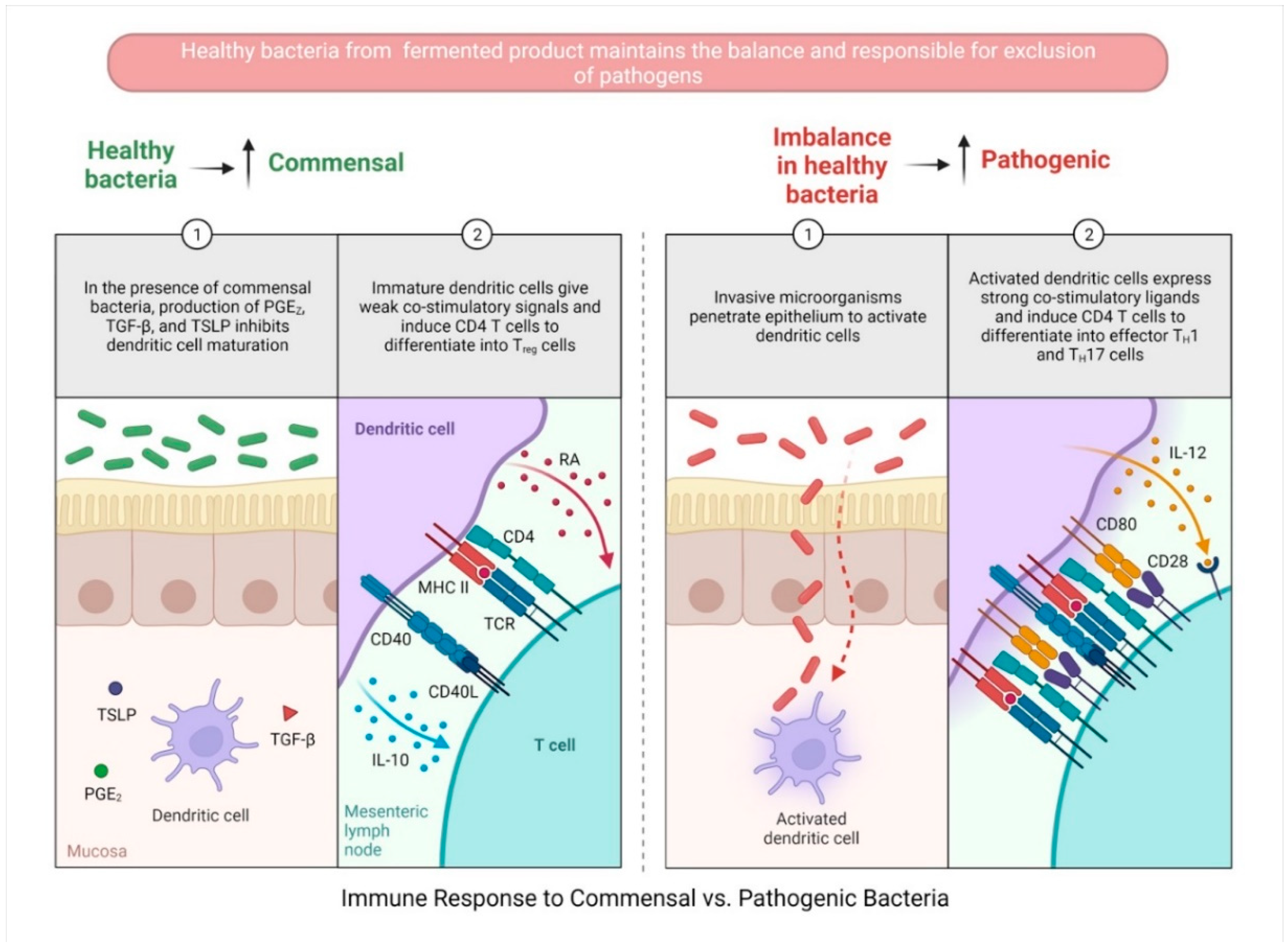{kind=link}
Table 1.
The studies demonstrating the effectiveness of fermented products containing healthy bacteria and health-promoting components against particular diseases and deficiencies.
Table 1.
The studies demonstrating the effectiveness of fermented products containing healthy bacteria and health-promoting components against particular diseases and deficiencies.
| Food Matrices | Microorganism | Health-Promoting Compounds | Disease | Reference |
|---|---|---|---|---|
| Kimchi | Lactobacillus plantarum (L. plantarum) | Several prebiotics and the presence of vitamins, minerals, and dietary fibers and O-linked β-N-acetylglucosamine (O-GlcNAc) | Colon cancer | [15] |
| Whey | L.rhamnosus MTCC 5897 | Probiotic released metabolites | Ulcerative colitis | [16] |
| Milk | Bifdobacterium breve | Bacteria and released components | Ulcerative colitis | [17] |
| Milk | L. rhamnosus NCDC 17 and L. rhamnosus GG | Metabolites | STZ-induced diabetic Wistar rats in high-fat diet (T2DM) | [18] |
| Koumiss | L. helveticus NS8 | Bacterial surface components | Gastrointestinal health | [19] |
| Skimmed milk | Lactococcus lactis strain NRRL B-50571 | ACE 3 inhibitory peptides and GABA | Anti-hypertensive activity | [20] |
| Milk, yogurt | LAB and Bifidobacteria species | Folate (vitamin B9), vitamin K, riboflavin (vitamin B2) | Vitamin deficiency | [21,22] |
| Cashew apple juice, kimchi | 5 probiotic strains (Lb. acidophilus, Lb. casei, Lb. plantarum, Lb. mesenteroides and B. longum) | Vitamin K (Phylloquinone) | Vitamin deficiency | [23,24] |
| Milk | L. casei DN-114 00 | - | Common infectious diseases (CIDs) | [25] |
| Milk | L. acidophilus and L. casei | - | Alleviation of lactose intolerance | [26] |
| Milk | L. helveticus LH511 | Bioactive components in supernatant | Barrier dysfunction | [27] |
| Shrikhand | L. acidophilus NCDC14 | - | Gastrointestinal health | [28] |
| Kefir | Lactobacillus lactis subs., Leuconostoc subs., Streptococcus thermophilus, Lactobacillus subs., and yeast of kefir | - | Inflammatory bowel disease (IBD) | [29] |
| Milk | L. paracasei CNCM I 1518 | - | Cirrhosis | [30] |
| Milk | L. fermentum NCDC 400 and L. rhamnosus NCDC 610 | - | Obesity and other related metabolic syndromes | [31] |
| Milk | L. rhamnosus MTCC 5897 | - | Hypercholestrolemia | [32] |
| Milk | L. acidophilus, L. amylovorus, L. bravis, L. bulgaricus, L. casei, L. curvatus, L. fermentum, L. gasseri, L. helveticus, L. lactis, L. paracasei, L. plantarum, L. rhamnosus, L. reuteri, L. salivarius, B. lactis, B. breve, B. bifidum, B. longum, B. infantis, Pediococcus pentosaceus, and Streptococcus thermophilus. | - | Cardiometabolic diseases (CMDs) | [33] |
| Yogurt, capsules | L. rhamnosus GG (ATCC 53013) | Long chain fatty acids | Acute pediatric diarrhea, antibiotic-associated diarrhea | [34] |
| Squacquerone cheese | L. crispatus BC4 | Calcium, vitamins with antimicrobial properties | Vaginal infections | [35] |
| Cream cheese | L. chungangensis CAU28 | Short chain fatty acid | Atopic dermatitis | [36] |
| Cheddar cheese | Lacticaseibacillus rhamnosus and Lc. paracasei | γ aminobutyric acid (GABA) | Gastrointestinal digestion | [37] |
| Milk | B. longum subsp. longum YS108R; L. paracasei PS23 | Exoplysaacharide (EPS); short chain fatty acids (SCFA) | Ulcerative colitis | [38,39] |
| Milk | Lactobacillus spp. | Free amino acids and lactate | Bone health | [40] |
| Milk | L. brevis DL1-11 | γ aminobutyric acid (GABA), short-chain fatty acids (SCFAs), such as butyric acid | Insomnia | [41] |
| Milk | L. acidophilus | Phenolic compounds, GABA 1, peptides, CLA 2, folates (vitamin B9) | Antioxidant activity related disorders | [42] |
Table 2.
The beneficial effects of probiotic-fermented products on human health showing in vivo and in vitro studies.
Table 2.
The beneficial effects of probiotic-fermented products on human health showing in vivo and in vitro studies.
| Formulation | Probiotic Strain | Associated Mechanism | Disorder | Model Organism | Reference |
|---|---|---|---|---|---|
| Kefir drink | Commercial kefir starter culture (Danisco®) | Fermented kefir ameliorates the colitis-induced symptoms in dose-dependent manner, relieves from disease-induced diarrhea and macroscopic damages caused in mucosal wall | Inflammatory bowel disease (IBD) | Rat | [29] |
| Whey drink | L. rhamnosus MTCC 5897 | Increases the anti-inflammatory cytokines to ameliorate the colitis-induced inflammation, enhances the tight junctional epithelial barrier integrity in mice | Ulcerative colitis | Albino mice | [16] |
| Fermented milk | B. longum subsp. longum YS108R | Reduces inflammatory cytokine concentrations in serum as well as their mRNA expression levels in the colon by increasing tight junction protein and MUC2 expression | DSS-induced colitis | C57BL/6J mice | [38] |
| Fermented milk | L. rhamnosus MTCC: 5957 and L. rhamnosus MTCC: 5897 | Reduces the high-cholesterol diet (HCD)-induced body weight, hyperlipidemia, and hepatic lipids (total cholesterol and triacylglycerol), decreases the TNF-alpha, IL-6 in the liver | Diet-induced hypercholesterolemia | Wistar rats | [32] |
| Fermented milk | L.paracasei PS23 | Strengthened tight junction through the modification of specific cecal bacteria and upregulation of short-chain fatty acids | DSS-induced colitis | Mice | [39] |
| Fermented milk | L. fermentum | Bioavailability of zinc | Zinc depletion | Wistar rats | [53] |
| Milk; fermented milk | L. fermentum | Improvement in the hypercholesterolemic effect | Hypercholesterolemic | Wistar rats | [54] |
| Cream cheese | L. chungangensis CAU28 | Maintains Th1 and Th2 balance as well as reduces IgE immunoglobulin associated with immune homeostasis | Atopic dermatitis | Balb/c mice | [36] |
| Fermented milk | L. paracasei CNCM I 1518 | Decreases bacterial translocation, gut dysbiosis, and ileal oxidative damage and increases ileal β-defensin-1 expression in rats treated with CCl4 | Cirrhosis | Sprague-Dawley rats | [55] |
| Fermented milk | Lactobacillus spp. | Improves amino acid absorption that in turn enhances postprandial MPS via Akt-independent mechanisms | Skeletal muscle protein anabolism | Sprague-Dawley rats | [40] |
| Yogurt and fermented milk | FMP: Lactobacillus helveticus; -Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus | Greek-style yogurt protein (YP) increased the expression of genes involved in jejunal and ileal immunity and integrity in WT mice; as well as YP also improved insulin sensitivity by 65% in LRKO mice, Lactobacillus helveticus fermented milk protein (FMP) and YP both modulate the intestinal microbiota | Cardiometabolic markers and intestinal microbiota | C57BL/6J wild-type (WT) and atherosclerotic (LRKO) male mice | [56] |
| Fermented milk | Lacticaseibacillus casei CRL431 | Probiotic-fermented milk (PFM) had the highest percentages for CD4+ and CD4+ CD8+ cells. While IL-10, TNF-α, IFN-γ, and IL-6 concentrations decreased significantly in the PFM group that increased in mice from the milk group | Breast cancer | Female BALB/c mice | [57] |
| Fermented milk | Lactobacillus brevis DL1-11 | Modulates the composition of the gut microbiota, such as significant increases in the relative abundances of Ruminococcus, Adlercreutzia, and Allobaculum. Moreover, these changes may be associated with significant increases in the production of SCFAs in the intestine | Insomnia | Rat | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kaur, H.; Kaur, G.; Ali, S.A. Dairy-Based Probiotic-Fermented Functional Foods: An Update on Their Health-Promoting Properties. Fermentation 2022, 8, 425. https://doi.org/10.3390/fermentation8090425
AMA Style
Kaur H, Kaur G, Ali SA. Dairy-Based Probiotic-Fermented Functional Foods: An Update on Their Health-Promoting Properties. Fermentation. 2022; 8(9):425. https://doi.org/10.3390/fermentation8090425
Chicago/Turabian StyleKaur, Harpreet, Gurjeet Kaur, and Syed Azmal Ali. 2022. "Dairy-Based Probiotic-Fermented Functional Foods: An Update on Their Health-Promoting Properties" Fermentation 8, no. 9: 425. https://doi.org/10.3390/fermentation8090425
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.