

Microalgal Biodiesel: A Challenging Route toward a Sustainable Aviation Fuel

1
School of Architecture Technology and Engineering, University of Brighton, Cockcroft Building, Brighton BN2 4G, UK
2
Department of Mechanical, Biomedical and Design Engineering, College of Engineering and Physical Sciences, Aston University, Birmingham B4 7ET, UK
3
Internal Combustion Engineering Division, Department of Mechanical Engineering, Anna University, Chennai 600 025, India
4
Chemical Engineering & Applied Chemistry, College of Engineering and Physical Sciences, Aston University, Birmingham B4 7ET, UK
*
Author to whom correspondence should be addressed.
Fermentation 2023, 9(10), 907; https://doi.org/10.3390/fermentation9100907
Submission received: 14 July 2023
/
Revised: 4 October 2023
/
Accepted: 8 October 2023
/
Published: 16 October 2023
(This article belongs to the Special Issue Algae—the Medium of Bioenergy Conversion: 2nd Edition)
Abstract
:By 2050, aviation-related carbon emissions are expected to quadruple to over 3000 million tonnes of carbon dioxide, so finding sustainable alternative solutions to minimise pollution is a key scientific challenge. Aviation gasoline and kerosene are currently used to power most jet engines. While battery-powered planes and planes that could utilise a cleaner fuel, such as hydrogen, are possible, the time scale required to improve and implement these technologies is distant, with air fleet turnover taking some 30 years. Existing jet engines could be modified to run on biodiesel, and considering the close similarity in fuel density to kerosene, could be a less disruptive approach to the industry. The sheer volume of biodiesel required remains a challenge, and certainly, using plant-derived oils grown on arable land is not acceptable, as it competes with food production. However, high-lipid-yielding microalgae (where productivity is an order of magnitude greater than oilseeds), grown on marginal land, such as desert or semi-desert areas of the world, could be possible. Indeed, to replace 30% of fossil fuel with algal-derived biodiesel would require 11,345 km2 of land. Biodiesel preparation is well understood, but what is lacking is proven technology aimed at optimising microalgal production of oil at a much larger scale. Here, a synergic review of the current state-of-the-art in algal production, that includes strain selection, possible production sites, culturing costs, and harvesting to identify the bottlenecks in meeting the ASTM specifications for the aviation industry, is presented.
1. Introduction
The rapid growth of the aviation industry, in recent decades, as well as airport expansions, has exacerbated aviation’s role in climate change and environmental protection [1]. Annually, about 3.1 billion people and 51.7 million tonnes of cargo are operated by 1000 commercial airlines, using approximately 15,000 aircrafts [2]. This has led to an annual 3.5–4.5% increase in hydrocarbon fuel consumption [3]. To meet the 2050 carbon reduction targets, improving aircraft fuel carbon-neutrality and emissions are critical [4].
Aircraft engine exhaust comprises up to 8% CO2 (approximately 900 Mt CO2) water vapour, NOx (0.03%), UHC, CO, and SO2 [5]. In 2010, the share of total global GHG emissions from the air transport sector was 4% and 6%, respectively, for domestic and international flights [6]. Although these numbers seem relatively low because the emissions are at high altitude, the NOx generated in the upper atmosphere leads to ozone production and is, therefore, likely to be more damaging than these figures initially suggest. Hence, to reduce the aviation carbon emission with sustainable energy efficiency, sustainable aviation fuel (SAF), such as algal-based biofuel, will be the key for this industry [7]. Such an alternative could potentially reduce the life cycle of CO2 emissions by up to 80% by 2050, compared to 2005 [6]. Fuel sustainability criteria must include: (1) suitability for existing aircraft engines, (2) no modifications in infrastructure, (3) must produce low life cycle carbon emissions, as compared to fossil-based jet fuel, (4) food and ecology should not be affected by energy crops, and (5) deforestation should not be the reason [8]. The SAF should be produced from biological feedstocks, such as non-edible oil crops (e.g., Jatropha, Palm, and Camelina), algae, municipal sewage and agricultural wastes [7,8]. The demand for jet fuel is 5–6 million barrels/day and is expected to increase by 15% by 2030 and account for 3.4% of the global CO2 emissions. Of the required global fuel supply of 6–7 billion litres/year, 30% is expected to be from bio-sources by 2030 according to the International Air Transport Association (IATA) [9,10]. In this context, offsetting CO2 emissions using microalgae is attractive since 1 tonne of microalgae can absorb 1.8 tonnes of CO2 from the 29 gigatons of CO2 that is released into the atmosphere annually from burning fossil fuels [9,10]. Improvements in other areas, such as lighter materials, improved engine design, and aerodynamics, have resulted in a 70% reduction in aircraft traveller/kilometre and tonne/kilometre consumption [11].
Implementing aircraft engine design takes time since most are powered by kerosene and changing the fuel source introduces significant design challenges. Indeed, it typically takes up to 10 years to design an aircraft and 30 years to roll over a fleet [12]. This time scale already takes us beyond 2050, the target date by which implementation of CO2 reduction targets need to be met [12,13]. In essence, this means that decarbonising aviation fuel should have started some time ago. Kerosene is the main fuel type to power flights, and engines are optimised to utilise this fuel source [14]. If radical engine design is to be avoided, then a fuel type close to the fuel density characteristics of kerosene (0.78–0.81 g cm3) needs to be developed. In this regard, the biodiesel fuel density is close to kerosene (0.88–0.9 g cm3); therefore, adopting this fuel would mitigate the need for a radical shift in aircraft design [14,15]. Other alternatives, such as battery-powered aircraft, are currently unfeasible for long-haul, high-payload flights [16]. They may, however, have a future for light aircraft undertaking short- to medium-range destinations [17]. Hydrogen has also been proposed as a possible fuel [18,19], but considering its low density and the intrinsic difficulties of handling it both as a liquid and in gas form, it would require significant modification to the aircraft design [20]. The time to achieve such a drastic change implies that keeping to hydrocarbon-based fuel would be highly beneficial. However, as with other biofuel types, being able to generate the high volumes needed for the industry remains a significant challenge [20]. Clearly, using plant oils derived from crops grown on agricultural land is unfeasible due to the sheer area required to support the industry, and this coupled with the relatively low oil yield per hectare makes it less attractive (see Section 3). However, the possibility of using other sources to generate biodiesel, such as microalgae grown on marginal land and/or desert/semi-desert areas, could be feasible [21]. In this review, we have attempted, for the first time, to consider microalgae as a potential source of aviation biodiesel by drawing together information on the requirements that would be needed to achieve this goal. This includes recent developments in the understanding of algal oil biosynthesis, cost implications for mass culture, and the land area required to facilitate such a large-scale production. Potential desert sites are suggested, together with an assessment of their suitability.
2. Current Aviation Fuel Production
For jet fuel, the aviation industry primarily uses two types of fuel: aviation gasoline and aviation kerosene [22]. Aviation gasoline, also known as AVGAS, is derived from the distillation of crude oil in the range between 30 and 170 °C [23,24]. This fuel mainly consists of isoparaffins with C5 to C9 carbon and a small amount of aromatic carbon. Alkylates are a mix of high-octane, low-vapour-pressure, branched paraffinic hydrocarbons used for blending into gasoline [23,25]. AVGAS is normally used in small aircraft, such as private aviation, agricultural, and pilot training. Aviation kerosene is also a petroleum distillation product obtained in the range between 150 and 300 °C [23,25]. This fuel is more suitable in aircraft engines for energy generation through combustion, used in many commercial and military aircraft [26]. Many developed countries are following the jet fuel specifications, which are compatible with the Aviation Fuel Requirement for Operated System (AFQRJOS) (Table 1).
2.1. Aviation Fuel Specification
Aviation fuel properties and compositions should meet the fuel specifications standards [27]. They should have the lowest energy density by mass, the highest permitted freeze point, the highest allowable viscosity, the highest allowable sulfuret and aromatic content, the highest allowable acidity, and the lowest allowable flash point [28,29,30]. Parameters for the certified international standards for aviation jet fuel are presented in Table 2 and agree closely with one another in many respects.
The chemical composition of Jet A and A1 fuels is presented in Table 3. Straight-chain hydrocarbons, C8 (octanes), and cyclic derivatives, both aromatic and non-aromatic, are major components.
2.2. Kerosene
Kerosene is a clear hydrocarbon-rich liquid obtained from petroleum distillation over a temperature range of 150 to 300 °C [14]. It is less viscous than biodiesel with a density of 0.78–0.81 g/mL and is composed of hydrocarbons predominantly from C10 to C16, although the range can extend to C6–C20. The main constituents are branched and straight-chain alkanes, as well as cycloalkanes, which account for at least 70% of the total volume [29]. Studies on synthetic alternative jet fuels with predominantly linear and lightly branched alkane content provide stronger low-temperature ignition characteristics, while other types of synthetic alternative jet fuels with a high content of highly branched alkanes exhibit weaker, low-temperature ignition characteristics, when compared to conventional jet fuels [31]. Aromatic hydrocarbons such as alkyl benzenes and alkyl naphthalene are rarely seen in concentrations greater than 25% by volume. Alkene content is low and typically makes up less than 5% of the total volume. Kerosene has a flash point of 37 to 65 °C and an autoignition temperature of 220 °C. Kerosene has a freezing point of −47 °C; however, this varies depending on the quality of the aviation fuel requirements [32]. Kerosene has a lower heating value of around 43 MJ/kg and a higher heating value of around 46 MJ/kg, similar to fossil diesel. The ASTM D-3699-78 recognises two grades of kerosene: 1-K (less than 0.04% sulphur by weight) and 2-K (0.3% sulphur by weight). The 1-K grade is better than the 2-K grade, providing cleaner combustion and fewer toxic emissions [32].
2.3. Routes to SAF Production
Several conversion routes have been developed to convert biomass into aviation fuel, and some key features are highlighted in Table 4, which include hydro-processing of esters and fatty acids (HEFA), Fischer-Tropsch (FT), and alcohol-to-jet (ATJ) [33]. In this regard, the only commercially demonstrated process for 100% SAF is the UOP-Eco-refining™ technology developed by Honeywell (USA).
In this review, we will focus attention on the production of biodiesel from algae as a suitable replacement for kerosene since it has the closest fuel density properties. An overview of the process is outlined in Figure 1.
3. Microalgae as a Potential Fuel Feedstock
Many microalgae have a high oil content, ranging from 20% to 77% (Table 5). Particularly high levels are found in Schizochytrium sp., Botryococcus braunii, Nanochloropsis sp., and Neochloris oleoabundans [34]. The lipid is largely in the form of triacylglycerol, which can be readily converted into biodiesel through transmethylation.
In terms of productivity per area, microalgae outperform their closest vegetable oil crop (Palm) by a factor of nearly 16 times (Table 6). Although Soy is one of the major oil crops in terms of global productivity, it is grown for protein, while the oil (about 20%) is a by-product [27].
The use of vegetable oils for fuel raises a strong ethical debate on the appropriate use for agricultural land use [25]. Many oils are considered non-edible, e.g., Tung and Jatropha could be used for biodiesel production [25]. Indeed, Jatropha also has the benefit of being able to grow on marginal land, which is rather nutrient-poor yet produces a high oil yield, and when water irrigation is provided to this crop, oil yields can be triple those quoted in Table 6. Palm cultivation, despite its high oil yield, has caused extensive destruction of rainforest, particularly in Indonesia, raising serious environmental concerns about its suitability considering its high negative impact on biodiversity [30,36]. Indeed, many food producers now state that their products are made without the use of palm oil, thereby allowing consumers to make informed choices. In contrast, microalgal oil production, estimated to be ca. 95,000 L/hectare/year, is significantly higher than Palm [37,38]. Other common oilseed crops are significantly lower (Table 6).
In 2019, the aviation industry consumed ca. 360 billion L of fuel [39,40]. To achieve a 30% replacement from algal sources, 108 billion litres would be required. An alga with an oil content of 50% would yield 86,582 L biodiesel/ha/year [3,4]. The land required to produce the 30% replacement of fossil fuel is equivalent to 11,345 km2 (Table 7).
4. Potential Areas for Growing Microalgae
The necessity to avoid using arable land for fuel production raises the question as to where large-scale microalgal cultivation could be achieved [41]. In this regard, one of the most under-utilised land areas on Earth are deserts, which cover approximately one-fifth of the surface area [43]. Desertification is an ongoing process, and aridification of the earth land surface could rise to 30% in the coming decades as a consequence of several factors, including urbanisation and climate change [44]. Indeed, the Sahara Desert expands 7600 km2 per year and has increased in size by 10% over the last century. Deserts are defined as areas of low rainfall (typically <250 mm/year), and the reasons for this are varied, giving rise to their classification [44] as follows: (1) subtropical, (2) coastal, (3) rain shadow, (4) interior, and (5) polar.
Some of the major deserts are listed in Table 8, together with some of their characteristics, including area, rainfall, temperature, and some key topographical summaries [35,44,45,46]. Clearly, there are many regional variations in temperature and rainfall, even within a given desert area, and the complexities are not possible to capture here [45]. However, sufficient trends and features can be seen that could inform the identification of possible mass algal production sites [35]. Some deserts can be ruled out, such as the Antarctic and Arctic Deserts, due to their exceptionally low temperatures for most of the year and because they are not readily amenable to human habitation and infrastructure development [46]. Others, such as the Namib Desert, are unfavourable due to low temperatures, thick coastal fogs, and high sand dunes (300 m), formed by Atlantic winds. On the Arabian Peninsula, temperatures often reach 50 °C in the summer months [35]. High temperatures and low rainfall mean that the Arabian Desert would be unsuitable to cultivate microalgae in open raceway ponds due to evaporation-related problems [35]. This would be particularly problematic since relatively shallow water columns are required for the penetration of light for photosynthesis (typically in the range of 10 to 50 cm) [41]. The Atacama Desert is the driest place on Earth, with many years of no rainfall.
The Kalahari Desert, apart from being of great significance for wildlife, is 900 m above sea level, and since a 6% energy cost would be incurred for every 100 m of elevation [47], this would make it less attractive. Considering this cost implication, ideally, the desert will be adjacent to the sea, be of flat topography, and will not suffer from extremes of temperature [42]. To this end, The Great Rann of Kutch is a seasonal salt marsh in the Thar Desert (200,000 km2) in the Kutch District of Gujarat in India, with an area of 7500 km2, and could produce up to 66% of the required volume, or 100% at 20% fossil fuel replacement [35,45]. Using algae that are able to produce only a modest 30% by weight of oil would require 38,000 km2 of land [45]. The average temperature of the desert is 28 °C, and the desert receives approximately 140 mm year−1 of rainfall. Here, the topography of the land is flat and thus suited for microalgae cultivation and worthy of further investigation as a potential site [45].
In Europe, only one area is designated as a desert, the Tabernas Desert in Almeria, Spain, with an area of 258 km2 [42,46]. The temperature is mild at 16–29 °C, with an average of 18 °C, and rainfall in the range of 150–220 mm/year. Apart from the limitation in size, the area is elevated up to 900 m above sea level so, again, pumping sea water would come at a high energy cost [41,45].
5. Current Challenges in Cultivating High-Oil Microalgae
Despite the theoretical appeal of microalgal oil yield, translating lab- and pilot-scale results into large-scale production is not straightforward. While stable conditions can be achieved in the laboratory, in outdoor settings, many variables can arise that negatively impact algal growth [46,48]; for example, sharp changes in temperature, dilution of growth media due to rain, or conversely, concentration due to evaporation caused by high temperatures. In open raceway ponds, contamination by microbes in the environment can occur, causing culture collapse [46,48]. Additionally, with time, strain selection can set in, which can impact predictions based on the use of the initial strain used to inoculate the system. Attempts to model this transition from laboratory- to larger-scale production have been undertaken [46,48]. There are several current bottlenecks in the process of scale-up to produce the sheer volume of algal oil required for the aviation sector [49]. The key targets are strain selection, nutrient costs, growing platforms, harvesting, and optimising oil production [50].
5.1. Strain Selection
Microalgae grow in a wide range of environments, from arctic regions to tropical and desert areas [51]. Higher productivity is associated with higher temperature and growth rates, which favour sites closer to the equator. For biodiesel production, algae with a high oil content are necessary [51]. The level of oil can vary significantly, dependent on environmental conditions, whereby potential stress triggers such as N depletion can stimulate oil production within certain limits [52]. Other triggers, such as exposure to the trace metal thallium, have been shown to induce lipid accumulation in microalgae operating through a nitrous oxide (NO) pathway [53]. In stationary phase cells, ideally, the lipid content should be at least 40% by weight of the wet cells. In terms of fatty acid composition, an oil rich in the monounsaturated fatty acid, oleic acid (18:1(Δ9)), is preferred over those with a high content of saturated fatty acids and polyunsaturated fatty acids [52]. This is because biodiesels prepared form saturated fatty acids tend to crystalise at low temperatures and would cause blockages in fuel lines in regions where the ambient temperature was low (<−20 °C) [54]. Conversely, oils rich in polyunsaturated fatty acids, such α-linoleic acid (18:2(Δ9,12)) and linolenic acid (18:3(Δ9,12,15)), eicosapentaenoic acid (EPA) (20:5 (Δ5,8,11,14,17)), and docosapentaenoic acid (DHA) (22:6 (Δ4,7,10,13,16,19)), are prone to oxidation, leading to the formation of secondary products such as aldehydes and ketones. These compounds have deleterious effects on engine performance and life span [55].
Since microalgae grow best at optimal temperatures typically around 25–30 °C, attempts to cultivate microalgae in regions where temperatures exceed 40 °C will require strain selection for tolerance to elevated temperatures [41]. Such high temperatures are likely to be met in semiarid or desert areas in many parts of the world, as previously discussed in Section 4 [41].
Another challenge for strain selection is tolerance to high-salt conditions [56]. Using saline water is a cheaper option than freshwater, particularly if coastal regions are selected to set up the algal production site. Many algae can be conditioned to adapt to increasing salinity through changes in their osmoregulatory functions [56]. In oil-producing countries, one barrel of crude oil results in the production of nine barrels of contaminated water. Tolerance to potentially toxic chemicals in the water would be advantageous and would provide an ideal use for this readily available source of water. Indeed, the microalgae may even play a key role in bioremediation, rendering the contaminated water more useful for other applications [57].
Screening for strains that are adapted to the local conditions could be advantageous. The use of high-throughput screening methods to assess growth under a variety of conditions can be carried out in small volumes (ca. < 1 mL) in microplates and assessing growth spectrophotometrically based on chlorophyll (or other pigment) absorption [58]. Alternatively, choosing strains that are known to have a high lipid content from stock culture centres and subjecting them to challenging conditions that are likely to be encountered in the field could also be profitable [58].
5.2. Nutrient Cost
As with land plants, the application of nutrients (fertilisers) enhances algal growth [59], and without these additions, growth rates are significantly lowered. In this regard, microalgae can be cultivated in autotrophic (i.e., only requiring sunlight source), heterotrophic (external carbon supply), or mixotrophic modes (combination of autotrophic and heterotrophic modes) [60]. Autotrophic cultivation is the cheapest mode, as the microalgae use energy provided by sunlight to fix CO2 and absorb H2O, to make oils [60]. However, a drawback with this cultivation method is the requirement for high sunlight levels and generally the slow growth compared with heterotrophic/mixotrophic approaches [61]. Generally, higher growth rates can be achieved when an external carbon source (e.g., acetate, glucose) is supplied; however, this carries an additional cost, but this is offset by the requirement for lower light intensities [61]. Indeed, cultivating Chlorella heterotrophically shows a 5.5 times quicker growth rate compared to cultures grown autotrophically, since the former mode is not limited by sunlight, and final biomass yields of 50–100 g/L, compared with 30 g/L, respectively [62]. When the sunlight intensity is too low, the storage lipids produced during photosynthesis are used in photorespiration, leading to a decreased oil yield [62]. Some of the advantages and disadvantages are summarised for the two major cultivation methods in Table 9.
Determination of the land area required for the two modes of growth revealed that heterotrophically cultivated microalgae require 12 times less land for cultivation than autotrophic microalgae [63]. In conclusion, therefore, growing microalgae heterotrophically is recommended for cultivating microalgae on an industrial scale.
The three major macronutrients are nitrogen, phosphorus, and potassium. Twenty-five million tonnes of N and four million tonnes of P would be required to replace fossil fuels with algal biomass in the EU alone. This is more than double the current EU capacity [63]. This situation is currently exacerbated by the Ukraine–Russia conflict, which has significantly driven up the price of fertilisers in 2022 [64]. Trace elements (micronutrients including Mo, K, Co, Fe, Mg, Mn, B, and Zn) are also required to support many enzymatic functions in algal cells [65]. Carbon dioxide is also required for photosynthesis, and this compound currently accounts for about 50% of the base costs for algal production, estimated to be ~1.6–1.8 EUR/kg [65]. Use of wastewater, e.g., from industrial poultry production and by-products from the milk industry, could help to supply nutrients and reduce costs, as well as reduce environmental pollution [65]. Due to the high sensitivity of distance to the refinery, the location of the facility has a significant impact on emissions [65]. Ideally, algal farms should have other industries built alongside them that generate a large amount of CO2, such as concrete manufacturing plants [63,65]. Microalgae that are able to utilise CO2 directly from flue gas would be desirable, since no prior removal of potential toxic gases from the gas mixture would be required, e.g., SO2, which forms sulphuric acid in aqueous solution [65]. In petroleum-rich countries, wastewater generated from the extraction of oil is produced at a 9:1 ratio [65]. This water is unfit for human consumption but could potentially be used for algal production (after bacterial bioremediation to remove toxic compounds), and thereby produce oil for biodiesel or high-value products such as pigments, proteins, and polyunsaturated fatty acids [66].
5.3. Growing Platforms and Harvesting
In essence, there are two principal methods by which algae can be mass cultivated, open raceway ponds and enclosed photobioreactors. A summary of the advantages and disadvantages of both methods is outlined in Table 10.
The expression net energy ratio (NER) of biomass products is produced by the sum of the energy used for cultivation, harvesting, and drying divided by the energy content of the dry biomass. NER values < 1 indicate a positive energy outcome, whereas values > 1 indicate net imbalance [49]. In general, raceway ponds are typically more energy-efficient than photobioreactors. To some extent, this depends on the value of the product, so with high-value products, photobioreactors may be more cost-effective [67]. For fuel, however, which must have a large volume, low-cost products such as raceway ponds may be more cost-effective [67].
Even in the stationary phase, when the cells have stopped dividing and the water looks intensely green, the actual biomass present in the water sample is low and requires concentration [68]. Typical dry-weight biomasses for a range of microalgae, including Chlorella, Scenedesmus, Monoraphidium, Carteria, and Nannochloropsis, are in the range of 0.1–0.5 g/L, but generally higher in photo-bioreactors at 0.5–0.8 g/L. Recently, a strain of Parachlorella kessleri was shown to accumulate biomass to 6.9 g/L, with an oil content of 45% and a wide tolerance to pH [69]. In addition, developments in biofilm reactors [70] have shown that significantly improved yields can be achieved with yield reports in the order of 8 g m2 day−1 [71], and even up to 16 g m−2 day−1 [72]. While suitable for wastewater remediation and biorefinery activities, application of this technology for large-scale biofuel production remains to be established. Harvesting costs are estimated to be around 20–30% of the total production costs [68]. While in the laboratory a bench-top centrifugation is applicable, such a technique would be unfeasible for the large volumes of water that would be required to be processed [68]. Other methods of concentrating the algal cells will have to be sought. Among these, precipitation with flocculants is possible, in which the added chemical causes the cells to aggregate together and sediment out of the water column. Examples of flocculants include chitosan (derived from crustacean shells) and alum, a hydrated sulphate salt of aluminium XAl (SO4)2. 12H2O [67]. Froth flotation is also possible, in which the hydrophobic surface chemistry of the cells interacts with air bubbles to produce a foam [54]. Magnetic nanoparticles coated with cationic polymers have also been suggested [54]. Detailed life cycle analysis of the potential use of such harvesting agents is required in order to assess their commercial viability, and this is currently ongoing [73].
5.4. Optimising Oil Production
A high-oil-yielding and fast-growing microalgal strain is highly desirable. To improve the oil yield, the factors that control the partitioning of metabolites into oils, carbohydrates, and proteins need to be understood. In this regard, it has been demonstrated that it is possible to re-direct metabolites in Nanochloropsis gaditana [20]. This alga usually accumulates ca. 20% oil; however, by using reverse genetics to negate the impact of transcription factors that adversely affect oil accumulation, oil levels up to 40–55% have been reported [20]. Further targets to improve oil yields could be achieved if the metabolic pathways for oil synthesis can be more fully understood.
Nitrogen starvation is known to stimulate oil biosynthesis in microalgae [74,75,76] at the expense of vegetative growth, with a lag phase typically around 72 h. This has potential cost implications for the biomass yield versus oil yield that accompany these physiological changes. High-throughput mass spectrometry integration of protein and metabolite profiles revealed that cell growth and N metabolism were clearly linked by the branched-chain amino acids, suggesting an important role in this stress [77]. Lipid accumulation was also tightly correlated to the COP II protein, involved in vesicle and lysosome coating, and a major lipid droplet protein. This protein, together with other key proteins mediating signal transduction and adaption, constitutes a series of metabolic and regulatory targets that could be exploited to improve oil yields.
Triacylglycerols are synthesised via the acylation of glycerol 3-phosphate in two steps to produce phosphatidic acid (PA, reactions 1 and 2, Figure 2). Genetic manipulation of the first acylating enzyme (reaction 1, lysophosphatidic acid acyltransferse), in combination with reaction 5 catalysed by diacylglycerol acyltransferase (DAGAT), resulted in an increase in triacylglycerol production in Neochloris [78]. A phosphatidate phosphohydrolase hydrolyses the PA to generate diacylglycerol (DAG, reaction 3), which is subsequently acylated at the sn-3 position to yield triacylglycerol (TAG) (reaction 5). Separate acyltransferases use acyl-CoA substrates to catalyse reactions 1 and 2. A phosphatidylcholine:diacylglycerol acyltransferase (PDAT) catalyses reaction 4, which produces TAG and lyso-phosphatidylcholine (LPC). This reaction has been shown to be responsible for the transfer of EPA from phospholipids to TAG in Nanochloropsis [50]. Transacylation of diacylglycerol (DAG) with another diacylglycerol molecule generates TAG and monoacylglycerol (MAG). The acylation of DAG with acyl-CoA to create TAG is catalysed by the enzyme DGAT (reaction 6). TAG production has been shown to increase when this enzyme is overexpressed [72,79]. However, in algae, the relative relevance of the various pathways to TAG production has yet to be determined, and it may likely change from species to species and be dependent on environmental stressors [80]. Heat stress in Chlamydomonas, for example, led to the activation of a direct pathway for the conversion of membrane lipids to store lipids [81,82]. Transcription factors responsible for regulating lipid synthesis in green algae have been identified and represent a significant step forward in our understanding of the control processes involved in lipid partitioning [83].
Additional reactions to generate polyunsaturated fatty acids have been demonstrated in plants [84,85] and include:
- (i)
- Diacylglycerol and phosphosphatidylcholine (PC) interconversion catalysed by cholinephosphotransferase.
- (ii)
- Acyl exchange between the acyl-CoA pool and position sn-2 of PC catalysed by lyso phosphtidylcholine:acyl-CoA acyltransferase.
However, reactions (i) and (ii) have yet to be demonstrated in algae that accumulate PUFA-rich triacylglycerols, although it is likely they occur. Such reactions would not be desirable in microalgae destined for biodiesel production because of the oxidative instability of the PUFAs generated [86]. Indeed, downregulating the enzymes responsible for these reactions would be possible genetic targets in order to increase the level of oleic acid in the triacylglycerol, and thereby increase oxidative stability [87].
5.5. Fatty Acid Composition and Oxidative Stability of Derived Biodiesels
In contrast with kerosene, algal oils are predominantly composed of triacylglycerol (ca. 95%), with minor quantities of diacylglycerol and phospholipids (ca. 5% combined) [52]. The ester bonds generated between the carboxylic acid of the fatty acid and the glycerol backbone in these natural compounds include oxygen. Due to its high boiling and flash temperatures, triacylglycerol is commonly converted to biodiesel for use in combustion engines [41]. This entails the transesterification of the oil under anhydrous circumstances in the presence of a catalyst, commonly using methanol (either a base such as NaOH or an acid such as H2SO4). Fatty acid methyl esters (FAMEs) are extracted from the glycerol by-product, rinsed to remove any remaining catalyst, dried, and utilised as biodiesel [88]. The chain lengths are typically C16–C18, with trace amounts of chain lengths either side of this range (C14–C20). Palmitic acid (C16:0), oleic acid (C18:1 Δ9), and linoleic acid (C18:2 Δ9, 12) are the main fatty acid (FA) compounds in biodiesel. Dependent on the species, significant amounts of polyunsaturated fatty acids (PUFAs) may be present, such as α-linolenic acid (LNA) (C18:3 Δ9,12,15), γ-linolenic acid (GLA) (C18:3 Δ6,9,12) [89], arachidonic acid (AA) (20:4 Δ5,8,11,14), eicosapentaenoic acid (EPA) (20:5 Δ5,8,11,14,17), and docosahexaenoic acid (DHA) (22:6 Δ5,8,11,14,17,19) (Figure 3). While environmental conditions can affect the level of PUFAs to some extent, e.g., nitrogen starvation and elevated temperatures, it is likely that a significant reduction in these components will require genetic manipulation to reduce the expression of the desaturase enzymes responsible for double-bond formation. Some progress has been made in identifying these genes in algae, and this has recently been reviewed [90].
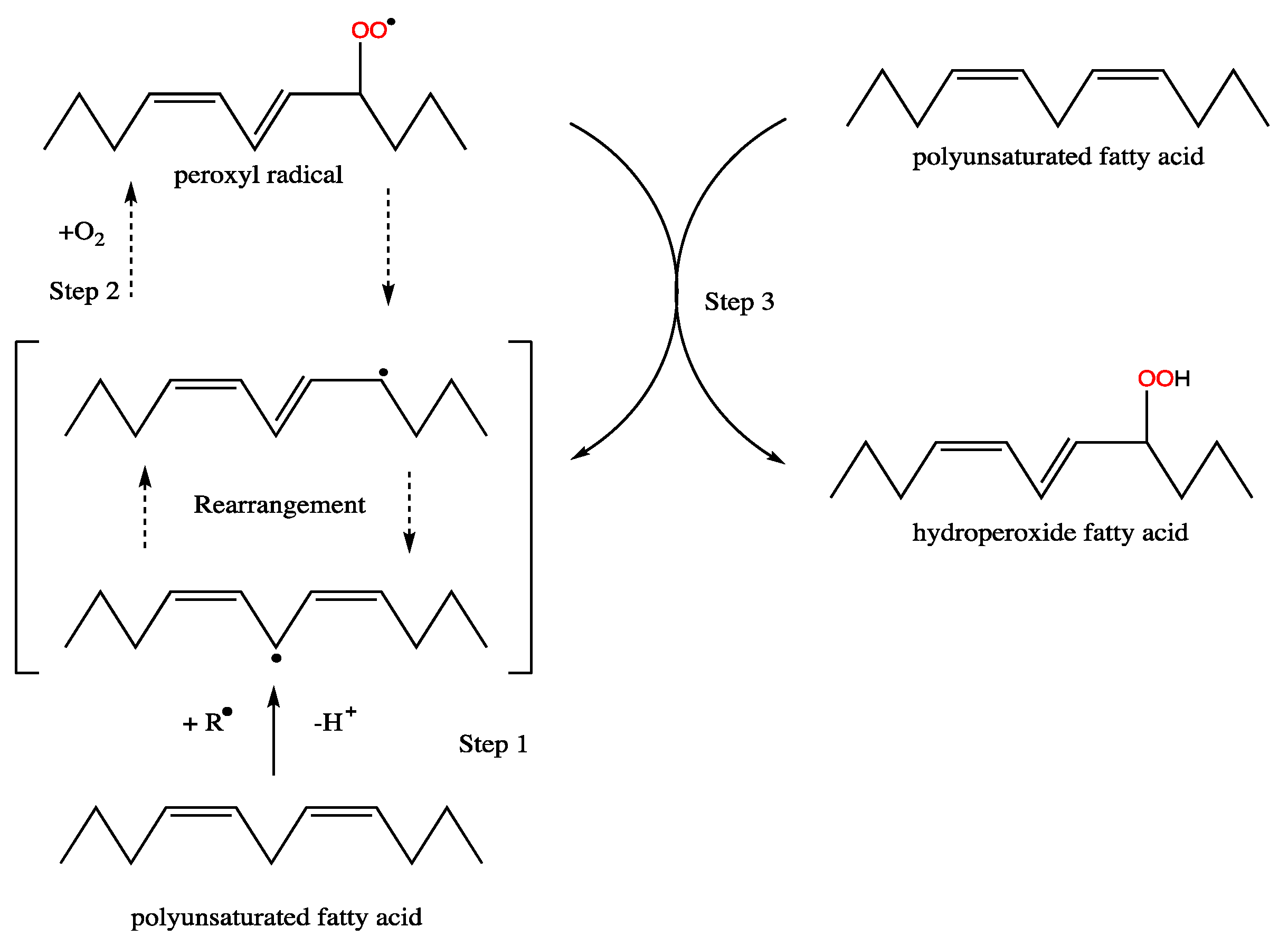
Polyunsaturated fatty acids, while of pharmaceutical value [91], are undesirable components of biodiesel due to their oxidative instability that results in the formation of short-chain aldehydes and ketones with corrosive characteristics [92]. Linoleic acid C18:2 (Δ9,12) is about 40 times more reactive than C18:1(Δ9) based on the relative rates of oxidation of a set of simple C18 unsaturated fatty acids, and α-18:3 (Δ9,12,15) is 2.4 times more reactive than C18:2 (Δ9,12) [93]. The oxidative stability of 18:2, 18:3, 20:4, and 22:6 is proportional to the number of bis-allylic motifs, revealing that the oxidising ability rises by around two-fold for each active bis-allylic methylene group, making 22:6 five times more unstable than 18:2. For each addition of C=C, two major oxygenated products are formed, which are classified into a series of derivatives depending on the oxygenation site [94]. The locations of proton abstraction and carbon radical production are determined by the number and position of methylene carbons. Regarding the carboxyl end of the molecule, radical rearrangement occurs in a radical shift of +2 or −2 to a carbon up-chain or down-chain from this original location [93]. The newly produced carbon radical core is subsequently attacked by molecular oxygen, leading to the production of a peroxy-radical (Figure 4). A high-oil-producing microalga rich in C18:1(Δ9) would be the optimal feedstock for biodiesel synthesis based on the oxidative stability.
5.6. Non-Triacylglycerol Lipid Sources from Microalgae
While the bulk of oils generated by algae are triacylglycerols, the ‘oil’ recovered from the Chlorophyte, Botryococcus braunii, is a significant exception [95]. This species accumulates a range of linear and monocyclic terpenes, termed botryococcenes, with up to ten compounds (CnH2n; n = 30–34) constituting ca. 25–40% of the dry weight of Botryococcus (Figure 5).
These compounds can be hydrocracked to generate 15% aviation fuel, 15% diesel fuel, and approximately 67% gasoline, with density and aromatic contents of aviation being achieved [16,96]. Potential applications of Botryococcus lipid products have been reviewed [97].
B. braunii has received significant attention as a potential SAF, and algae grown at IHI (Japan) have recently been processed by the Honeywell Eco-refining system in the USA to produce a blend to power a commercial flight (see Section 7 below).
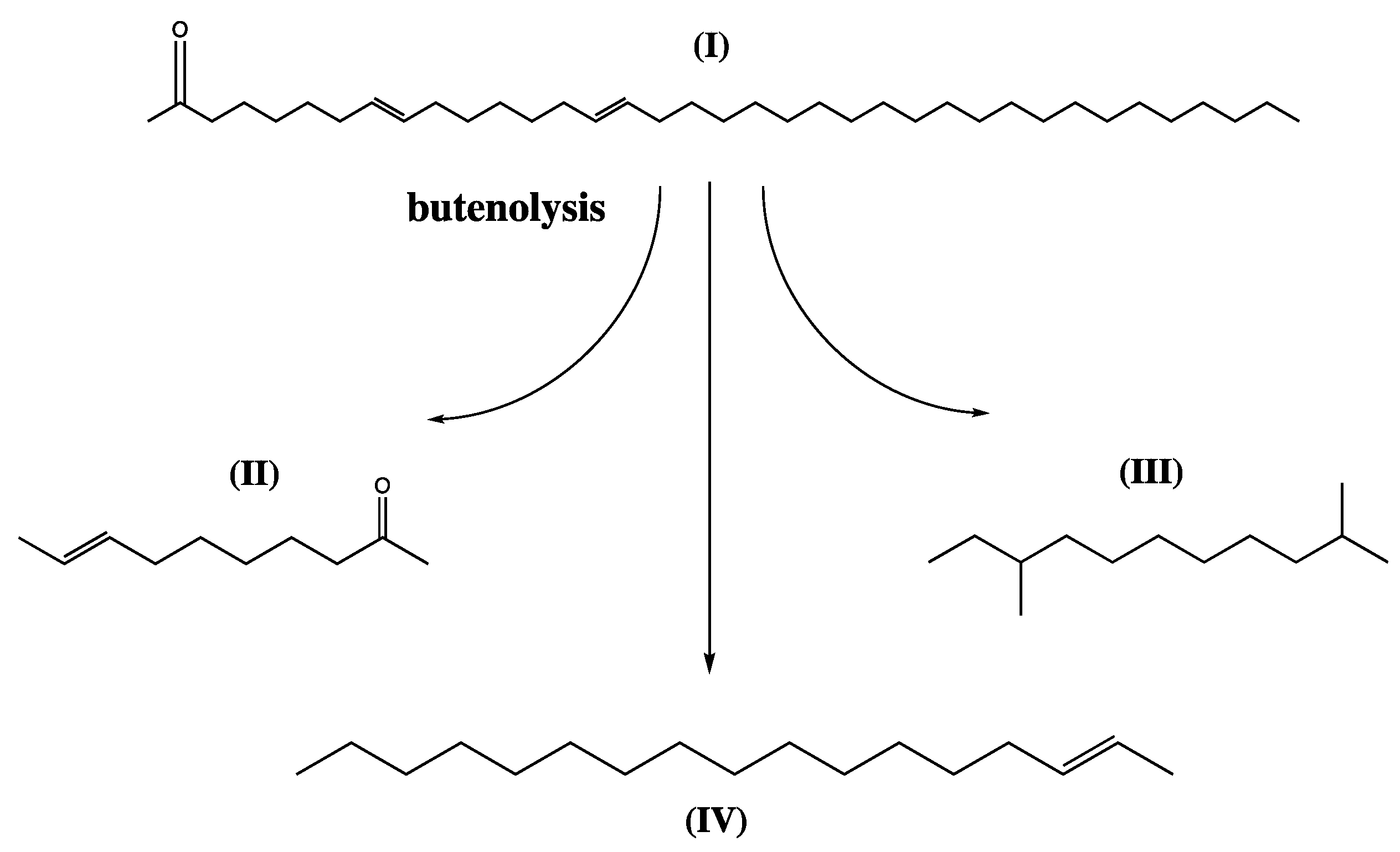
Another group of compounds restricted to a few species of the Prymnesiophyceae (Haptophyta) are long-chain alkenones [98]. The chain length is often in the C35–C41 region, which is roughly double that of conventional acyl fatty acids. Furthermore, they have trans double-bonds that are five carbons apart, as opposed to fatty acids’ methylene-interrupted systems, which likewise favour the cis orientation (Figure 3). Although alkenones are unsuitable for biodiesel production due to their high melting points (ca. 70 °C), butenolysis yields a range of jet fuel hydrocarbons, namely 2,9-undecane (43%), 2-heptadecene (28%), and 8-decen-2-one (12%) (Figure 6) [15].
6. Energy and Economics of Algae-Based Aviation Biofuels
One method of determining the competitiveness of a bio-based fuel for comparison with petroleum is to calculate the highest price that could possibly be paid for bio-oil, assuming that oil can be acquired at a particular price as a source of energy [99]. Equation (1) can be used to calculate the energy equivalent of biomass (in M, tonnes) equal to a barrel of crude oil:
where, EOil is the energy of a one barrel of oil in MJ, a is the volume of the biogas, b is the biodiesel yield, x is the oil content of the biomass in mass percentage, EBiogas is the energy of biogas in MJ/m3, and EBiodiesel is the average energy of the biodiesel fuel in MJ [99].
For the value of x chosen, M may be determined using Equation (1) (assuming that the cost of transporting one barrel of oil is roughly the same as the cost of transforming M tonnes of biomass into electricity). The maximum acceptable cost for biomass can be calculated from Equation (2):
To compete with fossil diesel, a biomass with 55% oil content, for example, should be produced for less than US$340/tonne [99]. According to current estimates, microalgal biomass may be generated for around US$3000 per tonne [99]. As a result, the cost of producing biomass must be decreased for biofuel generation from algae to be practical. It is important to note that this study disregards the potential of biomass residuals (such as high-value products) and the lower cost of converting biomass to oil [99,100].
Base commodities, such as fuels/energy, feed, and bioremediation, are considered high volume, low cost (<GBP£1/kg), while added-value chemicals can fetch GBP£1–£5, and speciality products GBP£5–£1000/kg, or in the case of nutraceutical/cosmeceutical applications, often >GBP£2000/kg. Fuel produced from vegetable sources such as Camelina, Soy, Jatropha, and waste cooking oils are, typically, <6 US$/gallon using HEFA processing (Table 11), compared to microalgae at US$31.98 per gallon. Ideally, if the microalgae produced a high-value-added product, such as a pigment or a protein with a value of US$2000–7000/kg, as well as a high oil content, then the economics of production would change significantly [101]. Commercial production of astaxanthin, for example, a xanthophyll carotenoid produced by the unicellular green alga, Haematococcus pluvialis, with a content of 1–5%, is the only commercially produced source of this pigment [102]. Other pigments include phycoerythrin and phycocyanin, which are protein–pigment complexes [103]. Other high-value lipids include polyunsaturated fatty acids and squalene [20]. Where high-value products are obtained, one could view the energy product (triacylglycerol) as a by-product.
7. Aviation Flights Undertaken Using Microalgal Biofuel
For more than a decade, several flights have been conducted using vegetable oils blended with those derived from microalgae. For example, in 2010 at the Farnborough Air Show, one of two Austro AE300 engines on an EADS demonstrator Diamond aircraft DA42 was powered by an algal-derived biofuel [104]. Exhaust emissions analysis revealed that up to eight times less hydrocarbons were emitted using the biofuel in comparison with the other engine powered by standard fuel. In 2021, All Nippon Airways (ANA), Japan’s largest airline, used a liquid hydrocarbon fuel obtained from hydrogenating crude oil extracted from Botryococcus braunii in a Boeing 787-8 flight between Tokyo and Osaka [12,28]. The blend was 3.8% algal-based mixed conventional fuel and met ASTM D7566 Annex7 specifications. The algal-derived oil was processed using UOP-Eco-refining™ technology and blended 50:50 with petroleum-based jet fuel. This is the only commercially demonstrated process for 100% SAF production currently in existence [12,28]. In 2022, Embraer’s E175 aircraft undertook a 1 h flight from Shizuoka to Nagoya using microalgal oil obtained from Euglena and blended with cooking oil [12,28]. The Euglena SAF, termed SUSTEO, is certified by ASTM International. Turkish Airlines also plan to test microalgal-derived SAF by the end of 2022 using oil obtained from its demonstration plant at Bogazici University in Istanbul [12,28]. No details of the algal species and what blends might be produced are currently available.
8. Future Work
To obtain an alga with all the desired qualities to produce biodiesel at scale will require the following characteristics: a fast growth rate and a high oil yield (>50%), with oleic acid as the principal fatty acid component. Factors that potentially influence this have been reviewed elsewhere [105]. The alga should also exhibit temperature tolerance (up to 50 °C) and be able to grow in salt water and/or polluted fresh water. This will require a detailed understanding on the underlaying mechanisms of salinity stress responses [106], metallic toxicity tolerance [107], and temperature adaptation [108]. The hunt for such an ideal species should be performed at sites where the organism will be grown at scale. For example, do any algae fulfilling these criteria already exist in the Gujarat area of India, which has been identified as a potential site for large-scale cultivation? High-throughput screening on microplates [44] could be performed to assess many of these features and combined with gas chromatography analysis of the lipids to assess the quantity and composition of the oils. Alternatively, environmental adaptive laboratory evolution of algal strain improvement could be performed on known species that already exhibit many of the features outlined above [109]. The possibility also exists to genetically modify traits for enhanced performance, as is the case with Nanochloropsis engineered with high levels of oils by ExxonMobil. Random DNA shuffling by using genetic backgrounds from multiple sources can be performed to produce enhanced characteristics where this feature is lacking in the existing gene pool [110]. While many other technical problems remain challenging, as outlined above, the ability to generate an alga with robust physiology and a high oil yield will prove valuable in advancing the technological innovation required to convert algal biodiesel into a real commercial success.
9. Conclusions
Based on fuel density and the requirement to not radically shift the aircraft design, biodiesel is an attractive alternative to kerosene. Based on the arguments around fuel versus food for land uses, oilseed crops are unfeasible. Since microalgae are more efficient in producing oil per hectare (by an order of magnitude), growing them in areas of current low productivity, such as deserts, close to coasts would be desirable. Ideally, there should be the least possible elevation from the sea to the site of production to minimise the cost of pumping seawater. This could be further facilitated by choosing sites where there is significant tidal range (i.e., the difference between high- and low-tide marks) that could offset pumping costs.
Strain selection, nutrient costs, harvesting, and oil optimisation are still key, but not insurmountable, challenges. Indeed, to replace 30% of fossil fuel with algal-derived biodiesel would require 11,345 km2 of land. This area could be further reduced with the development of new strains, as demonstrated recently, that are capable of higher biomass productivity by an order of magnitude on previous levels (c.f. 6.5 g/L to 0.6 g/L). Further yield enhancement could be achieved by using newer technologies, such as biofilm reactors, if costs for large-scale implementation could be reduced. Another key cost issue is the supply of carbon dioxide. To facilitate this would require the development of heavy industry plants at the sites of algal production, for example, cement works and steel manufacturing.
Finally, understanding the transcription control factors that regulate metabolite partitioning to oil or starch biosynthesis in microalgae will lead to the ability to engineer algae through biotechnological means. The challenges remain regrading biodiesel production from algae, but with investment to address them and the political will to implement them, these challenges could be overcome.
Author Contributions
Conceptualisation G.G., Algal biochemistry G.G., Fuel characteristics and engine performance sections V.S., A.K.H. and G.D. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the UKIERI project (Grant Number: DST-UKIERI 18-19-04): Waste to Energy—Low Temperature Combustion of Sustainable Green Fuels.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AA | Arachidonic acid |
| ATAG | Air Transport Action Group |
| ASTM | American Society for Testing and Materials |
| ATJ | Alcohol-to-jet |
| GHG | Greenhouse gas |
| AVGAS | Aviation gasoline |
| CO2 | Carbon dioxide |
| CO2 eq | Carbon dioxide equivalent |
| CORSIA | Carbon Offsetting and Reduction Scheme for International Aviation |
| CH | Catalytic hydrotreating |
| CCS-APR | Catalytic conversion of sugars by aqueous phase reforming |
| DAG | Diacylglycerol |
| DAGAT | Diacylglycerol acyltransferase |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| EU ETS | EU Emissions Trading Scheme |
| EFR | Effective radiative forcing |
| FAME | Fatty acid methyl esters |
| FT-SPK | Fischer-Tropsch synthetic paraffinic kerosene |
| GCAM | Global change assessment model |
| GLA | Gamma linolenic acid |
| HFS-SIP | Hydro-processing of fermented sugars–synthetic isoparaffinic kerosene |
| HEFA | Hydro-processed esters and fatty acids |
| ICAO | International Civil Aviation Organization |
| IEA | International Energy Agency |
| ILUC | Indirect land use change |
| LA | Linoleic acid |
| LNA | Linolenic acid |
| LPA | Lyso-phosphatidic acid |
| LPC | Lyso-phophatidylcholine |
| MAG | Monoacylglycerol |
| Mt | Million tonnes |
| MJSP | Minimum jet fuel selling price |
| PA | Phosphatidic acid |
| PC | Phosphatidylcholine |
| PDAT | Phosphatidylcholine: diacylglycerol acyltransferase |
| PUFAs | Polyunsaturated fatty acids |
| RF | Radiative forcing |
| SPK | Synthetic paraffinic kerosene |
| SAF | Sustainable aviation fuel |
| TAGs | Triacylglycerols |
| UCO | Used cooking oil |
References
- Lokke, S.; Aramendia, E.; Malskaer, J. A review of public opinion on liquid biofuels in the EU: Current knowledge and future challenges. Biomass Bioenergy 2021, 150, 106094. [Google Scholar]
- Varotsos, C.; Krapivin, V.; Mkrtchyan, F.; Zhou, X.R. On the effects of aviation on carbon-methane cycles and climate change during the period 2015–2100. Atmos. Pollut. Res. 2021, 12, 184–194. [Google Scholar]
- Chiaramonti, D. Sustainable Aviation Fuels: The challenge of decarbonization. Innov. Solut. Energy Transit. 2019, 158, 1202–1207. [Google Scholar]
- Chiaramonti, D.; Talluri, G.; Scarlat, N.; Prussi, M. The challenge of forecasting the role of biofuel in EU transport decarbonisation at 2050: A meta-analysis review of published scenarios. Renew. Sustain. Energy Rev. 2021, 139, 110715. [Google Scholar]
- Carlsson, F.; Hammar, H. Incentive-based regulation of CO2 emissions from international aviation. J. Air Transp. Manag. 2002, 8, 365–372. [Google Scholar]
- Hassan, M.; Pfaender, H.; Mavris, D. Probabilistic assessment of aviation CO2 emission targets. Transp. Res. Part D Transp. Environ. 2018, 63, 362–376. [Google Scholar]
- Bailis, R.E.; Bake, J.E. Greenhouse Gas Emissions and Land Use Change from Jatropha Curcas-Based Jet Fuel in Brazil. Environ. Sci. Technol. 2010, 44, 8684–8691. [Google Scholar]
- Lehr, U.; Nitsch, J.; Kratzat, M.; Lutz, C.; Edler, D. Renewable energy and employment in Germany. Energ. Policy 2008, 36, 108–117. [Google Scholar] [CrossRef]
- Agency, I.E. Market Report Series Renewables 2018, Analysis and Forecast to 2023. 2022. Available online: https://www.iea.org/reports/renewables-2018 (accessed on 25 May 2023).
- IATA Forecast Predicts 8.2 Billion Air Travelers in 2037. 2018. Available online: https://www.iata.org/en/pressroom/pr/2018-10-24-02/ (accessed on 20 March 2023).
- Abrantes, I.; Ferreira, A.F.; Silva, A.; Costa, M. Sustainable aviation fuels and imminent technologies—CO2 emissions evolution towards 2050. J. Clean. Prod. 2021, 313, 127937. [Google Scholar]
- Sustainable Aviation Fuel Report. Sustainable Aviation Fuel Road-Map 2020. 2020. Available online: https://www.sustainableaviation.co.uk/wp-content/uploads/2020/02/SustainableAviation_FuelReport_20200231.pdf (accessed on 1 April 2023).
- Biofuelwatch. ICAO Aviation Biofuels Report. 2017. Available online: https://www.biofuelwatch.org.uk/wp-content/uploads/Aviation-biofuels-report.pdf (accessed on 20 April 2023).
- CMR GWONGa. Jet Biofuels from Algae, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Mitici, M.; Pereira, M.; Oliviero, F. Electric flight scheduling with battery-charging and battery-swapping opportunities. EURO J. Transp. Logist. 2022, 11, 100074. [Google Scholar]
- O’Neil, G.W.; Culler, A.R.; Williams, J.R.; Burlow, N.P.; Gilbert, G.J.; Carmichael, C.A.; Nelson, R.K.; Swarthout, R.F.; Reddy, C.M. Production of Jet Fuel Range Hydrocarbons as a Coproduct of Algal Biodiesel by Butenolysis of Long-Chain Alkenones. Energy Fuels 2015, 29, 922–930. [Google Scholar]
- Paek, S.W.; Kim, S.; Raj, R.V. Optimal Endurance and Range of Electric Aircraft with Battery Degradation. Trans. Jpn. Soc. Aeronaut. Space Sci. 2020, 63, 62–65. [Google Scholar] [CrossRef]
- Bauen, A.; Bitossi, N.; German, L.; Harris, A.; Leow, K. Sustainable Aviation Fuels Status, challenges and prospects of drop-in liquid fuels, hydrogen and electrification in aviation. Johns. Matthey Technol. Rev. 2020, 64, 263–278. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, J. Estimation of Liquid Hydrogen Fuels in Aviation. Aerospace 2022, 9, 564. [Google Scholar]
- Ajjawi, I.; Verruto, J.; Aqui, M.; Soriaga, L.B.; Coppersmith, J.; Kwok, K.; Peach, L.; Orchard, E.; Kalb, R.; Xu, W.; et al. Lipid production in Nannochloropsis gaditana is doubled by decreasing expression of a single transcriptional regulator. Nat. Biotechnol. 2017, 35, 647–652. [Google Scholar]
- Aliyu, A.; Lee, J.G.M.; Harvey, A.P. Microalgae for biofuels via thermochemical conversion processes: A review of cultivation, harvesting and drying processes, and the associated opportunities for integrated production. Bioresour. Technol. Rep. 2021, 14, 100676. [Google Scholar]
- Deane, P.G.B.; Shea, R.O. Biofuels for Aviation: Technology Brief; International Renewable Energy Agency (IRENA): Masdar City, United Arab Emirates, 2017. [Google Scholar]
- Jia, T.H.; Zhang, X.W.; Liu, Y.; Gong, S.; Deng, C.; Pan, L.; Zou, J.-J. A comprehensive review of the thermal oxidation stability of jet fuels. Chem. Eng. Sci. 2021, 229, 116157. [Google Scholar]
- Qavi, I.; Jiang, L.L.; Akinyemi, O.S. Near-field spray characterization of a high-viscosity alternative jet fuel blend C-3 using a flow blurring injector. Fuel 2021, 293, 120350. [Google Scholar]
- Beal, C.M.; Cuellar, A.D.; Wagner, T.J. Sustainability assessment of alternative jet fuel for the U.S. Department of Defense. Biomass Bioenergy 2021, 144, 105881. [Google Scholar] [CrossRef]
- Dutta, S.; Madav, V.; Joshi, G.; Naik, N.; Kumar, S. Directional synthesis of aviation-, diesel-, and gasoline range hydrocarbon fuels by catalytic transformations of biomass components: An overview. Fuel 2023, 347, 128437. [Google Scholar]
- Wang, W.C.; Tao, L. Bio-jet fuel conversion technologies. Renew. Sustain. Energy Rev. 2016, 53, 801–822. [Google Scholar]
- US Department of Energy; Sustainable Aviation Fuel; Review of Technical Pathways. 2020. Available online: https://www.energy.gov/sites/prod/files/2020/09/f78/beto-sust-aviation-fuel-sep-2020.pdf (accessed on 4 March 2023).
- Vozka, P.; Kilaz, G. A review of aviation turbine fuel chemical composition-property relations. Fuel 2020, 268, 117391. [Google Scholar]
- Wei, H.J.; Liu, W.Z.; Chen, X.Y.; Yang, Q.; Li, J.S.; Chen, H.P. Renewable bio-jet fuel production for aviation: A review. Fuel 2019, 254, 115599. [Google Scholar]
- Kang, D.; Kim, D.; Kalaskar, V.; Violi, A.; Boehman, A.L. Experimental characterization of jet fuels under engine relevant conditions-Part 1: Effect of chemical composition on autoignition of conventional and alternative jet fuels. Fuel 2019, 239, 1388–1404. [Google Scholar]
- Ardo, F.M.; Lim, J.W.; Ramli, A.; Lam, M.K.; Kiatkittipong, W.; Abdelfattah, E.A.; Shahid, M.K.; Usman, A.; Wongsakulphasatch, S.; Sahrin, N.T. A review in redressing challenges to produce sustainable hydrogen from microalgae for aviation industry. Fuel 2022, 330, 125646. [Google Scholar]
- Prussi, M.; O’Connell, A.; Lonza, L. Analysis of current aviation biofuel technical production potential in EU28. Biomass Bioenergy 2019, 130, 105371. [Google Scholar] [CrossRef]
- Mathimani, T.; Mallick, N. A comprehensive review on harvesting of microalgae for biodiesel—Key challenges and future directions. Renew. Sustain. Energy Rev. 2018, 91, 1103–1120. [Google Scholar]
- Rajvanshi, S.; Sharma, M.P. Micro Algae: A Potential Source of Biodiesel. J. Sustain. Bioenergy Syst. 2012, 2, 49–59. [Google Scholar]
- Shokravi, Z.; Shokravi, H.; Atabani, A.E.; Lau, W.J.; Chyuan, O.H.; Ismail, A.F. Impacts of the harvesting process on microalgae fatty acid profiles and lipid yields: Implications for biodiesel production. Renew. Sustain. Energy Rev. 2022, 161, 112410. [Google Scholar]
- Fasaei, F.; Bitter, J.H.; Slegers, P.M.; van Boxtel, A.J.B. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018, 31, 347–362. [Google Scholar]
- Lin, C.H.; Chen, Y.K.; Wang, W.C. The production of bio-jet fuel from palm oil derived alkanes. Fuel 2020, 260, 116345. [Google Scholar]
- Doliente, S.S.; Narayan, A.; Tapia, J.F.D.; Samsatli, N.J.; Zhao, Y.R.; Samsatli, S. Bio-aviation Fuel: A Comprehensive Review and Analysis of the Supply Chain Components. Front. Energy Res. 2020, 8, 110. [Google Scholar]
- Yu, W.B.; Zhao, F.Y. Formulating of model-based surrogates of jet fuel and diesel fuel by an intelligent methodology with uncertainties analysis. Fuel 2020, 268, 117393. [Google Scholar]
- Bosnjakovic, M.; Sinaga, N. The Perspective of Large-Scale Production of Algae Biodiesel. Appl. Sci. 2020, 10, 8181. [Google Scholar]
- Zheng, X.S.R.D. Fuel Burn of New Commercial Jet Aircraft: 1960 to 2019; Int Counc Clean Transp: Washington, DC, USA, 2020. [Google Scholar]
- Bwapwa, J.K.; Anandraj, A.; Trois, C. Possibilities for conversion of microalgae oil into aviation fuel: A review. Renew. Sustain. Energy Rev. 2017, 80, 1345–1354. [Google Scholar]
- Griffiths, G.; Hossain, A.K.; Sharma, V.; Duraisamy, G. Key Targets for Improving Algal Biofuel Production. Clean. Technol. 2021, 3, 711–742. [Google Scholar]
- Li, P.Y.; Wang, X.; Luo, Y.Q.; Yuan, X.G. Sustainability evaluation of microalgae biodiesel production process integrated with nutrient close-loop pathway based on emergy analysis method. Bioresour. Technol. 2022, 346, 126611. [Google Scholar]
- Patnaik, R.; Mallick, N. Microalgal Biodiesel Production: Realizing the Sustainability Index. Front. Bioeng. Biotech. 2021, 9, 620777. [Google Scholar]
- Craggs, R.J.; Heubeck, S.; Lundquist, T.J.; Benemann, J.R. Algal biofuels from wastewater treatment high rate algal ponds. Water Sci. Technol. 2011, 63, 660–665. [Google Scholar]
- Zhang, L.; Wang, S.; Han, J.C.; Yang, G.P.; Pan, K.H.; Xu, J.L. Manipulation of triacylglycerol biosynthesis in Nannochloropsis oceanica by overexpressing an Arabidopsis thaliana diacylglycerol acyltransferase gene. Algal Res. 2022, 61, 102590. [Google Scholar]
- Paul, T.; Sinharoy, A.; Baskaran, D.; Pakshirajan, K.; Pugazhenthi, G.; Lens, P.N.L. Bio-oil production from oleaginous microorganisms using hydrothermal liquefaction: A biorefinery approach. Crit. Rev. Environ. Sci. Technol. 2022, 52, 356–394. [Google Scholar] [CrossRef]
- Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13601. [Google Scholar]
- Chu, R.Y.; Hu, D.; Zhu, L.D.; Li, S.X.; Yin, Z.H.; Yu, Y.J. Recycling spent water from microalgae harvesting by fungal pellets to re-cultivate Chlorella vulgaris under different nutrient loads for biodiesel production. Bioresour. Technol. 2022, 344, 126227. [Google Scholar] [CrossRef] [PubMed]
- Vasistha, S.; Khanra, A.; Clifford, M.; Rai, M.P. Current advances in microalgae harvesting and lipid extraction processes for improved biodiesel production: A review. Renew. Sustain. Energy Rev. 2021, 137, 110498. [Google Scholar]
- Song, X.T.; Kong, F.Y.; Liu, B.F.; Song, Q.Q.; Ren, N.Q.; Ren, H.Y. Thallium-mediated NO signaling induced lipid accumulation in microalgae and its role in heavy metal bioremediation. Water Res. 2023, 239, 120027. [Google Scholar]
- Mofijur, M.; Ashrafur Rahman, S.M.; Nguyen, L.N.; Mahlia, T.M.I.; Nghiem, L.D. Selection of microalgae strains for sustainable production of aviation biofuel. Bioresour. Technol. 2022, 345, 126408. [Google Scholar] [CrossRef]
- Ahn, Y.; Park, S.; Ji, M.K.; Ha, G.S.; Jeon, B.H.; Choi, J. Biodiesel production potential of microalgae, cultivated in acid mine drainage and livestock wastewater. J. Environ. Manage. 2022, 314, 115031. [Google Scholar]
- Patlakas, P.; Stathopoulos, C.; Flocas, H.; Kalogeri, C.; Kallos, G. Regional Climatic Features of the Arabian Peninsula. Atmosphere 2019, 10, 220. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Ali, S.S.; Ramadan, H.; El-Aswar, E.I.; Eltawab, R.; Ho, S.-H.; Elsamahy, T.; Li, S.; El-Sheekh, M.M.; Schagerl, M.; et al. Microalgae-based wastewater treatment: Mechanisms, challenges, recent advances, and future prospects. Environ. Sci. Ecotechnol. 2023, 13, 100205. [Google Scholar]
- Awad, N.; Vega-Estévez, S.; Griffiths, G. Salicylic acid and aspirin stimulate growth of Chlamydomonas and inhibit lipoxygenase and chloroplast desaturase pathways. Plant Physiol. Biochem. 2020, 149, 256–265. [Google Scholar] [CrossRef]
- Arora, K.; Kaur, P.; Kumar, P.; Singh, A.; Patel, S.K.S.; Li, X.; Yang, Y.-H.; Bhatia, S.K.; Kulshrestha, S. Valorization of Wastewater Resources Into Biofuel and Value-Added Products Using Microalgal System. Front. Energy Res. 2021, 9, 646571. [Google Scholar]
- Muhammad, G.; Alam, M.A.; Mofijur, M.; Jahirul, M.I.; Lv, Y.; Xiong, W.; Ong, H.C.; Xu, J. Modern developmental aspects in the field of economical harvesting and biodiesel production from microalgae biomass. Renew. Sustain. Energy Rev. 2021, 135, 110209. [Google Scholar]
- Kim, B.-H.; Choi, J.-E.; Cho, K.; Kang, Z.; Ramanan, R.; Moon, D.-G.; Kim, H.-S. Influence of Water Depth on Microalgal Production, Biomass Harvest, and Energy Consumption in High Rate Algal Pond Using Municipal Wastewater. J. Microbiol. Biotechnol. 2018, 28, 630–637. [Google Scholar] [CrossRef]
- Singh, G.; Patidar, S.K. Microalgae harvesting techniques: A review. J. Environ. Manag. 2018, 217, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Suganya, T.; Varman, M.; Masjuki, H.H.; Renganathan, S. Macroalgae and microalgae as a potential source for commercial applications along with biofuels production: A biorefinery approach. Renew. Sustain. Energy Rev. 2016, 55, 909–941. [Google Scholar]
- Bharathiraja, B.; Iyyappan, J.; Gopinath, M.; Jayamuthunagai, J.; PraveenKumar, R. Transgenicism in algae: Challenges in compatibility, global scenario and future prospects for next generation biofuel production. Renew. Sustain. Energy Rev. 2022, 154, 111829. [Google Scholar] [CrossRef]
- Bellou, S.; Baeshen, M.N.; Elazzazy, A.M.; Aggeli, D.; Sayegh, F.; Aggelis, G. Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 2014, 32, 1476–1493. [Google Scholar] [CrossRef]
- Sun, X.M.; Geng, L.J.; Ren, L.J.; Ji, X.J.; Hao, N.; Chen, K.Q.; Huang, H. Influence of oxygen on the biosynthesis of polyunsaturated fatty acids in microalgae. Bioresour. Technol. 2018, 250, 868–876. [Google Scholar] [CrossRef]
- Carlsson, A.S.; Yilmaz, J.L.; Green, A.G.; Stymne, S.; Hofvander, P. Replacing fossil oil with fresh oil—With what and for what? Eur. J. Lipid Sci. Technol. 2011, 113, 812–831. [Google Scholar] [CrossRef]
- Xu, J.; Li, T.; Li, C.-L.; Zhu, S.-N.; Wang, Z.-M.; Zeng, E.Y. Lipid accumulation and eicosapentaenoic acid distribution in response to nitrogen limitation in microalga Eustigmatos vischeri JHsu-01 (Eustigmatophyceae). Algal Res. 2020, 48, 101910. [Google Scholar] [CrossRef]
- Song, X.T.; Liu, B.F.; Kong, F.Y.; Song, Q.Q.; Ren, N.Q.; Ren, H.Y. Lipid accumulation by a novel microalga Parachlorella kessleri R-3 with wide pH tolerance for promising biodiesel production. Algal Res. 2023, 69, 102925. [Google Scholar] [CrossRef]
- Osorio, J.H.M.; Pollio, A.; Frunzo, L.; Lens, P.N.L.; Esposito, G. A Review of Microalgal Biofilm Technologies: Definition, Applications, Settings and Analysis. Front. Chem. Eng. 2021, 3, 737710. [Google Scholar] [CrossRef]
- Fica, Z.T.; Sims, R.C. Algae-based biofilm productivity utilizing dairy wastewater: Effects of temperature and organic carbon concentration. J. Biol. Eng. 2016, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Q.; Zhang, L.J.; Xu, G.; Li, F.; Li, X.K. A review on biodiesel production from microalgae: Influencing parameters and recent advanced technologies. Front. Microbiol. 2022, 13, 737710. [Google Scholar] [CrossRef]
- Mediboyina, M.K.; Banuvalli, B.K.; Chauhan, V.S.; Mudliar, S.N. Comparative life cycle assessment of autotrophic cultivation of Scenedesmus dimorphus in raceway pond coupled to biodiesel and biogas production. Bioprocess. Biosyst. Eng. 2020, 43, 233–247. [Google Scholar] [CrossRef]
- Arguelles, E.D.L.R. NITROGEN STARVATION INDUCED LIPID ACCUMULATION BY Chlorococcum infusionum (EAU-10) AS POTENTIAL RENEWABLE SOURCE OF LIPID FOR BIODIESEL PRODUCTION. J. Microbiol. Biotechnol. Food Sci. 2022, 11, e1931. [Google Scholar] [CrossRef]
- Chu, F.; Cheng, J.; Zhang, X.; Ye, Q.; Chen, S.; Zhou, J.; Cen, K. Transcriptome and key gene expression related to carbon metabolism and fatty acid synthesis of Chlorella vulgaris under a nitrogen starvation and phosphorus repletion regime. J. Appl. Phycol. 2019, 31, 2881–2893. [Google Scholar] [CrossRef]
- Mou, Y.; Liu, N.; Su, K.; Li, X.; Lu, T.; Yu, Z.; Song, M. The growth and lipid accumulation of Scenedesmus quadricauda under nitrogen starvation stress during xylose mixotrophic/heterotrophic cultivation. Environ. Sci. Pollut. Res. 2022, 30, 98934–98946. [Google Scholar] [CrossRef]
- Valledor, L.; Furuhashi, T.; Recuenco-Muñoz, L.; Wienkoop, S.; Weckwerth, W. System-level network analysis of nitrogen starvation and recovery in Chlamydomonas reinhardtii reveals potential new targets for increased lipid accumulation. Biotechnol. Biofuels 2014, 7, 171. [Google Scholar] [CrossRef]
- Chungjatupornchai, W.; Fa-aroonsawat, S. Enhanced triacylglycerol production in oleaginous microalga Neochloris oleoabundans by co-overexpression of lipogenic genes: Plastidial LPAAT1 and ER-located DGAT2. J. Biosci. Bioeng. 2021, 131, 124–130. [Google Scholar] [CrossRef]
- Azizi, S.; Bayat, B.; Tayebati, H.; Hashemi, A.; Pajoum Shariati, F. Nitrate and phosphate removal from treated wastewater by Chlorella vulgaris under various light regimes within membrane flat plate photobioreactor. Env. Prog. Sustain. 2021, 40, e13519. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [PubMed]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Sci. Rep. 2019, 9, 13935. [Google Scholar] [CrossRef] [PubMed]
- Jareonsin, S.; Pumas, C. Advantages of Heterotrophic Microalgae as a Host for Phytochemicals Production. Front. Bioeng. Biotech. 2021, 9, 628597. [Google Scholar]
- Shi, M.C.; Yu, L.H.; Shi, J.A.; Liu, J. A conserved MYB transcription factor is involved in regulating lipid metabolic pathways for oil biosynthesis in green algae. New Phytol. 2022, 235, 576–594. [Google Scholar] [CrossRef]
- Sagun, J.V.; Yadav, U.P.; Alonso, A.P. Progress in understanding and improving oil content and quality in seeds. Front. Plant Sci. 2023, 14, 1116894. [Google Scholar]
- Zafar, S.; Li, Y.-L.; Li, N.-N.; Zhu, K.-M.; Tan, X.-L. Recent advances in enhancement of oil content in oilseed crops. J. Biotechnol. 2019, 301, 35–44. [Google Scholar]
- Sun, X.-M.; Ren, L.-J.; Zhao, Q.-Y.; Ji, X.-J.; Huang, H. Enhancement of lipid accumulation in microalgae by metabolic engineering. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 552–566. [Google Scholar]
- Brar, A.; Kumar, M.; Soni, T.; Vivekanand, V.; Pareek, N. Insights into the genetic and metabolic engineering approaches to enhance the competence of microalgae as biofuel resource: A review. Bioresour. Technol. 2021, 339, 125597. [Google Scholar]
- Riaz, I.; Shafiq, I.; Jamil, F.; Al-Muhtaseb, A.H.; Akhter, P.; Shafique, S.; Park, Y.-K.; Hussain, M. A review on catalysts of biodiesel (methyl esters) production. Catal. Rev. 2022, 1–53. [Google Scholar] [CrossRef]
- Acien, F.G.; Fernandez, J.M.; Magan, J.J.; Molina, E. Production cost of a real microalgae production plant and strategies to reduce it. Biotechnol. Adv. 2012, 30, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.F.; Südfeld, C.; Naduthodi, M.I.; Weusthuis, R.A.; Barbosa, M.J.; Wijffels, R.H.; D’adamo, S. Genetic engineering of microalgae for enhanced lipid production. Biotechnol. Adv. 2021, 52, 107836. [Google Scholar]
- Fivga, A.; Speranza, L.G.; Branco, C.M.; Ouadi, M.; Hornung, A. A review on the current state of the art for the production of advanced liquid biofuels. Aims Energy 2019, 7, 46–76. [Google Scholar]
- Avdeev, Y.G.; Kuznetsov, Y.I. Inhibitor protection of steel corrosion in acid solutions at high temperatures. A review. Part 2. Int. J. Corros. Scale Inhib. 2020, 9, 867–902. [Google Scholar]
- Frankel, E.N. Lipid Oxidation; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Shahidi, F.; Hossain, A. Role of Lipids in Food Flavor Generation. Molecules 2022, 27, 5014. [Google Scholar] [CrossRef]
- Cheng, P.; Okada, S.; Zhou, C.; Chen, P.; Huo, S.; Li, K.; Addy, M.; Yan, X.; Ruan, R.R. High-value chemicals from Botryococcus braunii and their current applications—A review. Bioresour. Technol. 2019, 291, 121911. [Google Scholar]
- Chacko, A.R.; Amster, D.E.; Johnson, T.E.; Newman, S.R.; Gladchuk, A.V.; Sohn, C.J.; Prunkard, D.E.; Yakelis, N.A.; Freeman, J.O. High-throughput screen for sorting cells capable of producing the biofuel feedstock botryococcene. Org. Biomol. Chem. 2019, 17, 3195–3201. [Google Scholar] [CrossRef]
- Estevam, B.R.; Pinto, L.F.R.; Maciel, R.; Fregolente, L.V. Potential applications of Botryococcus terribilis: A review. Biomass Bioenergy 2022, 165, 106582. [Google Scholar]
- Liao, S.A.; Wang, K.J.; Huang, Y.S. Extended chain length alkenoates differentiate three Isochrysidales groups. Org. Geochem. 2021, 161, 104303. [Google Scholar] [CrossRef]
- Yin, S.; Jin, W.; Zhou, X.; Han, W.; Gao, S.; Chen, C.; Ding, W.; He, Z.; Chen, Y.; Jiang, G. Enhancing harvest of biodiesel-promising microalgae using Daphnia domesticated by amino acids. Environ. Res. 2022, 212, 113465. [Google Scholar]
- Lin, W.Z.; Chen, L.N.; Tan, Z.X.; Deng, Z.Q.; Liu, H. Application of filamentous fungi in microalgae-based wastewater remediation for biomass harvesting and utilization: From mechanisms to practical application. Algal Res. 2022, 62, 102614. [Google Scholar] [CrossRef]
- Said, Z.; Nguyen, T.H.; Sharma, P.; Li, C.; Ali, H.M.; Nguyen, V.N.; Pham, V.V.; Ahmed, S.F.; Van, D.N.; Truong, T.H. Multi-attribute optimization of sustainable aviation fuel production-process from microalgae source. Fuel 2022, 324, 124759. [Google Scholar] [CrossRef]
- Haghpanah, T.; Sobati, M.A.; Pishvaee, M.S. Multi-objective superstructure optimization of a microalgae biorefinery considering economic and environmental aspects. Comput. Chem. Eng. 2022, 164, 107894. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Y.; He, Y.; Liu, B.; Mou, H.; Chen, F.; Yang, S. Microalgae-Derived Pigments for the Food Industry. Mar. Drugs 2023, 21, 82. [Google Scholar] [CrossRef]
- Vazquez-Romero, B.; Perales, J.A.; Pereira, H.; Barbosa, M.; Ruiz, J. Techno-economic assessment of microalgae production, harvesting and drying for food, feed, cosmetics, and agriculture. Sci. Total Environ. 2022, 837, 155742. [Google Scholar] [CrossRef]
- Morales, M.; Aflalo, C.; Bernard, O. Microalgal lipids: A review of lipids potential and quantification for 95 phytoplankton species. Biomass Bioenergy 2021, 150, 106108. [Google Scholar]
- Shetty, P.; Gitau, M.M.; Maróti, G. Salinity Stress Responses and Adaptation Mechanisms in Eukaryotic Green Microalgae. Cells 2019, 8, 1657. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.-S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.-P.; Han, B.; Yu, X. Coupling of abiotic stresses and phytohormones for the production of lipids and high-value by-products by microalgae: A review. Bioresour. Technol. 2019, 274, 549–556. [Google Scholar] [CrossRef]
- LaPanse, A.J.; Krishnan, A.; Posewitz, M.C. Adaptive Laboratory Evolution for algal strain improvement: Methodologies and applications. Algal Res. 2021, 53, 102122. [Google Scholar] [CrossRef]
- Fajardo, C.; De Donato, M.; Carrasco, R.; Martínez-Rodríguez, G.; Mancera, J.M.; Fernández-Acero, F.J. Advances and challenges in genetic engineering of microalgae. Rev. Aquac. 2020, 12, 365–381. [Google Scholar] [CrossRef]
Figure 1.
An overview of the cultivation, extraction, and preparation of aviation biodiesel from microalgae.
Figure 1.
An overview of the cultivation, extraction, and preparation of aviation biodiesel from microalgae.

Figure 2.
Biosynthesis of triacylglycerols in algae.

Figure 3.
Structures of major fatty acids in microalgae.

Figure 4.
Radical formation and double-bond rearrangement in fatty acids during formation of hydroperoxides.
Figure 4.
Radical formation and double-bond rearrangement in fatty acids during formation of hydroperoxides.
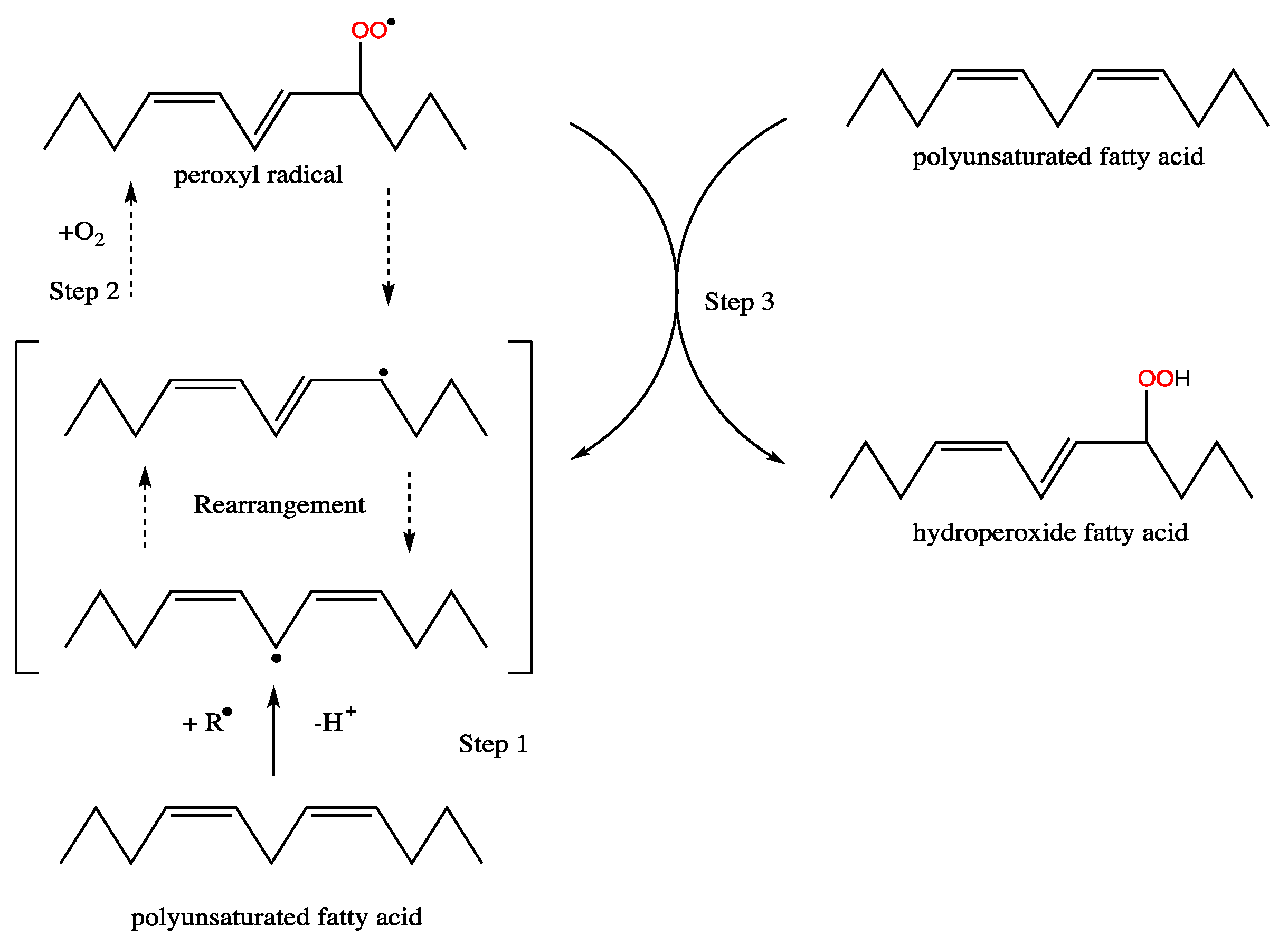
Figure 5.
Structure of a C34 botryococcene.

Figure 6.
Butenolysis of alkenones (I) generates a range of products, predominantly (II) 8-decen-2-one 2,9-dimethyl undecane (III) and 2-heptadene (IV).
Figure 6.
Butenolysis of alkenones (I) generates a range of products, predominantly (II) 8-decen-2-one 2,9-dimethyl undecane (III) and 2-heptadene (IV).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Regulatory agency and standards of aviation fuel around the world [25].
Table 1.
Regulatory agency and standards of aviation fuel around the world [25].
| Regulating Agency | Country | Standards/Resolution | Commercial Name |
|---|---|---|---|
| Agência Nacional do Petróleo, Gás Natural Biocombustíveis | Brazil | Resolution No. 37 | Jet A-1 |
| Federal Aviation Administration | USA | ASTM D1655/ASTM 6615 | Jet A, Jet A1/Jet B |
| Transport Canada Civil Aviation | Canada | CAN/CGSB-3.23 CAN/CGSB-3.22 | Jet A/A1/Jet B |
| Civil Aviation Authority | UK | DefStan 91–91 | Jet A1 |
| European Aviation Safety Agency | EU | AFQRJOS | Jet A1 |
| Federal Air Transport Agency | Russia | GOST 10,227/GOST R 52,050 | TS-1/Jet A1 |
| Civil Aviation Administration of China | China | GB 6537 | No. 3 |
| Properties | Unit | ASTM 1655-4a | Def Stan 91-91 | ANPn°37 |
|---|---|---|---|---|
| Density | g/mL | 0.775–0.840 | 0.775–0.840 | 0.771–0.836 (20 °C) |
| Viscosity at 20 °C | mm2/s | 8.0 max | 8.0 max | 8.0 max |
| Acid value | mgKOH/g | 0.100 | 0.0012 | 0.015 |
| Flash point | °C | 38 min | 38 min | 38–40 min |
| Heat of combustion | MJ/kg | 42.8 min | 42.8 min | 42.8 min |
| Freezing point | °C | −47 | −47 | −47 |
| Sulphur | % | 0.3 | 0.3 | 0.3 |
| Aromatics | % | 25 | 25 | 25 |
| Smoke point | Mm | 25 min | 25 min | 25 min |
| JFTOT Delta P (260 °C) | mmHg | 25 | 25 | 25 |
| Conductivity | pS/m | 50–450 | 50–600 | 50–600 |
| Maximum boiling point | °C | 300 max | 300 max | 300 max |
| Compound | Formula | Type | Chemical Structure |
|---|---|---|---|
| n-octane | C8H18 | n-paraffin |  |
| 2-Methylheptane | C8H18 | Isoparaffin | 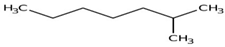 |
| 1-Methyl-1-ethylcyclopentane | C8H14 | Cycloparaffin |  |
| Ethyl-cyclohexane | C8H16 | Cycloparaffin | 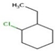 |
| o-Xylene | C8H10 | Aromatic | 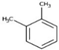 |
| p-Xylene | C8H10 | Aromatic | 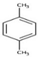 |
| Cis-Decalin | C10H18 | Cycloparaffin | 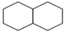 |
| Tetralin | C10H12 | Aromatic |  |
| Naphthalene | C10H8 | Aromatic | 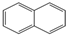 |
| n-Dodecane | C12H26 | n-paraffin | 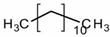 |
| 2-Methylundecane | C12H26 | Isoparaffin | 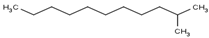 |
| 1-Ethylnaftalene | C12H12 | Aromatic | 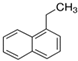 |
| n-Hexylbenzene | C12H18 | Aromatic | 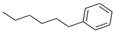 |
| n-Hexadecane | C16H34 | n-paraffin | 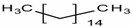 |
| 2-Methylpentadecane | C16H34 | Isoparaffin | 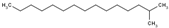 |
| n-Decylbenzene | C16H26 | Aromatic |  |
Table 4.
Sustainable aviation fuel (SAF) pathways [33].
Table 4.
Sustainable aviation fuel (SAF) pathways [33].
| Methods | Description |
|---|---|
| FT-SPK | Fischer-Tropsch synthetic paraffinic kerosene (FT-SPK). Biomass is converted into syngas and then biofuels via the FT process. ASTM approved the approach in 2009, and the UK MOD Def-Stan (91-91) approved it in 2010. FT-SPK aviation biofuel can be blended up to 50% with fossil jet fuel. |
| HEFA | Hydro-processed fatty acid esters and free fatty acid (HEFA). Hydrogen is used to transform liquid feedstock, including vegetable oils, cooking oil, and tallow, into green diesel, which can then be isomerised and separated to produce a jet fraction. In 2011, the route was certified for a 50% blend with fossil jet fuel. |
| HFS-SIP | Hydro-processing of fermented sugars–synthetic isoparaffinic kerosene (HFS-SIP). Sugars can be transformed to hydrocarbons using modified yeasts. The current permitted technique creates a C15 hydrocarbon terpenoid, farnesene. ASTM authorised this technology in 2014, and it can be combined with fossil jet fuel up to 10%. |
| FT-SPK/A | This is a modified FT-SPK process. Light aromatics are alkylated to yield a hydrocarbon mix with an aromatic component. This method was authorised in 2015 and can blend up to 50%. |
| ATJ-SPK | Alcohol-to-jet-synthetic paraffinic kerosene (ATJ-SPK). Hydro-processing, dehydration, and oligomerisation are used to convert alcohols (iso-butanol) into hydrocarbon. A certified process allows a maximum 50% blending. |
| Co-processing | Biological liquid feedstock, such as fats, oil, and other residues, can be blended with fossil crude oil by 5% (v/v) to carry out the refining process. This process was approved in April 2018 by ASTM and certified with ASTM D1655. |
| CCS-APR | Catalytic conversion of sugars by aqueous phase reforming. |
| CH | Catalytic hydrotreating of liquid to jet fuels. |
| CATJ-SKA | Catalytic upgrading of alcohol intermediate-catalytic ATJ-synthetic kerosene with aromatics. |
| ATJ-SPK expansion | Catalytic upgrading of ethanol. |
| HEFA expansion | Direct use of a wider cut of HEFA with renewable diesel. |
| HDCJ UOP-Eco-refining™ | Pyrolysis-hydrotreated de-polymerised cellulose. Blending vegetable biodiesels with petroleum-based fuels. |
| Microalgal Species | Oil Content (wt. % Dry Weight) |
|---|---|
| Schizochytrium sp. | 50–77 |
| Botryococcus braunii | 25–75 |
| Nannochloropsis sp. | 31–68 |
| Neochloris oleoabundans | 35–54 |
| Nitzschia sp. | 45–47 |
| Cylindrotheca sp. | 16–37 |
| Nannochloris sp. | 20–35 |
| Isochrysis sp. | 25–33 |
| Phaeodactylum tricornutum | 20–30 |
| Crop | Oil (L/Hectare) |
|---|---|
| Microalgae | 95,200 |
| Palm | 5950 |
| Jatropha | 4348 |
| Coconut | 2689 |
| Castor | 1413 |
| Sunflower | 952 |
| Tung | 640 |
| Soy | 446 |
| Oil Content of Algal Cells (% Dry Weight) | Oil Yield L/ha/Year | Biodiesel L/ha/Year | Land Area (km2) Required for 30% Replacement of Fossil Fuel |
|---|---|---|---|
| 30 | 58,700 | 51,927 | 11,345 |
| 50 | 97,833 | 86,582 | 6805 |
| 70 | 136,967 | 121,104 | 4863 |
| Name | Type | Area | Temp. Range (°C) | Rainfall (mm/Year) | Topography |
|---|---|---|---|---|---|
| Antarctic | Polar | 14,000,000 | +10 to −89 | 50–165 | Almost entirely covered in ice sheets. |
| Sahara | Subtropical | 9,200,000 | +47 to +5 | 25–120 | Extensive gravel-covered plains, abrupt mountains, sand dunes, large oasis depressions, and shallow seasonally inundated basins. |
| Great Australian | Subtropical | 2,700,000 | +40 to 16 | 40–300 | Salt lakes, sandy areas, grassy plains, and rocky areas. |
| Arabian | Subtropical | 2,330,000 | +55 to +8 | 20–500 | Vast light-coloured sand terrain, mountain ranges, black lava flows, and desert dunes. |
| Gobi | Interior | 1,295,000 | +45 to −40 | 152–194 | Cold desert with frost on plateau, 900–1500 m in height. |
| Kalahari | Subtropical | 900,000 | +45 to −12 | 100–500 | Featureless. Gently undulating sand covered 900 m above sea level. |
| Patagonia | Rain shadow | 673,000 | +13 to −33 | 127–203 | Alternating tablelands and massifs dissected by river valleys and canyons. |
| Sonoran | Subtropical | 310,000 | +48 to +4 | 30–265 | Mountain ranges and valleys. Trees common on rocky slopes and native cacti. Saguaros abundant. |
| Thar | Subtropical | 200,000 | +50 to 0 | 100–500 | Undulating sandy dunes separated by sandy plains and low barren hills, which rise abruptly from surrounding plains. |
| Namib | Coastal | 160,000 | +35 to +1 | 5–85 | Broad expanse of hyper-arid gravel and dunes that stretch along the entire coastline. Fog is frequent. |
| Atacama | Coastal | 140,000 | +32 to −2 | 0–1 | Stony terrain, salt lakes, sand, and felsic lava. |
| Danakil | Subtropical | 137,000 | +50 to +25 | 25–200 | Volcanic lakes formed by lava blockages of valleys, and salt lakes flanked by mountains to the east. |
| Mojave | Subtropical | 124,000 | +36 to +2 | 89–250 | Mountains and basins with spare vegetation. Dry riverbeds, towering sand dunes, and lava flows. |
| Ferlo | Subtropical | 70,000 | +43 to +32 | 300–400 | Plains and sand dunes, scattered rocks, and small valleys with clay soil. Traversed by tributaries of Senegal River. |
| Autotrophic Microalgae | Heterotrophic Microalgae | |
|---|---|---|
| Advantages | Low initial costs, as CO2 and H2O are relatively inexpensive, natural resources. Can reduce the amount of atmospheric CO2. Easier to genetically modify than other microalgae. | Higher yield of lipids; therefore, the cost of extracting and harvesting oils is lower. Growth is not limited by sunlight. Yield is 5.5 times larger compared to autotrophic microalgae. Shorter scale-up time than autotrophic microalgae. Requires 12 times less land for cultivation. |
| Disadvantages | High sunlight for photosynthesis is needed for growth. Areas where microalgae are grown are restricted to latitudes 30° above and below the equator and coastal regions with a flat topography. Mutual shading of cells. Lower yields of lipids extracted. | High initial cost to provide organic carbon sources, such as glucose and acetate. |
| Methods of Cultivating Microalgae | ||
|---|---|---|
| Open Raceway Pond | Photobioreactors | |
| Design characteristics | Closed-loop oval-shaped channels. Channels filled with water to a depth of 0.25–0.4 m, constructed using concrete. Circulation of nutrients using stirrer paddles. | Array of transparent tubes. Tubes have a diameter of 0.1 m, made from plastic or glass. Circulation of nutrients using a pump or airlift technology. Use an external light source. |
| Advantages | Easy to construct. Low construction, operating, and maintenance costs. Lower energy demands. Can use non-arable land. | Increased exposure to light. Larger biomass and lipid productivity. Low water consumption. Low risk of contamination. |
| Disadvantages | Susceptible to contamination, predators, and harsh weather conditions. Requires a large amount of land. Water loss is an issue. Hard to control the temperature. | High construction, operating, and maintenance costs. |
Table 11.
Minimum jet fuel selling price (MJSP) from the HEFA pathway [30].
Table 11.
Minimum jet fuel selling price (MJSP) from the HEFA pathway [30].
| Feedstock | MJSP (US$/Gallon) |
|---|---|
| Camelina oil | 1.63–4.62 |
| Soybean oil | 3.82–4.39 |
| Jatropha oil | 5.42–5.74 |
| Waste oil | 2.36–4.73 |
| Microalgae | 31.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharma, V.; Hossain, A.K.; Duraisamy, G.; Griffiths, G. Microalgal Biodiesel: A Challenging Route toward a Sustainable Aviation Fuel. Fermentation 2023, 9, 907. https://doi.org/10.3390/fermentation9100907
AMA Style
Sharma V, Hossain AK, Duraisamy G, Griffiths G. Microalgal Biodiesel: A Challenging Route toward a Sustainable Aviation Fuel. Fermentation. 2023; 9(10):907. https://doi.org/10.3390/fermentation9100907
Chicago/Turabian StyleSharma, Vikas, Abul Kalam Hossain, Ganesh Duraisamy, and Gareth Griffiths. 2023. "Microalgal Biodiesel: A Challenging Route toward a Sustainable Aviation Fuel" Fermentation 9, no. 10: 907. https://doi.org/10.3390/fermentation9100907
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.