
Enhanced Phycocyanobilin Production in Escherichia coli by Fusion-Expression of Apo-Proteins with Signal Peptides

1
Key Laboratory of Marine Drugs, Chinese Ministry of Education, School of Medicine and Pharmacy, Ocean University of China, Qingdao 266003, China
2
Laboratory for Marine Drugs and Bioproducts, Pilot National Laboratory for Marine Science and Technology, Qingdao 266237, China
3
Laboratory of Marine Medicinal Resources Discovery, Marine Biomedical Research Institute of Qingdao, Qingdao 266075, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Fermentation 2023, 9(9), 851; https://doi.org/10.3390/fermentation9090851
Submission received: 4 August 2023
/
Revised: 10 September 2023
/
Accepted: 16 September 2023
/
Published: 18 September 2023
(This article belongs to the Special Issue Production of Nutritional and Functional Properties in Genetically Engineered Microorganisms)
Abstract
:Phycocyanobilin (PCB) is the bioactive chromophore attached to Phycocyanin (PC) that is of special interest for nutraceutical and therapeutic applications. However, the production of PCB from the heterologous host Escherichia coli is still very low. To facilitate subsequent application of PCB, improving its production in microbial hosts is still a challenge to be solved. In this paper, a strategy involving fusion-expression of apo-proteins with signal peptides was adopted to improve PCB production in E. coli. First, we reconstructed the PCB biosynthesis pathway in E. coli and then optimized its culture media. Subsequently, one PC α (CpcA) subunit and one PC β (CpcB) subunit, which can capture free PCB, were introduced and increased the yield of PCB. Finally, CpcA was fused with seven signal peptides to generate recombinant proteins, among which, the signal peptide N20 fused with CpcA protein drastically improved PCB production in E. coli, providing a maximum flask output of 8.47 ± 0.18 mg/L. The results of this study demonstrate that PCB distribution and transporting manners in E. coli could affect the heterologous production efficiency. By fusing apo-proteins with signal peptides, the secretion of phycocyanin was refined and the production of PCB was successfully enhanced by 3.7-fold, compared with the starting strain (1.80 ± 0.12 mg/L). This work provided an alternative method for improving the production of PCB and other phycobilins.
1. Introduction
Phycocyanobilin (PCB) is a linear tetrapyrrolic chromophore which was originally discovered from the blue–green algae Spirulina platensis. In the producing organisms, PCB is usually covalently bonded to apo-proteins through a specific cysteine residue with a thioether linkage and absorbs light in the red region of the electromagnetic spectrum, thus exhibiting a vivid blue color [1,2]. Recent studies have demonstrated that PCB exhibits significant bioactivities including anti-cancer [3], anti-inflammatory [4], atheroprotective [5], anti-oxidant and nephroprotective [6,7] effects, which make it of great interest to food and pharmaceutical industries. In addition, due to the special optical properties, it also shows great application potential in immunodiagnostics and photodynamic therapy [8].
Biosynthetic studies have revealed that the molecule of PCB is derived from the cyclic tetrapyrrole of heme via a cascade of redox reactions where Ho1, a ferredoxin-dependent heme oxygenase, promotes the oxidative cleavage of the tetracyclic structure of heme and produces the intermediate of biliverdin IXα (BV), which is then converted to PCB, catalyzed by PcyA, which is also known as phycocyanobilin: ferredoxin oxidoreductase [9,10,11]. In cyanobacteria, PCB always attaches to apo-allophycocyanin (apo-APC) and apo-phycocyanin (apo-PC), the key pigments assisting photosynthesis, to form the fluorescent proteins of APC and PC [12,13]. Both APC and PC are assembled as oligomers composed from α and β units. For instance, C-phycocyanin (CPC), one of the major phycobiliproteins, is composed of α and β subunits in the form of (αβ)3 or (αβ)6 aggregates [13,14]. The subunits are covalently bonded to the PCB chromophore through thioethers with catalysis by the chromophores lyases, among which CpcE and CpcF are responsible for the attachment of PCB to the PC α subunit [15,16], and CpcS or CpcT catalyzes the attachment of PCB to the chromophore binding site of Cys-β82 or Cys-β153, respectively, to the PC β unit [17,18].
In light of the great potential for application in the food industry and healthcare, good efforts have been devoted to developing heterologous expression systems that facilitate the production of PCB and associated phycobiliproteins. The key enzymes of the ho1 and pcyA genes were first co-expressed in Escherichia coli BL21 (DE3) by Ge et al., and PCB was successfully produced with around 3 mg/L [11]. Meanwhile, higher production of PCB has been achieved by Tooley and Wu via expressing apoproteins in combination, like apo-PC α or β subunits, apo-phycoerythrocyanin and streptavidin, together with PCB, in E. coli BL21 (DE3) as a heterologous host [14,19,20,21]. The metabolic flux was also optimized by up-regulating hemB, hemG, hemH, hemA and hemL and provided higher supply of heme, the cyclic tetrapyrrole precursor of PCB. Instead, the accumulation of heme, as well as the product of PCB, will cause not only negative feedback to the biosynthetic process, but metabolic burden and even toxicity to the heterologous host cells, which greatly constrict the productivity of PCB in E. coli [22].
Cells of Gram-negative bacteria like E. coli can be divided into four regions: cytoplasm, inner membrane, periplasm and outer membrane. Non-intracellular proteins such as periplasm proteins, outer membrane proteins and even heterologously expressed proteins must be carefully translocated and exported across the inner membrane to maintain the homeostasis of the intracellular structural space, and most importantly, to deliver the functional proteins to their specific destinations [23]. For the translocation process of the periplasmic and outer membrane proteins, the amino-terminal signal sequences play key roles in directing these precursor proteins across the inner membranes [24]. In light of this, different signal peptide systems have been well-developed to engineer the cell wall permeability and secretion process in E. coli, which resulted in the improved production of a series of recombinant proteins like penicillin G acylases [25], lipoxygenase [26], phytase [27] and human leptin [28].
In this study, we developed a strategy that increases PCB production via accelerating translocation and secretion of phycobiliproteins, achieved by constructing recombinant apoproteins fused with signal peptides. For a start, the two key genes ho1 and pcyA were expressed in E. coli BL21 (DE3), which resulted in the biosynthesis of PCB with a production of 1.80 mg/L. Then, the production of PCB was improved to 3.52 mg/L by co-expressing the α and or β subunits of PC (apo-CpcA and apo-CpcB) in one cell. Finally, the apo-CpcA and apo-CpcB proteins were fitted with seven signal peptides which have been widely used for the efficient secretory production of recombinant proteins in E. coli. As a result, the strain expressing apo-CpcA fused with the signal peptide N20 provided an improved PCB production of 8.47 mg/L, which is 3.7-fold higher than the starting yield (the strain with ho1 and pcyA).
2. Materials and Methods
2.1. Cyanobacterial Cultivation and DNA Extraction
Synechocystis sp. PCC6803 was obtained from East China University of Science and Technology. The alga was cultured in BG11 medium at 30 °C, at 130 rpm and with a light supply, until the OD730 reached 0.5~1.5. The alga was collected by centrifugation (5000 rpm) at the logarithmic phase. The Spirulina genomic DNA was extracted using TRIzol reagent (TIANGEN, Beijing, China) according to the manufacturer’s instructions.
2.2. Obtaining of the Gene Encoding Phycocyanin
The primers used to amplify the cpcA gene (GenBank: BAA17231.1) were 5′-GGA ATT CCA TAT GAA AAC CCC TTT AAC TGA AG-3′ and 5′-ACG CGT CGA CCG TAC GAC TAG TCT AGC TCA GAG-3′. The resulting 510 bp product was digested with restriction enzymes NdeI and SalI and cloned into the NdeI- and SalI-digested cloning vector, pET28a(+), giving the plasmid pET28a-cpcA. The other genes (cpcE, cpcF, cpcB, cpcS, cpcT, ho1, pcyA) were synthesized by BGI (Beijing, China) with codon optimization (Table S4), for improved expression, and cloned into pET28a(+).
2.3. Bacterial Strains and Culture Conditions
E. coli strain DH5α (TransGen) was used for recombinant DNA manipulation. E. coli BL21 (DE3) (TransGen) was used as a host for the expression. Cells were cultured in 5 mL Luria Broth (LB) medium for 12 h at 37 °C. The seed culture was inoculated into 100 mL Terrific Broth (TB) medium in a 250 mL flask. To induce the expression of the desired proteins, Isopropyl-β-D-thiogalactopyranoside (IPTG) was added to a final concentration of 0.1 mM when the OD600 reached 0.6. After induction, cells were cultured at 25 °C for 22 h. Antibiotics were added as follows: kanamycin (Kan) 30 mg/mL and ampicillin (Amp) 100 mg/mL.
2.4. Plasmid Construction
All plasmids (Table S2) were constructed by using a double enzyme digestion and ligation method. The pET28a-cpcF-cpcE-N20-cpcA plasmid construction was taken as an example. First, the signal peptide N20 was fused to the cpcA gene by overlapping PCR and digested with the restriction enzymes NdeI and XhoI, and ligated into pET28a(+), which was digested with the same restriction enzymes, gaining pET28a-N20-cpcA. Then, the pET28a-cpcE was digested with the restriction enzymes XbaI and XhoI, and ligated into pET28a-cpcF, which was digested with the restriction enzymes SpeI and XhoI, resulting in the plasmid pET28a-cpcF-cpcE. The pET28a-cpcF-cpcE-N20-cpcA was constructed in the same method. The primers are listed in Table S3.
2.5. PCB Standard Curve
PCB was purified by HPLC with a YMC-Pack ODS-A column (10 mm × 250 mm, 5 μm) for the standard curve preparation. Purified PCB powder was measured and dissolved in a specific volume of methanol to make a PCB solution. Next, this PCB solution was serially diluted. The peak area of each dilution was measured at 687 nm on HPLC analysis with a constant mobile phase of methanol–water at 45% over 20 min at 1 mL/min. Based on the correlation between the peak area and concentration, a standard curve of PCB was prepared to calculate the yields in the optimization experiments. All crude extracts were dissolved in methanol and their peak area at 687 nm were measured on the HPLC. The yields were calculated based on the measured data and standard curve. All samples were repeated for three replicates.
2.6. SDS–PAGE Analysis
Cells were harvested by centrifugation at 12,000 rpm for 1 min and then resuspended in ddH2O. The protein samples were heated for 10 min in loading buffer at 100 °C, and centrifugated at 12,000 rpm for 2 min, before loading onto the gel. Proteins were separated using 12% SDS–PAGE and protein bands were detected by a gel-imaging system after staining with Coomassie brilliant blue R-250.
2.7. Alcoholysis
Cell pellets were collected and then mixed with 20 mL ethanol 95% v/v in a 50 mL round bottom flask paired with a reflux condenser. The flask was equipped with a magnetic stirrer and submerged in a silicon oil bath heated to a temperature 10 °C above the boiling point of the reagent. Running tap water was used to cool the condenser. The reflux reaction was stopped after 4 h by submerging the flask in an ice bath for 15 min. The reaction mixture was filtered and the deep blue solution was collected and analyzed by HPLC, while the green solid protein residue was discarded.
2.8. Abbreviations
PCB: phycocyanobilin; PC: phycocyanin; Ho1: ferredoxin-dependent heme oxygenase; PcyA: phycocyanobilin: ferredoxin oxidoreductase; apo-APC: apo-allophycocyanin; apo-PC: apo-phycocyanin; APC: allophycocyanin; CPC: C-phycocyanin; CpcE: chromophores lyases; CpcF: chromophores lyases; CpcS: chromophores lyases; CpcT: chromophores lyases; pHP: pET21c-ho1-pcyA; pFEA: pET28a-cpcF-cpcE-cpcA; pBS: pET28a-cpcB-cpcS; pBT: pET28a-cpcB-cpcT; pFENA: pET28a-cpcF-cpcE-N20-cpcA; pPBT: pET28a-PhoA-cpcB-cpcT; pMBT: pET28a-MalE-cpcB-cpcT; OmpA: outer membrane protein A; pelB: pectate lyase B; Lpp: murein lipoprotein; PhoA: alkaline phosphatase; MalE: maltose-binding protein; IPTG: Isopropyl β-D-Thiogalactoside.
3. Results
3.1. Heterologous Production of PCB in E. coli through the Heme Catabolic Pathway
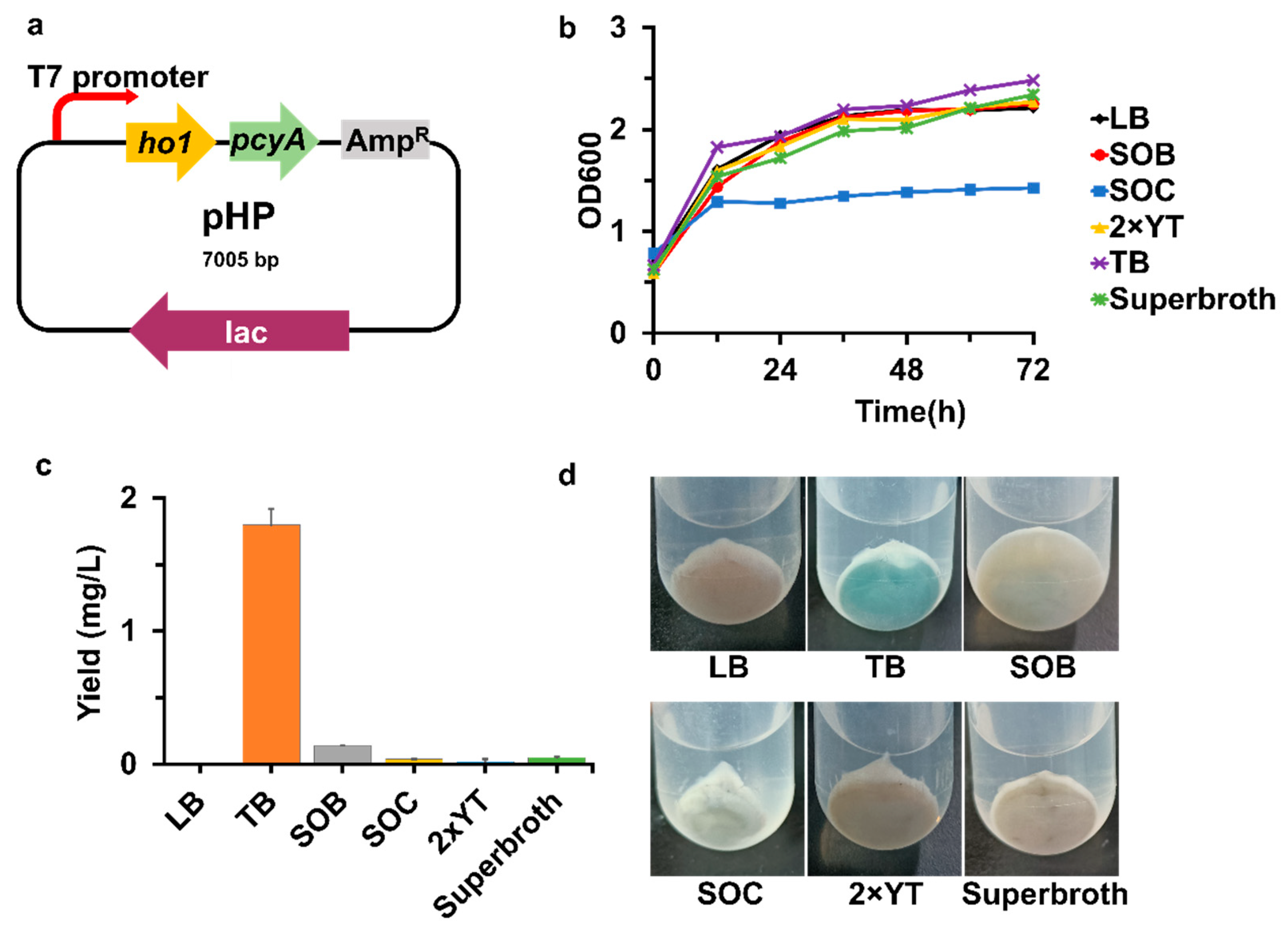
Two enzymes, ferredoxin-dependent heme oxygenase (Ho1) and phycocyanobilin: ferredoxin oxidoreductase (PcyA), are committed to phytochrome chromophore biosynthesis from heme [9,10,11]. We established a basic PCB biosynthesis pathway in E. coli by introducing the two genes ho1 and pcyA (Figure 1). First, the genes ho1 and pcyA were amplified and digested with the restriction enzymes NdeI and XhoI, and ligated into pET21c(+), which was digested with the same restriction enzymes, gaining pET21c-ho1 and pET21c-pcyA, respectively. Then, pET21c-pcyA was digested with the restriction enzymes XbaI and XhoI, and ligated into pET28a-ho1, which was digested with the restriction enzymes SpeI and XhoI, resulting in the plasmid pHP (Figure 2a). The gaining strain BL21/pHP was cultivated with LB medium and induced with IPTG after the plasmid pPH was transformed into E. coli BL21 (DE3). PCB was extracted from cell pellets and analyzed by HPLC at 687 nm. The results showed that the extracted sample had the same retention time and absorption as the authentic standard, suggesting the successful production of PCB (Figure S1). However, the PCB yield was too low to calculate by using a standard curve.
To improve the production of PCB in E. coli BL21 (DE3), an array of media including LB, SOB, SOC, 2 × YT, TB and superbroth media (recipes for media are in Table S1) were then used to produce PCB. As a result, the TB medium provided the optimal PCB production with a yield of 1.80 ± 0.12 mg/L (Figure 2c). Additionally, only TB medium contains glycerol, which has been shown to increase the production of carotenoid [29] and PCB [11]. TB medium also had the highest cell density, which may explain why PCB is produced more effectively in TB medium (Figure 2b).
3.2. Enhancing the PCB Yield by Introducing Apo-α or -β Protein
E. coli strains have been reported for a relatively low level of PCB production by expressing only ho1 and pcyA genes, which may be due to the feedback inhibition of PCB to the key enzymes in heme biosynthesis [22]. In view of this, co-expression of apoproteins by a combinatorial biosynthetic method is usually adopted to combine free PCB and reduce the negative feedback, which demonstrates an efficient way to improve the production of PCB. As aforementioned, PCB is usually attached with apo-proteins catalyzed by a set of chromophore lyases including CpcE, CpcF, CpcS and CpcT. By recombining the above genes with cpcA, which codes for the PC α subunit, and cpcB, which codes for the PC β subunit, we constructed the three plasmids pET28a-cpcF-cpcE-cpcA (pFEA), pET28a-cpcB-cpcS (pBS) and pET28a-cpcB-cpcT (pBT), among which pFEA is capable of producing PCB-CpcA-type phycocyanin, and pBS together with pBT is able to produce PCB-CpcB-type phycocyanins (Figure 3a and Figure 4a).
The plasmid pFEA, pBS or pBT was co-transformed with pHP into E. coli BL21 (DE3) to give the strains BL21/pFEA/pHP, BL21/pBS/pHP and BL21/pBT/pHP, respectively. We then examined the PCB productivity by culturing the above strains in TB medium. All of the three strains exhibited a distinct blue–green color (Figure 3d), which indicated the formation of phycocyanins. Among those transformants, the BL21/pFEA/pHP strain showed a significant bright blue color, suggesting it might have better potential for PCB production. After alcoholysis with ethanol, PCB was collected from the phycocyanin products of PCB-CpcA, PCB-CpcB (C-82) and PCB-CpcB (C-153) and then subjected to HPLC analysis. The results showed that the BL21/pFEA/pHP strain maintains the highest production of PCB with a yield of 3.52 ± 0.16 mg/L (Figure 3c), which is 0.9-fold higher than the PCB production in the E. coli BL21 (DE3) strain harboring only the pHP plasmid. However, for the BL21/pBS/pHP and BL21/pBT/pHP strains, the production of PCB is even lower than that in BL21/pHP strain, which may be mainly due to the fact that the production of CpcB is tightly regulated in E. coli [30] and the free PCB could not be saturated by the apo-protein, so that the negative feedback of PCB is still maintained at a relatively high level.
3.3. Enhancing PCB Production by Fusion-Expression of Apo-α Protein with Signal Peptide N20
To further improve the production of PCB, seven different signal peptides (Table 1) which were widely used for efficient secretory production of recombinant proteins in E. coli were selected for conducting and mediating phycocyanin secretion. We then constructed 13 recombinant expression plasmids carrying the corresponding signal peptides to be fused with apo-phycocyanins (7 for apo-CpcA and 6 for apo-CpcB). After 16 h of fermentation, the production of recombinant signal peptide-apo-CpcA or signal peptide-apo-CpcB was tested by SDS-PAGE. As shown in Table 1 and Figure S3 for apo-CpcA, 5 signal peptides, N20, pelB, PhoA, MalE and Kp-SP, could successfully produce the recombinant protein of signal peptide-apo-CpcA. However, compared with native apo-CpcA, only the production of N20-apo-CpcA increased significantly; the other 4 recombinant proteins showed little ability in assisting apo-CpcA production. As shown in Table 1 and Figure S4, for apo-CpcB, only 2 signal peptides, PhoA and MalE, could successfully produce PhoA-apo-CpcB and MalE-apo-CpcB, whose protein productions were comparable to native apo-CpcB. On the contrary, the other signal peptides even inhibited the expression of apo-CpcA or apo-CpcB when fused with them. The reason may be that the signal peptides not only guided transmembrane transport but also affected protein translation [31]. Therefore, the fusion proteins N20-apo-CpcA, PhoA-apo-CpcB and MalE-apo-CpcB were selected for further study.
To test whether these fused proteins could bind to the chromophore PCB and affect the final production of PCB, we constructed an N20-apo-CpcA-producing plasmid by inserting N20-cpcA, cpcE and cpcF into the pET28a(+) backbone (pFENA, Figure 4a). The plasmid pFENA was then co-transformed with pHP into E. coli BL21 (DE3) to form the strain BL21/pFENA/pHP. After induction with IPTG, the transformants harboring pFENA and pHP exhibited a bright blue color (Figure 4d), which implied a higher production of PCB-N20-CpcA. For comparison, two genes phoA-cpcB and malE-cpcB, which are responsible for producing the recombinant apo-CpcB proteins of PhoA-apo-CpcB and MalE-apo-CpcB, were also integrated with the C-153 lyase of cpcT (binding to C-153 site of CpcB) on the pET28a(+) backbone to form the plasmids pPBT and pMBT, which were co-transformed with pHP into E. coli BL21 (DE3). And, conceivably, both the strains BL21/pPBT/pHP and BL21/pMBT/pHP only presented a slight visible blue color, which indicated the low level expression of CpcB recombinant-related phycocyanins. The above results showed that the signal peptide of N20 could be accommodated well together with CpcA and once co-expressed in E. coli, the signal peptides systems could provide improved production of the phycocyanin.
To verify whether the production of PCB was increased as well by the N20 signal peptide system, the strain BL21/pFENA/pHP was regrown in TB media at 25 °C, 220 rpm for 22 h. After fermentation, the pellets were collected by centrifugation, then PCB-N20-CpcA was cleaved from the cell pellets by alcoholysis and analyzed by HPLC. The results showed that the yield of PCB was 6.57 ± 0.28 mg/L, which was 2.6-fold higher than that of free PCB (Figure 4c).
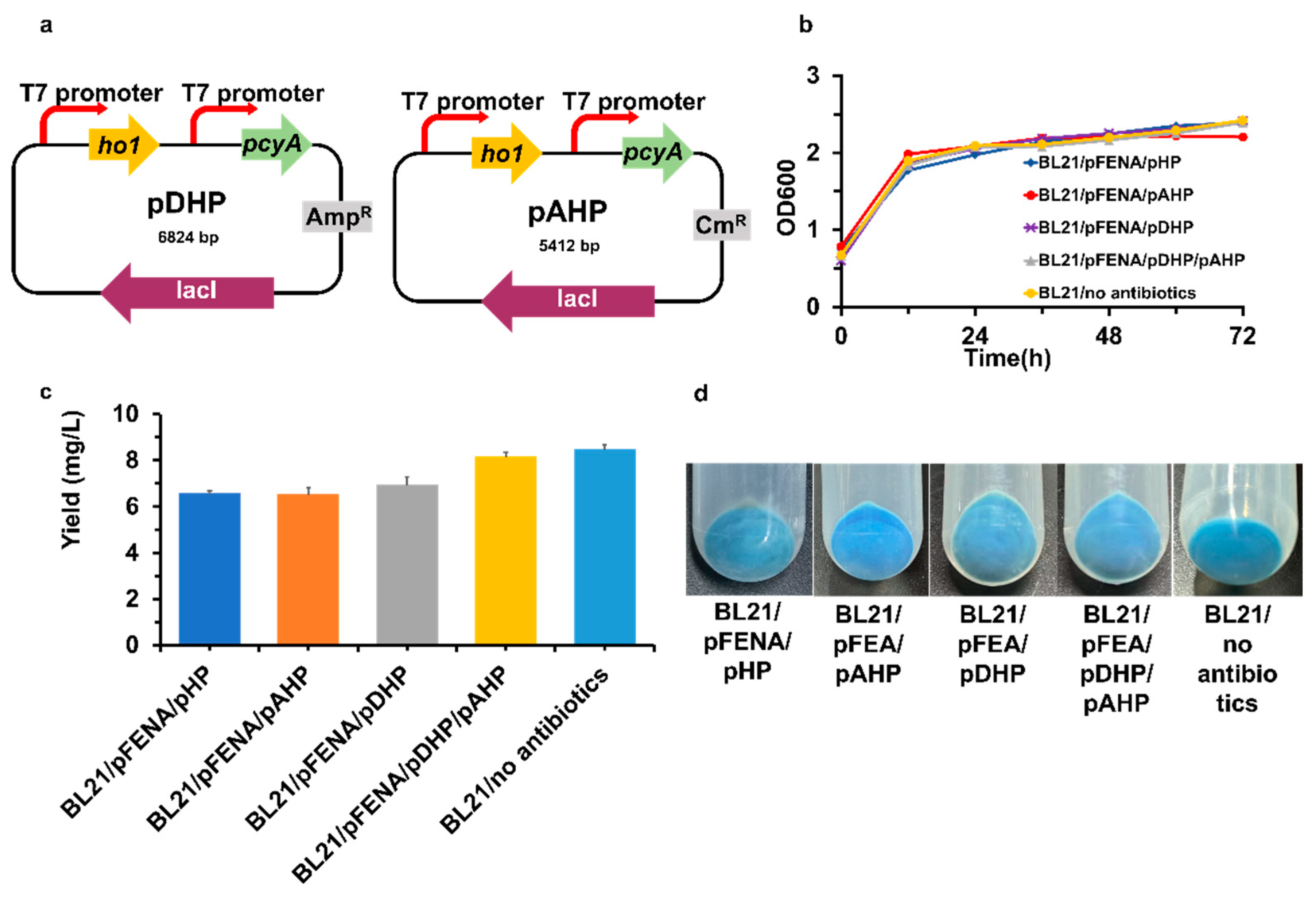
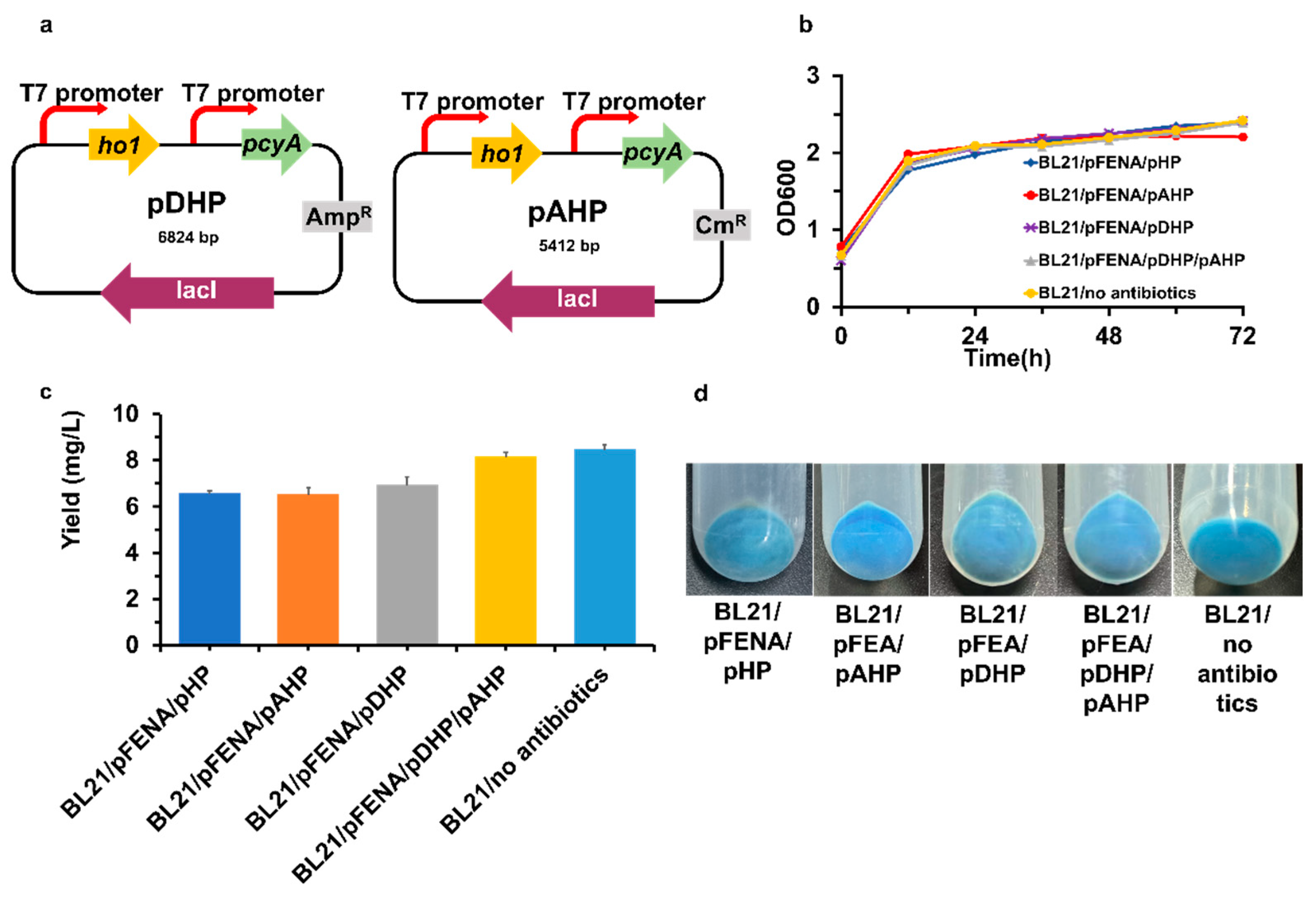
Since the pBR322-derived replicon is present in both the pET28a(+) and pET21c(+) vector backbones, two antibiotics—kanamycin and ampicillin—should be administered to the culture to ensure that these two plasmids are stably maintained in a single cell [34,35]. However, the use of multiple antibiotics could cause burden to the metabolic process of the chassis cell, and having multiple antibiotics applied is not an economic approach for sustainable industrial production. Therefore, we tried to modify our existing plasmid system by replacing pET21c(+) with pETDuet-1 (ColE1 origin) and/or pACYCDuet-1 (p15A origin), which carry different replicons from pET28a(+), and to be able to maintain multiple plasmids stably in antibiotic-free fermentation. We then constructed two PCB-producing plasmids by inserting ho1 and pcyA into pETDuet-1 (pDHP, Figure 5a) and pACYCDuet-1 (pAHP, Figure 5a), respectively. The plasmid pDHP and/or pAHP was then co-transformed with pFENA into E. coli BL21 (DE3) to form the strains BL21/pFENA/pDHP, BL21/pFENA/pAHP and BL21/pFENA/pDHP/pAHP. To test their ability to produce PCB-N20-CpcA, these three strains were cultured with the corresponding antibiotics. After induction with IPTG, all three transformants exhibited a bright blue color (Figure 5d), implying the PCB-N20-CpcA-producing ability of the modified expression systems. Then, we depleted the antibiotics in a new culture of the strain BL21/pFENA/pDHP/pAHP in TB medium (the culture named BL21/no antibiotics). As a result, the cell pellets exhibited a bright blue color like the cell pellets from strain BL21/pFENA/pDHP/pAHP, demonstrating that the PCB-N20-CpcA can be produced more efficiently without additional antibiotics. After fermentation and alcoholysis, the final expression system showed that the yield of PCB was 8.47 ± 0.18 mg/L, which was improved by 3.7-fold compared with the starting cells, which expressed only the ho1 and pcyA genes (Figure 5c).
4. Discussion
Due to its wide applications in food and medicine, PCB has drawn increasing attention for its large-scale preparation and applications. Despite the fact that it was possible to produce PCB in E. coli by co-expression of the key genes ho1 and pcyA, which are required to produce PCB from cyclic tetrapyrrole, the yield of PCB is still limited [11]. This demonstrated that the conversion of precursors is not the limiting step in PCB production and that the failure to transport PCB from within the cell and the subsequent inhibition of productivity by the accumulated product should be the main factor. In this case, improving and accelerating the PCB secretion process could be a solution to weaken the feedback inhibition of free PCB and benefit the PCB-producing system.
Despite many successful examples [36,37,38], the secretory production of heterologous proteins in E. coli still needs to deal with some obstacles: (1) the improper processing of signal sequences; (2) low secretion efficiency due to the characteristics of proteins; (3) undesirable interaction with the cellular components and compositions; and (4) formation of inclusion bodies due to overproduction or protein misfolding. In this study, we developed a PCB secretion system by overexpressing apoproteins fused with a signal peptide, which could bind PCB and facilitate the transportation of the phycocyanin products into the periplasmic space.
First, we built the basic PCB production strain BL21/pHP, which contains two genes, ho1 and pcyA. Our results clearly showed that the strain BL21/pHP could express PCB successfully and the culture media have a significant influence on the expression level of PCB (Figure 2). Then, we tested the binding ability of two types of apo-proteins, the apo-PC α subunit (CpcA) and the apo-PC β subunit (CpcB), with PCB. Compared with the β subunit, the α subunit exhibited a much higher binding efficiency to PCB; thus, the strain BL21/pFEA/pHP exhibited a much deeper bright blue color and higher production (3.52 ± 0.16 mg/L) than the BL21/pBS/pHP and BL21/pBT/pHP strains (Figure 3). This indicated that the α subunit could alleviate the feedback inhibition of free PCB and improve the final PCB production.
We then amplified seven reported signal peptides (e.g., pelB, Kp-SP, PhoA, OmpA, MalE, Lpp and N20) and constructed recombinant proteins by fusing them with CpcA. However, only the recombinant protein with N20 increased PCB production (6.57 ± 0.28 mg/L, Figure 4), while the other recombinant proteins even decreased PCB production compared with the control that expressed wild type CpcA. To overcome the above obstacles, additional fusion proteins derived from the recombination of a broader range of apo-proteins (e.g., ApcA, ApcB, CpeA and CpeB) and signal peptides (e.g., OmpC, OmpF, and LamB) should be constructed in further study and tested for better secretion capability.
5. Conclusions
This study reports the successful optimization of PCB biosynthesis in an E. coli microbial chassis. Intracellular accumulation and negative feedback of PCB were identified as critical bottlenecks for PCB production. This was alleviated through introducing a signal peptide-directed PCB secretion system. First, the ho1 and pcyA genes were heterologously expressed in E. coli BL21 (DE3), which provided the PCB production of 1.80 mg/L. Then the α subunit of C-phycocyanin CpcA was co-expressed in E. coli BL21 (DE3) in combination with ho1 and pcyA, which improved the PCB production by 0.9-fold (3.52 mg/L). Finally, by fusing CpcA with a signal peptide N20, which guided the transportation of the phycobiliprotein PCB-CpcA, we achieved an optimized PCB production of 8.47 mg/L, which is 4.7-fold higher than the starting yield. This work confirmed that the distribution and transporting manners of PCB in E. coli could affect its production and provided a new method for the production of PCB and other phycobilins such as phycoviolobilin and phycoerythrobilin.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation9090851/s1, Figure S1: HPLC analysis of BL/pHP strain; Figure S2: Standard curve for PCB using HPLC at 687 nm; Figure S3: SDS-PAGE of expression of pET28a-sp-cpcA; Figure S4: SDS-PAGE of expression of pET28a-sp-cpcB; Table S1: Recipes of the media used in this study; Table S2: Plasmids and strains used in this work; Table S3: Primers used in this study; Table S4: Codon optimized gene sequences used in this study.
Author Contributions
X.L. and J.Y. performed the experiments, drafted the manuscript, analyzed the data and prepared the figures. Q.C., T.Z. and D.L. contributed to the critical reading of the manuscript; G.Z. designed the study, supervised the laboratory work, and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (41976105, 81991522), Qingdao Marine Science and Technology Center (2022QNLM030003-2), NSFC-Shandong Joint Fund (U1906212), Hainan Provincial Joint Project of Sanya Yazhou Bay Science and Technology City (2021CXLH0012), Taishan Scholar Youth Expert Program in Shandong Province (tsqn 202103153), Major Basic Research Programs of Natural Science Foundation of Shandong Province (ZR2021ZD28).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this article and its Supplementary Materials. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Jianhua Fan from East China University of Science and Technology, for providing Synechocystis sp. PCC6803. We thank Xiaoying Bian from Shandong University for providing plasmids pETDuet-1 and pACYCDuet-1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Biswas, A.; Vasquez, Y.M.; Dragomani, T.M.; Kronfel, M.L.; Williams, S.R.; Alvey, R.M.; Bryant, D.A.; Schluchter, W.M. Biosynthesis of cyanobacterial phycobiliproteins in Escherichia coli: Chromophorylation efficiency and specificity of all bilin lyases from Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2010, 76, 2729–2739. [Google Scholar] [CrossRef] [PubMed]
- Malwade, C.R.; Roda-Serrat, M.C.; Christensen, K.V.; Fretté, X.; Christensen, L.P. Kinetics of Phycocyanobilin Cleavage from C-Phycocyanin by Methanolysis. Comput. Aided Chem. Eng. 2016, 38, 61–66. [Google Scholar] [CrossRef]
- Koníčková, R.; Vaňková, K.; Vaníková, J.; Váňová, K.; Muchová, L.; Subhanová, I.; Zadinová, M.; Zelenka, J.; Dvořák, A.; Kolář, M. Anti-cancer effects of blue-green alga Spirulina platensis, a natural source of bilirubin-like tetrapyrrolic compounds. Ann. Hepatol. 2014, 13, 273–283. [Google Scholar] [CrossRef]
- Marín-Prida, J.; Pavón-Fuentes, N.; Llópiz-Arzuaga, A.; Fernández-Massó, J.R.; Delgado-Roche, L.; Mendoza-Marí, Y.; Santana, S.P.; Cruz-Ramírez, A.; Valenzuela-Silva, C.; Nazábal-Gálvez, M. Phycocyanobilin promotes PC12 cell survival and modulates immune and inflammatory genes and oxidative stress markers in acute cerebral hypoperfusion in rats. Toxicol. Appl. Pharmacol. 2013, 272, 49–60. [Google Scholar] [CrossRef]
- Strasky, Z.; Zemankova, L.; Nemeckova, I.; Rathouska, J.; Nachtigal, P. Spirulina platensis and phycocyanobilin activate atheroprotective heme oxygenase-1: A possible implication for atherogenesis. Food Funct. 2013, 4, 1586–1594. [Google Scholar] [CrossRef]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from Spirulina platensis protect against diabetic nephropathy by inhibiting oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R110–R120. [Google Scholar] [CrossRef]
- Garcia-Pliego, E.; Franco-Colin, M.; Rojas-Franco, P.; Blas-Valdivia, V.; Serrano-Contreras, J.I.; Pentón-Rol, G.; Cano-Europa, E. Phycocyanobilin is the molecule responsible for the nephroprotective action of phycocyanin in acute kidney injury caused by mercury. Food Funct. 2021, 12, 2985–2994. [Google Scholar] [CrossRef]
- Encarnação, T.; Pais, A.A.C.C.; Campos, M.G.; Burrows, H.D. Cyanobacteria and Microalgae: A Renewable Source of Bioactive Compounds and Other Chemicals. Sci. Prog. 2015, 98, 145–168. [Google Scholar] [CrossRef]
- Beale, S.I. Biosynthesis of phycobilins. Chem. Rev. 1993, 93, 785–802. [Google Scholar] [CrossRef]
- Dammeyer, T.; Michaelsen, K.; Frankenberg-Dinkel, N. Biosynthesis of open-chain tetrapyrroles in Prochlorococcus marinus. FEMS Microbiol. Lett. 2007, 271, 251–257. [Google Scholar] [CrossRef]
- Ge, B.; Li, Y.; Sun, H.; Zhang, S.; Hu, P.; Qin, S.; Huang, F. Combinational biosynthesis of phycocyanobilin using genetically-engineered Escherichia coli. Biotechnol. Lett. 2013, 35, 689–693. [Google Scholar] [CrossRef]
- Craig, I.W.; Carr, N.G. C-phycocyanin and allophycocyanin in two species of blue-green algae. Biochem. J. 1968, 106, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Wu, X.J.; Yang, H.; Chen, Y.T.; Li, P.P. Biosynthesis of Fluorescent beta Subunits of C-Phycocyanin from Spirulina subsalsa in Escherichia coli, and Their Antioxidant Properties. Molecules 2018, 23, 1369. [Google Scholar] [CrossRef]
- Guan, X.; Qin, S.; Su, Z.; Zhao, F.; Ge, B.; Li, F.; Tang, X. Combinational biosynthesis of a fluorescent cyanobacterial holo-alpha-phycocyanin in Escherichia coli by using one expression vector. Appl. Biochem. Biotechnol. 2007, 142, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Li, P.; Chen, X.; Chao, X. Combinatorial biosynthesis of Synechocystis PCC6803 phycocyanin holo-alpha-subunit (CpcA) in Escherichia coli and its activities. Appl. Microbiol. Biotechnol. 2016, 100, 5375–5388. [Google Scholar] [CrossRef]
- Zhao, K.H.; Su, P.; Tu, J.M.; Wang, X.; Liu, H.; Plöscher, M.; Eichacker, L.; Yang, B.; Zhou, M.; Scheer, H. Phycobilin: Cystein-84 biliprotein lyase, a near-universal lyase for cysteine-84-binding sites in cyanobacterial phycobiliproteins. Proc. Natl. Acad. Sci. USA 2007, 104, 14300–14305. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Xu, D.; Zang, X.; Yuan, D.; Zhao, B.; Tang, L.; Tan, Y.; Zhang, X. Lyase activities of heterologous CpcS and CpcT for phycocyanin holo-β-subunit from Arthrospira platensis in Escherichia coli. J. Ocean. Univ. China 2014, 13, 497–502. [Google Scholar] [CrossRef]
- Tooley, A.J.; Glazer, A.N. Biosynthesis of the Cyanobacterial Light-Harvesting Polypeptide Phycoerythrocyanin Holo-α Subunit in a Heterologous Host. J. Bacteriol. 2002, 184, 4666–4671. [Google Scholar] [CrossRef]
- Tooley, A.J.; Cai, Y.A.; Glazer, A.N. Biosynthesis of a fluorescent cyanobacterial C-phycocyanin holo-alpha subunit in a heterologous host. Proc. Natl. Acad. Sci. USA 2001, 98, 10560–10565. [Google Scholar] [CrossRef] [PubMed]
- Ge, B.; Lin, X.; Chen, Y.; Wang, X.; Chen, H.; Jiang, P.; Huang, F. Combinational biosynthesis of dual-functional streptavidin-phycobiliproteins for high-throughput-compatible immunoassay. Process Biochem. 2017, 58, 306–312. [Google Scholar] [CrossRef]
- Ge, B.; Chen, Y.; Yu, Q.; Lin, X.; Li, J.; Qin, S. Regulation of the heme biosynthetic pathway for combinational biosynthesis of phycocyanobilin in Escherichia coli. Process Biochem. 2018, 71, 23–30. [Google Scholar] [CrossRef]
- Danese, P.N.; Silhavy, T.J. Targeting and assembly of periplasmic and outer-membrane proteins in Escherichia coli. Annu. Rev. Genet. 1998, 32, 59–94. [Google Scholar] [CrossRef] [PubMed]
- von Heijne, G. The signal peptide. J. Membr. Biol. 1990, 115, 195–201. [Google Scholar] [CrossRef]
- Pan, X.; Yu, Q.; Chu, J.; Jiang, T.; He, B. Fitting replacement of signal peptide for highly efficient expression of three penicillin G acylases in E. coli. Appl. Microbiol. Biotechnol. 2018, 102, 7455–7464. [Google Scholar] [CrossRef]
- Pang, C.; Liu, S.; Zhang, G.; Zhou, J.; Du, G.; Li, J. Enhancing extracellular production of lipoxygenase in Escherichia coli by signal peptides and autolysis system. Microb. Cell Fact. 2022, 21, 42–53. [Google Scholar] [CrossRef]
- Miksch, G.; Kleist, S.; Friehs, K.; Flaschel, E. Overexpression of the phytase from Escherichia coli and its extracellular production in bioreactors. Appl. Microbiol. Biotechnol. 2002, 59, 685–694. [Google Scholar] [CrossRef]
- Jeong, K.J.; Lee, S.Y. Secretory production of human leptin in Escherichia coli. Biotechnol. Bioeng. 2000, 67, 398–407. [Google Scholar] [CrossRef]
- Fidan, O.; Zhan, J. Discovery and engineering of an endophytic Pseudomonas strain from Taxus chinensis for efficient production of zeaxanthin diglucoside. J. Biol. Eng. 2019, 13, 66–83. [Google Scholar] [CrossRef]
- Cao, X.; Zang, X.; Liu, Z.; Jin, Y.; Sun, D.; Guo, Y.; Wang, Z.; Zhang, F.; Lin, J. Molecular cloning of the cpeT gene encoding a bilin lyase responsible for attachment of phycoerythrobilin to Cys-158 on the β-subunit of phycoerythrin in Gracilariopsis lemaneiformis. J. Appl. Phycol. 2019, 31, 3331–3340. [Google Scholar] [CrossRef]
- Voss, M.; Schröder, B.; Fluhrer, R. Mechanism, specificity, and physiology of signal peptide peptidase (SPP) and SPP-like proteases. Biochim. Biophys. Acta. 2013, 1828, 2828–2839. [Google Scholar] [CrossRef]
- Cui, Y.; Meng, Y.; Zhang, J.; Cheng, B.; Yin, H.; Gao, C.; Xu, P.; Yang, C. Efficient secretory expression of recombinant proteins in Escherichia coli with a novel actinomycete signal peptide. Protein Expr. Purif. 2017, 129, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Luan, Y.; Liang, Q.; Qi, Q. Exploring the N-terminal role of a heterologous protein in secreting out of Escherichia coli. Biotechnol. Bioeng. 2016, 113, 2561–2567. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, B.A.; Admiraal, S.J.; Gramajo, H.; Cane, D.E.; Khosla, C. Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science 2001, 291, 1790–1792. [Google Scholar] [CrossRef]
- Liu, X.; Hua, K.; Liu, D.; Wu, Z.; Wang, Y.; Zhang, H.; Deng, Z.; Pfeifer, B.A.; Jiang, M. Heterologous Biosynthesis of Type II Polyketide Products Using E. coli. ACS Chem. Biol. 2020, 15, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Chen, R. Extracellular recombinant protein production from Escherichia coli. Biotechnol. Lett. 2009, 31, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, H.; Zhou, C.; Ma, Y.; Li, J.; Song, J. Cloning, expression and characterization of a pectate lyase from Paenibacillus sp. 0602 in recombinant Escherichia coli. BMC Biotechnol. 2014, 14, 18–29. [Google Scholar] [CrossRef]
- Teng, C.; Jia, H.; Yan, Q.; Zhou, P.; Jiang, Z. High-level expression of extracellular secretion of a β-xylosidase gene from Paecilomyces thermophila in Escherichia coli. Bioresour. Technol. 2011, 102, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Three-step improvement of PCB production. (a) The basic PCB biosynthesis pathway. The black arrows indicate the endogenous pathway for heme production, the red arrows indicate heterologous expression genes for PCB production and the PCB is exhibited by blue color (blue objects). Ho1, ferredoxin-dependent heme oxygenase; PcyA, phycocyanobilin: ferredoxin oxidoreductase; (b) Modified biosynthesis with apo-α or β proteins. P1 plasmid carrying heterologous expression genes for PC apo-α or -β subunit production and then covalently bonded to the PCB with catalysis by the chromophores lyases CpcE/CpcF, CpcS and CpcT; (c) Enhanced PCB production by fusion-expression of apo-proteins with signal peptides. P2 plasmid carrying heterologous expression genes for PC apo-α or -β subunits fused with signal peptide production and then covalently bonded to the PCB. The signal peptide could guide PC α or β subunit secretion. The red box represents the signal peptides library, including N20, OmpA, pelB, LPP, PhoA, MalE and Kp-SP. PC, phycocyanin; PCB, phycocyanobilin.
Figure 1.
Three-step improvement of PCB production. (a) The basic PCB biosynthesis pathway. The black arrows indicate the endogenous pathway for heme production, the red arrows indicate heterologous expression genes for PCB production and the PCB is exhibited by blue color (blue objects). Ho1, ferredoxin-dependent heme oxygenase; PcyA, phycocyanobilin: ferredoxin oxidoreductase; (b) Modified biosynthesis with apo-α or β proteins. P1 plasmid carrying heterologous expression genes for PC apo-α or -β subunit production and then covalently bonded to the PCB with catalysis by the chromophores lyases CpcE/CpcF, CpcS and CpcT; (c) Enhanced PCB production by fusion-expression of apo-proteins with signal peptides. P2 plasmid carrying heterologous expression genes for PC apo-α or -β subunits fused with signal peptide production and then covalently bonded to the PCB. The signal peptide could guide PC α or β subunit secretion. The red box represents the signal peptides library, including N20, OmpA, pelB, LPP, PhoA, MalE and Kp-SP. PC, phycocyanin; PCB, phycocyanobilin.
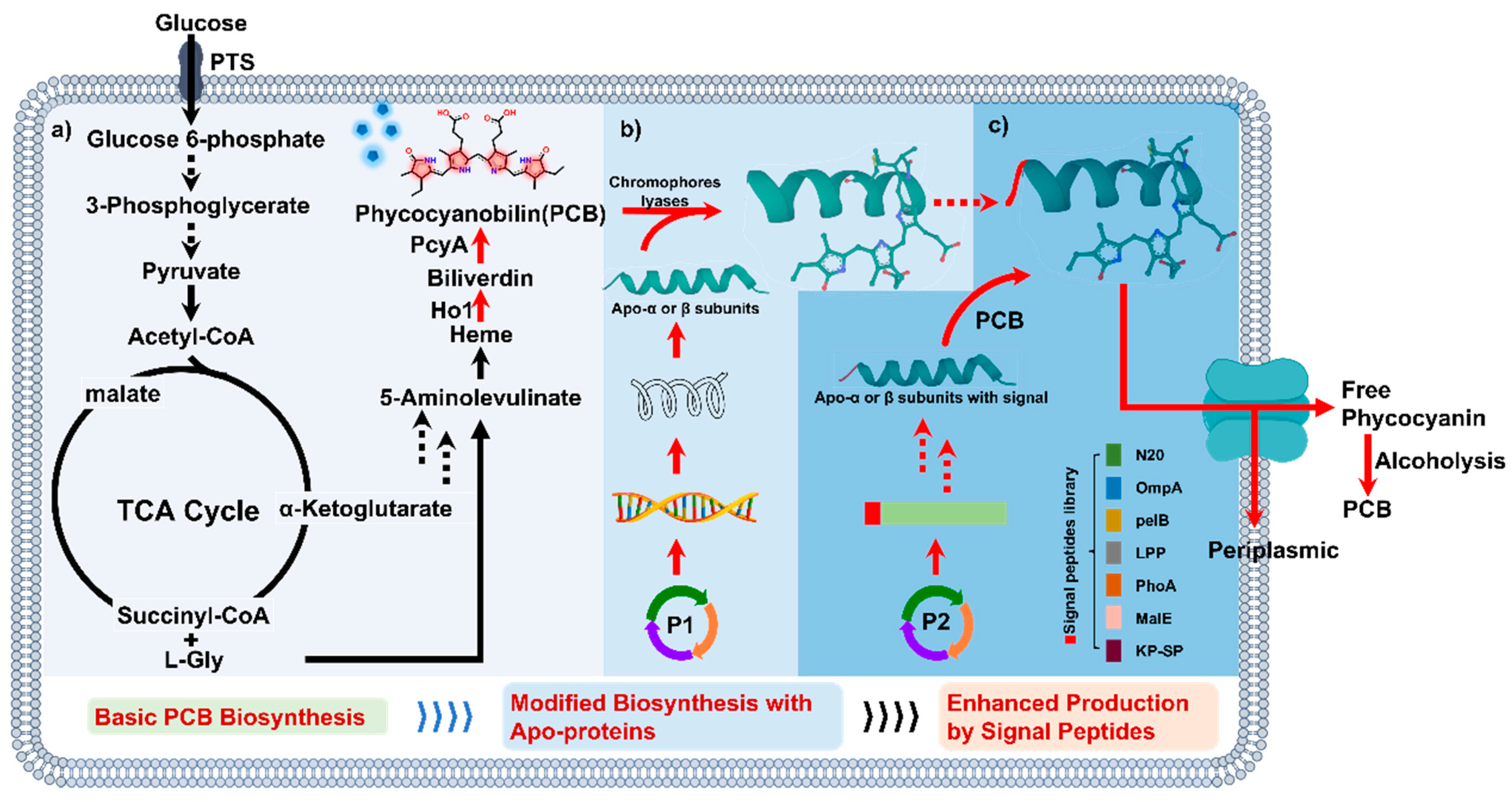
Figure 2.
The effects of different culture media on the yield of PCB. (a) Schematic representation of pHP; (b) The OD600 of strain BL/pHP under different culture media; (c) The PCB production of strain BL/pHP under different culture media; (d) The cell pellets of strain BL/pHP under different culture media. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
Figure 2.
The effects of different culture media on the yield of PCB. (a) Schematic representation of pHP; (b) The OD600 of strain BL/pHP under different culture media; (c) The PCB production of strain BL/pHP under different culture media; (d) The cell pellets of strain BL/pHP under different culture media. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
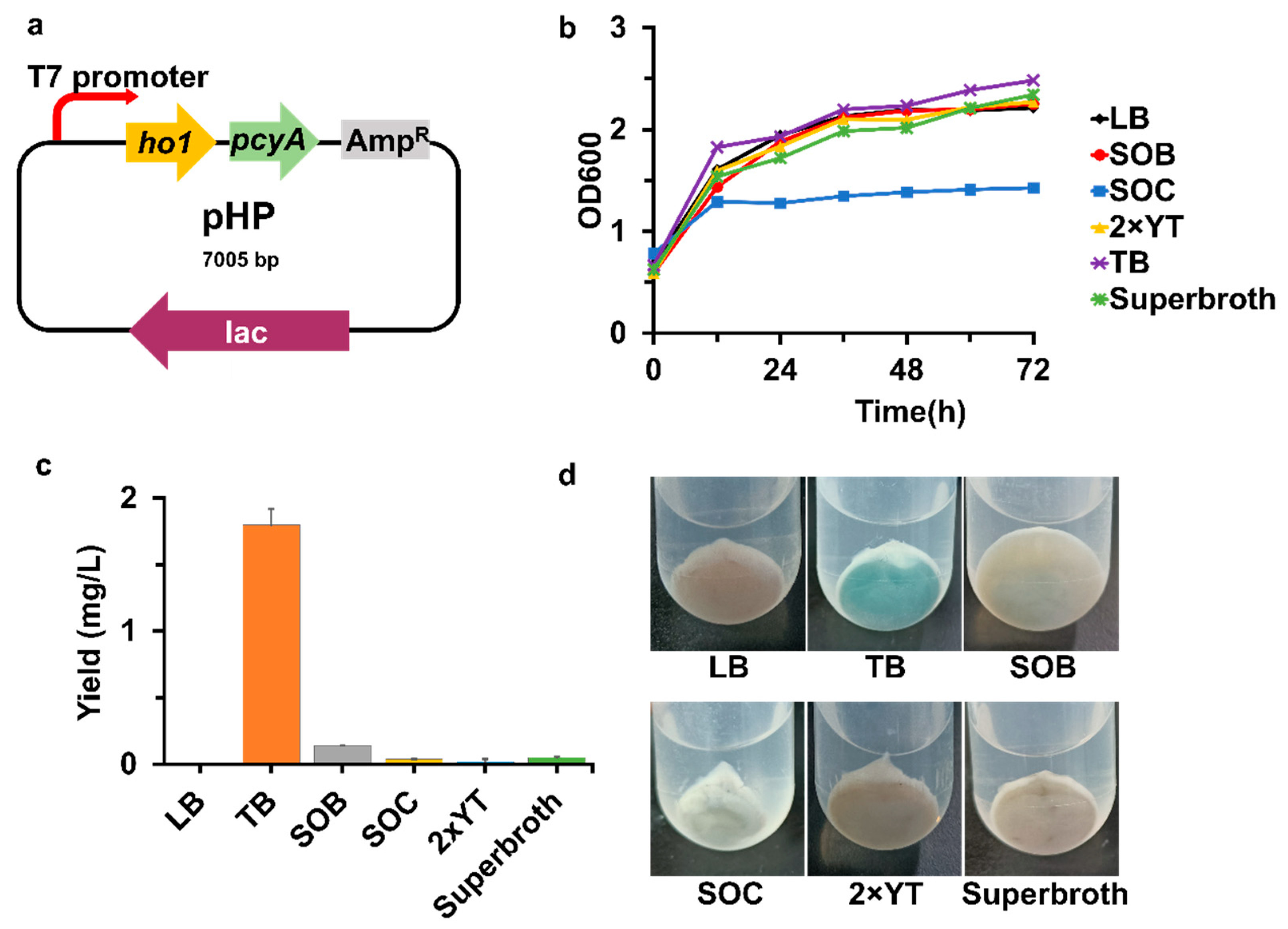
Figure 3.
The effects of different apoproteins on the yield of PCB. (a) Schematic representation of pBS and pBT; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
Figure 3.
The effects of different apoproteins on the yield of PCB. (a) Schematic representation of pBS and pBT; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
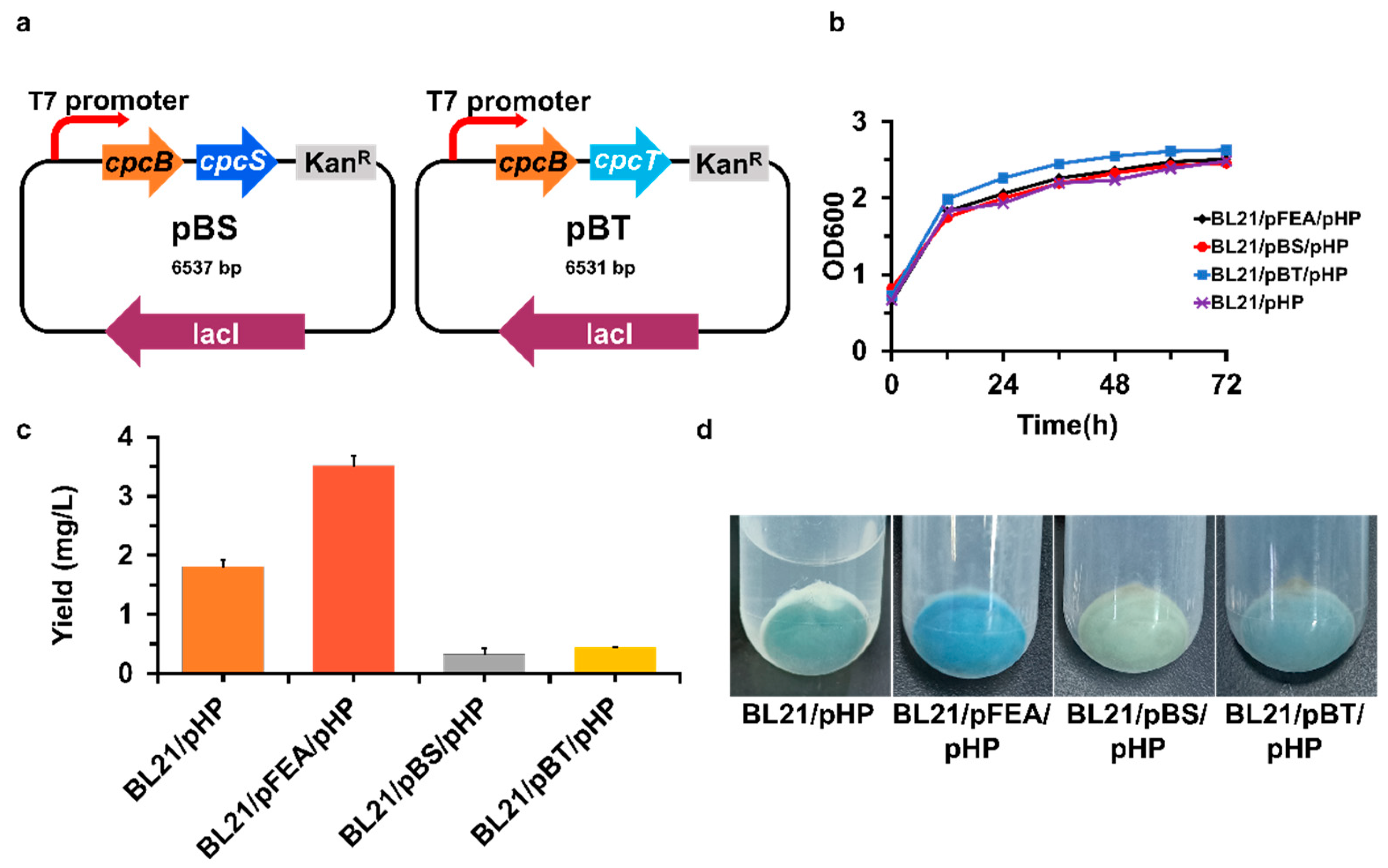
Figure 4.
The effects of signal peptides on the yield of PCB. (a) Schematic representation of pFEA and pFENA; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
Figure 4.
The effects of signal peptides on the yield of PCB. (a) Schematic representation of pFEA and pFENA; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.

Figure 5.
The PCB production was improved in the antibiotic-free system, by the yield of PCB. (a) Schematic representation of pDHP and pAHP; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.
Figure 5.
The PCB production was improved in the antibiotic-free system, by the yield of PCB. (a) Schematic representation of pDHP and pAHP; (b) The OD600 of different recombinant strains; (c) The PCB production of different recombinant strains; (d) The cell pellets of different recombinant strains. OD600 represents cell density. Induction time was the time of lysis protein expression after adding IPTG. The color of cell pellets represents: the darker the blue, the more PCB manufacture.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Signal sequences used for the secretory production of CpcA/CpcB and their expression level.
Table 1.
Signal sequences used for the secretory production of CpcA/CpcB and their expression level.
| Signal Sequences | Amino Acid Sequences | Expression Level | |
|---|---|---|---|
| CpcA | CpcB | ||
| pelB (pectate lyase B) [25] | MKYLLPTAAAGLLLLAAQPAMA | + | − |
| Kp-SP [32] | MSRRAPLLRAGAATAVAVLYLTAVPQAASA | + | − |
| PhoA (alkaline phosphatase) [25] | MKQSTIALALLPLLFTPVTKA | + | + |
| OmpA (outer membrane protein A) [25] | MKKTAIAIAVALAGFATVAQA | − | − |
| MalE (maltose-binding protein) [25] | MKIKTGARILALSALTTMMFSASALA | + | ++ |
| Lpp (murein lipoprotein) [25] | MKATKLVLGAVILGSTLLAG | − | − |
| N20 [33] | MEGNTREDNFKHLLGNDNVKRPSEA | +++ | |
| Native apo-CpcA | ++ | ||
| Native apo-CpcB | ++ | ||
“−” represents no protein production; “+”, “++” and “+++” represent less protein expression output from low to high.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, X.; Yu, J.; Che, Q.; Zhu, T.; Li, D.; Zhang, G. Enhanced Phycocyanobilin Production in Escherichia coli by Fusion-Expression of Apo-Proteins with Signal Peptides. Fermentation 2023, 9, 851. https://doi.org/10.3390/fermentation9090851
AMA Style
Liu X, Yu J, Che Q, Zhu T, Li D, Zhang G. Enhanced Phycocyanobilin Production in Escherichia coli by Fusion-Expression of Apo-Proteins with Signal Peptides. Fermentation. 2023; 9(9):851. https://doi.org/10.3390/fermentation9090851
Chicago/Turabian StyleLiu, Xiaolin, Jing Yu, Qian Che, Tianjiao Zhu, Dehai Li, and Guojian Zhang. 2023. "Enhanced Phycocyanobilin Production in Escherichia coli by Fusion-Expression of Apo-Proteins with Signal Peptides" Fermentation 9, no. 9: 851. https://doi.org/10.3390/fermentation9090851
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.