
Research Progress in Understanding the Molecular Biology of Cordyceps militaris
by

Lihong Wang
1,
Ganghua Li
2,
Xueqin Tian
1,3,
Yitong Shang
1,3,
Huanhuan Yan
1,3,
Lihua Yao
1,3 and
Zhihong Hu
1,3,4,* 1
College of Life Science, Jiangxi Science & Technology Normal University, Nanchang 330013, China
2
Hubei Key Laboratory of Edible Wild Plants Conservation and Utilization, Hubei Normal University, Huangshi 435002, China
3
Jiangxi Key Laboratory of Bioprocess Engineering, Jiangxi Science & Technology Normal University, Nanchang 330013, China
4
Key Laboratory of Microbial Resources and Metabolism of Nanchang City, Nanchang 330013, China
*
Author to whom correspondence should be addressed.
Fermentation 2024, 10(3), 167; https://doi.org/10.3390/fermentation10030167
Submission received: 18 January 2024
/
Revised: 12 March 2024
/
Accepted: 13 March 2024
/
Published: 15 March 2024
(This article belongs to the Special Issue Production of Nutritional and Functional Properties in Genetically Engineered Microorganisms)
Abstract
:Cordyceps militaris (C. militaris) is a valued medicinal fungus that can be traced back thousands of years in traditional Chinese medicine (TCM). Both TCM and modern scientific research have confirmed the positive effects of C. militaris on human health. In recent years, C. militaris has gained wide popularity; unfortunately, strains often degrade during cultivation, resulting in a decline in fruiting bodies and active components that negatively impacts the development of C. militaris in the health food and medicine industries. This review summarizes the current progresses in research on the genomic, transcriptomic, proteomic, and genetic manipulation of C. militaris and discusses its primary metabolites and strain degradation mechanisms. The current challenges and future prospects of C. militaris research are also discussed.
1. Introduction
Cordyceps militaris (C. militaris), belonging to Cordycipitaceae (Hypocreales, Ascomycota), is a well-known entomopathogenic fungus with rich nutritional value [1,2,3]. In Asia, especially in China, C. militaris is a famous tonic food and traditional Chinese medicine (TCM); it contains essential amino acids, with the most abundant being tyrosine and leucine [4]. C. militaris also contains a higher content of polysaccharides, vitamins, carotenoids, and other compounds compared to common plant foods. Considering its medicinal value, it has been used in alternative medicines to treat kidney, liver, neurological, and cardiovascular diseases, as well as to treat tumors and improve immunity [5,6]. Although a variety of compounds have been extracted from C. militaris, it is not the most popular member of the Cordyceps family. Similar to C. militaris, Cordyceps sinensis (C. sinensis) has been used to improve renal, hepatic, and respiratory functions; delay fatigue; and reduce type 2 diabetes symptoms. It has also been used as an aphrodisiac [7]. In China, C. sinensis has been listed as an endangered species due to a sharp decline in production and over-harvesting caused by high market demand. For these reasons, there is an active search for alternative sources that are already being produced under large-scale conditions, and C. militaris is used as an artificial cultured alternative due to its ease of production. As a model Cordyceps species, C. militaris is relatively more widely studied compared to C. sinensis [8]. There has been increasing attention paid to its unique pharmacological properties. More research has been conducted on the active metabolites of C. militaris [9], such as cordycepin (COR), cordycepic polysaccharide, and cordycepic acid [10,11,12,13,14,15,16,17], which are not significantly different from those in C. sinensis. Among these compounds, COR is regarded as an important active secondary metabolite with therapeutic potential. Pharmacological studies have reported that COR has antibacterial [18,19], tumor-relieving [20,21], antioxidant [22,23], anti-inflammatory [24,25], immune-regulatory [22], and lipid-lowering [26] properties. It also shows antiviral effects towards several viruses including influenza virus [27], HIV [28], dengue virus [29], Epstein–Barr virus (EBV) [30,31], and hepatitis C virus (HCV) [32]. Recent studies indicate that COR may also have effective treatment potential against coronavirus disease 2019 (COVID-19) [33,34]. Notably, the COR content in C. militaris is higher than that in wild C. sinensis while its price is much lower. In recent years, novel carotenoids with good water solubility have been found in C. militaris fruiting bodies, which have different characteristics from fat-soluble carotenoids. These carotenoids have expanded the application of C. militaris in healthcare, where it is considered to be a safe natural drug without side effects. Its fruiting body can be easily cultivated and contains more metabolites than the vegetative mycelium. C. militaris fruiting body powder and capsules have been found to improve micturition and prostate size in patients with benign prostatic hyperplasia [35].
The popularity and demand for C. militaris have led to its scarcity, also leading it to be listed as a protected endangered species. To meet the demand, significant research has been conducted on artificial cultivation and fermentation. Several challenges encountered during cultivation, especially strain degradation, affect its large-scale production. Strain degradation in the subculture process usually manifests as the absence or reduction of fruiting body production [36], which not only leads to a decrease in the yield of C. militaris metabolites but also affects the strain quality and causes economic losses [37]. To eliminate these issues, it is necessary to study the degradation mechanism and related genes that affect the biosynthesis of important metabolites. Many studies have focused on the molecular analysis of C. militaris, including manipulation of its genome, transcriptome, proteome, and genetics. Genetic engineering is a positive choice to increase production performance. However, the understanding of its molecular biology has been limited due to the lack of genome sequence information. It was not until 2011 that Zheng et al. [38] analyzed the complete molecular mechanism of COR biosynthesis through bioinformatics and gene function analysis of 18 Cordyceps species, including C. militaris, C. sinensis, and Cordyceps cicadae (C. cicadae) [39,40]. There has been great progress in transcriptomic and proteomic studies on C. militaris. Advances in multi-omics technology have enabled the analysis of the secondary metabolite biosynthesis pathways and their regulatory mechanisms, and the published genome has facilitated further molecular biological study [38].
This review primarily focuses on recent findings from genomic, transcriptomic, proteomic, and genetic manipulation studies of C. militaris. Additionally, the biosynthesis mechanisms of COR and carotenoids are discussed along with recent advances in understanding strain degradation. This study highlights the recent advancements in C. militaris research and its application prospects, providing a reference for further exploration of the mechanisms and interactions of its active components.
2. C. militaris Genome Analysis
In 2011, Zheng et al. [38] performed second-generation high-throughput whole-genome sequencing of the C. militaris strain Cm01. This strain has seven chromosomes ranging between 2.0 and 32.2 Mb that encode 9684 protein-producing genes. The publication of the genome of this strain has greatly promoted C. militaris research using other omics and molecular biology techniques. Cm01 has a similar genome structure to other Cordyceps strains (Table 1). In 2017, Kramer and Nodwell [41] used third-generation single-molecule sequencing to conduct chromosome-level assembly of the genome of the C. militaris strain ATCC 34164, which contained seven chromosomes ranging between 1.9 and 33.6 Mb with 9371 genes. In 2019, Chen et al. [42] conducted whole-genome sequencing of the C. militaris strain HN, assembling a total of 14 contigs ranging between 0.35 and 4.58 Mb with an N50 of 2.86 Mb and a genome size of 32.6 Mb. This strain was predicted to contain approximately 10,095 genes, which is higher than that found in Cm01 and ATCC 34164. Through comparison with Cm01 and ATCC 34164, it was found that there might be interchromosomal translocation among different strains of Cordyceps. Identification of genomic structural variants of Cordyceps showed that there are 406 structural variations between HN and Cm01, and 1114 structural variations between HN and ATCC 34164. The translocation and structural variants often lead to changes in the number and order of related genes, resulting in changes in the function of these genes that might be associated with secondary metabolism [43]. These findings contribute to our understanding of Cordyceps biology and the biosynthesis pathways of bioactive components.
Cordyceps and Metarhizium species are well known entomopathogenic fungi that have independently evolved insecticidal properties [44]. Until recently, the genes responsible for insect pathogenicity and bioactive component biosynthesis have been identified through comparative genome analysis; this provides a basis for studying the biological characteristics of C. militaris and will promote the development of the medicinal compounds it produces. Through the analysis and comparison of the genomes of several entomopathogenic fungi, it was found that the number of protein-coding genes in C. militaris is lower than that in Metarhizium anisopliae (M. anisopliae, 10817) [45], Metarhizium robertsii (M. robertsii, 10582) [46], and Metarhizium acridum (M. acridum, 9849) (Table 2) [47]. However, C. militaris has more species-specific genes (1328, 13.7%) compared to M. acridum (343, 3.5%) and M. anisopliae (509, 4.8%) [38]. Significant differences were observed in gene contents between these two genera that might be related to different survival strategies. In 2014, Staats et al. [45] compared the genome sizes of these entomopathogenic fungi and found that the C. militaris genome (32.2 Mb) is smaller than that of M. robertsii (39.04 Mb), M. anisopliae (38.5 Mb), and M. acridum (38.05 Mb); it also has the highest number of species-specific genes (2245) compared to M. acridum (875), M. anisopliae (690), and M. robertsii (603). This is consistent with the findings of Zheng et al. [38]. Gene family expansions of proteases and chitinases in C. militaris were identified with comparison to Metarhizium species. These two types of enzymes can degrade insect body cell walls, indicating that the pathogenic strategy of C. militaris is to invade insects by breaking down their epidermis. The pathogenic strategies of Cordyceps and Metarhizium species may differ due to differences in gene content. In addition, Xia et al. [39] conducted a comparative analysis of C. militaris with Aspergillus nidulans (A. nidulans, another COR-producing fungus) to identify four physically linked genes (CCM_04436–CCM_04439 vs. AN3333–AN3329), respectively, which were associated with COR biosynthesis (discussed in detail in Section 6).
3. C. militaris Transcriptome Analysis
C. militaris shows typical sexual and asexual stages, which manifest as fruiting bodies and mycelia, respectively. The transcriptome profiles of C. militaris at specific developmental stages are distinct (Table 3) [48]. Differentially expressed gene (DEG) analysis was performed, and a transcription-level comparison between mycelia and fruiting bodies showed significant differences: C. militaris TBRC6039 had 1877 DEGs, among which 935 and 942 genes were up-regulated in the mycelia and fruiting bodies, respectively [49]. In addition, genes involved in lipid metabolism were up-regulated in fruiting bodies compared to mycelia. It has been reported that the neutral lipids in lipid droplets were required during COR accumulation in C. militaris [50]. This might explain why the phenomenon of high COR content was associated with lipid accumulation in the fruiting body stage. These studies suggest that these stage-specific genes may have important functions in fungal development; however, they did not give insights into many other biological processes, such as strain degradation.
Strain degradation is a common phenomenon during C. militaris subculture. Several complex physiological and biochemical changes occur during strain degradation, along with changes in the expression of related genes. Overcoming strain degradation has become a primary target for the factory production of C. militaris. Yin et al. [36] subcultured the C. militaris strain YCC for six successive generations and compared the growth of its fruiting bodies. They found that C. militaris begins to retrogress from the third generation, with its fruiting bodies being underdeveloped in the fourth generation and absent in the fifth generation. Transcriptome profiles of these six generations revealed 1892, 2498, 2006, 2273, and 2188 DEGs in the second, third, fourth, fifth, and sixth generations, respectively, compared with the first generation. The number of DEGs in the third generation was the highest, indicating that an increase in passage numbers (and, consequently, an increase in DEGs) exacerbates strain degradation. The morphological changes and mechanisms of C. militaris strain degradation will be discussed in detail in Section 8.
Mycelium growth and bioactive metabolite biosynthesis are also affected by medium and culture conditions. In our previous study, C. militaris growth was inhibited by salt treatments, but COR production increased significantly with an increase in salt concentration. Transcriptome analysis of the salt-treated and control C. militaris groups revealed 3885 DEGs, among which DEGs associated with amino sugar and nucleotide sugar pathways were significantly enriched in the salt-treated group [51]. Salt treatment might increase energy molecule production and further promote COR accumulation. There exists sufficient evidence that COR biosynthesis is related to different carbon sources, but its regulatory mechanisms have rarely been studied. To explore these, Raethong et al. [52] cultured the C. militaris strain TBRC6039 using sucrose, glucose, and xylose as different carbon sources. It was found that the growth rate and biomass of xylose were much lower than those of sucrose and glucose. Thus, the cultivation time in xylose was much longer than that in sucrose and glucose for COR measurement, and the results showed the extracellular COR titer (mg/L) in xylose culture was the highest. However, the extracellular COR productivity (mg/L/day) was still lower than that of sucrose and glucose due to the prolonged cultivation in xylose culture [52]. Consistent with the results, Wongsa et al. [53] also found xylose inhibited mycelial growth and biomass. However, the xylose culture showed the highest yield of extracellular COR on dry biomass, although it was a poor carbon source for mycelial growth [53]. Transcriptome analysis showed that there were much fewer DEGs between glucose and sucrose cultures (858) than in cultures grown with xylose versus sucrose (1799) and xylose versus glucose (2098). This finding might be related to the hydrolysis of sucrose to glucose and fructose [54]. Overall, the effects of xylose metabolism on COR production might be a complex process, and further studies are needed to gain a better understanding.

{kind=link}
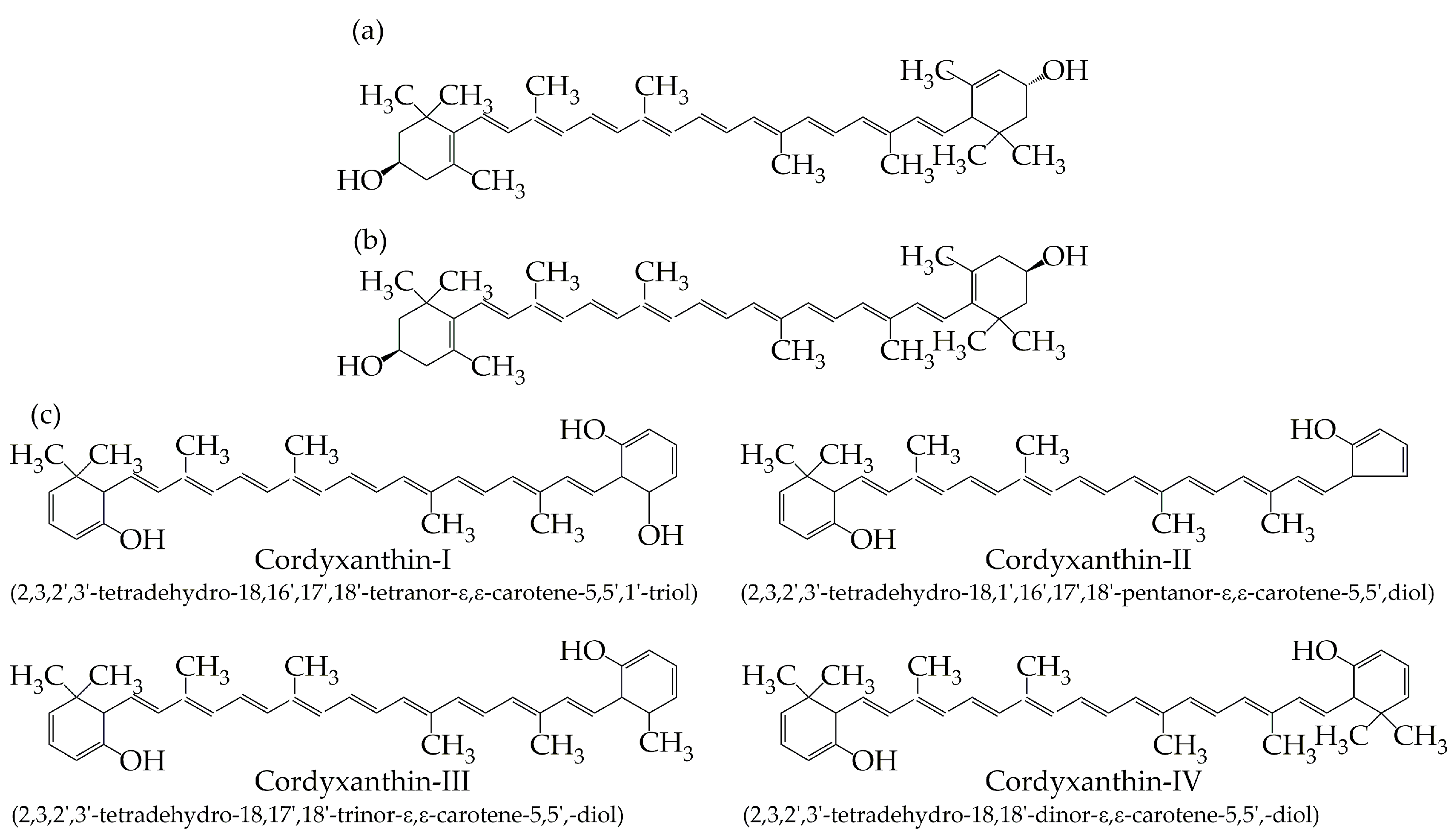{kind=link}
{kind=link}
{kind=link}
Table 3.
Transcriptome of C. militaris.
| Strain | Sample Information | Main Results | Year | Ref. |
|---|---|---|---|---|
| C. militaris TBRC6039 | Mycelia and fruiting bodies | Compared the metabolic functions and regulation of C. militaris relevant to its developmental stages | 2023 | [49] |
| C. militaris YCC | Mycelia from 6 generations of subculture | Compared the changes in C. militaris for six generations during fruiting body growth | 2017 | [36] |
| C. militaris CM10 | Mycelia (cultured under light and dark conditions) | Discovered the genes related to the biosynthesis of C. militaris carotenoid | 2021 | [55] |
| C. militaris Cm01 | Mycelia with different salt treatment | Provided a global transcriptome characterization of the salt treatment adaptation process in C. militaris | 2021 | [51] |
| C. militaris TBRC6039 | Mycelia (cultured under three carbon sources) | Uncovering sucrose- and glucose-mediated changes in the transcriptional regulation of central carbon metabolism in C. militaris | 2018 | [52] |
| C. militaris TBRC7358 | Mycelia (cultured under three carbon sources) | Proposed putative alternative routes in formation of precursors for COR production on xylose | 2020 | [53] |
4. C. militaris Proteome Analysis
Presently, proteomic studies on C. militaris are limited. One proteomic study found 359 and 214 proteins in the C. militaris mycelium and fruiting body, respectively [56], among which 98 were expressed at both stages. There were also significant differences in protein expression between the two stages; this is consistent with the results of the transcriptomic analysis by Zheng et al. [38]. The C. militaris strain Cm01 contains 73 glycosylated phospholipid acyl-anchored proteins and is predicted to have 301 secretion-related proteins, accounting for 3.1% of the total number of proteins (9651) [45]. Similarly, Cordyceps kyushensis Kob (C. kyushuensis Kob) has 445 differentially expressed proteins [57], among which 224 are up-regulated and 221 are down-regulated between the two stages. These findings provide insights into the molecular mechanisms underlying the different life stages of C. militaris.
5. Genetic Manipulation of C. militaris
Commonly used methods for fungal transformation include Agrobacterium tumefaciens-mediated transformation (ATMT), polyethylene glycol (PEG)-mediated protoplast transformation (PMPT), CRISPR-Cas9, electroporation transformation, and gene gun-mediated transformation. At present, ATMT is the most developed technology and has been widely used in C. militaris. As early as 2011, Zheng et al. [58] constructed a C. militaris mutant library and identified the functions of these genes. He et al. [59] obtained a defective mutant using ATMT with a colony phenotype that was white after light exposure; the results showed that the Cmcrf1 gene was involved in fruiting body development and carotenoid biosynthesis. In 2019, Lou Haiwei et al. [55] transformed Cmtns into a deleted mutant (ΔCmtns) using ATMT, discovering that Cmtns significantly affects carotenoid biosynthesis; this provides an important basis for the future study and promotion of carotenoid production. It is necessary to further study these genes involved in carotenoid biosynthesis and their interaction with other genes in order to better understand the growth and development of C. militaris. ATMT is not only an effective tool to study gene functions, but it is also an important method to increase the production of secondary metabolites. Chen et al. [60] transferred ergothioneine (EGT) synthesis genes into the C. militaris genome using ATMT, increasing the production of EGT and COR. They also found that the overexpression of EGT synthesis genes in degraded strains of C. militaris could restore their production capacity, verifying its protective effect in degraded strains. PMPT methods using protoplasts as receptor cells have also been reported. Choi et al. [61] obtained green fluorescent protein (GFP) mutants using PMPT and established the genetic transformation system in C. militaris. Most transformation methods still face problems with low editing efficiency due to the difficulty of foreign gene transformation. Recently, CRISPR-Cas9 has been known as a site-specific genome editing method and is more accurate and efficient than ATMT and PMPT. In 2021, an optimized RNP-based method increased transformation efficiency in C. militaris, with editing efficiency reaching 100% and demonstrating the feasibility of gene editing in this species [62]. In summary, ATMT has more applications and higher transformation efficiency, followed by PMPT, and great challenges are faced in the application of CRISPR-Cas9 technology to fungal genome editing. However, it is not necessary to extract protoplasts for electroporation and gene gun-mediated transformation; their transformation efficiencies are low and they are rarely applied to fungal genetic transformation. The above studies provide important information for the establishment and development of fungal genetic transformation systems, contributing to gene isolation and functional identification in order to engineer C. militaris strains.
6. COR Biosynthesis in C. militaris
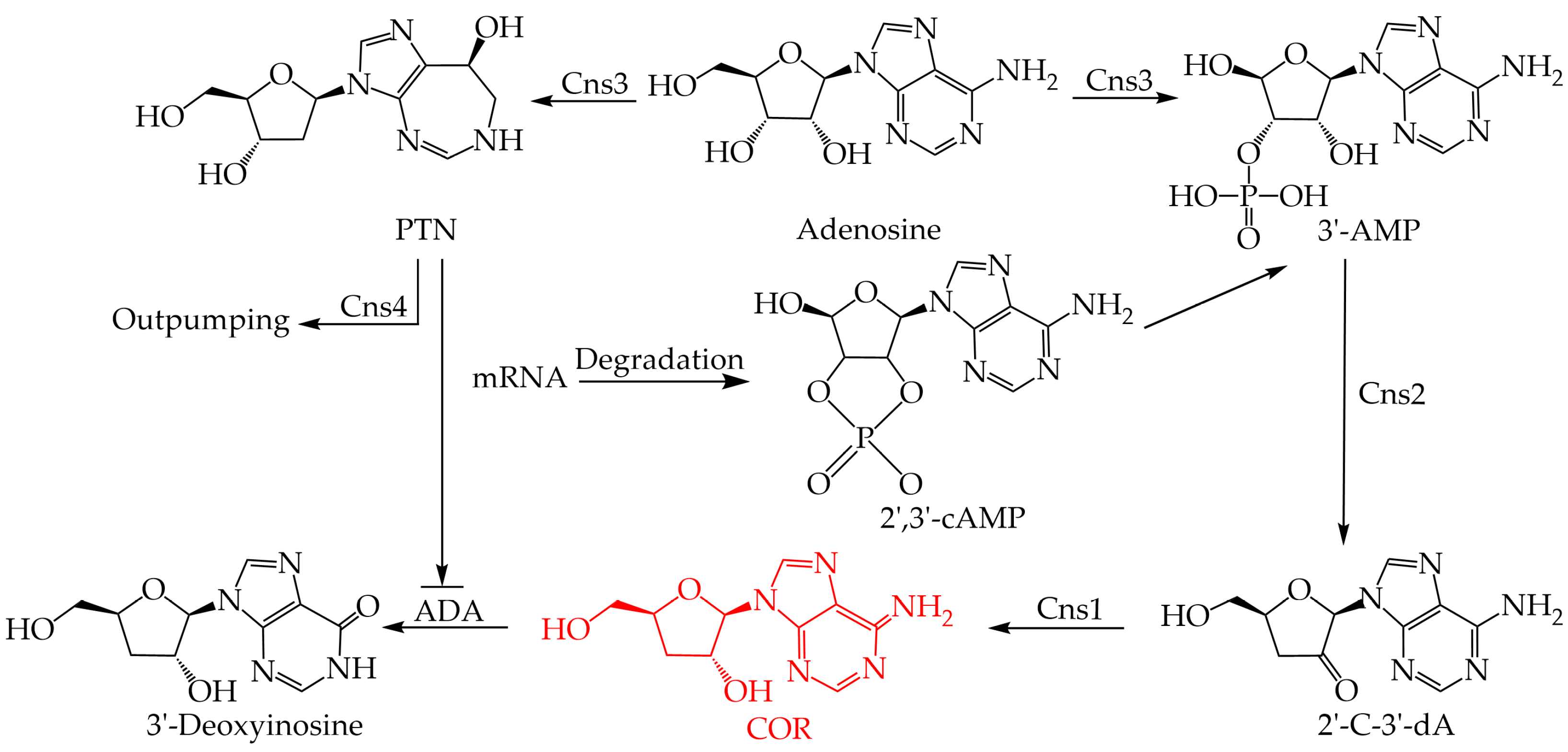
COR was isolated and identified from cultures of C. militaris for the first time in 1950 [63]. Over the past half-century, there has been great interest in detailing the COR biosynthesis pathway. However, few complete biosynthesis pathways have been uncovered due to the lack of C. militaris genomic data; it was not until 2011 that the whole genome of C. militaris was sequenced [38]. Subsequent studies identified gene clusters responsible for COR biosynthesis through comparative genomic analysis with A. nidulans and proposed a novel COR biosynthesis pathway (Figure 1) [39]. The COR biosynthesis gene cluster includes Cns1, Cns2, Cns3, and Cns4. COR is synthesized from adenosine and phosphorylated by Cns3 to produce the intermediate product adenosine-3′-monophosphate (3′-AMP). Subsequently, 3′-AMP is dephosphorylated by Cns2 and eventually reduced by Cns1 to the final product COR. In addition, C. militaris also has a self-detoxification mechanism that is used to regulate the excessive production of COR. The gene cluster of C. militaris can also synthesize penstatin (PTN, deoxycoformycin), which can inhibit the degradation of COR into 3′-deoxyinosine (3′-dI). When the intracellular COR content is too high and causes cytotoxicity, the intracellular PTN content decreases, and COR is catalyzed to produce non-toxic 3′-dI through adenosine deaminase (ADA) intracellular deamination. Similar biosynthesis pathways for COR have been found in other COR-producing fungi such as C. kyushensis Kob [57] and C. cicadae [40]. Four related genes, ck1-ck4, were identified in C. kyushensis Kob as being involved in the synthesis of COR, similar to Cns1-Cns4 in C. militaris, suggesting that there may be other routes involved in the COR biosynthesis pathway in C. militaris.
Differences in mycelial growth and COR biosynthesis are seen with different carbon sources. As described above, Raethong [51] and Wongsa et al. [53] found xylose inhibited the growth and biomass of C. militaris but increased COR biosynthesis on dry biomass. Consistent with this, transcriptome sequencing found that xylose significantly up-regulated genes involved in cell growth and COR biosynthesis, suggesting an important role for xylose in the formation of the precursor 3′-AMP. In addition, alternative routes involved in COR biosynthesis were proposed via transcriptome analysis of this two strains of C. militaris; this is consistent with the results of Xia et al. [39], who found that the COR biosynthesis pathway is highly correlated with the formation of the precursor 3′-AMP, which can be synthesized by 2′,3′-cyclic monophosphate (2′,3′-cAMP), a byproduct of mRNA degradation. However, the biochemical function and catalytic mechanism of the enzymes required for this pathway remain unknown. Further research is needed to prove whether this pathway is also present in other fungi.
7. Carotenoid Biosynthesis in C. militaris
C. militaris contains six types of pigments, including lutein, zeaxanthin, and four cordyxanthins (Figure 2) [64,65,66]. Lutein and zeaxanthin are insoluble in water, while cordyxanthin I, II, III, and IV are water-soluble carotenoids; they contain 36, 35, 37, and 38 carbon atoms, respectively, compared to the 40 carbon atoms usually found in most carotenoids [66]. As a class of rare water-soluble carotenoids, they potentially have higher physiological activity, which will expand the application potential of the C. militaris pigments used in food, cosmetics, and other fields. Until now, the content of carotenoids in C. militaris has remained low, and still cannot meet the market demand. It is necessary to study the biosynthesis pathways of C. militaris carotenoids and the related genes to improve the content of carotenoids through genetic engineering.
C. militaris pigments are considered to be carotenoids. However, there are few reports on carotenoid biosynthesis genes in C. militaris; only two genes at the end of the carotenoid biosynthesis pathway have been identified that have little effect on the entire pathway [67,68]. The pathway of carotenoid biosynthesis in Neurospora crassa (N. crassa) has been well studied. In N. crassa, carotenoid biosynthesis involves at least three enzymes, among which geranylgeranyl diphosphate synthase genes (CCM_03059, CCM_03697, and CCM_06355) are present in C. militaris [69]; the other two key enzymes, phytoene synthetase and phytoene dehydrogenase, are not found in C. militaris, indicating that its carotenoid biosynthesis pathway is different [55]. Based on the above, the absence of genes involved in the biosynthesis of carotenoids in C. militaris suggests that its biosynthesis pathway might be different from that of other species. Because of this, it is challenging to characterize the carotenoid biosynthesis pathway in C. militaris and explore the underlying mechanisms to solve the problem of low carotenoid yield.
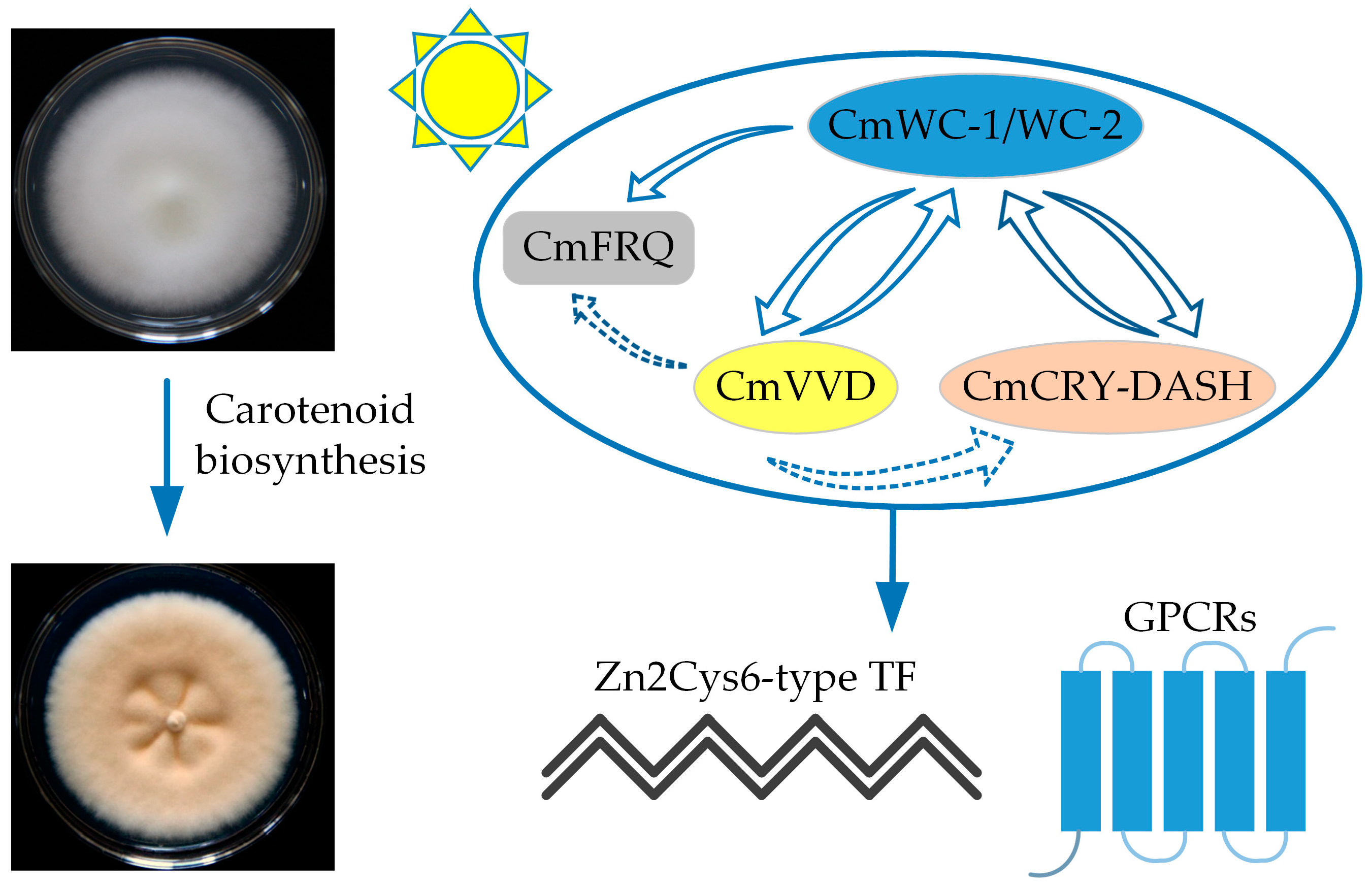
Light has a significant influence on C. militaris pigment formation [70]. The circadian clock protein FREQUENCY (FRQ) and blue light receptors White Colar (WC)-1 and WC-2 are important photoreceptors in fungi. Shrestha et al. [71] found that wild-type C. militaris colonies are orange or orange-yellow. The colony color is associated with the light intensity during culturing, which influences carotenoid production of C. militaris, resulting in orange pigmentation [67,72,73]. A subsequent study found an albino strain (505CGMCC) of C. militaris with a mutation in the photoreceptor gene Cmwc-1 [68]; the colony of the albino strain remained white under light while that of the wild-type strain (498CGMCC) changed from white to yellow. The carotenoid content was significantly lower in this mutant compared with the wild-type strain, but the COR content was high. This finding suggested that the formation of carotenoids in C. militaris was dependent on the photoreceptor CmWC-1. Yang et al. also found the colony color of the albino strain and a degraded one did not change or changed slower after exposure to light, and the light-induced transcription of Cmwc-1 is significantly increased in C. militaris [74]. This is consistent with the findings of Wang et al. [68]. Additionally, Cmwc-1 knockout strains have albinism, reduced fruiting body production, and significantly reduced carotenoid and COR production [67]. These results suggested that the Cmwc-1 gene is involved in the regulation of fruiting body development and carotenoid biosynthesis in C. militaris. In 2020, it was found that the interaction of CmWC-1 and CmFRQ promoters regulates fruiting body development and carotenoid and COR production [75]. These studies indicate that light greatly influences the production of C. militaris pigments and the development of its fruiting bodies (Figure 3) [76].
8. C. militaris Strain Degradation Mechanisms
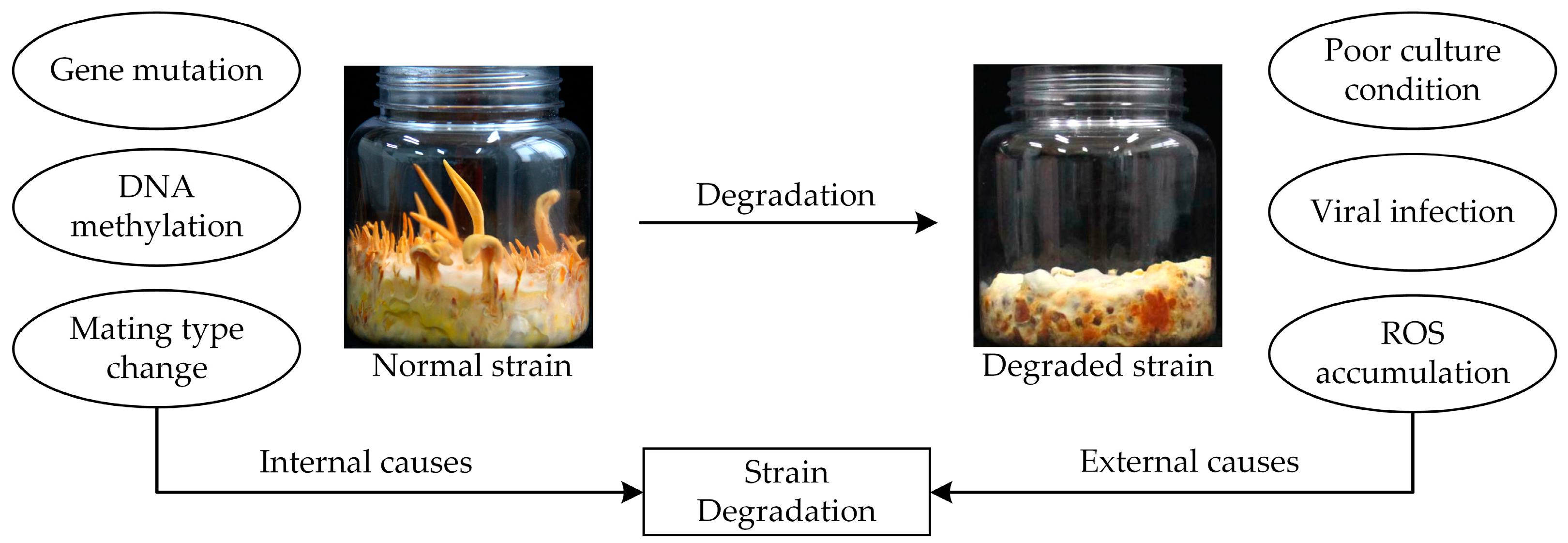
Filamentous fungi usually undergo degradation during the subculture process, leading to a sharp decline in fungal production that causes severe economic losses and drastically affects quality control. The colony morphology and physiological characteristics of degraded strains differ from those of wild-type strains [37]. This affects external and internal mycelial morphology. C. militaris degradation usually leads to the absence or reduction of fruiting body production [36,77] and decreased sporulation [78]. Studies have found significant decreases in the dehydrogenase activity and decolorization ability of degraded strains [79]; while wild-type strains gradually turn orange after light exposure, degraded strains only slightly change color or do not change color at all. In addition, strain degradation also leads to slower mycelial growth and metabolic capacity, and product yield decline, affecting production efficiency [37].
Strain degradation is a complex process involving many factors, including mating type, external environmental factors, and alternative splicing (AS) events [37]. Fungi can be divided into homokaryotic or heterothallic based on mating type; homokaryotic strains contain only one mating-type locus, whereas heterokaryotic strains contain both loci [80]. Wang et al. [81] found that wild-type strains have both mating-type loci, while degraded strains only have one mating-type locus and cannot produce fruiting bodies; this is consistent with the results of Yin et al. [14], who also found that degraded strains are homokaryotic. These results indicate that variations in the mating type may induce C. militaris degradation. It was unexpectedly observed in one study that the single mating strain C. militaris Cm01 could grow fruiting bodies without mating [38]. Additionally, only one mating-type locus has been identified in C. militaris, containing either MAT-α (MAT 1-1 gene) or MAT-hmg (MAT 1-2 gene) [82]. Overall, these results indicate that strain degradation is caused by many factors that may be associated with the change of mating type, though this needs to be further verified.
The external environment also influences C. militaris development and fruiting body production. Poor environmental conditions may lead to strain death or gene mutations that trigger strain degradation. Reactive oxygen species (ROS) are closely related to spontaneous degradation in C. militaris [37]. Exogenous melatonin (MT) is reported to scavenge oxygen-free radicals in fungal cells, and Zu et al. [83] found that the degradation characteristics of a degraded strain of C. militaris under MT treatment were significantly lessened. RNA-seq revealed that compared with the control, glutathione s-transferase genes were up-regulated under MT treatment. These studies indicate that ROS damage is one of the key factors affecting C. militaris strain degradation.
As mentioned above, over 5000 AS events were detected after each subculture of C. militaris; 6230, 6170, and 6320 AS events were detected in the second, fourth, and sixth generations, respectively. These results indicate an increased number of passages leads to enhanced strain degradation, suggesting that degradation may be directly associated with AS events [36]. In a previous study, the DNA methylation levels of a wild-type and degraded strain were found to be 0.48% and 0.56%, respectively, with 188 differential methylation regions (DMRs) [84]. DNA methylation has important effects on cell differentiation and development; the methylation level of wild-type strains was stable while that of the degraded strain was increased. This indicates that DNA methylation and recombination can induce C. militaris strain degradation.
Appropriate preservation methods can prevent fungal strain degradation. Since the number of generations is directly associated with enhanced strain degradation, maintaining the number of passages to ≤3 can be an appropriate strain preservation strategy. Additionally, repeated subculturing is prone to contamination, and genetic and physiological changes often occur during long-term preservation. Cryopreservation seems to be a very efficient way to maintain the purity and vitality of strains for a long time [85]; for example, the activity of C. militaris strains has been shown to be maintained for ≥1 year in sterile water or liquid nitrogen at −196 °C [86]. However, some studies have suggested that ultra-low-temperature freezing at −80 °C is not suitable for the preservation of C. militaris strains [87]. As results are contradictory, there is an urgent need to determine an appropriate culture and preservation method. Although freezing can delay degradation, it cannot restore the performance of degraded strains; breeding optimal strains of C. militaris is key to overcome strain degradation. In a previous study, cross-mating of a single ascospore from a degraded strain restored its ability to produce fruiting bodies, COR, and adenosine [77], indicating that cross-mating is an effective method to rejuvenate degraded strains.
In summary, gene mutation is the primary cause of strain degradation, followed by poor culture and strain preservation conditions, which can induce genotypic or phenotypic changes, affecting production performance (Figure 4) [36,88,89,90]. Although factory production in the C. militaris industry has been successfully established, the key technology to overcome strain degradation has not yet been discovered, and the mechanisms associated with degradation are still unknown; the above studies contribute to understanding the causes of strain degradation and the countermeasures that can be used to counteract and prevent it.
9. Conclusions
At present, artificial culture and large-scale production of C. militaris have somewhat alleviated issues of low availability and high prices in wild Cordyceps; however, low metabolite yield, strain degradation, and complex biosynthesis pathways continue to be challenges facing the Cordyceps industry. Although several attempts have been made to enhance C. militaris yield, most studies were conducted with different strains; the physiological characteristics of strains vary greatly, and it is not applicable to generalize the results from one strain to others, even within the same species. Additional studies examining multiple aspects of C. militaris should be carried out. There have been many studies on the cultivation of C. militaris; considering the strain degradation, it is necessary to systematically optimize factory production and further promote the healthy and stable development of the C. militaris industry. C. militaris has great application value in various industries, including medicine, beauty, and healthcare fields, making it a key organism of interest for further study. It contains multiple secondary metabolite gene clusters, indicating its potential for secondary metabolism studies. The development of molecular biology and genetic technology will allow researchers to explore new synthetic routes and obtain new compounds. In this study, recent findings in genomic, transcriptomic, proteomic, and genetic manipulation studies on C. militaris were discussed, along with its primary metabolites and strain degradation mechanisms. This review will help increase the understanding of the research developments and applications of C. militaris cultivation.
Author Contributions
Z.H. designed the manuscript. L.W. wrote the manuscript; L.Y. and G.L. participated in the collecting of data; X.T., Y.S. and H.Y. contributed to the revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (NSFC grant no. 32260009) and the Open Foundation of Hubei Key Laboratory of Edible Wild Plants Conservation and Utilization (EWPL202207).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
We thank Mingquan Yu for the funding support and revision suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wu, F.; Zhou, L.-W.; Yang, Z.-L.; Bau, T.; Li, T.-H.; Dai, Y.-C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Jiaojiao, Z.; Fen, W.; Kuanbo, L.; Qing, L.; Ying, Y.; Caihong, D. Heat and light stresses affect metabolite production in the fruit body of the medicinal mushroom Cordyceps militaris. Appl. Microbiol. Biotechnol. 2018, 102, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Kunhorm, P.; Chaicharoenaudomrung, N.; Noisa, P. Enrichment of cordycepin for cosmeceutical applications: Culture systems and strategies. Appl. Microbiol. Biotechnol. 2019, 103, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Aung, W.L.; Kyaw, M. Identification and Determination of Secondary Metabolites and Amino Acids in Cordyceps. Partn. Univers. Int. Innov. J. 2023, 1, 251–258. [Google Scholar]
- Song, Q.; Zhu, Z. Using Cordyceps militaris extracellular polysaccharides to prevent Pb 2+-induced liver and kidney toxicity by activating Nrf2 signals and modulating gut microbiota. Food Funct. 2020, 11, 9226–9239. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Shin, H.-S.; Leyva-Gómez, G.; Prado-Audelo, M.L.D.; Cortes, H.; Singh, Y.D.; Panda, M.K.; Mishra, A.P.; Nigam, M.; Saklani, S. Cordyceps spp.: A review on its immune-stimulatory and other biological potentials. Front. Pharmacol. 2021, 11, 2250. [Google Scholar] [CrossRef]
- Van Nguyen, T.; Chumnanpuen, P.; Parunyakul, K.; Srisuksai, K.; Fungfuang, W. A study of the aphrodisiac properties of Cordyceps militaris in streptozotocin-induced diabetic male rats. Vet. World. 2021, 14, 537–544. [Google Scholar] [CrossRef]
- Dong, C.; Guo, S.; Wang, W.; Liu, X. Cordyceps industry in China. Mycology 2015, 6, 121–129. [Google Scholar] [CrossRef]
- Wei, H.-P.; Ye, X.-L.; Chen, Z.; Zhong, Y.-J.; Li, P.-M.; Pu, S.-C.; Li, X.-G. Synthesis and pharmacokinetic evaluation of novel N-acyl-cordycepin derivatives with a normal alkyl chain. Eur. J. Med. Chem. 2009, 44, 665–669. [Google Scholar] [CrossRef]
- Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.-D.; Khanh, T.D.; Teschke, R. Cordycepin isolated from Cordyceps militaris: Its newly discovered herbicidal property and potential plant-based novel alternative to glyphosate. Molecules 2019, 24, 2901. [Google Scholar] [CrossRef]
- Lan, D.; Thu, N.; Lan, P.; Nha, P.; Tung, B. Cordyceps militaris (L.) link: Chemical bioactive compounds and pharmacological activities. J. Pharm. Nutr. Sci. 2016, 6, 153–159. [Google Scholar] [CrossRef]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An overview of its chemical constituents in relation to biological activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.-Y.; Liu, X.-C.; Dong, F.-Y.; Guo, M.-Z.; Wang, X.-T.; Wang, Z.; Zhang, Y.-M. Influence of fermentation conditions on polysaccharide production and the activities of enzymes involved in the polysaccharide synthesis of Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 3909–3921. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Xin, X.D.; Weng, Y.J.; Li, S.H.; Jia, J.Q.; Gui, Z.Z. Genotypic analysis of degenerative Cordyceps militaris cultured in the pupa of Bombyx mori. Entomol. Res. 2018, 48, 137–144. [Google Scholar] [CrossRef]
- Wang, F.; Song, X.; Dong, X.; Zhang, J.; Dong, C. DASH-type cryptochromes regulate fruiting body development and secondary metabolism differently than CmWC-1 in the fungus Cordyceps militaris. Appl. Microbiol. Biotechnol. 2017, 101, 4645–4657. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Chen, S.; Xie, Y.; Chen, D.; Su, J.; Shuai, O.; Jiao, C.; Zuo, D. Cordycepin, a characteristic bioactive constituent in Cordyceps militaris, ameliorates hyperuricemia through URAT1 in hyperuricemic mice. Front. Microbiol. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Wei, T.; Lin, Y.; Ye, Z.-W.; Lin, J.-F.; Guo, L.-Q.; Yun, F.; Kang, L. Developing a novel two-stage process for carotenoid production by Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms. 2019, 21, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lou, Z.; Wang, H.; Chen, C. Antimicrobial effect and proposed action mechanism of cordycepin against Escherichia coli and Bacillus subtilis. J. Microbiol. 2019, 57, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cai, G.; He, Y.; Tong, G. Separation of cordycepin from Cordyceps militaris fermentation supernatant using preparative HPLC and evaluation of its antibacterial activity as an NAD+-dependent DNA ligase inhibitor. Exp. Ther. Med. 2016, 12, 1812–1816. [Google Scholar] [CrossRef]
- Jeong, M.-H.; Lee, C.-M.; Lee, S.-W.; Seo, S.-Y.; Seo, M.-J.; Kang, B.-W.; Jeong, Y.-K.; Choi, Y.-J.; Yang, K.-M.; Jo, W.-S. Cordycepin-enriched Cordyceps militaris induces immunomodulation and tumor growth delay in mouse-derived breast cancer. Oncol. Rep. 2013, 30, 1996–2002. [Google Scholar] [CrossRef]
- Wu, W.-C.; Hsiao, J.-R.; Lian, Y.-Y.; Lin, C.-Y.; Huang, B.-M. The apoptotic effect of cordycepin on human OEC-M1 oral cancer cell line. Cancer Chemother. Pharmacol. 2007, 60, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, Y.; Cui, Y.; Liu, H.; Dong, C.; Sun, Y. Structural characterization, antioxidant and immunomodulatory activities of a neutral polysaccharide from Cordyceps militaris cultivated on hull-less barley. Carbohydr. Polym. 2020, 235, 115969. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Gu, S.; Pan, L.; Sun, H.; Gong, E.; Zhu, Z.; Wen, T.; Daba, G.M.; Elkhateeb, W.A. Structure analysis and antioxidant activity of polysaccharide-iron (III) from Cordyceps militaris mycelia. Int. J. Biol. Macromol. 2021, 178, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Won, S.-Y.; Park, E.-H. Anti-inflammatory and related pharmacological activities of cultured mycelia and fruiting bodies of Cordyceps militaris. J. Ethnopharmacol. 2005, 96, 555–561. [Google Scholar] [CrossRef]
- Phull, A.-R.; Ahmed, M.; Park, H.-J. Cordyceps militaris as a bio functional food source: Pharmacological potential, anti-inflammatory actions and related molecular mechanisms. Microorganisms 2022, 10, 405. [Google Scholar] [CrossRef]
- Gao, J.; Lian, Z.-Q.; Zhu, P.; Zhu, H.-B. Lipid-lowering effect of cordycepin (3’-deoxyadenosine) from Cordyceps militaris on hyperlipidemic hamsters and rats. Yao Xue Xue Bao Acta Pharm. Sin. 2011, 46, 669–676. [Google Scholar]
- Li, R.-F.; Zhou, X.-B.; Zhou, H.-X.; Yang, Z.-F.; Jiang, H.-M.; Wu, X.; Li, W.-J.; Qiu, J.-J.; Mi, J.-N.; Chen, M. Novel fatty acid in Cordyceps suppresses influenza A (H1N1) virus-induced proinflammatory response through regulating innate signaling pathways. ACS Omega 2021, 6, 1505–1515. [Google Scholar] [CrossRef]
- Jiang, Y.; Wong, J.; Fu, M.; Ng, T.; Liu, Z.; Wang, C.; Li, N.; Qiao, W.; Wen, T.; Liu, F. Isolation of adenosine, iso-sinensetin and dimethylguanosine with antioxidant and HIV-1 protease inhibiting activities from fruiting bodies of Cordyceps militaris. Phytomedicine 2011, 18, 189–193. [Google Scholar] [CrossRef]
- Panya, A.; Songprakhon, P.; Panwong, S.; Jantakee, K.; Kaewkod, T.; Tragoolpua, Y.; Sawasdee, N.; Lee, V.S.; Nimmanpipug, P.; Yenchitsomanus, P.-T. Cordycepin inhibits virus replication in dengue virus-infected vero cells. Molecules 2021, 26, 3118. [Google Scholar] [CrossRef]
- Ryu, E.; Son, M.; Lee, M.; Lee, K.; Cho, J.Y.; Cho, S.; Lee, S.K.; Lee, Y.M.; Cho, H.; Sung, G.-H. Cordycepin is a novel chemical suppressor of Epstein-Barr virus replication. Oncoscience 2014, 1, 866–881. [Google Scholar] [CrossRef]
- Du, Y.; Yu, J.; Du, L.; Tang, J.; Feng, W.-H. Cordycepin enhances Epstein–Barr virus lytic infection and Epstein–Barr virus-positive tumor treatment efficacy by doxorubicin. Cancer Lett. 2016, 376, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Mori, K.; Satoh, S.; Dansako, H.; Ikeda, M.; Kato, N. Anti-HCV activity of the Chinese medicinal fungus Cordyceps militaris. Biochem. Biophys. Res. Commun. 2014, 447, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K. Cordycepin: A bioactive metabolite of Cordyceps militaris and polyadenylation inhibitor with therapeutic potential against COVID-19. J. Biomol. Struct. Dyn. 2022, 40, 3745–3752. [Google Scholar] [CrossRef] [PubMed]
- Dubhashi, S.; Sinha, S.; Dwivedi, S.; Ghanekar, J.; Kadam, S.; Samant, P.; Datta, V.; Singh, S.; Chaudry, I.H.; Gurmet, P. Early Trends to Show the Efficacy of Cordyceps militaris in Mild to Moderate COVID Inflammation. Cureus 2023, 15, e43731. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.-A.; Lin, T.-H.; Wang, J.-S.; Chen, J.-J.; Hsu, W.-K.; Ying, L.-C.; Liang, Z.-C. The effects of Cordyceps militaris fruiting bodies in micturition and prostate size in benign prostatic hyperplasia patients: A pilot study. Pharmacol. Res.-Mod. Chin. Med. 2022, 4, 100143. [Google Scholar] [CrossRef]
- Yin, J.; Xin, X.; Weng, Y.; Gui, Z. Transcriptome-wide analysis reveals the progress of Cordyceps militaris subculture degeneration. PLoS ONE 2017, 12, e0186279. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-J.; Deng, C.-H.; Zhang, L.-Y.; Hu, K.-H. Molecular analysis and biochemical characteristics of degenerated strains of Cordyceps militaris. Arch. Microbiol. 2017, 199, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal cordycepin biosynthesis is coupled with the production of the safeguard molecule pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Z.; Yao, X.; Huang, Y.; Qu, Q.; Shi, X.; Zhang, H.; Shi, X. Identification of cordycepin biosynthesis-related genes through de novo transcriptome assembly and analysis in Cordyceps cicadae. R. Soc. Open Sci. 2018, 5, 181247. [Google Scholar] [CrossRef]
- Kramer, G.J.; Nodwell, J.R. Chromosome level assembly and secondary metabolite potential of the parasitic fungus Cordyceps militaris. BMC Genom. 2017, 18, 912. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, Y.; Liu, L.; Feng, J.; Zhang, T.; Qin, S.; Zhao, X.; Wang, C.; Li, D.; Han, W. Study of the whole genome, methylome and transcriptome of Cordyceps militaris. Sci. Rep. 2019, 9, 898. [Google Scholar] [CrossRef]
- Massonnet, M.; Morales-Cruz, A.; Minio, A.; Figueroa-Balderas, R.; Lawrence, D.P.; Travadon, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Whole-genome resequencing and pan-transcriptome reconstruction highlight the impact of genomic structural variation on secondary metabolite gene clusters in the grapevine esca pathogen Phaeoacremonium minimum. Front. Microbiol. 2018, 9, 1784. [Google Scholar] [CrossRef]
- Brunner-Mendoza, C.; Reyes-Montes, M.d.R.; Moonjely, S.; Bidochka, M.J.; Toriello, C. A review on the genus Metarhizium as an entomopathogenic microbial biocontrol agent with emphasis on its use and utility in Mexico. Biocontrol. Sci. Technol. 2019, 29, 83–102. [Google Scholar] [CrossRef]
- Staats, C.C.; Junges, A.; Guedes, R.L.M.; Thompson, C.E.; de Morais, G.L.; Boldo, J.T.; de Almeida, L.G.P.; Andreis, F.C.; Gerber, A.L.; Sbaraini, N. Comparative genome analysis of entomopathogenic fungi reveals a complex set of secreted proteins. BMC Genom. 2014, 15, 822. [Google Scholar] [CrossRef] [PubMed]
- Qiang, G.; Kai, J.; Sheng-Hua, Y.; Yongjun, Z.; Guohua, X.; Yanfang, S.; Zhibing, D.; Xiao, H.; Xue-Qin, X.; Gang, Z.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar]
- Xiao, G.; Ying, S.-H.; Zheng, P.; Wang, Z.-L.; Zhang, S.; Xie, X.-Q.; Shang, Y.; St Leger, R.J.; Zhao, G.-P.; Wang, C. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Suparmin, A.; Kato, T.; Dohra, H.; Park, E.Y. Insight into cordycepin biosynthesis of Cordyceps militaris: Comparison between a liquid surface culture and a submerged culture through transcriptomic analysis. PLoS ONE 2017, 12, e0187052. [Google Scholar] [CrossRef] [PubMed]
- Thananusak, R.; Laoteng, K.; Raethong, N.; Koffas, M.; Vongsangnak, W. Dissecting metabolic regulation in mycelial growth and fruiting body developmental stages of Cordyceps militaris through integrative transcriptome analysis. Biotechnol. Bioprocess Eng. 2023, 28, 406–418. [Google Scholar] [CrossRef]
- Qin, P.; Wang, Z.; Lu, D.; Kang, H.; Li, G.; Guo, R.; Zhao, Y.; Han, R.; Ji, B.; Zeng, Y. Neutral lipid content in lipid droplets: Potential biomarker of cordycepin accumulation in cordycepin-producing fungi. Molecules 2019, 24, 3363. [Google Scholar] [CrossRef]
- Lv, G.; Zhu, Y.; Cheng, X.; Cao, Y.; Zeng, B.; Liu, X.; He, B. Transcriptomic Responses of Cordyceps militaris to Salt Treatment during Cordycepins Production. Front. Nutr. 2021, 8, 793795. [Google Scholar] [CrossRef]
- Raethong, N.; Laoteng, K.; Vongsangnak, W. Uncovering global metabolic response to cordycepin production in Cordyceps militaris through transcriptome and genome-scale network-driven analysis. Sci. Rep. 2018, 8, 9250. [Google Scholar] [CrossRef] [PubMed]
- Wongsa, B.; Raethong, N.; Chumnanpuen, P.; Wong-Ekkabut, J.; Laoteng, K.; Vongsangnak, W. Alternative metabolic routes in channeling xylose to cordycepin production of Cordyceps militaris identified by comparative transcriptome analysis. Genomics 2020, 112, 629–636. [Google Scholar] [CrossRef]
- Shendurse, A.; Khedkar, C. Glucose: Properties and analysis. In Encyclopedia of Food and Health; Academic Press: Oxford, UK, 2016; Volume 3, pp. 239–247. [Google Scholar]
- Lou, H.-W.; Zhao, Y.; Tang, H.-B.; Ye, Z.-W.; Wei, T.; Lin, J.-F.; Guo, L.-Q. Transcriptome analysis of Cordyceps militaris reveals genes associated with carotenoid synthesis and identification of the function of the Cmtns gene. Front. Microbiol. 2019, 10, 2105. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Yu, G.; Chen, Y.; Jiang, S.; Wang, M.; Jin, Y.; Lan, X.; Liang, Y.; Sun, H. Genome-wide transcriptome and proteome analysis on different developmental stages of Cordyceps militaris. PLoS ONE 2012, 7, e51853. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, G.; Li, C.; Ling, J. Cordycepin and pentostatin biosynthesis gene identified through transcriptome and proteomics analysis of Cordyceps kyushuensis Kob. Microbiol. Res. 2019, 218, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Huang, C.; Cao, L.; Xie, C.; Han, R. Agrobacterium tumefaciens-mediated transformation as a tool for insertional mutagenesis in medicinal fungus Cordyceps militaris. Fungal Biol. 2011, 115, 265–274. [Google Scholar] [CrossRef]
- He, R.; Zhang, L.; Lan, J.; Mei, S.; Li, Y. Cmcrf1, a Putative Zn2Cys6 Fungal Transcription Factor, Is Involved in Conidiation, Carotenoid Production, and Fruiting Body Development in Cordyceps militaris. Biology 2022, 11, 1535. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-X.; Xue, L.-N.; Wei, T.; Ye, Z.-W.; Li, X.-H.; Guo, L.-Q.; Lin, J.-F. Enhancement of ergothioneine production by discovering and regulating its metabolic pathway in Cordyceps militaris. Microb. Cell Factories 2022, 21, 169. [Google Scholar] [CrossRef]
- Choi, H.; Park, S.-W.; Oh, J.; Kim, C.S.; Sung, G.-H.; Sang, H. Efficient disruption of CmHk1 using CRISPR/Cas9 ribonucleoprotein delivery in Cordyceps militaris. FEMS Microbiol. Lett. 2023, 370, fnad072. [Google Scholar] [CrossRef]
- Zou, G.; Xiao, M.; Chai, S.; Zhu, Z.; Wang, Y.; Zhou, Z. Efficient genome editing in filamentous fungi via an improved CRISPR-Cas9 ribonucleoprotein method facilitated by chemical reagents. Microb. Biotechnol. 2021, 14, 2343–2355. [Google Scholar] [CrossRef]
- Cunningham, K.; Manson, W.; Spring, F.; Hutchinson, S. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Bao, H.; Bau, T. Isolation and identification of one natural pigment from cultured Cordyceps militaris. Mycosystema 2010, 29, 777–781. [Google Scholar]
- Chen, C.; Bao, H.; Bau, T. Chemical composition analysis of cultured Cordyceps militaris. Food Sci. 2013, 34, 36–40. [Google Scholar]
- Dong, J.Z.; Wang, S.H.; Ai, X.R.; Yao, L.; Sun, Z.W.; Lei, C.; Wang, Y.; Wang, Q. Composition and characterization of cordyxanthins from Cordyceps militaris fruit bodies. J. Funct. Foods 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Yang, T.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, Q.; Zhang, J.; Liu, K.; Li, K.; Liu, G.; Dong, C. Comparative transcriptome analysis between a spontaneous albino mutant and its sibling strain of Cordyceps militaris in response to light stress. Front. Microbiol. 2018, 9, 1237. [Google Scholar] [CrossRef]
- Lian, T.; Dong, C.; Yang, T.; Sun, J. Three types of geranylgeranyl diphosphate synthases from the medicinal caterpillar fungus, Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms. 2014, 16, 115–124. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Yang, Y.; Wang, Y.; Dong, C. CmVVD is involved in fruiting body development and carotenoid production and the transcriptional linkage among three blue-light receptors in edible fungus Cordyceps militaris. Environ. Microbiol. 2020, 22, 466–482. [Google Scholar] [CrossRef]
- Shrestha, B.; Lee, W.-H.; Han, S.-K.; Sung, J.-M. Observations on some of the mycelial growth and pigmentation characteristics of Cordyceps militaris isolates. Mycobiology 2006, 34, 83–91. [Google Scholar] [CrossRef]
- Avalos, J.; Carmen Limón, M. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef]
- Bovier, E.R.; Hammond, B.R. A randomized placebo-controlled study on the effects of lutein and zeaxanthin on visual processing speed in young healthy subjects. Arch. Biochem. Biophys. 2015, 572, 54–57. [Google Scholar] [CrossRef]
- Yang, T.; Dong, C. Photo morphogenesis and photo response of the blue-light receptor gene Cmwc-1 in different strains of Cordyceps militaris. FEMS Microbiol. Lett. 2014, 352, 190–197. [Google Scholar] [CrossRef]
- Zhang, H.; Yu-Xian, W.; Xin-Xin, T.; Yokoyama, W.; Jing, C.; Fang, W.; Cheng, P.; Jin-Lin, G. Overexpression of ribonucleotide reductase small subunit, RNRM, increases cordycepin biosynthesis in transformed Cordyceps militaris. Chin. J. Nat. Med. 2020, 18, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Takano, H. The regulatory mechanism underlying light-inducible production of carotenoids in nonphototrophic bacteria. Biosci. Biotechnol. Biochem. 2016, 80, 1264–1273. [Google Scholar] [CrossRef]
- Chen, A.; Wang, Y.; Shao, Y.; Huang, B. A novel technique for rejuvenation of degenerated caterpillar medicinal mushroom, Cordyceps militaris (Ascomycetes), a valued traditional Chinese medicine. Int. J. Med. Mushrooms. 2017, 19, 87–91. [Google Scholar] [CrossRef]
- Wang, C.; Butt, T.M.; Leger, R.J.S. Colony sectorization of Metarhizium anisopliae is a sign of ageing. Microbiology 2005, 151, 3223–3236. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.-L.; Qiu, X.-H.; Han, R.-C. Identification of the genes involved in the fruiting body production and cordycepin formation of Cordyceps militaris fungus. Mycobiology 2015, 43, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Li, M.; Kang, J.; He, J. A molecular genetic study on fruiting-body formation of Cordyceps militaris. Afr. J. Microbiol. Res. 2012, 6, 5215–5221. [Google Scholar]
- Wang, H.; Wei, J.; Lin, N.; Feng, A.; Chen, M.; Bao, D. Distribution of mating-type genes in fruiting and non-fruiting forms of Cordyceps militaris. Acta Edulis Fungi. 2010, 17, 1–4. [Google Scholar]
- Wang, H.; Cai, T.; Wei, J.; Feng, A.; Lin, N.; Bao, D.-P. Molecular markers to detect the formation of heterokaryon and homokaryon from asexual spores of the caterpillar medicinal mushroom, Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms. 2015, 17, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Zu, Z.; Wang, S.; Zhao, Y.; Fan, W.; Li, T. Integrated enzymes activity and transcriptome reveal the effect of exogenous melatonin on the strain degeneration of Cordyceps militaris. Front. Microbiol. 2023, 14, 1112035. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Yin, J.; Zhang, B.; Li, Z.; Zhao, S.; Gui, Z. Genome-wide analysis of DNA methylation in subcultured Cordyceps militaris. Arch. Microbiol. 2019, 201, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Homolka, L. Preservation of live cultures of Basidiomycetes—Recent methods. Fungal Biol. 2014, 118, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, F.; Liu, K.; Dong, C. Influence of strain preservation methods on fruiting body growth and metabolite production by the medicinal mushroom Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms. 2018, 20, 1003–1011. [Google Scholar] [CrossRef]
- Lin, Q.; Qiu, X.; Zheng, Z.; Xie, C.; Xu, Z.; Han, R. Characteristics of the degenerate strains of Cordyceps militaris. Mycosystema 2010, 29, 670–677. [Google Scholar]
- Hu, X.; Lv, A.; JN, U. Discussion of the degenerate strains of Cordyceps militaris. Edible Fungi China 2015, 34, 1–3. [Google Scholar]
- Wang, Y.-L.; Wang, Z.-X.; Liu, C.; Wang, S.-B.; Huang, B. Genome-wide analysis of DNA methylation in the sexual stage of the insect pathogenic fungus Cordyceps militaris. Fungal Biol. 2015, 119, 1246–1254. [Google Scholar] [CrossRef]
- Zhang, G.; Liang, Y. Improvement of fruiting body production in Cordyceps militaris by molecular assessment. Arch. Microbiol. 2013, 195, 579–585. [Google Scholar] [CrossRef]
Figure 1.
The biosynthesis pathway of cordycepin. Abbreviations: PTN: pentostatin; 3′-AMP: adenosine-3′-monophosphate; 2′,3′-cAMP: 2′,3′-cyclic monophosphate; 2′-C-3′-dA: 2′-carbonyl-3′-deoxyadenosine; COR: cordycepin; ADA: adenosine deaminase.
Figure 1.
The biosynthesis pathway of cordycepin. Abbreviations: PTN: pentostatin; 3′-AMP: adenosine-3′-monophosphate; 2′,3′-cAMP: 2′,3′-cyclic monophosphate; 2′-C-3′-dA: 2′-carbonyl-3′-deoxyadenosine; COR: cordycepin; ADA: adenosine deaminase.

Figure 2.
Lutein (a) and zeaxanthin (b) in C. militaris; (c) chemical structures of four cordyxanthins in C. militaris.
Figure 2.
Lutein (a) and zeaxanthin (b) in C. militaris; (c) chemical structures of four cordyxanthins in C. militaris.

Figure 3.
Putative model for light-induced carotenoid biosynthesis in C. militaris. White Colar (WC)-1/2, Drosophila-Arabidopsis-Synechocystis-human type cryptochromes (CRY-DASH), VIVID (VVD), FREQUENCY (FRQ). Photoreceptors CmWC-1/2, CmFRQ, and CRY-DASH perceive and transmit the light signal. The CmWC-1/2 complex activated the transcription of photoreceptor VVD gene cascades. Additionally, Zn2Cys6-type TFs and GPCRs functioned downstream of the CmWC-1/2 complex to regulate carotenoid biosynthesis.
Figure 3.
Putative model for light-induced carotenoid biosynthesis in C. militaris. White Colar (WC)-1/2, Drosophila-Arabidopsis-Synechocystis-human type cryptochromes (CRY-DASH), VIVID (VVD), FREQUENCY (FRQ). Photoreceptors CmWC-1/2, CmFRQ, and CRY-DASH perceive and transmit the light signal. The CmWC-1/2 complex activated the transcription of photoreceptor VVD gene cascades. Additionally, Zn2Cys6-type TFs and GPCRs functioned downstream of the CmWC-1/2 complex to regulate carotenoid biosynthesis.

Figure 4.
Manifestations and causes of C. militaris strain degradation. Strain degradation was manifested as loss of the ability to produce fruiting bodies. Several factors might lead to strain degradation of C. militaris including external and internal causes.
Figure 4.
Manifestations and causes of C. militaris strain degradation. Strain degradation was manifested as loss of the ability to produce fruiting bodies. Several factors might lead to strain degradation of C. militaris including external and internal causes.

Table 1.
Information on C. militaris genome sequencing.
| Strain | Size/Mb | Coverage/Fold | Assembly Level | GenBank Assembly Accession | Ref. |
|---|---|---|---|---|---|
| Cm01 | 32.2 | 147× | 33 scaffolds | GCA_000225605 | [38] |
| ATCC34164 | 33.6 | 180× | 7 chromosomes | GCA_008080495 | [41] |
| HN | 32.6 | 300× | 14 contigs | GCA_003332165 | [42] |
Table 2.
Comparison of C. militaris genome with other fungi.
| Strain | Size/Mb | Coverage /Fold | %G + C Content | Protein- Coding Genes | NCBI Accession | Ref. |
|---|---|---|---|---|---|---|
| C. militaris Cm01 | 32.2 | 147× | 51.4 | 9684 | AEVU00000000 | [38] |
| M. robertsii ARSEF23 | 39.0 | 100× | 51.5 | 10582 | ADNJ00000000 | [46] |
| M. acridum CQMa102 | 38.1 | 107× | 50.0 | 9849 | ADNI00000000 | [45,47] |
| M. anisopliae E6 | 38.5 | 19× | 51.0 | 10817 | AZNF00000000 | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Li, G.; Tian, X.; Shang, Y.; Yan, H.; Yao, L.; Hu, Z. Research Progress in Understanding the Molecular Biology of Cordyceps militaris. Fermentation 2024, 10, 167. https://doi.org/10.3390/fermentation10030167
AMA Style
Wang L, Li G, Tian X, Shang Y, Yan H, Yao L, Hu Z. Research Progress in Understanding the Molecular Biology of Cordyceps militaris. Fermentation. 2024; 10(3):167. https://doi.org/10.3390/fermentation10030167
Chicago/Turabian StyleWang, Lihong, Ganghua Li, Xueqin Tian, Yitong Shang, Huanhuan Yan, Lihua Yao, and Zhihong Hu. 2024. "Research Progress in Understanding the Molecular Biology of Cordyceps militaris" Fermentation 10, no. 3: 167. https://doi.org/10.3390/fermentation10030167
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.