
Tenacibaculum ovolyticum 16S rDNA Quantitative-PCR Assay Development and Field Testing
by
 , , ,
, , ,
Joseph P. Nowlan
1,2,*,
Brianna M. Heese
1,
Matthew J. Wilson
1,
Scott R. Britney
1,2,
John S. Lumsden
2 and
Spencer Russell
1 1
Center for Innovation in Fish Health, Vancouver Island University, Nanaimo, BC V9R 5S5, Canada
2
Department of Pathobiology, Ontario Veterinary College, University of Guelph, Guelph, ON N1G 2W1, Canada
*
Author to whom correspondence should be addressed.
Fishes 2022, 7(6), 303; https://doi.org/10.3390/fishes7060303
Submission received: 27 September 2022
/
Revised: 18 October 2022
/
Accepted: 20 October 2022
/
Published: 25 October 2022
(This article belongs to the Special Issue Infections of Aquatic Animals)
Abstract
:In British Columbia (BC; Canada) Atlantic salmon (Salmo salar L.) production, Tenacibaculum members are associated with ‘mouthrot’ and disease identification is based on gross observation and clinical data. Genomic similarities (i.e., putative virulence factors) between T. ovolyticum and other better-characterized agents of mouthrot could imply potential pathogenicity. While T. ovolyticum has not been directly linked to salmon mortality events in BC, it has been isolated from diseased marine fish. To investigate T. ovolyticum’s pathogenicity in situ, a T. ovolyticum 16S rDNA qPCR assay targeting a ~155 bp amplicon was developed. The assay was used to screen 67 biotic and 33 abiotic samples collected from a BC Atlantic salmon (Salmo salar L.) net-pen site before, during, and after a mouthrot outbreak. The assay was specific, quantifiable and detectable for T. ovolyticum over 6-log and 8-log units, respectively. However, cycle quotients differed between the BC isolate and type strain of T. ovolyticum, suggesting that qualitative use of the qPCR assay in field samples would be more accurate. Only two out of 100 samples were T. ovolyticum-positive, indicating limited involvement in this particular outbreak. However, the ecological role of T. ovolyticum and its involvement in the pathogenesis of other mouthrot outbreaks in Atlantic salmon is unknown.
Keywords:
Tenacibaculum; tenacibaculosis; mouthrot; bacterial stomatitis; yellow mouth; aquaculture; qPCR; TaqMan1. Introduction
Members of the bacterial genus Tenacibaculum (e.g., T. maritimum, T. dicentrarchi, and T. finnmarkense) are putative agents responsible for tenacibaculosis in mariculture fishes; a disease characterized by epidermal ulcerations, which can be accompanied by yellow plaques, and changes in fish behaviour [1,2,3,4]. Mouthrot, a regional variant of tenacibaculosis in British Columbia (BC; Canada), can induce mortality in Atlantic salmon (Salmo salar L.) post-smolts after transfer into saltwater [5,6] to a size of ~500 g [7] and presents with variably sized, yellow, oral plaques and ulcerations. Mitigating these mortalities necessitates antimicrobial treatments [5] incurring greater associated production costs and the potential for bacteria to develop antibiotic resistance [8]. In 2022, mouthrot outbreaks in BC have been treated using per os antibiotics (e.g., florfenicol, trimethoprim, and sulfadiazine) based on gross pathological findings and increased daily mortality. Since several Tenacibaculum species and isolates, including T. ovolyitucm (T.ovo), can be cultured from diseased BC Atlantic salmon, it is unclear which species or isolates are the causative agent(s) for individual mouthrot outbreaks.
Although characterization of T.ovo as a pathogen is limited, clinically, T.ovo has been found in lesions of sardine (Sardina pilchardus W.) eggs [9], American lobster (Homarus americanus H.) [10], halibut (Hippoglossus L.) fry [11], and Atlantic salmon [12]. In vivo exposure trials also identified that T.ovo could dissolve the chorion and zona radiata of halibut eggs through exoproteolytic activity when the bacteria comprised over 30% of the epiflora [13]. Phylogenetically, T.ovo is similar to other putative pathogens (i.e., T. dicentrarchi and T. finnmarkense) using 16S rDNA sequencing and multi-locus sequence analysis [11,14]. Beyond short amplicon comparisons, whole-genome sequencing identified that T.ovo encoded potential virulence factors [12,15] similar to other potentially pathogenic Tenacibaculum species (e.g., T. maritimum [16], T. dicentrarchi, and T. finnmarkense [17,18]). Overall, T.ovo’s genomic similarity to pathogenic Tenacibaculum species and correlations to disease in other fish indicates that more research is needed to understand the potential of T.ovo as a pathogen to cultured Atlantic salmon and other aquatic organisms. Thus, the primary objectives of this study were to develop a quantitative-PCR (qPCR) assay to identify and quantify T.ovo, and subsequently use the assay on samples [4] collected before, during, and after a mouthrot outbreak at an Atlantic salmon net-pen site.
2. Materials and Methods
2.1. Assay Development
2.1.1. Isolates and DNA Used
A T.ovo isolate (20-4135-2 [in-house name]) was obtained by swabbing a mouth lesion on an Atlantic salmon that exhibited mouthrot at a BC marine net-pen site in the Broughton Archipelago, and was cultured on Flexibacter maritimus media supplemented with kanamycin (50 µg·mL−1) at 12 °C. T.ovo 20-4135-2 (Genbank Accession #: OP629685) was most similar to T.ovo da5A-8 based on 16S rDNA sequencing using universal 27F (5′- AGAGTTTGATCATGGCTCAG -3′) and 1492R (5′- GGTTACCTTGTTACGACTT -3′) primers (Table 1). T.ovo 20-4135-2 was also sequenced using MinION nanopore long-read technologies [19]. A FastANI [20] comparison against the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov [accessed on 1 August 2022]) sequences of T.ovo da5A-8 (GCF_001641405.1_ASM164140v1), T.ovo EKD 002T (DSM 18103) (GCF_000430545.1_ASM43054v1), and T.ovo To-7Br (GCF_021852385.1_ASM2185238v1) were over 95% similar, passing the threshold for Tenacibaculum species-level identification [19,21,22]. Recent work on the complete genome of T.ovo 20-4135-2 identified six copies of 16S rDNA and a single nucleotide polymorphism [19]. Additional bacterial species, including T.ovo EKD 002T (DSM 18103), and Atlantic salmon DNA were also used in this study (Table 1). The DNA extraction of samples was accomplished using the Omega E.Z.N.A. Tissue extraction kit (Omega Bio-tek, Inc., Norcross, GA, USA) according to manufacturer’s guidelines and DNA was stored at −20 °C. All extractions had an A260/280 of 1.8–2.0 and an A260/230 of 2.0–2.3.
2.1.2. Alignments and Oligonucleotide Generation
Tenacibaculum spp. 16S rDNA sequences were obtained from the NCBI database before being aligned using MUSCLE in MEGAX [23]. The resulting alignment, in conjunction with the primer (MAR-4 Forward [Fw], MAR Reverse [Rv]) and probe (MAR) placement [24], were used to design the primers and probe for a T.ovo-specific qPCR assay (Table 2).
The developed primers denoted as Tenaci-G forward (Fw) and Tenaci-G reverse (Rv), are in the same genomic location as the MAR primers [24] (Figure 1). Degenerate base pairs were used for the primers to allow a 100% match to all compared Tenacibaculum species (Figure 1), and the probe denoted as ‘OVO probe’ was designed to be 100% homologous to all 16S rDNA sequences described as T.ovo on NCBI, including isolates da5A-8, NCIMB 13127, To7 Br, NBRC 15947, IFO 15947, and IAM14318 (Figure 1). The probe was also 100% similar to T.ovo EKD 002T [15]. Primers and the TaqMan hydrolysis probe were obtained from Sigma-Aldrich and Eurofins Genomics. Primers and the probe used together (Table 2) will be collectively termed the OVO assay.
2.1.3. Temperature Gradient Test and Amplicon Sequencing
A temperature gradient test (TGT) was completed using two separate PCR master-mixes. In the first, each reaction (i.e., well) had a 10 µL Light-Cycler SYBR-Green Master kit (Roche Diagnostics, Laval, QC, Canada), 1 µL of 10 µM Tenaci-G Fw, 1 µL of 10 µM Tenaci-G Rv, 7 µL of 20-4135-2 isolate DNA (14.5 ng·µL−1 [~100 ng per reaction]), and 1 µL of PCR grade water (Roche Diagnostics, Laval, QC, Canada). The second was identical, except that SYBR-Green was substituted with the Probes Master kit (Roche Diagnostics, Laval, QC, Canada), and 1 µL of 2.5 µM OVO probe replaced the water. Both TGTs used a CFX96 thermocycler (BIO-RAD, Hercules, CA, USA) with the following thermal profile: 95 °C (5 min), 40 cycles of 95 °C (30 s), variable annealing temperatures (60, 59.4, 58.3, 56.3, 53.9, 52, 50.7, and 50 °C; 30 s) and 72 °C (30 s). All temperatures for each master-mix were run in triplicate, including no-template controls, with a cut-off cycle quotient (Cq) of 35.
All qPCR products were cleaned (MinElute® Reaction Cleanup Kit, QIAGEN, Hilden, Germany) and subjected to bidirectional Sanger sequencing (University of Alberta, Molecular Biology Facility) using the Tenaci-G Fw and Rv primers. Sequences were aligned in MEGAX, and the resulting consensus sequence was used for NCBI BLAST comparison.
After the temperature gradient tests, all qPCR tests used the following, unless mentioned otherwise, and will be referred to as the optimized master-mix: each reaction (well) had a volume of 20 µL comprising 10 µL of probes master solution (Roche Diagnostics, Laval, QC, Canada); 1 µL of the OVO probe (final concentration 0.125 µM); 1 µL each of the Tenaci-G Fw and Rv primers (final concentration 0.5 µM each); 7 µL of template DNA (100 ng total, final concentration 5 ng·µL−1); 52 °C was selected as the annealing temperature; and no-template controls and positive controls were included. All samples were run in triplicate, and the cut-off Cq was set to 35.
2.1.4. Primer and Probe Optimization
The Tenaci-G Fw and Rv primers underwent optimization in factorial using final concentrations of 1, 0.75, 0.5, and 0.25 µM while the final probe (0.125 µM) and template concentration (20-4135-2, 5 ng·µL−1) remained constant. Similarly, the OVO probe was optimized at several final concentrations (0.25, 0.125, 0.05, and 0.025 µM) while the final primer (0.5 µM) and template concentration (20-4135-2, 5 ng·µL−1) remained constant. ANOVAs and Tukey HSD tests [25] compared Cq values from the primer and probe optimizations. Prior tests were used to interpret reagent efficiency.
2.1.5. Standard Curves (Sensitivity and Amplification Efficiency Testing)
Standard curves to determine the OVO assay’s sensitivity and amplification efficiency were performed using 8-log units (1000, 100, 10, 1, 0.1, 0.01, 0.001, and 0.0001 ng) of T.ovo 20-4135-2 with or without 100 ng of S. salar DNA (muscle or kidney).
Rough estimates for the limit of detection (LOD), were based on the complete genome length (~4.1–4.2 Mb) of T.ovo EKD 002T (DSM 18103) (NCBI: GCF_000430545.1_ASM43054v1), da5A-8 (NCBI: GCF_001641405.1_ASM164140v1), and 20-4135-2 (19), and the DNA Copy Number, and Dilution Calculator (https://www.thermofisher.com/ca/en/home/brands/thermo-scientific/molecular-biology/molecular-biology-learning-center/molecular-biology-resource-library/thermo-scientific-web-tools/dna-copy-number-calculator.html [accessed on 10 September 2021]) using a ‘Custom DNA Fragment’ and ‘Genome Length’ of 4.15 Mb, indicating ~223,345 copies of the genome per·ng−1.
2.1.6. Outgroup Testing and Fluorescence in Mixed Cultures
To assess the OVO assay’s specificity, outgroup testing using 100 ng of the bacterial species genomic DNA mentioned in Table 1 was conducted.
To assess the possibility of reduced Cq values from shifted fluorescence in mixed cultures of Tenacibauclum species, samples of T.ovo 20-4135-2 at 100 ng, 50 ng, and T.ovo 20-4135-2 at 50 ng mixed with 50 ng of either T. ovolyticum EKD 002T (DSM 18103), T. dicentrarchi 20-4116-9, T. finnmarkense 20-4106-2, T. maritimum R-2T (DSM 17995), T. gallaicum A37.1T (DSM 18841), or T. discolor LL04 11.1.1T (DSM 18842) isolate were prepared and run under the same aforementioned conditions. An ANOVA and Tukey HSD test [25] compared Cq values between samples.
2.2. Net-Pen Sample Screening
2.2.1. Sample Descriptions
Samples were previously collected and provided [4] (Table 3). Selected samples were from the Midsummer (MS) commercial net-pen site located in the Broughton Archipelago, BC. Samples included collections before the introduction of Atlantic salmon (collection 1 [C1]), one week after smolt entry (collection 2 [C2]), and both during and after treatments for mouthrot (collections 4 [C4] and 6 [C6], respectively). Outbreak status was previously defined [4]. Triplicates of water (0 m, 5 m, 10 m), invertebrate (Mytilus sp.), and fish tissues (euthanized and dead) were selected for each stage of a single outbreak (Table 3). Eight bacterial isolates that previously tested negative for T. maritimum and T. dicentrarchi [4] were also chosen to attempt to fill diagnostic gaps. There were 100 samples: 67 biotic samples consisting of 59 samples from the net-pen and 8 isolates; and 33 abiotic net-pen samples.
2.2.2. DNA Extractions
DNA was previously extracted [4] (Table 3) and stored at −20 °C. Frozen DNA outside acceptable parameters (i.e., A260/280 and an A260/230 of 1.8–2.0 and 2.0–2.3, respectively) or lacking frozen DNA were re-extracted from samples stored in RNALater (Invitrogen™, Waltham, MA, USA) at −20 °C using the Omega E.Z.N.A. Tissue extraction kit (Omega Bio-tek, Inc., Norcross, GA, USA) with the following modifications: DNA was eluted using 100 µL of provided elution buffer; and water samples collected on 0.22 µm filters were homogenized using a Fisherbrand™ Bead Mill 24 (Fisherbrand, Pittsburgh, PA, USA).
2.2.3. qPCR Application
All net-pen DNA samples were normalized to 14.5 ng·µL−1 if possible; in several instances, water samples were below target values and were directly used for qPCR. The optimized master-mix and thermal profile as described in ‘Assay Development: Temperature Gradient Test and Amplicon Sequencing’ was used. Detection of T.ovo was reported as the mean Cq ± standard deviation.
3. Results
3.1. Assay Development
3.1.1. Temperature Gradient Test and Amplicon Sequencing
All tested temperatures resulted with amplification and fluorescence for both the SYBR-Green and the probes master PCR mix. Melt curve analysis from the SYBR-Green TGT showed a single product with a Tm of 75.5–76 °C. An NCBI BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi [accessed on 1 September 2022]) comparison of a 127 bp section of the 16S rDNA amplicon sequence from the Tenaci-G primers indicated the closest match was T. ovolyticum da5A-8 (max and total score: 235, query cover: 100%, E-value: 6e-58, Percent Identity: 100%).
3.1.2. Primer and Probe Optimization
An ANOVA indicated that the Fw primer and Rv primer explained the variation between resultant Cq values (Fw: F3,23 = 311.4, p < 2 × 10−16; Rv: F3,23 = 8.5, p = 5.5 × 10−4); however, there was no interactive influence (Fw·Rv: F9,23 = 1.8, p < 1.3 × 10−1). An ANOVA also indicated that the probe also explained variation in Cq values (F3,7 = 23,496, p = 2.3 × 10−14). The largest concentrations for the final forward (1 µM) and reverse primer (1 µM), and the probe (2 µM) provided the lowest Cq values (Table 4). However, given other Cq values, fluorescence, reagent efficiency, and protocol simplicity, the final concentrations of 0.5, 0.5, and 0.125 µM were used for the forward and reverse primer, and probe, respectively, in further testing (Table 4).
3.1.3. Standard Curves (Sensitivity and Amplification Efficiency Testing)
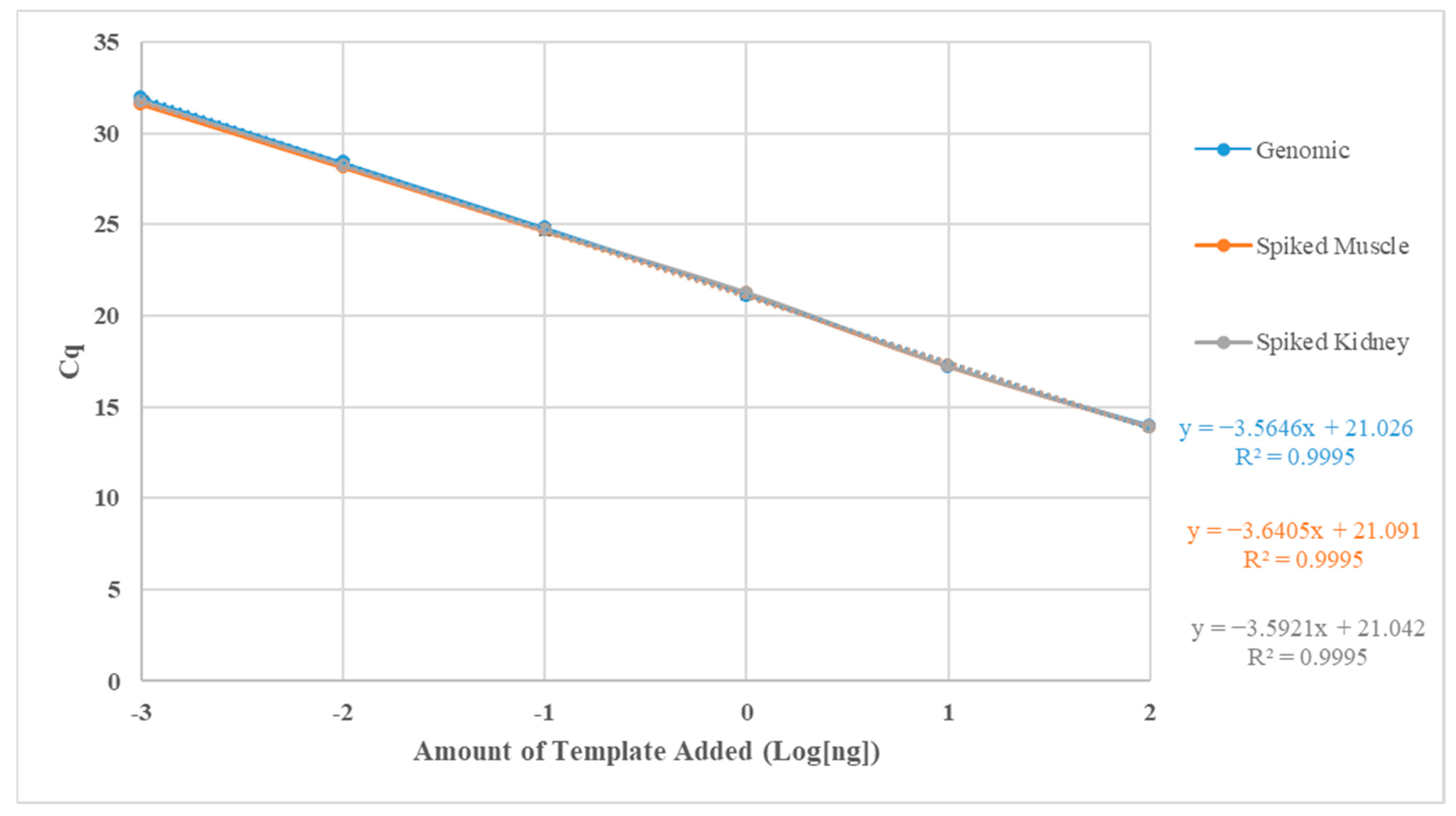
T.ovo 20-4135-2 could be identified over 8-log units; however, only 6-log units could be used for quantification (0.001–100 ng) based on deviations in the slope (Table 5). From the standard curves, using the limit of quantifiable detection (LOQD), correlation coefficients were above 0.99 and amplification efficiencies were above 88% (Figure 2, Table 5). The minimum LOD and LOQD were estimated to be 22.3 and 223 bacteria, respectively (Table 5).
3.1.4. Outgroup Testing and Fluorescence in Mixed Cultures
Outgroup testing indicated no false positives with the bacteria tested and only the positive control (T.ovo 20-4135-2 [mean Cq = 15.3 ± 0.25]) and the T.ovo EKD 002T (mean Cq = 20.40 ± 0.043) amplified.
An ANOVA on mixed Tenacibaculum cultures identified differences in Cq values (F7,13 = 16.39, p = 1.64 × 10−5). An ANOVA identified that the 100 ng T.ovo sample was different from all other tested samples (p < 4.19 × 10−2), with a Cq one less than 50 ng of the same isolate. The sample with two isolates of T.ovo (i.e., 50 ng of 20-4135 and 50 ng of EKD 002T) was marginally different than 100 ng of T.ovo 20-4135-2 (p = 4.19 × 10−2). The sample with only 50 ng of T.ovo 20-4135-2 was not significantly different from all mixed cultures (p > 0.05). The sample with two isolates of T.ovo (i.e., 20-4135 and EKD 002T) had a lower mean Cq than the 50 ng of T.ovo 20-4135-2; however, it was statistically similar (p = 6.6 × 10−1). The only recorded difference when using only 50 ng of T.ovo 20-4135-2 was when it was combined with T. dicentrarchi and T. maritimum (p = 0.04). The only differences when using two isolates of T.ovo (i.e., 20-4135 and EKD 002T), aside from what was mentioned, was between the 50 ng of T.ovo with 50 ng of T. maritimum (p = 5.23 × 10−3) isolate.
3.2. Net-Pen Sample Screening
All qPCR plates were successfully completed as identified by statistically similar Cq’s between T.ovo-positive controls. T.ovo was detected in two out of 100 samples: a water sample at 10 m from before the introduction of fish (water, mean Cq = 25.85 ± 0.083) and in a gill tissue sample taken one week after smolt entry (euthanized, mean Cq = 32.88 ± 0.044).
4. Discussion
4.1. Assay Development
The OVO assay is the first qPCR assay to identify and quantify T.ovo from both biotic and abiotic substances, and the first assay to attempt to quantify an amount of T.ovo in situ. The tested assay was sensitive (LOD and LOQD of 22.3 and 223 bacteria, respectively), specific (only positive for known T.ovo cultures), and had amplification efficiencies (88–91%) that are described in other qPCR studies [26,27,28]. Amplification efficiencies are within ranges described by standard guidelines [29,30,31]; however, with amplification efficiencies on the low end of the acceptable range, qualitative results may be more accurate.
Previous research has demonstrated that 16S rDNA sequencing can lead to misinterpreted bacterial species-level designations [18], and 16S rDNA qPCR can lead to false positives when nucleotide polymorphisms occur on the targeted section of the bacteria [28]. This indicates that other genomic targets may bear more utility. For qPCR, however, when fluorescence caused by the target region is distinct from other species and few to no polymorphisms between copies occur (as interpreted by complete genome sequencing on T.ovo 20-4135-2 [19]), it could indicate that 16S rDNA may still be situationally useful [24].
Genomic studies with incomplete de novo T.ovo sequences describe only a single copy of 16S rDNA [12,15]; however, full genome investigations with T.ovo 20-4135-2 describe six copies of 16S rDNA [19]. A potential reason for the discrepancy between copy numbers could be based on the sequencing technologies used; short-read sequencing assembly [12,15] versus long-read sequencing assembly [19]. When using short-read sequencing technologies, highly similar DNA sequences could be aggregated as one sequence [32]. However, with the Cq difference between T. ovolyticum 20-4135-2 and EKD 002T, it would support that different isolates have different copy numbers of 16S rDNA. Varying extraction efficiencies could also lead to further differences between the number of 16S copies within a sample when normalized to a set concentration. With this discrepancy between copy numbers and Cq, the qualitative use of the OVO assay would be more appropriate.
This work is currently based on two T.ovo isolates, and ideally, more isolates would be utilized as with previous work on T. dicentrarchi [22]. However, few T.ovo isolates are commercially available or have been identified. Traditional methods to identify isolates include but are not limited to Sanger sequencing and MLSA; however, both methods are time consuming and can be costly when screening multiple isolates. An economically feasible tool (i.e., qPCR) to identify T.ovo isolates may help overcome this issue and provide a larger set of isolates to test.
4.2. Net-Pen Sample Screening
The abiotic and biotic factors influencing microbial communities associated with mouthrot are multifaceted [33]. Variations in environmental conditions, such as salinity and temperature, may have the potential to influence T.ovo distributions and ecological niches. The single environmental water sample positive for T.ovo (1 out of 100) was collected before the introduction of fish (C1) at 10 m in spring/early summer when the water column had a salinity of 30 ppt and was ~8.25 °C [4]. Greater salinity concentrations have been shown to promote the growth of T.ovo (>50% seawater media [13]; 70-100% seawater media [34]) and T.ovo can grow at cooler temperatures in contrast to other Tenacibaculum species (i.e., 4–25 °C [34]). T.ovo was also isolated from deep seawater (i.e., 344 m), which grew optimally at cooler temperatures (10–20 °C) [15]. During subsequent collections (C2, C4, C6) at the net-pen site, the water was not only higher in salinity but also warmer suggesting that growth and presence could be multifaceted.
While T.ovo has been speculated to be a fish pathogen [12,13,15], the absence of T.ovo in 98 out of 100 samples tested in this study suggests that it was not directly responsible for the reported mouthrot outbreak in Atlantic salmon production in BC [4]. Another study described T.ovo with low mean abundance (<0.1%) in the oral cavity of BC produced Atlantic salmon that were deemed healthy even though affected with mouthrot [33]. Reasons for the lack of detection could be due to the number of target bacteria being below the OVO assay’s LOD, DNA degradation in samples experiencing multiple freeze–thaw cycles since their collection, and sample selection bias where target bacteria could be present in other samples collected during other mouthrot outbreaks [4]. More work is needed to interpret if T.ovo is an opportunistic pathogen or has an important role in the microbiome of marine fish, which could be accomplished using experimental infection trials with shedders and cohabitants as with T. maritimum and T. dicentrarchi [5,6].
Fish health status and life stage can also influence microbial distribution [11,33]. Mouthrot is commonly associated with smolts that have recently transferred to saltwater [5,6,33]. The only other detected presence of T.ovo in this study was in the gill tissue of a euthanized fish one week after smolt entry (C2). Stressful events, such as saltwater transfer post-smoltification, result in novel microbiomes being established and increased susceptibility to environmental pathogens. During this environmental transition, the skin and mucosal microbiota of Atlantic salmon have been known to experience increased proportions of Tenacibaculum species on healthy and diseased salmon [33,35,36]. Exposing Atlantic salmon in vivo and tracking the change in T.ovo using qPCR or high-throughput sequencing could help clarify if T.ovo is commensal to the microbiome or if it is an opportunistic pathogen.
The absence of T.ovo during and after outbreaks could be related to dysbiosis in the skin and external gill tissue of the Atlantic salmon microbiome. Similar niche requirements within the genus Tenacibaculum can lead to a sole species outcompeting others. Members of the Tenacibaculum genus can competitively influence the abundance of other species in healthy and diseased Atlantic salmon [33,35], but co-infections with multiple Tenacibaculum genotypes have also been reported [11]. In this study, T. dicentrarchi was presumed to be a contributing agent to the recorded mouthrot outbreaks [4], where an increase in the abundance of T. dicentrarchi may have reduced the abundance of T.ovo. Previous research exposing Atlantic salmon to salmonid alphavirus identified a dysbiotic event, where an increased abundance of T.ovo was recorded [37]. This increase in T.ovo could have occurred as there were no competitors within the genus that were identified that have the same niche requirements. Understanding the relationship between T.ovo and dysbiotic events (whether it is purely opportunistic or contributes to pathogenicity) is important for future research. More work is needed to understand the microbial ecology associated with T.ovo as there are still gaps in understanding its role in the environment, in hosts, and mouthrot outbreaks.
5. Conclusions
A 16S rDNA qPCR assay for T.ovo was developed for research purposes, where qualitative results appear to be more accurate than quantitative results. Upon application of the assay, two out of 100 samples were positive for T.ovo before an outbreak at a marine net-pen site; and it was concluded that T.ovo was not an agent contributing to the specific outbreak. A multispecies approach might be the next best step in understanding environmental influences and Tenacibaculum spp. community dynamics.
Future studies could include testing the non-specific Tenaci-G primers with T. maritimum [24] and T. dicentrarchi, and the T. finnmarkense [28]-specific 16S rDNA probes for a multiplex-PCR screening of BC samples similar to previous work with Tenacibaculum spp. in Australia [38]. Studies screening the remaining samples from [4] and other net-pen sites to quantify the fluctuation of T.ovo in the ecosystem during the early stages of mouthrot outbreaks would also be helpful to understand this bacteria’s role in future disease outbreaks.
Author Contributions
Conceptualization, J.P.N., B.M.H. and S.R.; methodology, J.P.N., B.M.H. and M.J.W.; software, J.P.N. and B.M.H.; validation, J.P.N. and B.M.H.; formal analysis, J.P.N. and B.M.H.; investigation, J.P.N., B.M.H., M.J.W. and S.R.; resources, S.R.; data curation, J.P.N., B.M.H. and M.J.W.; writing—original draft preparation, J.P.N., B.M.H. and M.J.W.; writing—review and editing, J.P.N., B.M.H., M.J.W., S.R.B., J.S.L. and S.R.; visualization, J.P.N., B.M.H. and S.R.; supervision, S.R. and J.P.N.; project administration, S.R. and J.S.L.; funding acquisition, J.P.N., S.R. and J.S.L. All authors have read and agreed to the published version of the manuscript.
Funding
Sources of funding include an NSERC Engage and Engage Plus grant (Grant number: 411309784) in conjunction with Mowi Canada West and a Vancouver Island University VIURAC grant (Grant number: 100863), and a Vancouver Island University Publish Grant (Grant number: 101543).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All collected data are available through contacting the corresponding author or the principle investigator.
Acknowledgments
Aquaculture company Grieg Seafood BC Ltd., and Tim Green from Vancouver Island University supplied several bacterial isolates. Mowi Canada West supported research efforts throughout the project when samples were collected.
Conflicts of Interest
All authors declare that there are no conflict of interest.
References
- Hikida, M.; Wakabayash, H.; Egusa, S.; Masumura, K. Flexibacter sp., a gliding bacterium pathogenic to some marine fishes in Japan. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 421–428. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Toranzo, A.E.; Magariños, B. Tenacibaculosis infection in marine fish caused by Tenacibaculum maritimum: A review. Dis. Aquat. Org. 2006, 71, 255–266. [Google Scholar] [CrossRef]
- Nowlan, J.P.; Lumsden, J.S.; Russell, S. Advancements in characterizing Tenacibaculum infections in Canada. Pathogens 2020, 9, 1029. [Google Scholar] [CrossRef] [PubMed]
- Nowlan, J.P.; Britney, S.R.; Lumsden, J.S.; Russell, S. Application of quantitative-PCR to monitor netpen sites in British Columbia (Canada) for Tenacibaculum species. Pathogens 2021, 10, 414. [Google Scholar] [CrossRef] [PubMed]
- Frisch, K.; Småge, S.B.; Vallestad, C.; Duesund, H.; Brevik, J.; Klevan, A.; Olsen, R.H.; Sjaatil, S.T.; Gauthier, D.; Brudeseth, B. Experimental induction of mouthrot in Atlantic salmon smolts using Tenacibaculum maritimum from Western Canada. J. Fish. Dis. 2018, 41, 1247–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowlan, J.P.; Britney, S.R.; Lumsden, J.S.; Russell, S. Experimental induction of tenacibaculosis in Atlantic salmon (Salmo salar L.) using Tenacibaculum maritimum, T. dicentrarchi, and T. finnmarkense. Pathogens 2021, 10, 1439. [Google Scholar] [CrossRef]
- Hewison, T. Yellow mouth—The Grieg Seafood experience in BC. In Tenacibaculum 2: Current Knowledge and Future Directions Workshop; Maritime Heritage Centre: Campbell River, BC, Canada, 2019; Available online: https://www.cahs-bc.ca/wp-content/uploads/2019/10/4Tim_Yellow_Mouth_The_Grieg_Seafood_BC_Experience.pdf (accessed on 24 September 2022).
- Irgang, R.; Mancilla, M.; Avendaño-Herrera, R. Florfenicol and oxytetracycline susceptibility patterns in Chilean isolates of Tenacibaculum dicentrarchi: An emerging pathogen for farmed salmonids. J. Fish Dis. 2021, 44, 1043–1046. [Google Scholar] [CrossRef]
- Míguez, B.; Combarro, M.P. Bacteria associated with sardine (Sardina pilchardus) eggs in a natural environment (Ría de Vigo, Galicia, northwestern Spain). FEMS Microbiol. Ecol. 2003, 44, 329–334. [Google Scholar] [CrossRef]
- Quinn, R.A.; Cawthorn, R.J.; Summerfield, R.L.; Smolowitz, R.; Chistoserdov, A.Y. Bacterial communities associated with lesions of two forms of shell disease in the American lobster (Homarus americanus, Milne Edwards) from Atlantic Canada. Can. J. Microbiol. 2013, 59, 380–390. [Google Scholar] [CrossRef]
- Olsen, A.B.; Gulla, S.; Steinum, T.; Colquhoun, D.J.; Nilsen, H.K.; Duchaud, E. Multilocus sequence analysis reveals extensive genetic variety within Tenacibaculum spp. associated with ulcers in sea-farmed fish in Norway. Vet. Microbiol. 2017, 205, 39–45. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Saldarriaga-Córdoba, M.; Irgang, R. Draft Genome Sequence of Tenacibaculum ovolyticum To-7Br, Recovered from a Farmed Atlantic Salmon (Salmo salar). Microbiol. Resour. Announc. 2022, 11, e00254-22. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.H.; Bergh, O.; Michaelsen, J.; Knappskog, D. Flexibacter ovolyticus sp. nov., a pathogen of eggs and larvae of Atlantic halibut, Hippoglossus hippoglossus L. Int. J. Syst. Bacteriol. 1992, 42, 451–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, C.; Houel, A.; Lunazzi, A.; Bernardet, J.F.; Olsen, A.B.; Nilsen, H.; Toranzo, A.E.; Castro, N.; Nicolas, P.; Duchaud, E. Multilocus sequence analysis of the marine bacterial genus Tenacibaculum suggests parallel evolution of fish pathogenicity and endemic colonization of aquaculture systems. Appl. Environ. Microbiol. 2014, 80, 5503–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, M.; Zhai, Z.; Komatsu, A.; Shibayama, K.; Suzuki, M. Genome sequence of the psychrophilic bacterium Tenacibaculum ovolyticum strain da5A-8 isolated from deep seawater. Genome Announc. 2016, 4, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pascual, D.; Lunazzi, A.; Magdelenat, G.; Rouy, Z.; Roulet, A.; Lopez-Roques, C.; Larocque, R.; Barbeyron, T.; Gobet, A.; Michel, G. The complete genome sequence of the fish pathogen Tenacibaculum maritimum provides insights into virulence mechanisms. Front. Microbiol. 2017, 8, 1542. [Google Scholar] [CrossRef] [Green Version]
- Grothusen, H.; Castillo, A.; Henríquez, P.; Navas, E.; Bohle, H.; Araya, C.; Bustamante, F.; Bustos, P.; Mancilla, M. First complete genome sequence of Tenacibaculum dicentrarchi, an emerging bacterial pathogen of salmonids. Genome Announc. 2016, 4, e01756-15. [Google Scholar] [CrossRef] [Green Version]
- Bridel, S.; Olsen, A.B.; Nilsen, H.; Bernardet, J.F.; Achaz, G.; Avendaño-Herrera, R.; Duchaud, E. Comparative genomics of Tenacibaculum dicentrarchi and “Tenacibaculum finnmarkense” highlights intricate evolution of fish-pathogenic species. Genome Biol. Evol. 2018, 10, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Nowlan, J.P.; Sies, A.N.; Britney, S.R.; Cameron, A.D.S.; Siah, A.; Russell, S. Genomic Investigation into Tenacibaculum species in British Columbia, Canada. Pathogens, 2022; submitted. [Google Scholar]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.B.; Spilsberg, B.; Nilsen, H.K.; Lagesen, K.; Gulla, S.; Avendaño-Herrera, R.; Irgang, R.; Duchaud, E.; Colquhoun, D.J. Tenacibaculum piscium sp. nov., isolated from skin ulcers of sea-farmed fish, and description of Tenacibaculum finnmarkense sp. nov. with subdivision into genomovars finnmarkense and ulcerans. Int. J. Syst. Evol. Microbiol. 2020, 70, 6079–6090. [Google Scholar] [CrossRef]
- Saldarriaga-Córdoba, M.; Irgang, R.; Avendaño-Herrera, R. Comparison between genome sequences of Chilean Tenacibaculum dicentrarchi isolated from red conger eel (Genypterus chilensis) and Atlantic salmon (Salmo salar) focusing on bacterial virulence determinants. J. Fish Dis. 2021, 44, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Fringuelli, E.; Savage, P.D.; Gordon, A.; Baxter, E.J.; Rodger, H.D.; Graham, D.A. Development of a quantitative real-time PCR for the detection of Tenacibaculum maritimum and its application to field samples. J. Fish Dis. 2012, 35, 579–590. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R. Version 3.6.1 (2019-07-05) “Action of the Toes”; RStudio, PBC: Boston, MA, USA. Available online: http://www.rstudio.com/ (accessed on 10 September 2021).
- Zhang, Q.; Wang, J.; Deng, F.; Yan, Z.; Xia, Y.; Wang, Z.; Ye, J.; Deng, Y.; Zhang, Z.; Qiao, M. TqPCR: A touchdown qPCR assay with significantly improved detection sensitivity and amplification efficiency of SYBR green qPCR. PLoS ONE 2015, 10, e0132666. [Google Scholar] [CrossRef]
- Rayl, J.M.; Wellehan, J.F., Jr.; Bunick, D.; Allender, M.C. Development of reverse-transcriptase quantitative PCR assays for detection of the cytokines IL-1β, TNF-α, and IL-10 in chelonians. Cytokine 2019, 119, 16–23. [Google Scholar] [CrossRef]
- Nowlan, J.P.; Lumsden, J.S.; Russell, S. Quantitative PCR for Tenacibaculum dicentrarchi and T. finnmarkense. J. Fish Dis. 2021, 44, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Wittwer, C.T. MIQE: A step toward more robust and reproducible quantitative PCR. Clin. Chem. 2017, 63, 1537–1538. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The ultimate qPCR experiment: Producing publication quality, reproducible data the first time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Oxford Nanopore Technologies (ONT). Nanopore Sequencing: The Advantages of Long Reads for Genome Assembly; pp. 1–16. 2017. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjPkviI-a_5AhXDI30KHV6WA34QFnoECAoQAw&url=https%3A%2F%2Fnanoporetech.com%2Fsites%2Fdefault%2Ffiles%2Fs3%2Fwhite-papers%2FWGS_Assembly_white_paper.pdf%3FsubmissionGuid%3D40a7546b-9e51-42e7-bde9-b5ddef3c3512&usg=AOvVaw1S_sBMns-u-WlA4qyAZoRS (accessed on 24 September 2022).
- Wynne, J.W.; Thakur, K.K.; Slinger, J.; Samsing, F.; Milligan, B.; Powell, J.F.F.; McKinnon, A.; Nekouei, O.; New, D.; Richmond, Z. Microbiome profiling reveals a microbial dysbiosis during a natural outbreak of tenacibaculosis (yellow mouth) in Atlantic salmon. Front. Microbiol. 2020, 11, 586387. [Google Scholar] [CrossRef]
- Suzuki, M.; Nakagawa, Y.; Shigeaki, H.; Yamamoto, S. Phylogenetic analysis and taxonomic study of marine Cytophaga-like bacteria: Proposal for Tenacibaculum gen. nov. with Tenacibaculum maritimum comb. nov. and Tenacibaculum ovolyticum comb. nov., and description of Tenacibaculum mesophilum sp. nov. and Tenacibaculum amylolyticum sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 1639–1652. [Google Scholar] [CrossRef]
- Karlsen, C.; Ottem, K.F.; Brevik, Ø.J.; Davey, M.; Sørum, H.; Winther Larsen, H.C. The environmental and host-associated bacterial microbiota of arctic seawater-farmed Atlantic salmon with ulcerative disorders. J. Fish Dis. 2017, 40, 1645–1663. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; Leadbeater, S.; Garcia, C.; Sylvain, F.E.; Custodio, M.; Ang, K.P.; Powell, F.; Carvalho, G.R.; Creer, S.; Elliot, J. Parasitism perturbs the mucosal microbiome of Atlantic Salmon. Sci. Rep. 2017, 7, 43465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, K.M.; Patel, S.; Robinson, A.J.; Bu, L.; Jarungsriapisit, J.; Moore, L.J.; Salinas, I. Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.) post-smolts. PLoS ONE 2017, 12, e0172856. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.K.; Douglas, M.; Dunn, V. First identification in Tasmania of fish pathogens Tenacibaculum dicentrarchi and T. soleae and multiplex PCR for these organisms and T. maritimum. Dis. Aquat. Org. 2019, 136, 219–226. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
In silico 16S rDNA alignment of several Tenacibaculum species from the NCBI website and T. ovolyticum 20-4135-2 on MEGAX using MUSCLE for the Tenaci-G forward primer, OVO probe, and Tenaci-G reverse primer. Yellow highlighted boxes represent sequences that match the oligonucleotide.
Figure 1.
In silico 16S rDNA alignment of several Tenacibaculum species from the NCBI website and T. ovolyticum 20-4135-2 on MEGAX using MUSCLE for the Tenaci-G forward primer, OVO probe, and Tenaci-G reverse primer. Yellow highlighted boxes represent sequences that match the oligonucleotide.

Figure 2.
OVO assay qPCR standard curves using T. ovolyticum (T.ovo) 20-4135-2 template DNA. All standard curves display the limit of quantifiable detection using the cycle quotient (Cq) against the log-transformed amount of template DNA added (ng). Blue is the standard curve generated using only template T.ovo DNA. Orange and gray standard curves are generated using template T.ovo DNA spiked with 100 ng of Atlantic salmon muscle or kidney DNA. Standard deviation is displayed for each point and was below 0.3.
Figure 2.
OVO assay qPCR standard curves using T. ovolyticum (T.ovo) 20-4135-2 template DNA. All standard curves display the limit of quantifiable detection using the cycle quotient (Cq) against the log-transformed amount of template DNA added (ng). Blue is the standard curve generated using only template T.ovo DNA. Orange and gray standard curves are generated using template T.ovo DNA spiked with 100 ng of Atlantic salmon muscle or kidney DNA. Standard deviation is displayed for each point and was below 0.3.

{kind=link}
{kind=link}
Table 1.
Bacterial isolates and DNA used. ‘NA’ indicates the data are not available; ‘-’ indicates a negative OVO assay qPCR reaction using 100 ng of bacterial DNA. Bacterial designation is the identifier provided by the supplier, or the most similar NCBI BLAST comparison describing the gene used, *—length of the amplicon, query cover (%), similarity (%), and E-value.
Table 1.
Bacterial isolates and DNA used. ‘NA’ indicates the data are not available; ‘-’ indicates a negative OVO assay qPCR reaction using 100 ng of bacterial DNA. Bacterial designation is the identifier provided by the supplier, or the most similar NCBI BLAST comparison describing the gene used, *—length of the amplicon, query cover (%), similarity (%), and E-value.
| In-House Sample Name | Bacterial Designation or Most Similar NCBI BLAST Comparison | NCBI BLAST Comparison | Obtained from | Grown on | Grown at °C | |||
|---|---|---|---|---|---|---|---|---|
| 16S rDNA Amplicon Length (bp) * | Query Cover, Similarity, E-Value | |||||||
| 20-4135-2 | Tenacibaculum ovolyticum da5A-8 | 1368 | 100 | 99.9 | 0 | BC Atlantic salmon | FMM+K | 12 |
| T.mar 2.1C | Tenacibaculum maritimum NLF-15 | 1366 | 99 | 100 | 0 | BC Atlantic salmon | FMM+K | 12 |
| 20-4116-9 | Tenacibaculum dicentrarchi TdChD04 | 1409 | 99 | 98.8 | 0 | BC Atlantic salmon | FMM+K | 12 |
| 20-4106-2 | Tenacibaculum finnmarkense Tsp.2 | 1426 | 100 | 99 | 0 | BC Atlantic salmon | FMM+K | 12 |
| DSM 17995 | Tenacibaculum maritimum R-2T | NA | NA | DSMZ | MA | 30 | ||
| DSM 18103 | Tenacibaculum ovolyticum EKD 002 T | NA | NA | DSMZ | MA | 15 | ||
| DSM 18841 | Tenacibaculum gallaicum A37.1T | NA | NA | DSMZ | MA | 28 | ||
| ATCC BAA-459™ | Tenacibaculum skagerrakense D30T | NA | NA | ATCC | MA | 12 | ||
| DSM 18842 | Tenacibaculum discolor LL04 11.1.1T | NA | NA | DSMZ | MA | 28 | ||
| FP | Flavobacterium sp. | 1317 | 99 | >95 | 0 | Environmental sample | CA | 12 |
| ATCC 43844™ | Polaribacter glomeratus UQM 3055T | NA | NA | ATCC | MA | 30 | ||
| ATCC 23079™ | Flexibacter flexilis CR-63T | NA | NA | ATCC | CA | 21 | ||
| F.flex Contam | Dermacoccus sp. | 589 | >91 | >88 | <1 × 10−78 | Culture contaminate | CA | 21 |
| Pcocus | Paracoccus sp. | 1184 | 100 | >96 | 0 | Culture contaminate | FMM+K | 12 |
| Beluga HI TSA 1 | Pseudomonas sp. CC11J | 1434 | 99 | 99.8 | 0 | BC White Sturgeon | TSA/CA | 12 |
| Beluga HI TSA 2 | Flavobacterium sp. T69L.09.B.RBT.MI.W. Kidney | 1386 | 99 | 99.9 | 0 | BC White Sturgeon | TSA/CA | 12 |
| LI C4 P1 | Vibrio splendidus BST398 | 1375 | 100 | 99.7 | 0 | Environmental sample | FMM+K | 12 |
| LI C3 PCB | Pseudoalteromonas sp. NBRC 107703 | 1357 | 100 | 99.7 | 0 | Environmental sample | FMM+K | 12 |
| MS7 F1 | Celluphaga sp. W5B | 1366 | 100 | 98.7 | 0 | BC Atlantic salmon | FMM+K | 12 |
| MS5 M2 | Dokdonia sp. 6a | 1358 | 100 | 99.1 | 0 | BC Atlantic salmon | FMM+K | 12 |
| MS5 F3 | Cellulophaga baltica NN015840 | 1349 | 100 | 99.8 | 0 | BC Atlantic salmon | FMM+K | 12 |
| Aero sp. kida | Aermonas sp. | NA | NA | BC Atlantic salmon | FMM+K | 12 | ||
| V.anguill | Vibrio anguillarum 155 5RH | NA | NA | BC Atlantic salmon | FMM+K | 12 | ||
| V.aest | Vibrio aestuarianus | NA | NA | Dr. Tim Green | NA | NA | ||
| Shewn.sp | Shewnanella sp. | NA | NA | Dr. Tim Green | NA | NA | ||
| P.unid | Pseudoalternomonas udina | NA | NA | Dr. Tim Green | NA | NA | ||
| E.coliTop10 | Similar to Escherichia coli DH10B™ | NA | NA | Invitrogen Topo TA Cloning Kit | LB | 37 | ||
T = Type strain; FMM+K = Flexibacter martimus media plus kanamycin (50 µg·mL−1), CA = Cytophaga agar, TSA = Tryptic soy agar, LB = Luria-Bertani media, MA = Marine agar, DSMZ = Deutsche Sammlung von Mikroorganismen und Zellkulturen, ATCC = American Type Culture Collection, BC = British Columbia (Canada), NCBI BLAST—National Center for Biotechnology Information Basic Local Alignment Search Tool. Amplicons for identification were generated using universal 16S rDNA primers (27F, 1492R).
Table 2.
Primers and probes used for the OVO assay based on 16S rDNA sequences.
| Primer or Probe Name | Sequence | Tm (°C) | Length (bp) | Amplicon Length (bp) |
|---|---|---|---|---|
| Tenaci-G Fw | TRC CTT STA CAK RRG GAT ARC C | 49.7 | 22 | ~155 |
| Tenaci-G Rv | CTA TCG THG CCA TGG TAA GCC G | 65.9 | 22 | |
| OVO Probe (FAM Fluorophore) | TGT TAA TTA GAG GCA TCT | 49.2 | 18 | NA |
Table 3.
Net-pen sample summary from the Midsummer site [4] used for qPCR analysis with number of biological replicates in parentheses. ‘C#’ refers to the collection. Water samples are described by sampling depth (m). Fish tissue samples were collected from euthanized (Euth.) and dead or moribund specimens (Dead).
Table 3.
Net-pen sample summary from the Midsummer site [4] used for qPCR analysis with number of biological replicates in parentheses. ‘C#’ refers to the collection. Water samples are described by sampling depth (m). Fish tissue samples were collected from euthanized (Euth.) and dead or moribund specimens (Dead).
| Sample Type: | C1—Pre-Fish Entry | C2—One Week Post-Fish Entry | C4—During Treatment | C6—After Treatment |
|---|---|---|---|---|
| Water | 0 m (2) | 0 m (3) | 0 m (3) | 0 m (3) |
| 5 m (2) | 5 m (3) | 5 m (3) | 5 m (3) | |
| 10 m (2) | 10 m (3) | 10 m (3) | 10 m (3) | |
| Fish Tissues (Euth.) | NA | Skin (3) | Skin (3) | Skin (3) |
| Gill (3) | Gill (3) | Gill (3) | ||
| Upper Jaw (3) | Upper Jaw (3) | Upper Jaw (3) | ||
| Kidney (3) | Kidney (3) | Kidney (3) | ||
| Fish Tissues (Dead) | NA | Skin (3) | Skin (3) | Skin (3) |
| Kidney (3) | Kidney (3) | Kidney (3) | ||
| Invertebrate | Mytilus sp. (1) | Mytilus sp. (1) | Mytilus sp. (2) | Mytilus sp. (1) |
Table 4.
T. ovolyticum primer and probe assay optimization with the cycle quotient (Cq) and Cq standard deviation (SD). Only the results from the same forward and reverse primer concentration are displayed. Significant differences (p < 0.05) between Cq are denoted by different superscript letters; different letter cases indicate separate statistical comparisons.
Table 4.
T. ovolyticum primer and probe assay optimization with the cycle quotient (Cq) and Cq standard deviation (SD). Only the results from the same forward and reverse primer concentration are displayed. Significant differences (p < 0.05) between Cq are denoted by different superscript letters; different letter cases indicate separate statistical comparisons.
| Assay | Tenaci-G Fw Primer (µM) | Tenaci-G Rv Primer (µM) | OVO Probe (µM) | Cq Mean | Cq SD |
|---|---|---|---|---|---|
| OVO | 0.25 | 0.25 | 0.125 | 16.99 a | 0.07 |
| OVO | 0.5 | 0.5 | 0.125 | 14.84 b | 0.30 |
| OVO | 0.75 | 0.75 | 0.125 | 13.97 c | 0.28 |
| OVO | 1 | 1 | 0.125 | 13.58 d | 0.06 |
| OVO | 0.5 | 0.5 | 0.025 | NA A | NA |
| OVO | 0.5 | 0.5 | 0.05 | 19.9 B | 0.11 |
| OVO | 0.5 | 0.5 | 0.125 | 15.22 C | 0.11 |
| OVO | 0.5 | 0.5 | 0.25 | 13.61 D | 0.12 |
Table 5.
Standard curves generated using T. ovolyticum 20-4135-2 DNA using the OVO assay. The limit of detection (LOD) and limit of quantifiable detection (LOQD) show the number of bacteria when a cut-off cycle of 35 is used. Correlation coefficients, slope, and amplification efficiencies are based on the LOQD.
Table 5.
Standard curves generated using T. ovolyticum 20-4135-2 DNA using the OVO assay. The limit of detection (LOD) and limit of quantifiable detection (LOQD) show the number of bacteria when a cut-off cycle of 35 is used. Correlation coefficients, slope, and amplification efficiencies are based on the LOQD.
| Standard Curve | LOD | LOQD | R2 | Slope | Amplification Efficiency (%) |
|---|---|---|---|---|---|
| Genomic | 2.23 × 101–2.23 × 108 | 2.23× 102–2.23 × 107 | 0.9995 | −3.56 | 90.78 |
| Spiked (S. salar muscle DNA) | 2.23 × 101–2.23 × 108 | 2.23 × 102–2.23 × 107 | 0.9995 | −3.64 | 88.23 |
| Spiked (S. salar head kidney DNA) | 2.23 × 101–2.23 × 108 | 2.23 × 102–2.23 × 107 | 0.9995 | −3.59 | 89.84 |
2.23 × 101 (1 × 104 ng)–2.23 × 108 (1 × 103 ng).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nowlan, J.P.; Heese, B.M.; Wilson, M.J.; Britney, S.R.; Lumsden, J.S.; Russell, S. Tenacibaculum ovolyticum 16S rDNA Quantitative-PCR Assay Development and Field Testing. Fishes 2022, 7, 303. https://doi.org/10.3390/fishes7060303
AMA Style
Nowlan JP, Heese BM, Wilson MJ, Britney SR, Lumsden JS, Russell S. Tenacibaculum ovolyticum 16S rDNA Quantitative-PCR Assay Development and Field Testing. Fishes. 2022; 7(6):303. https://doi.org/10.3390/fishes7060303
Chicago/Turabian StyleNowlan, Joseph P., Brianna M. Heese, Matthew J. Wilson, Scott R. Britney, John S. Lumsden, and Spencer Russell. 2022. "Tenacibaculum ovolyticum 16S rDNA Quantitative-PCR Assay Development and Field Testing" Fishes 7, no. 6: 303. https://doi.org/10.3390/fishes7060303