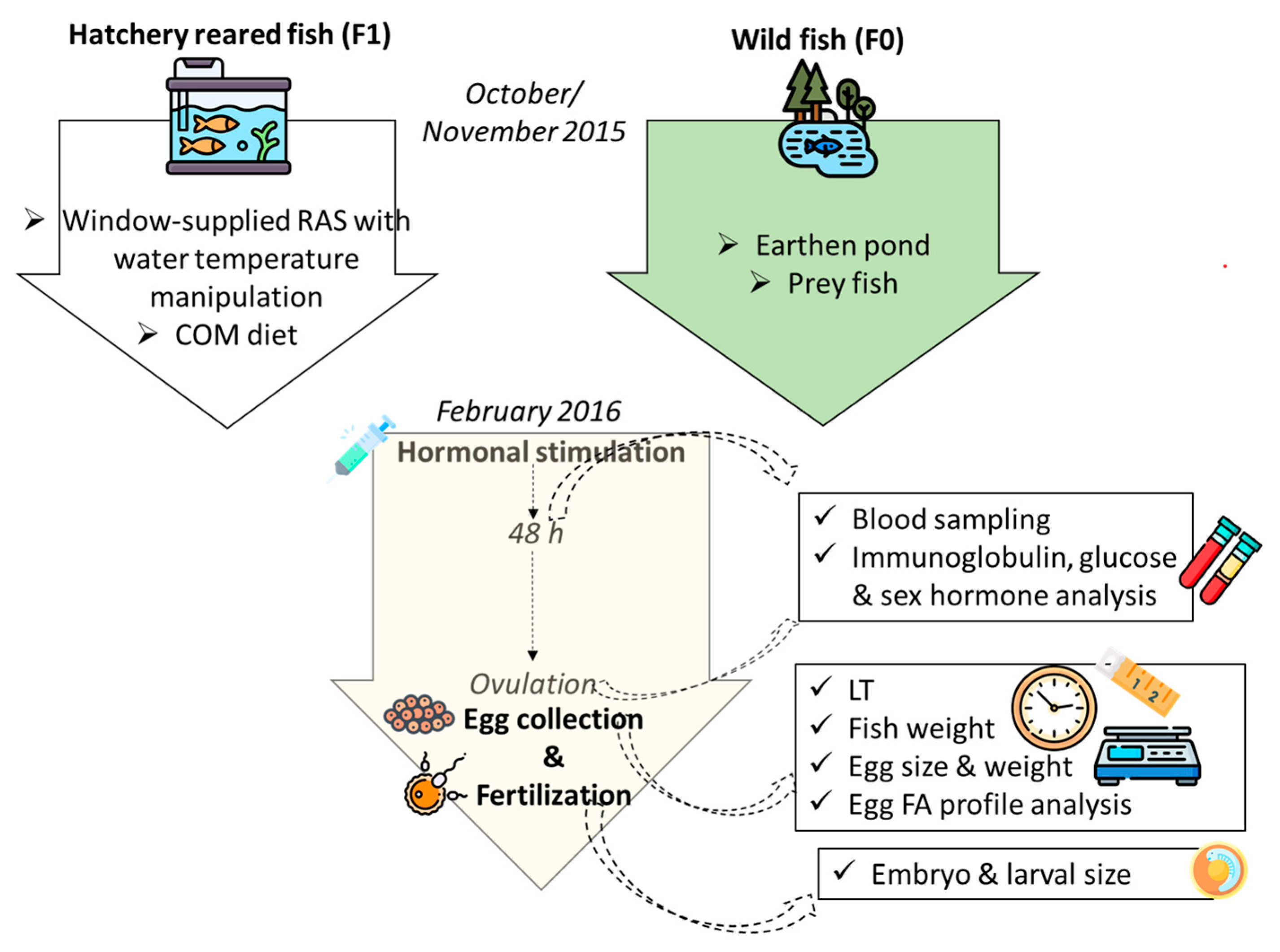
1. Introduction
Rising interest in percid cultivation across Europe has urged the need for complete control of the life cycle of these species. Due to their great adaptability to indoor farming, which is mainstream in resource-sparing fish production, percids—and particularly pike-perch (
Sander lucioperca)—have been identified as key fish species for the diversification of European aquaculture [
1]. Pike-perch has reached level four of domestication, but wild broodstock are still used as the source of reproductive material [
2]. The reasons behind the inferior reproductive performance of farmed pike-perch and other fish species have not been elucidated so far, though nutritional factors are suspected to play a critical role in oocyte growth and larval survival [
3,
4].
Oocytes’ lipid content is one of the main predictors of fish fertility. Lipids and fatty acids (FA) are the most important nutrients from the aspect of fish reproduction [
5]. Deposition of FA in finfish oocytes occurs during the previtellogenesis state when neutral lipids (NL) are deposited, and during the vitellogenesis state, when phospholipid (PL) accumulation occurs [
6]. Arachidonic acid (ARA) is the most important FA linked to gamete quality, and a deficit in this FA in cultured broodstock’s oocytes has been demonstrated in numerous fish species [
4,
7]. ARA is synthesized through desaturation and elongation reactions from linoleic acid (LA). It serves as a substrate for cyclooxygenase (COX) and 5-lipooxigenase (LOX), giving rise to prostaglandins (PG) and leukotrienes (LT), respectively. Aside from both pro- and anti-inflammatory activities, products of COX activity play a significant role in fish reproduction, though the exact mechanisms have not been elucidated yet [
8].
Fatty acids (FA) in oocytes come from dietary sources and de novo synthesis [
8]. In general, feed supplied to hatchery-reared fish throughout the on-growing and maturation periods is deficient in polyunsaturated fatty acids (PUFA), particularly ARA [
9]. Hence, deficiency in the feed may be one of the reasons for lower ARA accumulation reported in hatchery-reared broodstock’s oocytes [
7]. However, in a study performed in 2017, the reproductive potential of wild and hatchery pike-perch broodstock fed forage fish high in ARA [
10] was assessed. Although this study can have substantial implications in evaluating the role of fish physiology and/or feeding history in reproductive efficiency, it does not account for the lower feeding efficacy of F1 broodstock in comparison to F0 fish, due to different cognition and preying capacities between the two fish populations [
11,
12] Anyway, it indicated the potentially lower intrinsic capacity of F1 fish to accumulate ARA in oocytes. This finding was further confirmed when hatchery fish were fed with feed enriched with ARA at the same percentage as was present in live feed [
13].
Aside from ARA, docosahexaenoic (DHA) and eicosapentaenoic acid (EPA) have been considered to affect oocyte quality in maturing fish, which is the reason why broodstock diets are supplemented with DHA- and EPA-rich ingredients, predominantly fish oil [
14]. However, even formulated diets rich in DHA and EPA fail to provide satisfactory reproductive performance of hatchery-reared pike-perch broodstock, as shown in the previous study [
13]. Moreover, due to competition between DHA, EPA, and ARA for incorporation into membrane phospholipids and/or enzymes involved in eicosanoid synthesis [
15,
16], DHA/EPA/ARA ratios in diets may be more important than their absolute levels, both for fish growth and reproductive success [
17,
18]. Ratio 3:2:2 was found to be optimal for satisfactory egg and larval quality in Eurasian perch [
19], and diets high in marine ingredients do not necessarily meet this requirement [
13]. Hence, the application of high-cost diets may unduly increase farming costs, without any advantage to lower-cost, more sustainable diets with a lower inclusion of marine ingredients. The present study therefore aimed to assess the reproductive performance of hatchery pike-perch broodstock fed locally produced commercial feed with a significant substitution of fishmeal with vegetable protein sources, and to compare it to the performance of wild fish fed forage fish. It further aimed to estimate the potential influence of FA content in diet on the FA profile in oocytes after ovulation and to interpret the obtained results in the light of present and recent findings on fish adaptation to the farming environment.
4. Discussion
The present study corroborates the results of existing studies, demonstrating the inferior reproductive capacity of F1 pike-perch broodstock in comparison to wild breeders. However, the use of commercial feed, containing more plant products and low DHA and EPA levels (~1.1. and 0.5%), did not substantially reduce F1 fish’s reproductive performance, both in comparison to F1 fish administered a high-quality broodstock diet in the research from 2020 (~4.2 and 3.1% DHA and EPA) and breeders administered forage fish during the prespawn wintering in the research from 2017 (~6.5 and 3.9% DHA and EPA) (embryo survival of 36 ± 17% in F1 broodstock fed a high-quality broodstock diet vs. 42 ± 34% in F1 broodstock fed forage fish during wintering vs. 24 ± 23% in F1 broodstock fed a COM diet in the current study) [
10,
13]. DHA and EPA accumulation in oocytes seems to be directly affected by the levels of FAs in the diets (33.7 and 5.2% in F1 broodstock fed a high-quality broodstock diet vs. 31.7 and 5.7% in F1 broodstock fed forage fish vs. 18.0 and 1.55% in F1 fish fed the diet in the present study, in the polar lipid fraction) (
Supplementary Material) [
10]. Since F1 fish fed forage fish in the study from 2017 [
10] were not inferior in terms of DHA and EPA accumulation in comparison to F0 fish, there is possibly no physiological predisposition of F1 fish for lower DHA and EPA accumulation, nor any effects of feeding history during juvenile on-growth and maturation. There is, however, the possibility that some ingredients in formulated feed, may have contributed to the poor incorporation of DHA and EPA in oocytes’ lipids in the F1 fish, along with the lower levels of these FA in respective diets. Commercial feeds in general are rich sources of linoleic acid (LA), coming mostly from LA-rich vegetable oils incorporated in the diets [
33]. In the diet used in this research, levels of LA in the total lipid fraction were higher (~35%) in comparison to the high-quality broodstock diet (~15.6%) and forage fish (~10.4%) [
13]. LA may negatively affect DHA and EPA deposition through competition with alpha-linolenic acid (ALA) for desaturases needed for DHA and EPA synthesis [
34]. High levels of ALA, regardless of LA levels, have been demonstrated to restore the synthesis of HUFA [
35], so the replacement of LA-rich vegetable oils, e.g., sunflower oil, with ALA-rich oils, such as flaxseed oil, may potentially improve EPA and DHA deposition in hatchery-reared broodstock oocytes [
33].
Lower levels of DHA and EPA in the diet of F1 fish in this research did not appear to significantly affect ARA incorporation in oocytes’ lipids (~0.5% in F1 breeders fed a high-quality broodstock diet vs. ~2.8% in F1 breeders fed forage fish during prespawning vs. ~1.3% in F1 fish fed the diet from the current study, in the polar lipid fraction of eggs), with ARA levels in the total lipid fractions of respective diets being ~0.4%, ~3.4%, and ~0.6%. Since F1 fish were shown to be inferior in ARA deposition in comparison to F0 fish fed the same diet (forage fish), as stated above (~2.8% in F1 vs. ~5.6% in F0 fish in the polar lipid fraction), there may be a possibility that aside of the levels of ARA in the diet, some physiological factors specific to F1 fish may interfere with ARA deposition. In line with this, hCG injection, which was reported to act as both an immune and stress stimulator in fish [
36], caused a significantly higher immunoglobulin response in the plasma of F1 fish in comparison to wild breeders, implying an increased immunological response to external stimuli. This substantiates the results of previous research (Péter et al., under review), showing that higher stress and immunological response could be potential adaptive mechanisms of pike-perch to the farming environment due to their highly cannibalistic nature, though this contradicts the lower stress response generally observed in fish throughout the domestication process [
37]. Although glucose levels in F1 fish in the current research were also increased, potentially indicating a higher stress response, a firm conclusion on the difference in stress response between F0 and F1 breeders cannot be made. Hence, investigation of cortisol levels is needed to confirm this finding [
38].
Immune system activation can be linked to a higher use of ARA for the production of proinflammatory molecules, which can deploy ARA reserves. In support of this, a decline in the incorporation of ARA into membranes was observed after infection of rabbit cornea with the herpes virus [
39]. The alternative scenario is that due to intense stress and immunological reactions, F1 fish developed a protective mechanism to prevent the overproduction of proinflammatory molecules from ARA through the inhibition of ARA synthesis, as the enzymes involved in ARA synthesis have been attributed with potent inflammation-resolving capabilities [
40]. Indeed, in support of this assumption, the ARA to di-homo-gamma-linolenic acid (DGLA) ratio in eggs’ polar (PL) and neutral lipids (NL), standardized to the same ratio in food, was lower in hatchery-reared fish in comparison to wild fish. Nevertheless, additional investigation on the role of the inflammatory status of fish on oocytes’ ARA levels is needed.
Though intrinsic factors described above may potentially be involved in F1 broodstock’s inferior performance, both the diet administered during prespawn wintering and dietary history (feed provided to fish during both juvenile and maturation periods) may be important as well. In this sense, aside from the lower ARA percentage in commercial feed in comparison to live food, we identified two additional dietary factors that may have caused an ARA deficiency in hatchery broodstock’s oocytes:
1. Commercial feed is rich in oleic acid (OA), coming from plant oils or lard [
41], which can compete with ARA for binding sites in PL or can additionally inhibit the activities of desaturases and/or elongases involved in ARA synthesis [
42]. Hence, strategies to reduce OA accumulation throughout oocyte growth could potentially increase ARA levels. However, it seems that the different physiology of F1 fish may contribute to increased OA deposition as well. Eggs from F1 fish exhibited a higher OA:SA ratio (calibrated to OA:SA ratio in feed) in the PL fraction in comparison to wild fish. It is uncommon for OA to be accumulated in PL, as it rather accumulates in NL [
43]. Therefore, its high presence in PL seen in this study is probably the result of de novo synthesis in oocytes, which is believed to be the major means of deposition of FA in fish oocytes’ PL fraction [
6]. Interestingly, the synthesis of OA from SA may be aggravated by lower levels of DHA and EPA in the oocytes of F1 fish fed formulated feed during prespawn wintering, as reported by Bellenger et al., 2004 [
44].
2. LA, which is present in copious amounts in commercial feed, may be an additional reason for poor ARA incorporation in fish oocytes. LA is the precursor of ARA, and it can indirectly reduce ARA levels through stimulation of OA synthesis, which may be aggravated in the state of DHA and EPA deficiency present in F1 fish fed commercial feed, as stated above [
35]. Aside from indirectly reducing ARA accumulation, LA was reported to directly compete with ARA for acylation and incorporation in membrane PL [
45,
46]. Hence, the occupation of PL binding sites by LA may inhibit the accumulation of ARA, even after the replacement of LA-rich commercial feed with live food before spawning, as was the case in the study from 2017 [
10]. Since—as stated in the above text—DHA and EPA may suppress the conversion of SA to OA, and since ALA may restore the synthesis of DHA and EPA, the replacement of LA-rich plant sources with ALA-rich oils in formulated aqua feeds may be one of the solutions to potential interference of LA with ARA deposition.
OA accumulation in F1 fish, as noted in present study, may potentially lead to adverse effects of ARA, especially when externally supplied in larger amounts. OA has been described as an efficient competitive substrate for cyclooxygenase (COX), which is involved in the production of prostaglandins (PG), attributed with numerous pro- and anti-inflammatory activities, as well as hormone-like effects [
47]. This is because the occupation of COX by OA may leave more ARA substrate for 5-lipooxigenase (LOX), which produces leukotrienes (LT), predominantly proinflammatory molecules, commonly associated with various pathological processes [
48,
49]. This might have caused the observed detrimental effects of ARA supplementation in previous research, when hatchery fish administered commercial feed enriched with ARA had poorer performance in comparison to fish without ARA enrichment [
13]. This effect might have additionally been amplified by low levels of EPA and DHA, which are the sources of anti-inflammatory resolvins, in the diet of F1 fish [
50].
Lower pseudogonadosomatic index (PGSI) and embryo survival rate in F1 fish were, surprisingly, associated with higher estrogen (E2) levels after human chorionic gonadotropin (hCG) injection. Though levels of E2 were not evaluated throughout the reproductive season, previous studies have reported lower estrogen and estrogen-releasing hormone levels in farmed fish [
4,
51]. Given the importance of E2 in vitellogenesis, gonad development, and FA deposition in oocytes, it can be assumed that the rise in E2 levels observed in F1 fish after hCG injection does not reflect the real hormonal status of F1 fish [
8]. So, the difference in E2 secretion between the groups may rather be the consequence of slight desensitization of hCG receptors in wild broodstock, which otherwise has higher endogenous gonadotropins levels in a nonstimulated state, as mentioned above [
52]. However, the observed elevation of E2 after hCG injection was probably too late to positively regulate the PGSI and other major FAs such as ARA, EPA, or DHA.
We note here that, although we failed to link DHA and EPA levels in the diets with the reproductive performance of the fish, there is a possibility that the DHA:EPA:ARA ratio, which was close to optimal in the COM (~2:1:1) and forage fish (~2:1:1) diets but far from the ratio present in the high-quality broodstock diet (~11:8:1), may have counterbalanced potential detrimental effects of low DHA and EPA availabilities from COM feed [
16]. Anyway, additional research is needed to clarify the roles of dietary DHA, EPA, and DHA: EPA:ARA ratios in the reproductive performance of pike-perch.
An illustration of the above-provided assumptions on the effects of FA composition of the diets, as well as intrinsic factors, on ARA levels in eggs is given in
Figure 3.
The present study emphasized the role of FAs in pike-perch reproduction efficacy. It did not evaluate the potential effects of other nutrients that may affect the reproduction, which are mostly vitamins, including vitamin A, C, and E [
53]. The COM diet was optimized to meet the requirement of predatory fish in intensive rearing conditions for micronutrients, including vitamins, minerals, and amino acids. The forage fish utilized may similarly contain the required microelement, but this unfortunately was not determined from our samples. According to the data provided by the COM feed manufacturer, the contents of vitamin A, C, and E per 100 g of feed are 475 μg retinol equivalents (RE), 10 mg, and 7.7 mg per dry weight, respectively (
Table 1). Forage fish commonly preyed by pike-perch in the oxbow from which the breeders were harvested include common bleak (
Alburnus alburnus), roach (
Rutilus rutilus), rudd (
Scardinius erythrophthalmus), and bream (
Abramis brama). Though data on the nutritive values of the above fish are not available in the literature, according to the studies that evaluated the nutritive values of various fish species, mostly in fillets, the approximate levels of vitamin A and E per 100 g of wet weight are 50 μg and 2 mg, respectively, while vitamin C is present in trace amounts [
54,
55,
56]. Given the conversion factor of four to account for the difference in dry matter content between the COM and forage fish diets (
Table 1), the approximate levels of the above vitamins per 100 g of dry weight of forage fish are expected to be 200 μg and 8 mg, which is lower than or close to the levels present in the COM diet. Nevertheless, a separate study to evaluate the effects of proteins and vitamins should be set up, particularly because the protein content is higher in forage fish than in COM diet (~52% vs. ~41% per dry weight).
A major drawback of the present research is the failure to address the role of physiology in fish reproductive performance, though putative roles of different immunological and stress responses in F0 and F1 fish have been assumed. There are two reasons why clear differentiation between the roles of diet and physiology cannot be made: 1—the impossibility to eliminate dietary history, which is different for wild and F1 fish and which can significantly impact the deposition of FA in oocytes, particularly during the maturation period; and 2—the impossibility to equalize nutrient intake between the two fish populations during the prespawn wintering because of the potentially lower preying capacity of farmed fish, as stated above, and indigestibility issues related to use of dry diets by large wild fish. To the best of our knowledge and experience, the oldest wild fish to be habituated on a dry diet is a summer-old juvenile, and even so, with largely reduced success compared to the month-old pond-nursed fry. This goes along with different rearing conditions (indoor vs. pond) in two fish populations, since different systems are more suitable for different feeds (RAS for dry feed and pond for live food). Setting up the study with a two-way interaction design, where the major and simple main effects of diets and fish physiology are evaluated, would more closely reveal the effects of the two factors in fish reproductive performance.


 ,
, 




{kind=link}
{kind=link}
{kind=link}