Abstract
This study investigated the speciation, transformation, and availability of P during indigenous vegetable production by employing a combination of chemical and spectroscopic techniques. The study focused on sites in two ecozones of SSA, the dry savanna (lna, Republic of Benin) and rainforest (Ilesha, Nigeria). Both sites were cultivated with two indigenous vegetable species: local amaranth (Amaranthus cruentus (AC)) and African eggplant (Solanum macrocarpon (SM)). The soils were treated with 5 t/ha poultry manure and urea fertilizer at the rates of 0, 20, 40, 60, and 80 kg N/ha. Soil samples were collected before planting and after harvest. Phosphorus K-edge X-ray absorption near-edge structure (XANES) spectroscopy was used to determine P speciation in these soils. Quantitative analysis showed that adsorbed and organic P were the two dominant P species in the manure amended dry savanna (DS) soils before planting and after harvest in soils cultivated with both AC and SM, with the addition of urea (40 kg N/ha) causing an increase in the organic P form in dry savanna soils cultivated with AC. Soils of the rainforest (RF) cultivated with AC initially had large amounts of apatite P in the manure amended soils prior to planting, which was transformed to adsorbed and organic P after harvest. Urea addition to the rainforest soils shifted the dominant P species from organic P to adsorbed and apatite P, which was likely to limit P availability. Soils cultivated with SM had similar proportions of both organic and adsorbed P forms, with 40 kg N/ha addition slightly increasing the proportion of adsorbed P.
1. Introduction
Soils of Sub-Saharan Africa (SSA) are generally highly weathered, sandy textured, and characterized by poor fertility [1] and low phosphorus (P) and nitrogen (N) levels [2]. These soil characteristics pose limits on agricultural productivity, which in turn leads to food, household, and economic insecurity for West Africa [3,4,5]. With a rapid population growth rate of 3 to 5% [3], there is a need to intensify agricultural production sustainably to meet the food needs of this region. Accordingly, Integrated Soil Fertility Management (ISFM) is a package of agronomic recommendations designed to improve both soil fertility and agricultural yields. The ISFM incorporates the use of inorganic fertilizers and other organic amendments with improved crop varieties and crop rotations [6,7]. One of the ISFM fertilization techniques is fertilizer microdosing; the precise application of relatively small quantities of fertilizer to plants near their planting holes, which reduces the volume of fertilizers farmers need to apply due to improved nutrient uptake efficiency. This technique has proven to be economical and efficient in a wide variety of agricultural systems [2,8]. Indigenous leaf vegetables such as Amaranthus cruentus and Solanum macrocarpon are an integral part of agriculture in SSA; they contribute to human nutrition and traditional medicine and play a key role in the rural economy and family subsistence [9,10,11,12]. This MicroVeg project is incorporating the fertilizer microdosing technique to increase indigenous vegetable production in Nigeria and the Republic of Benin. Fertilizer microdosing has proven to increase yields in cereals across SSA, but has been much less frequently utilized for vegetable crops.
The practice of microdosing is not without negatives. Since farmers are applying reduced rates of inorganic fertilizer and experiencing large yield increases, there is a concern that repeated use of fertilizer microdosing may mine nutrients from the soil, reduce soil pH, and accelerate land degradation [13]. Specifically, in systems where microdosing uses urea instead of NPK fertilizer, nutrient mining of P may limit yields for microdosing. Phosphorus is essential for plant growth and development [14]. However, P is often strongly retained in the soil by chemical mechanisms such as adsorption to mineral surfaces, especially Al and Fe oxides [15,16] and precipitation as phosphate minerals with Al and Fe in acidic soils or with Ca and Mg in circumneutral soils [17]. As a result of these fixation reactions, plant available P in the inorganic orthophosphate form (HPO42− or H2PO4−) is sparingly found in soil solution, and instead, dissolved P is largely in the organic form [18], which can limit availability.
The application of manure to agricultural soils is beneficial in many ways: it provides essential macronutrients such as N and P [19,20,21,22], improves soil organic matter content [23], and helps to maintain soil quality [24]. However, manures can also alter the physical and chemical properties of the soil [25], with P minerals in the soil transforming into new phases even after short term manure application [26], and pose an environmental threat to surface waters if not properly managed. The bioavailability of P in manure and manure amended soils varies with the type of manure and soil, the solubility of the P compounds in them, and the reaction products formed upon their application [20,27,28]. Therefore, understanding how the applied manure and nitrogen fertilizer influences the forms and plant availability of P is important in minimizing its negative effects on the environment [29] in any agricultural system.
X-ray absorption near-edge structure (XANES) spectroscopy has been used by many researchers in studying the speciation of P in soils [26,30,31,32,33,34,35,36,37]. This direct speciation technique is preferred because it is element specific and non-destructive [38] and it avoids bias in distinguishing different P species that can arise from chemical fractionation methods [39,40]. Typically, P species in soil samples can be identified and quantified using linear combination fitting (LCF), which requires a robust spectral library of P reference compounds [41,42]. The spectral features of different phosphate standard materials have been reported by several researchers [30,31,34,41,43,44,45] and can be used to determine overall P speciation in soils amended with organic or inorganic P fertilizer.
The objective of this study is to identify P species present in the soil before and after the cultivation of two indigenous vegetables (Amaranthus cruentus and Solanum macrocarpon) with the MicroVeg agronomic practices in two ecozones (dry savanna and rainforest) of West Africa. This study utilizes a combination of both the direct solid state speciation via synchrotron X-ray absorption spectroscopy (XAS) and indirect speciation via chemical analysis to determine the linkages between availability and mobility of P and its speciation in soils.
2. Materials and Methods
The study sites were located in (a) Ina, Republic of Benin (9° 95 N, 2°72 E), which has an elevation of 380 m, an annual precipitation of 1073 mm, and a mean temperature of 26.5 °C, and (b) Ilesha, Nigeria (07°29′21.9″ N 004°34′17.3″ E), which has an altitude of 372 m, an annual precipitation of 1386 mm, and a mean temperature of 25.5 °C. Ina is in the dry savanna ecoregion, whereas Ilesha falls within the rainforest ecoregion. Plots from both sites were cultivated with two indigenous vegetable species shown in Figure 1: local amaranth (Amaranthus cruentus) and African eggplant (Solanum macrocarpon).

Figure 1.
Photo of Amaranthus cruentus and Solanum macrocarpon growing in (a) Ilesha and (b) Ina.
The experimental plots consisted of 1 control and 4 treatments: manure only (T0), as the control, manure + 20 kg/ha urea (T20), manure + 40 kg/ha urea (T40), manure + 60 kg/ha urea (T60), and no manure + 80 kg/ha urea (T80) using a randomized complete block design and replicated four times. Manure was applied at 5 t/ha, and it contained 20% C, 2.5% N, and 1.5% P by weight. Soil samples were randomly collected from 9 to 10 points on each plot before planting (BP) and after final harvest (AH, i.e., after 3 cycles of harvesting the vegetable leaves) at a 0–20 cm depth using a soil auger. A sub-sample of a mixed composite sample was air dried, ground to pass through a 2 mm sieve, packaged in airtight and sealed bags, and shipped to the University of Saskatchewan in April 2017. The soils were stored in airtight vials at room temperature and analyzed for pH, total P, available P, estimated cation exchange capacity (eCEC), and P XANES analysis. Due to the limited volume of soil samples shipped, bulk density could not be measured. Soil pH was measured in triplicate in a 2:1 water:soil suspension (Carter and Gregorich, 2008), and particle size analysis was done using the hydrometer method following dispersion with sodium hexametaphosphate [46]. Organic C was determined by combustion using a Leco C/N analyzer (Leco Corp., St. Joseph, MI, USA), and total P was measured colorimetrically with an auto-analyzer (Folio AA3) in triplicate following digestion using the H2SO4—H2O2 acid block digestion method of Thomas et al. (1967). Available P and eCEC were determined using the Mehlich-III extraction procedure, as described by Carter and Gregorich (2008). This method can extract multiple elements of interest to this study and is known to be appropriate for tropical soils [46]. Available P was determined from the Mehlich-III extracted soil solution on an auto-analyzer, and CEC was calculated by measuring the concentrations of exchangeable Ca2+, Mg2+, Na+, and K+ cations in vacuum filtered Mehlich-III solutions on a Microwave Plasma Atomic Absorption Spectrometer (MP-AES 4100, Agilent Technologies, Santa Clara, CA, USA). Estimated CEC was then calculated as the sum of exchangeable base cations. All chemical analyses were carried out in triplicate.
2.1. Phosphorus K-Edge XAS
Prior to XANES analysis, all triplicate soil samples of each treatment were mixed into one composite sample, finely ground, homogenized, sub-sampled, and spread as a thin film (area of 2 cm2 ± 0.5 cm) on graphite tape and then carefully mounted on a sample holder. Phosphorus K-edge XANES data were collected at the Soft X-ray Micro-characterization Beamline (SXRMB) of the Canadian Light Source (CLS), Saskatoon, Canada. Measurements were performed with an InSb (III) double crystal monochromator with an energy resolution of 0.5 eV in partial fluorescence mode (PFY) using a 4 element solid-state detector (Bruker Optics, Billerica, MA, USA) optimized for P emission. Two to three scans were taken for each sample and merged into one spectrum. The beamline energy was calibrated after every beam fill using a ZnPO4 reference compound. The acquisition time for the spectrum varied between 2 s and 5 s depending on the total P concentration in the sample. All spectra were baseline corrected with a linear pre-edge (energy region: 2115.5–2140.5 eV) and edge normalized (energy region: 2180.5–2200.5 eV) to a step height of 1.0 using the software package ATHENA (Ravel & Newville, 2005).
Linear combination fitting (LCF) as used by Peak (2002), Prietzel et al. (2013), Kar et al. (2017), and Hamilton et al. (2018) was used to fit the spectra of the unknown composition from the soil samples against the spectra of carefully selected reference materials compiled in previous studies by Hamilton et al., 2017 and 2018, and Kar et al., 2012 and 2017. The LCF approach assumes that the spectrum from a heterogeneous soil sample is an average of all the P species in the soil. Our previous studies suggested that a viable P speciation strategy includes one organic P phase (phytic acid was used as a representative compound), one adsorbed phosphate reference, a poorly crystalline calcium phosphate (brushite), a highly crystalline calcium phosphate (apatite), and an Al and Fe phosphate mineral. These phases were all included as potential components in the LCF of each sample, but a maximum of four standards per sample was allowed in the fits. Fits were accepted when the sum of the percentages of included standards were in the range of 90% to 110%, but then, the relative contribution was normalized to 100% for comparison. Goodness of fit values (as reduced chi2) in Athena were typically 0.002–0.005, suggesting a statistically significant fit.
Two samples that had very different bulk XANES speciation were also imaged using the tender X-ray microprobe of the SXRMB beamline using a 4 element Vortex detector, a 10 × 10 µm incident beam spot size with an overall 0.5 × 0.5 mm dimension, and a constant incident energy of 7200 eV to capture the regions of interest between Si and Fe. Soil samples were loaded as a dried, ground, thin layer on carbon tape. X-ray fluorescence map data analysis was processed using the SMAK software package (Sam Webb, Stanford Synchrotron Radiation Lightsource, Menlo Park, CA, USA) to subtract the inline beamline I0 and generate tricolour images to visualize co-locations of elements of interest in the samples.
2.2. Statistical Analysis
The physico-chemical analysis data were analyzed using the Statistical Package of Social Sciences (SPSS Inc.) at p ≤ 0.05 level. The significance of treatments and time effects, as well as the interaction of treatments and time effects was analyzed using repeated measure analysis of variance and determined significant at p ≤ 0.05 probability levels.
3. Results
3.1. Yields
The RF had higher vegetable yields than the DS for both vegetable species (Figure 2a,b). For the AC, the T40 had the highest yield in the RF, while the T60 gave the highest yield in the DS (Figure 2a). For the SM, the T60 gave the highest yield in both ecoregions (Figure 2b). However, for economic profitability, the T40 was considered the recommended rate.
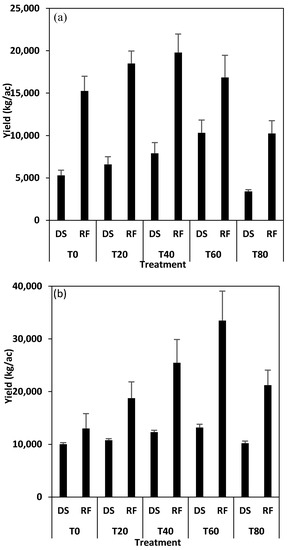
Figure 2.
Average treatment yields (kilogram per acre) of (a) Amaranthus cruentus and (b) Solanum macrocarpon in the dry savanna (DS) and rainforest (RF) sites. T0 = 0 kg urea + manure treatment, T20 = 20 kg N + manure, T40 = 40 kg N + manure, T60 = 60 kg N +manure, T80 = 80 kg N + manure.
3.2. Soil Physico-Chemical Analysis
Particle size analysis results showed that soils in both ecoregions were coarse textured, having sand content as high as 81% in Ina and 85% in Ilesha. However, the rainforest (RF) soils had more fine texture particles than those in dry savanna (DS), with clay contents as high as 33% in some samples (Table 1 and Table 2). The soils in the DS were slightly basic to neutral in pH (6.1 to 7.0) (Table 1), while the RF soils ranged from acidic, 3.5, to slightly basic, 6.7 (Table 2). Generally, for soils cultivated with Amaranthus cruentus (AC), the pH reduced over time in both the DS and RF ecoregions (Table 1 and Table 2). For the soils cultivated with Solanum macrocarpon (SM), the pH significantly increased after harvest in all treatments at the DS site (Table 1, Figure 3a), while in the RF, the pH significantly decreased over time except for T80, i.e., the samples with heavy urea addition (Table 2, Figure 4b).

Table 1.
Physico-chemical properties of the Ina (dry savanna) soils cultivated with Amaranthus cruentus and Solanum macrocarpon (n = 4). eCEC, estimated cation exchange capacity; BP, before planting; AH, after harvest.
Table 2.
Physico-chemical properties of the Ilesha (rainforest) soils cultivated with Amaranthus cruentus and Solanum macrocarpon (n = 4).
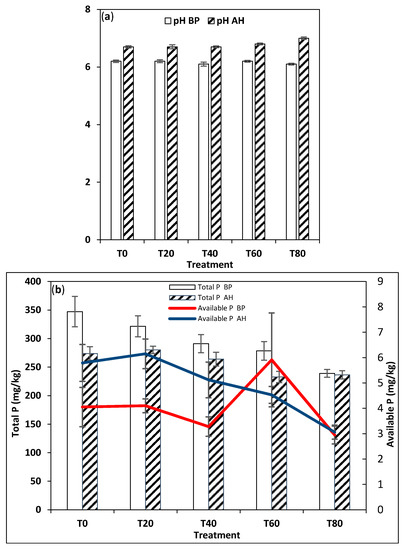
Figure 3.
Effects of fertilizer treatments on (a) pH and (b) total P and available P measured before planting (BP) and after harvest (AH) of Solanum macrocarpon cultivated on dry savanna soils.
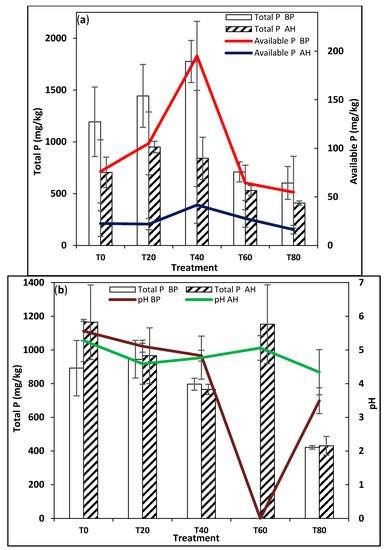
Figure 4.
Time and treatment effects on (a) total P and available P of rainforest soils cultivated with Amaranthus cruentus and (b) on pH and total P of rainforest soils cultivated with Solanum macrocarpon.
The DS soils had a significantly lower total P than the RF soils. The maximum total P measured in the DS was 347.25 mg kg−1 (Table 1), versus 1775.50 mg kg−1 in the RF (Table 2). At both sites, the available P was a small fraction of the total P, with only 3.3% and 11% of the total P being available in the DS and RF sites, respectively (Table 1 and Table 2). In the soils cultivated with AC, total and available P reduced after harvest compared to before planting at both sites (Figure 4a and Figure 5b), while in the soils cultivated with SM, total P reduced after harvest in all treatments as available P increased at the DS site (Figure 3b, Table 1). However, in the RF, for soils cultivated with SM, total P increased in all treatments except T40 (Table 2, Figure 4a), albeit within the margin of error, while available P had a different trend, increasing in all treatments except T20 at the RF site for soils cultivated with SM (Table 2, Figure 4a).
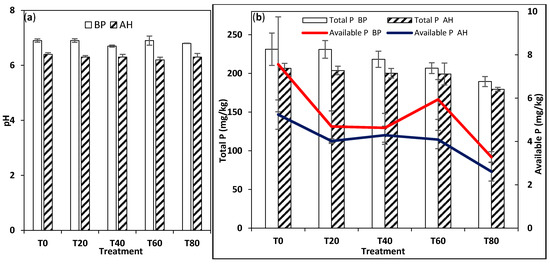
Figure 5.
Effects of fertilizer treatments on (a) pH and (b) total P and available P measured before planting (BP) and after harvest (AH) of Amaranthus cruentus cultivated on dry savanna soils.
In the DS, eCEC reduced with time in all treatments except for T80 in the soils cultivated with AC, while the eCEC increased with time for all treatments in the soils cultivated with SM (Table 1). In the RF, the eCEC reduced with time in all treatments in soils cultivated with AC (Table 2), and in the soils cultivated with SM, eCEC increased in T0 and T40 and reduced in T20 and T80 (Table 2). The trend observed in the eCEC of soils of both ecoregions cultivated with AC was almost similar to the trend observed in the pH, where both pH and eCEC reduced over time in treatments of both the DS (except for T80) and RF (except for T0) ecoregions for soils cultivated with AC (Table 1 and Table 2).
3.3. P XANES Analysis
For this study, we chose to compare the control soils, T0, with the T40 treatments as this represented the MicroVeg fertilizer recommendation in both ecoregions. The XANES spectra for all the DS soils were almost identical in shape and energy position. Visual inspection of the sample spectra presented in Figure 6a shows features that are characteristic of organic and adsorbed phosphates such as a post-white line peak at 2171.3 eV (Zuo et al., 2015). The LCF results of the DS in the AC control, AT0, showed that adsorbed P (64.9%) and organic P (35.1%) were the dominant P species (Table 3). Upon harvest however, both P forms slightly reduced by 1% and 2%, respectively, apatite accounting for the remaining 2.9% of P present in the soil. In the soils amended with 40 kg N/ha of urea (AT40), the P forms present prior to planting were adsorbed P (67.9%) and organic P (32.1%); after harvest, however, the adsorbed P was reduced by 27.2%, while the organic P increased by 26.1% (Table 3, Figure 6b).
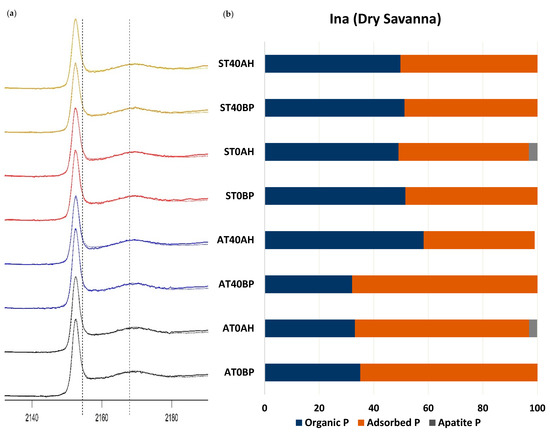
Figure 6.
(a) Normalized P K-edge X-ray absorption near-edge structure (XANES) spectra showing the linear combination fitting (LCF) results of dry savanna (Ina) soils cultivated with Amaranthus cruentus (A) and Solanum macrocarpon (S) amended with poultry manure alone (T0) and that amended with poultry manure and 40 kg N/ha of urea (T40). BP means before planting, and AH means after harvest. (b) Proportion of P compounds found in the LCF of soils in (a).
Table 3.
Proportion of P species in the amended soils from linear combination fitting (standard error of the fit in parentheses).
In the DS soils cultivated with SM, the LCF showed an almost equal distribution of organic P (51.6%) and adsorbed P (48.4%) (Figure 6a,b) as the P forms present prior to planting in ST0. After harvest, the adsorbed P and organic P reduced slightly to 47.8% and 52.7%, respectively, and there was the occurrence of some apatite P (3.1%) (Figure 6a,b). In the ST40 treatment, adsorbed and organic P had an even distribution as the dominant P form, accounting for 49% and 51%, respectively, of P forms in the soil before planting and having no significant change after harvest.
In the RF, visual inspection of the sample spectra presented in Figure 7a,b showed features that were characteristic of Ca phosphates such as apatite, a post-edge shoulder near 2157.5 eV, and a post-white line peak at 2164.7 [47], as prominent in the soils cultivated with AC. LCF results showed apatite and adsorbed P to be the P forms present in AT0 before planting, with both P forms accounting for 81% and 18.9% of P forms, respectively (Table 3, Figure 6a,b). Upon harvest, there was a major change in the P forms found in the soil: organic and adsorbed P were the dominant P forms found after harvest, accounting for 62.9% and 21.5% of P forms, respectively, while apatite accounted for 14.7% of the P forms present (Table 3, Figure 7a,b). In AT40, prior to planting, organic P accounted for 47.7% of the P present, while adsorbed and apatite P accounted for 42.1% and 8.1% of the P forms present, respectively (Table 3, Figure 7a,b). After harvest, there were notable changes in the P forms present, as apatite P more than tripled and accounted for 32.9% of P forms, adsorbed P decreased to 27.5%, while organic P saw a reduction to 37.5% of the P forms present in the soil (Figure 7a,b).
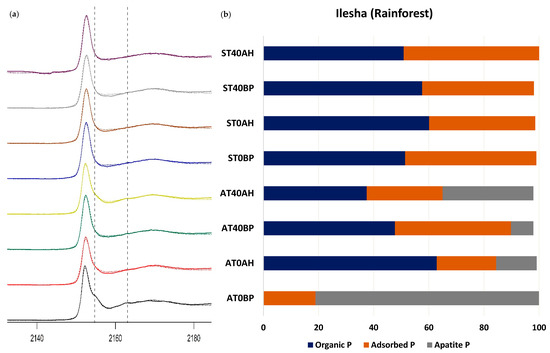
Figure 7.
(a) Normalized P K-edge X-ray absorption near-edge structure (XANES) spectra showing the linear combination fitting (LCF) results of rainforest (Ilesha) soils cultivated with Amaranthus cruentus (A) and Solanum macrocarpon (S) amended with poultry manure alone (T0) and that amended with poultry manure and 40 kg N/ha of urea (T40). BP means before planting, and AH means after harvest. (b) Proportion of P compounds found in the LCF of soils in (a).
Furthermore, in the rainforest soils cultivated with SM, the LCF showed adsorbed P and organic P as the dominant P forms in ST0 before planting, at 51.4% and 47.6%, respectively (Table 3, Figure 7a,b). After harvest (ST0AH), there were changes in the P speciation as organic P increased to 60.1%, while adsorbed P decreased to 38.5% (Table 3, Figure 7a,b). In ST40 treated soils, organic P accounted for 57.6% of the P present in the soil prior to planting while adsorbed P accounted for 40.5%. After harvest, organic P reduced slightly to 50.9% while adsorbed P increased, accounting for 49.1% of the P species present (Figure 7a,b).
3.4. X-Ray Fluorescence Imaging
Further investigation into the RF samples AT0Bp and AT40Ah on a microprobe showed that the AT0Bp samples had concentrated discrete particles that contained high levels of both Ca and P (turquoise “hotspots” in Figure 8a), which was consistent with the high apatite content observed by LCF analysis of the bulk XANES for this sample. In contrast, the AT40Ah image showed P that was both correlated with Fe (yellow regions in Figure 8b) and spread diffusely throughout the entire sample. This was what would be expected based on the LCF model that described the sample with P adsorbed on iron oxides and P of organic origin.
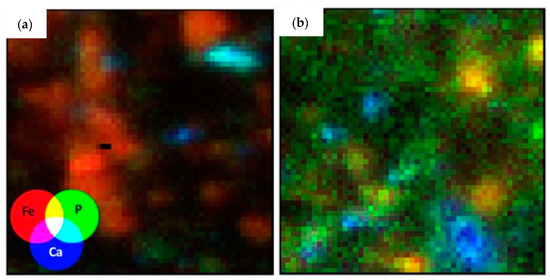
Figure 8.
A tri-color map showing the distribution of phosphorus across rainforest samples (a) AT0Bp and (b) AT40Ah.
4. Discussion
The RF soils were generally more acidic and had higher P content than the DS soils (Table 1 and Table 2). It is possible that the presence of Fe (as evident from the soil acidity) or higher clay content in the RF soils allowed the binding and retention of more P. The concentrations of extracted soil P forms were affected by the interaction of treatment and time, but the interaction between these two factors showed inconsistent effects for all P forms (Table 1 and Table 2). In the DS, total P significantly reduced with increasing rates of urea addition (Figure 3b and Figure 5b); T0 had significantly higher total and available P than T40 in soils cultivated with both AC and SM (Table 1), but the decline in total and available P was more in T0 than in T40 (Table 1), suggesting that urea addition slowed down P mining in these soils. In the RF, soil pH declined with increasing rates of urea addition (Figure 4b); the significant reduction in the soil pH of the T0 and T40 treatments in soils cultivated with AC correlated with the increment in total P (Table 2). This suggested that urea addition was contributing to soil acidification in these treatments and simultaneously increasing the soils’ total P content. There was also a significant decline in available P in both samples at harvest (Table 2). For soils cultivated with SM, the significant increment in the total and available P content of the soils over time in the T0 treatment suggested that the manure added to the soil raised the soils P content over time, whereas the decline in the total P in the urea addition (i.e., T40) and the significant increment in available P in that sample suggested that urea addition caused P mining in this soil.
The LCF results suggested the adsorption of P to mineral phases of Fe, organic P, and Ca phases (in some of the RF samples). These results were in line with other studies [26,31,33] that observed a dominance of adsorbed P in manure amended soils and highlighted the role of Fe in the chemistry of P in manure amended soils. In the DS, the marginal shift in the dominant P species in AT0, from 64.9% adsorbed P and 35.1% organic P (Table 3, Figure 6b) before planting to 63.9% adsorbed P and 33.1% organic P, and the occurrence of apatite P (2.9%) after harvest (Table 3, Figure 6a) were also accompanied by a reduction in the pH and total and available P after harvest (Table 3, Figure 5a,b). This reduction in both the adsorbed and organic P species and the formation of apatite suggested a slow but gradual conversion of P into mineral (apatite) P, which may become soluble as pH decreases by rhizosphere acidification [48]. Apatite can dissolve under acidic conditions that are achievable by rhizosphere acidification and be broken down to adsorbed P [49], which is plant accessible in alkaline and neutral soils because acidification increases the solubility of Ca-P minerals, making Ca-P and sorbed P phases preferentially extracted [49].
In the DS soils amended with manure and 40 kg N/ha, the dominant P form shifted from 67.9% adsorbed P and 32.1% organic P to 40.7% adsorbed P and 58.3% organic P, indicating a significant transformation of P over time from the adsorbed form to the organic form. This was also accompanied by a reduction in pH and total and available P (Table 1, Figure 5a,b). The shift in the P phase in AT40 from adsorbed P to organic P may be due to the microbial immobilization of the available P in the soil into the organic P phase (i.e., from adsorbed P—solution P—organic P). The accompanying reduction in the soil pH may be caused by the production of organic acids by the plant roots and/or soil microorganisms or a product of acidification from the process of nitrification of ammonium [34] in soils treated with urea.
The same speciation of adsorbed and organic P in the AC cultivated soils was also observed in the P K-edge XANES results of the DS soils cultivated with SM. In ST0, both organic and adsorbed P were the dominant P species, accounting for 51.6% and 48.4% of P species, respectively (Table 3, Figure 6b) prior to planting; however, upon harvest, there was a slight reduction in both P species, possibly because of its conversion into apatite P, hence its occurrence with an abundance of 3.1% after harvest (Figure 6a,b). The gradual shift in the P pool from adsorbed and organic P towards apatite P (Figure 7a,b) was consistent with the findings of Kar et al. (2017), who reported the gradual conversion of adsorbed P into apatite P, and Dendougui and Schwedt (2004), who noted the strong affinity of organic P to Ca minerals. This observed shift in P speciation may be due to precipitation of P with Ca2+ in the high CEC ST0Ah soil of the DS (Table 1), leading to a slow transformation of P into the mineral phase in this soil as the soil pH increased (Table 1, Figure 3a), increasing available P while reducing total P (Figure 3b). The ST0Ah LCF spectra however did not have a well pronounced shoulder feature around 2154 to 2158 eV typical of Ca phosphates [44], possibly due to the low P concentration in the soil.
In the ST40 treatment, there was only a marginal change in the P speciation prior to planting and at harvest, with adsorbed and organic P accounting for 40.5% and 57.6% of total P respectively before planting and 49.1% and 50.9% after harvest (Figure 6a,b), although the pH and available P increased as the total P reduced (Table 1, Figure 3a,b). This decrease in total P as the soil pH and P availability increased suggested P mining from the soil by the vegetables, hence explaining the optimum yield from the ST40 treatments (Figure 2). It is possible that as the soil microbes gain access to the N from the urea, they mineralize the organic P in the soil, making them more plant available, hence the reduction in the proportion of the organic P and total P while available P increased (Table 1). This could cause a potential increase in P susceptibility to loss by leaching or runoff in this soil.
In the RF, the major shift in the dominant P species in the AT0 treatment from 81% apatite P (possibly present due to poultry manure amendment, which is known to be high in Ca-P phases [43,50,51]) and 18.9% adsorbed P before planting to organic P (62.9%), adsorbed P (21.5%), and 14.7% apatite P (Table 3, Figure 7b) after harvest strongly suggested that the dominant P cycling process here was microbial mediated immobilization. The apatite P could be expected to weather slowly due to organic acid exudates from soil microbes and/or plant roots, and then, P becomes more plant available and thus taken up out of the soil. It is also possible that some of the P is lost via leaching/runoff as the P is transformed from the mineral phase into the adsorbed/organic phase, hence accounting for the reduced total and available P over time (Table 3).
In the RF AT40 treatment, prior to planting, organic P (47.7%), adsorbed P (42.1%), and apatite P (8.1%) were present. Upon harvest, the dominant P forms shifted to organic P (37.5%) and apatite P (32.9%), while adsorbed P decreased to 27.5% (Table 3, Figure 7b). This suggested a conversion of organic P to the mineral P phase; this is possibly a microbial mediated process of mineralization that makes P become more plant available. However, the XANES LCF showed that mineralized P rapidly formed precipitates with Ca, thereby reducing the availability of P in the soil (Table 3). Furthermore, the huge reduction in the total P after harvest (Table 2) suggested that some of the P may be lost to the environment through erosion and runoff. Furthermore, in the RF, the P K-edge XANES for ST0 soils showed adsorbed and organic P as the only two P species present in the soils, both accounting for 47.6% and 51.4% of the P species respectively before planting (Table 3, Figure 6b), with a small change in the P phase occurring after harvest as organic P increased by 8.7% while adsorbed P reduced by 9.1%. Explanations for this include (a) either some of the adsorbed P is desorbed and then becomes immobilized by the soil microbes or (b) as the manure remains in contact with the soil, it begins to get slowly immobilized by the soil microbes. The latter sounds more feasible, given that the soil pH slightly decreased between planting and harvest, which would favor P immobilization over P desorption. However, in the ST40 treatment, organic P (57.6%) and adsorbed P (40.5%) were the initial P forms (Table 3, Figure 7b), whereas organic P reduced to 50.9% after harvest while the adsorbed P increased to 49.1%. This shift in P phase suggested that as N becomes more available to plants and microbes in the soil, some of the organic P in the soil becomes mineralized and increases in availability (Table 2). As availability increases, some of the P may adsorb to Fe minerals given the low soil pH (Table 2) [52].
5. Conclusions
This study investigated the solid phase speciation of P in manure amended and urea fertilized soil of the rainforest ecozone of Nigeria and dry savanna ecozone of the Republic of Benin under cultivation with two indigenous vegetable species: local amaranth (Amaranthus cruentus) and African eggplant (Solanum macrocarpon), using P K-edge XANES spectroscopy. The quantitative analysis of the XANES spectra of the dry savanna soils of Ina, Republic of Benin, under cultivation by the local amaranth (AC) and the African eggplant (SM) showed that adsorbed P and organic P were the two dominant P species in the manure amended soils before and after planting. In the manure amended soils of the dry savanna, the formation of apatite after harvest as pH and total P reduced showed the gradual conversion of the organic and adsorbed P into mineral P (Ca-P), which may become soluble and dissolve under acidic conditions. It is however difficult to conclude on how N addition affected P speciation and transformation, as the addition of 40 kg N/ha caused an increase in the organic P form in soils cultivated with AC and an increase in adsorbed P in soils cultivated with SM.
In the rainforest soils of Ilesha, Nigeria, XANES analysis showed the presence of apatite P in the manure amended soils cultivated with AC, which was then transformed via microbial immobilization into organic P. When N was added to this soil (i.e., AT40), XANES analysis showed a shift in the dominant P species from organic P before planting to adsorbed P and apatite P, most likely through a combination of mineralization and precipitation processes. In the soils cultivated with SM, organic and adsorbed P were the two dominant P species in both the manure amended (ST0), and the manure amended + urea fertilized soils (ST40). However, at harvest, there was a slow immobilization of P from the adsorbed phase into the organic phase in the manure amended soils (ST0), with N addition reversing this effect and mineralizing the P pool from the organic P to adsorbed P. We concluded that the N addition to the soils of both ecoregions caused P to cycle differently, thereby suggesting that we may be mining P from these soils.
Author Contributions
The authors contributed to this work in the following way: conceptualization and resources, D.O., P.A., and D.P.; methodology, G.K. and A.O.; software, D.P. and A.O.; validation, D.O., G.K., D.P., and A.O.; formal analysis, P.A. and A.O.; writing, original draft preparation, A.O.; writing, review and editing, G.K. and D.P.; visualization, D.P. and A.O.; supervision, D.P.; project administration and funding acquisition, D.O., P.A., and D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the International Development Research Centre (IDRC), Project Number 107983, through the Canada International Food Security Research Fund (CIFSRF) of the Government of Canada through Foreign Affairs, Trade and Development Canada. The authors are extremely grateful.
Acknowledgments
The authors would like to thank all field extension officers, technical support staff, interpreters, and farmers for their significant work and contributions throughout the research. Furthermore, we thank the Canadian Light Source and their funding partners for granting us access to carry out spectroscopic analyses. Special thanks also go to members of the Environmental Soil Chemistry research group at the University of Saskatchewan for their support.
Conflicts of Interest
The authors declare no conflict of interest; the funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; nor in the decision to publish the results.
References
- Zingore, S.; Delve, R.J.; Nyamangara, J.; Giller, K.E. Multiple benefits of manure: The key to maintenance of soil fertility and restoration of depleted sandy soils on African smallholder farms. Nutr. Cycl. Agroecosyst. 2007, 80, 267–282. [Google Scholar] [CrossRef]
- Twomlow, S.; Mugabe, F.T.; Mwale, M.; Delve, R.; Nanja, D.; Carberry, P.; Howden, M. Building adaptive capacity to cope with increasing vulnerability due to climatic change in Africa- a new approach. Phys. Chem. Earth 2008, 33, 780–787. [Google Scholar] [CrossRef]
- United Nations (UN). World Population Prospects: The 2000 Revision; Department of Economic and Social Affairs: New York, NY, USA, 2001. [Google Scholar]
- Bationo, A.B.; Waswa, B.; Abdou, A.; Bado, B.V.; Bonzi, M.; Iwuafor, E.; Kibunja, C.; Kihara, J.; Mucheru, M.; Mugendi, D.; et al. Lessons Learned from Long-Term Soil Fertility Management Experiments in Africa; Springer: Dordrecht, The Netherlands, 2012; pp. 1–26. [Google Scholar]
- Callo-Concha, D.; Gaiser, T.; Webber, H.; Tischbein, B.; Muller, M.; Ewert, F. Farming in the West African Sudan Savanna: Insights in the context of climate change. Afr. J. Agric. Res. 2013, 8, 4693–4705. [Google Scholar] [CrossRef]
- Alliance for a Green Revolution in Africa (AGRA). Building on the New Momentum in African Agriculture: AGRA in 2008; Alliance for a Green Revolution in Africa: Nairobi, Kenya, 2009. [Google Scholar]
- Vanlauwe, B.; Kihara, J.; Chivenge, P.; Pypers, P.; Coe, R.; Six, J. Agronomic use efficiency of N fertilizer in maize-based systems in sub-Saharan Africa within the context of integrated soil fertility management. Plant Soil 2010, 339, 35–50. [Google Scholar] [CrossRef]
- International Crops Research Institute for the Semi-Arid Tropics (ICRISAT). Fertilizer Microdosing; ICRISAT Publ.: Patancheru, India, 2009; Available online: www.icrisat.org/fertilizermicrodosing (accessed on 3 May 2018).
- Adebooye, O.C. Proximate composition and nutrient analysis of six selected leaf vegetables of southwest Nigeria. IFE J. Agric. 1996, 18, 56–62. [Google Scholar]
- Adebooye, O.C. Wild Plants for Medicinal and Culinary Uses: Nigeria. Sharing Innovative Experiences; TWAS: Trieste, Italy, 2001; Volume 2, pp. 69–78. [Google Scholar]
- United States Department of Agriculture (USDA). USDA Nutrient Database for Standard Reference; Release 16; US. Department of Agriculture, Agriculture Research Service, Nutrient Data Laboratory: Beltsville, MD, USA, 2003. Available online: www.nal.usda.gov/fnic/foodcomp (accessed on 12 January 2016).
- Smith, I.F.; Eyzaguirre, P. African leafy vegetables: Their role in the world health organization’s global fruit and vegetables initiative. Afr. J. Food Agric. Nutr. 2007, 7, 3. [Google Scholar]
- Adams, A.M.; Gillespie, A.W.; Kar, G.; Koala, S.; Ouattara, B.; Kimaro, A.A.; Bationo, A.; Akponikpe, P.B.I.; Schoenau, J.J.; Peak, D. Long term effects of reduced fertilizer rates on millet yields and soil properties in the West-African Sahel. Nutr. Cycl. Agroecosyst. 2016, 106, 17–29. [Google Scholar] [CrossRef]
- Hammond, J.P.; White, P.J. Sucrose transport in the phloem: Integrating root responses to phosphorus starvation. J. Exp. Bot. 2008, 59, 93–109. [Google Scholar] [CrossRef]
- Pierzynski, G.M.; Sims, J.T.; Vance, G.F. Soils and Environmental Quality, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Richardson, A.E. Utilization of Soil Organic Phosphorus by Higher Plants; Organic Phosphorus in the Environment; CABI Publishing: Oxfordshire, UK, 2005; pp. 165–184. [Google Scholar]
- Castillo, A.E.; Benito, S.G.; Fernandez, J.A. Using organic manures as liming materials. Agrochimica 2003, 47, 14–20. [Google Scholar]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Nitrogen. In Principles of Plant Nutrition; Springer: Dordrecht, The Netherland, 2001; pp. 397–434. [Google Scholar]
- Cooperband, L.; Bollero, G.; Coale, F. Effect of poultry litter and composts on soil nitrogen and phosphorus availability and corn production. Nutr. Cycl. Agroecosyst. 2002, 62, 185–194. [Google Scholar] [CrossRef]
- Kashem, M.A.; Akinremi, O.O.; Racz, G.J. Phosphorus fractions in soil amended with organic and inorganic phosphorus sources. Can. J. Soil Sci. 2004, 84, 83–90. [Google Scholar] [CrossRef]
- Montgomery, D.R. Dirt the Erosion of Civilizations; University of California Press: Berkeley, CA, USA, 2007. [Google Scholar]
- Schoenau, J.J.; Carley, C.; Stumborg, C.; Malhi, S.S. Strategies for maximizing crop recovery of applied manure nitrogen in the Northern Great Plains of North America. In Recent Trends in Soil Science and Agronomy Research in the Northern Great Plains of North America; Malhi, S.S., Gan, Y., Schoenau, J.J., Lemke, R.L., Mark, A., Eds.; Liebig. Research Signpost: Kerala, India, 2010; pp. 95–108. [Google Scholar]
- Lemke, R.L.; Farrell, R.E.; Malhi, S.S. Greenhouse gas emissions from crop production systems in the Northern Great Plains of North America. In Recent Trends in Soil Science and Agronomy Research in the Northern Great Plains of North America; Malhi, S.S., Gan, Y., Schoenau, J.J., Lemke, R.L., Mark, A., Eds.; Liebig. Research Signpost: Kerala, India, 2010; pp. 95–108. [Google Scholar]
- Campbell, C.A.; Janzen, H.H.; Juma, N.G. Case studies of soil quality in the Canadian prairies: Long-term field experiments. Dev. Soil Sci. 1997, 25, 351–397. [Google Scholar]
- Assefa, B.A.; Schoenau, J.J.; Grevers, M.C.J. Effects of four annual applications of manure on Black Chernozemic soils. Can. Biosyst. Eng. 2004, 46, 39–46. [Google Scholar]
- Kar, G.; Schoenau, J.J.; Hilger, D.; Peak, D. Direct chemical speciation of soil Phosphorus in a Saskatchewan Chernozem after long- and short-term manure amendments. Can. J. Soil Sci. 2017, 97, 626–636. [Google Scholar] [CrossRef]
- Dormaar, J.F.; Chang, C. Effects of 20 annual applications of excess feedlot manure on labile soil phosphorus. Can. J. Soil Sci. 1995, 75, 507–512. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Racz, G.J. Laboratory characterization of P in fresh and oven-dried organic amendments. J. Environ. Qual. 2004, 33, 1062–1069. [Google Scholar] [CrossRef]
- Schoenau, J.J.; Davis, J.G. Optimizing soil and plant responses to land-applied manure nutrients in the Great Plains of North America. Can. J. Soil Sci. 2006, 86, 587–595. [Google Scholar] [CrossRef]
- Hesterberg, D.; Weiqing, Z.; Hutchison, K.J.; Beauchemin, S.; Sayers, D.E. XAFS study of adsorbed and mineral forms of phosphate. J. Synchrotron Rad. 1999, 6, 636–638. [Google Scholar] [CrossRef]
- Beauchemin, S.; Hesterberg, D.; Chou, J.; Beauchemin, M.; Simard, R.R.; Sayers, D.E. Speciation of phosphorus in phosphorus-enriched agricultural soils using X-ray absorption near-edge structure spectroscopy and chemical fractionation. J. Environ. Qual. 2003, 32, 1809–1819. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Hu, Y.; Flaten, D.N. Phosphorus speciation of sequential extracts of organic amendments using nuclear magnetic resonance and X-ray absorption near-edge structure spectroscopies. J. Environ. Qual. 2007, 36, 1563–1576. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Hu, Y.; Jurgensen, A. XANES Speciation of Phosphorus in Organically Amended and Fertilized Vertisol and Mollisol. Soil Sci. Soc. Am. J. 2008, 72, 1256–1262. [Google Scholar] [CrossRef]
- Kar, G.; Peak, D.; Schoenau, J.J. Spatial distribution and chemical speciation of soil phosphorus in a band application. Soil Sci. Soc. Am. J. 2012, 76, 2297–2306. [Google Scholar] [CrossRef]
- Prietzel, J.; Dümig, A.; Wu, Y.; Zhou, J.; Klysubun, W. Synchrotron-based P K-edge XANES spectroscopy reveals rapid changes of phosphorus speciation in the topsoil of two glacier foreland chronosequences. Geochim. Cosmochim. Acta 2013, 108, 154–171. [Google Scholar] [CrossRef]
- Alotaibi, K.D.; Schoenau, J.J.; Kar, G.; Peak, D.; Fonstad, T. Phosphorus speciation in a prairie soil amended with MBM and DDG ash: Sequential chemical extraction and synchrotron-based XANES spectroscopy investigations. Sci. Rep. 2018, 8, 3617. [Google Scholar] [CrossRef]
- Hamilton, J.G.; Grosskleg, J.; Hilger, D.; Bradshaw, K.; Carlson, T.; Siciliano, S.D.; Peak, D. Chemical speciation and fate of tripolyphosphate after application to a calcerous soil. Geochem. Trans. 2018, 19, 1. [Google Scholar] [CrossRef]
- Ingall, E.D.; Brandes, J.A.; Diaz, J.M.; de Jonge, M.D.; Paterson, D.; McNulty, I.; Elliott, W.C.; Northrup, P. Phosphorus Kedge XANES spectroscopy of mineral standards. J. Synchrotron Radiat. 2011, 18, 189–197. [Google Scholar] [CrossRef]
- Negassa, W.; Kruse, J.; Michalik, D.; Appathurai, N.; Zuin, L.; Leinweber, P. Phosphorus Speciation in Agro-Industrial Byproducts: Sequential Fractionation, Solution 31P NMR, and P K-and L2,3-Edge XANES Spectroscopy. Environ. Sci. Technol. 2010, 44, 2092–2097. [Google Scholar] [CrossRef]
- Condron, L.M.; Newman, S. Revisiting the fundamentals of phosphorus fractionation of sediments and soils. J. Soils Sediments 2011, 11, 830–840. [Google Scholar] [CrossRef]
- Khare, N.; Hesterberg, D.; Martin, J.D. XANES investigation of phosphate sorption in single and binary systems of iron and aluminum oxide minerals. Environ. Sci. Technol. 2005, 39, 2152–2160. [Google Scholar] [CrossRef]
- Kelly, S.D.; Hesterberg, D.; Ravel, B. Analysis of soils and minerals using X-ray absorption spectroscopy. Methods of Soil Analysis. Part 5. Mineral. Methods 2008, 5, 387–463. [Google Scholar]
- Peak, D. Solid-state speciation of natural and alum-amended poultry litter using XANES spectroscopy. Environ. Sci. Technol. 2002, 36, 4253–4261. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Solomon, D.; Hyland, C.; Ketterings, Q.M.; Lehmann, J. Phosphorus speciation in manure and manure-amended soils using XANES spectroscopy. Environ. Sci. Technol. 2005, 39, 7485–7491. [Google Scholar] [CrossRef] [PubMed]
- Kruse, J.; Leinweber, P. Phosphorus in sequentially extracted fen peat soils: A K-edge X-ray absorption near-edge structure (XANES) spectroscopy study. Environ. Sci. Technol. 2008, 39, 7485–7491. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis, 2nd ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2008. [Google Scholar]
- Zuo, M.; Renman, G.; Gustafsson, J.P.; Renman, A. Phosphorus removal performance and speciation in virgin and modified argon oxygen decarburisation slag designed for wastewater treatment. Water Res. 2015, 87, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.O.; Tighe, M.K.; Guppy, C.N.; Milham, P.J.; McLaren, T.I.; Schefe, C.R.; Lombi, E. XANES demonstrates the release of calcium phosphates from alkaline vertisols to moderately acidified solution. Environ. Sci. Technol. 2016, 50, 4229–4237. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, A.K.; Hillier, S.; Hesterberg, D.; Klysubun, W.; Ulén, B.; Gustafsson, J.P. Evolution of phosphorus speciation with depth in an agricultural soil profile. Geoderma 2016, 280, 29–37. [Google Scholar] [CrossRef]
- Klopfenstein, T.J.; Angel, R.; Cromwell, G.; Erickson, G.E.; Fox, D.G.; Parsons, C.; Satter, L.D.; Sutton, A.L.; Baker, D.H.; Lewis, A.; et al. Animal diet modification to decrease the potential for nitrogen and phosphorus pollution. In Faculty Papers and Publications in Animal Science; University of Nebraska Press: Lincoln, NE, USA, 2002; p. 518. Available online: http://digitalcommons.unl.edu/animalscifacpub/518 (accessed on 8 January 2020).
- Toor, G.S.; Peak, D.J.; Sims, J.T. Phosphorus speciation in broiler litter and turkey manure produced from modified diets. J. Environ. Qual. 2005, 34, 687–697. [Google Scholar] [CrossRef]
- Dendougui, F.; Schwedt, G. In vitro analysis of binding capacities of calcium to organic acid in different food samples. Eur. Food Res. Technol. 2004, 219, 409–415. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).