Impact of Carao (Cassia grandis) on Lactobacillus plantarum Immunomodulatory and Probiotic Capacity

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Tolerance to Bile Salt
2.3. Tolerance to pH
2.4. Tolerance to NaCl
2.5. Viability of Lactobacillus plantarum
2.6. Viability of Lactobacillus plantarum under Gastrointestinal Digestion
2.7. Effect of Carao on Intestinal Infection with Y. enterocolitica O9 In Vivo
2.7.1. Animals
2.7.2. Preparation of Fermented Milk Added with L. plantarum
2.7.3. Experimental Infection with Y. enterocolitica O9 IP383
2.7.4. Measurement of Cytokines and IgA
2.8. Statistical Analysis
3. Results
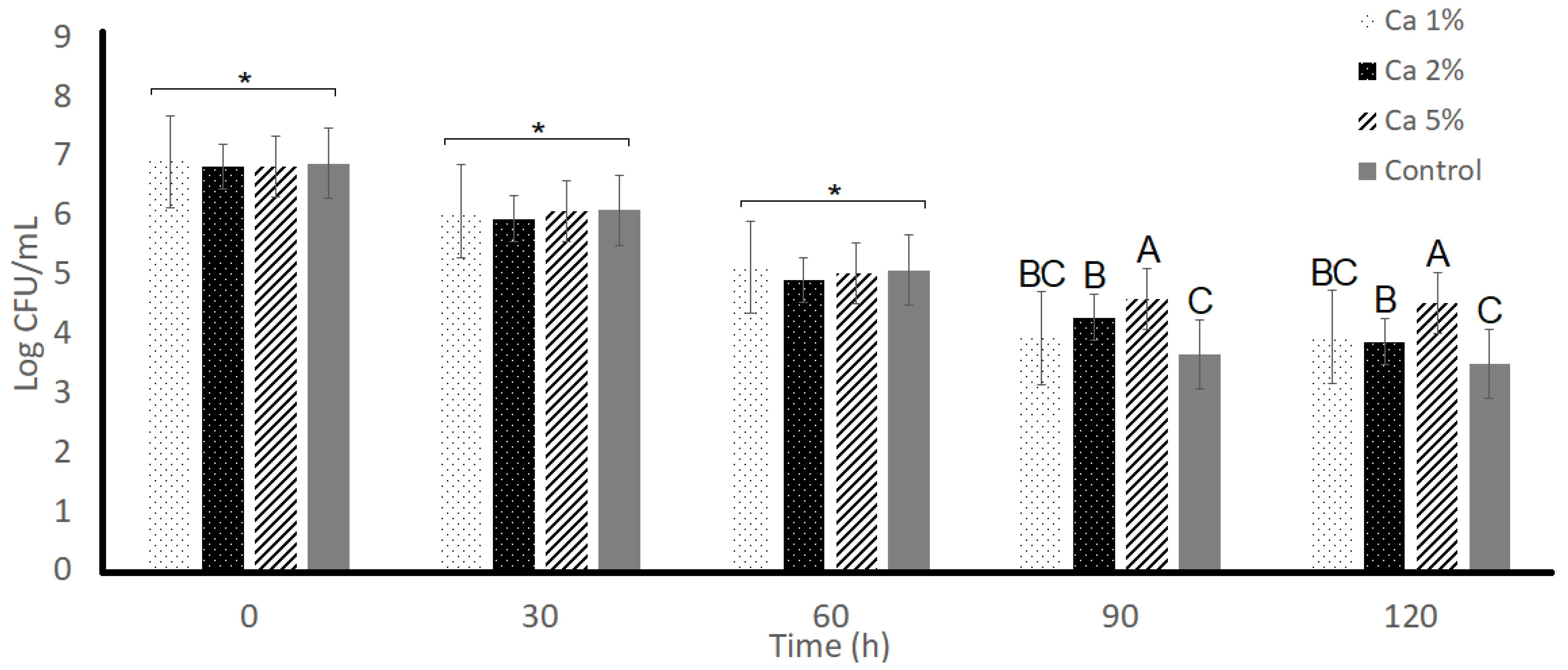
3.1. Bile Tolerance
3.2. Acid Tolerance
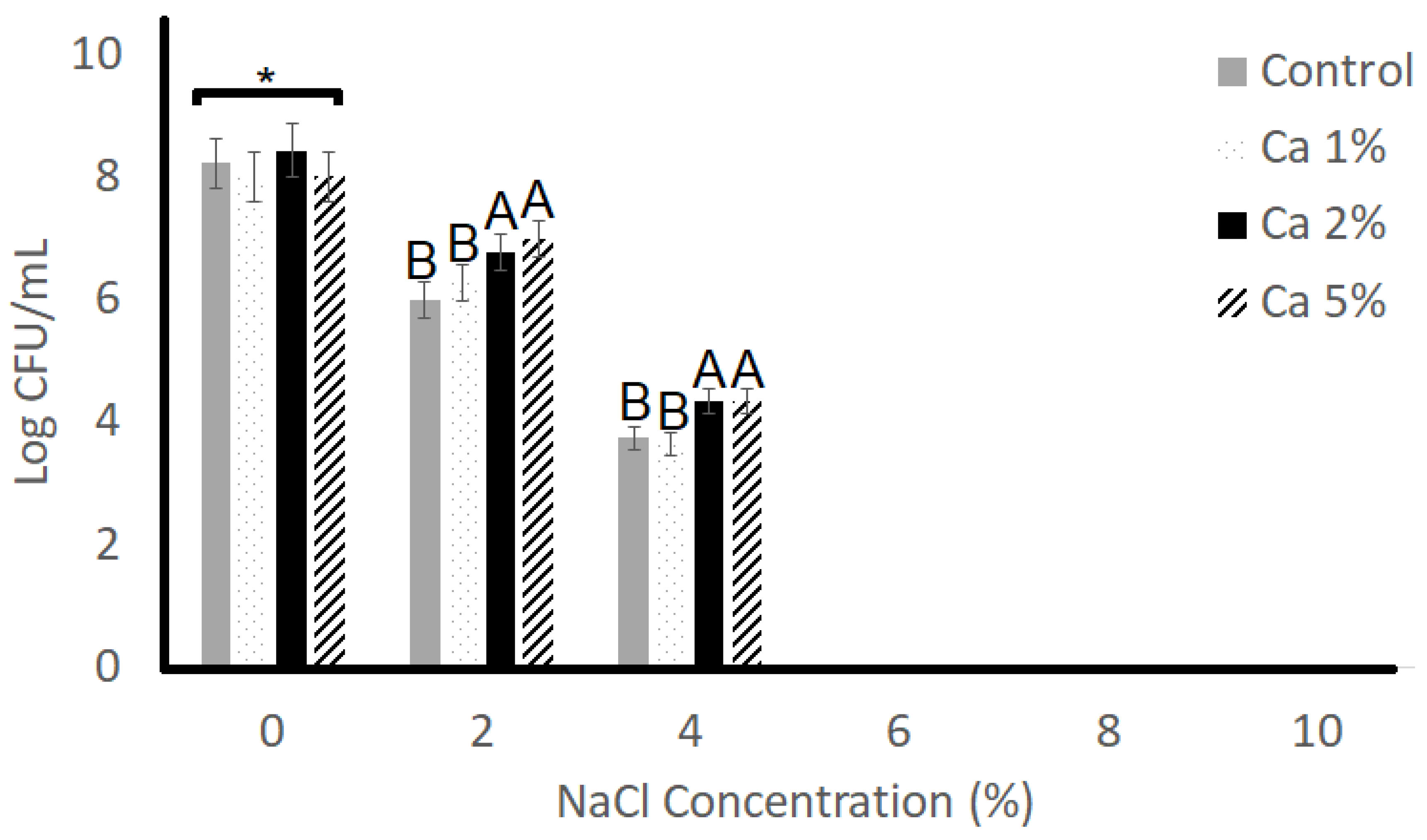
3.3. Resistance to NaCl
3.4. L. plantarum Viability Characteristics
3.5. L. plantarum Resistance to the Gastrointestinal Digestion Process In Vitro: Carao Influence
3.6. Effect of Treatment with L. plantarum with Carao on Experimental Infection with Y. enterocolitica O9 in Mice
3.7. Effect of Treatment with L. plantarum with Carao on Intestinal Cytokine and IgA Production in Uninfected Mice and Y. enterocolitica O9-Infected Mice
3.8. Effects of Treatment with L. plantarum with Carao on Plasma Cytokine Levels in Uninfected Mice and Y. enterocolitica O9-Infected Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gueimonde, M.; Collado, M.C. Metagenomics and probiotics. Clin. Microbiol. Infect. 2012, 18, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Levri, K.M.; Ketvertis, K.; Deramo, M.; Merenstein, J.H.; D’Amico, F. Do probiotics reduce adult lactose intolerance? J. Fam. Pract. 2005, 54, 613–620. [Google Scholar] [PubMed]
- Binns, N. Probiotics, Prebiotics and the Gut Microbiota; ILSI Europe: Brussels, Belgium, 2013; ISBN 978-90-78637-39-4. [Google Scholar]
- Corgneau, M.; Scher, J.; Ritie-Pertusa, L.; Le, D.T.L.; Petit, J.; Nikolova, Y.; Banon, S.; Gaiani, C. Recent Advances on Lactose Intolerance: Tolerance Thresholds and Currently Available Answers. Crit. Rev. Food Sci. Nutr. 2017, 57, 3344–3356. [Google Scholar] [CrossRef] [PubMed]
- Aronson, J.K. Defining ‘nutraceuticals’: Neither nutritious nor pharmaceutical. Br. J. Clin. Pharmacol. 2017, 83, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Hasler, C.M. Functional foods: Benefits, concerns and challenges—A position paper from the American Council on Science and Health. J. Nutr. 2002, 132, 3772–3781. [Google Scholar] [CrossRef] [PubMed]
- Govender, M.; Choonara, Y.E.; Kumar, P.; Du Toit, L.C.; Van Vuuren, S.; Pillay, V. A review of the advancements in probiotic delivery: Conventional vs. non-conventional formulations for intestinal flora supplementation. AAPS Pharm. Sci. Tech. 2014, 15, 29–43. [Google Scholar] [CrossRef]
- Marcía-Fuentes, J.; Santos-Aleman, R.; Borrás-Linares, I.; Sánchez, J.L. The Carao (Cassia grandis L.): Its Potential Usage in Pharmacological, Nutritional, and Medicinal Applications. In Innovations in Biotechnology for a Sustainable Future; Maddela, N.R., García, L.C., Eds.; Springer: Cham, Switzerland, 2021; pp. 403–427. [Google Scholar]
- Aleman, R.S.; Marcia, J.; Page, R.; Kazemzadeh Pournaki, S.; Martín-Vertedor, D.; Manrique-Fernández, V.; Montero-Fernández, I.; Aryana, K. Effects of Yogurt with Carao (Cassia grandis) on Intestinal Barrier Dysfunction, α-glycosidase Activity, Lipase Activity, Hypoglycemic Effect, and Antioxidant Activity. Fermentation 2023, 9, 566. [Google Scholar] [CrossRef]
- Fuentes, J.A.M.; Fernández, I.M.; Fernández, H.Z.; Sánchez, J.L.; Alemán, R.S.; Navarro-Alarcon, M.; Borrás-Linares, I.; Maldonado, S.A.S. Quantification of bioactive molecules, minerals and bromatological analysis in Carao (Cassia grandis). J. Agric. Sci. 2020, 12, 88–94. [Google Scholar] [CrossRef]
- Marcia, J.; Aleman, R.S.; Montero-Fernández, I.; Martín-Vertedor, D.; Manrique-Fernández, V.; Moncada, M.; Kayanush, A. Attributes of Lactobacillus acidophilus as Effected by Carao (Cassia grandis) Pulp Powder. Fermentation 2023, 9, 408. [Google Scholar] [CrossRef]
- Aleman, R.S.; Avila, D.; Avila, A.; Losso, J.N.; Picha, D.; Xu, Z.; Aryana, K. Chemical Characterization and Impact of Nipple Fruit (Solanum mammosum) on the Characteristics of Lactobacillus acidophilus LA K. Fermentation 2023, 9, 715. [Google Scholar] [CrossRef]
- Aleman, R.S.; Avila, D.; Avila, A.; Marcia, J.; Picha, D.; Aryana, K.; Montero-Fernández, I. Effects of Weevil (Rhynchophorus palmarum), Teosinte (Dioon mejiae) and Caesar’s Mushroom (Amanita caesarea) on the Properties of Lactobacillus acidophilus LA-K. Fermentation 2023, 9, 852. [Google Scholar] [CrossRef]
- Barragán, P.J.; Sánchez, Ó.J.; Henao-Rojas, J.C. Evaluation of the Growth Kinetics of Lactobacillus plantarum ATCC 8014 on a Medium Based on Hydrolyzed Bovine Blood Plasma at Laboratory and Bench-Scale Levels and Its Application as a Starter Culture in a Meat Product. Fermentation 2020, 6, 45. [Google Scholar] [CrossRef]
- Gbassi, K.G.; Vandamme, T.; Ennahar, S.; Marchioni, E. Microencapsulation of Lactobacillus plantarum spp. in an alginate matrix coated with whey proteins. Int. J. Food Microbiol. 2009, 129, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Bergillos-Meca, T.; Costabile, A.; Walton, G.; Moreno-Montoro, M.; Ruiz-Bravo, A.; Ruiz-López, M.D. In vitro evaluation of the fermentation properties and potential probiotic activity of lactobacillus plantarum c4 in batch culture systems. LWT—Food Sci. Technol. 2015, 60, 420–426. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Moreno, E.; Bergillos-Meca, T.; Lasserrot, A.; Ruiz-López, M.D.; Ruiz-Bravo, A.; Jiménez-Valera, M. A Lactobacillus plantarum strain isolated from kefir protects against intestinal infection with Yersinia enterocolitica O9 and modulates immunity in mice. Res. Microbiol. 2015, 166, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, S.J.; Schuurman-Wolters, G.K.; Poolman, B. Crystal Structure of the Substrate-Binding Domain from Listeria monocytogenes Bile-Resistance Determinant BilE. Crystals 2016, 6, 162. [Google Scholar] [CrossRef]
- Lara Mantilla, C.; Burgos Portacio, Á. Potencial probiótico de cepas nativas para uso como aditivos en la alimentación avícola. Rev. Colomb. De Biotecnol. 2012, 14, 31–40. [Google Scholar]
- Cueto-Vigil, M.C.; Yudtanduly, A.-M.; Valenzuela-Riaño, J. Evaluación in vitro del potencial probiótico de bacterias ácido lácticas aisladas de suero costeño. Actual. Biológicas 2012, 32, 129–138. [Google Scholar] [CrossRef]
- Khagwai, N.; Sharma, P.; Chand, D. Screening and evaluation of Lacobacillus spp. for the development of potential probiotics. Afr. J. Microbiol. Res. 2014, 8, 1573–1579. [Google Scholar]
- Martínez-Cárdenas, L.; Álvarez-González, C.A.; Hernández-Almeida, O.U.; Frías-Quintana, C.A.; Ponce-Palafox, J.T.; Castillo-Vargasmachuca, S. Partial Characterization of Digestive Proteases in the Green Cichlid, Cichlasoma beani. Fishes 2017, 2, 4. [Google Scholar] [CrossRef]
- Altieri, C.; Bevilacqua, A.; Sinigaglia, M. Prolonging the Viability of Lactobacillus Plantarum through the Addition of Prebiotics into the Medium. J. Food Sci. 2011, 76, M336–M345. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.J.; Lee, J.H.; Hutkins, R.W. Functional analysis of the fructooligosaccharide utilization operon in Lactobacillus paracasei 1195. Appl. Environ. Microbiol. 2007, 73, 5716–5724. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, H.; Hutkins, R.W. Fermentation of fructooligosaccharides by lactic acid bacteria and bifidobacteria. Appl. Environ. Microbiol. 2000, 66, 2682–2684. [Google Scholar] [CrossRef] [PubMed]
- Bujalance, C.; Moreno, E.; Jimenez-Valera, M.; Ruiz-Bravo, A. A probiotic strain of lactobacillus plantarum stimulates lymphocyte responses in immunologically intact and immunocompromised mice. Int. J. Food Microbiol. 2007, 113, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Bujalance, C.; Jiménez-Valera, M.; Moreno, E.; Ruiz-López, M.D.; Lasserrot, A.; Ruiz-Bravo, A. Lack of correlation between in vitro antibiosis and in vivo protection against enteropathogenic bacteria by probiotic lactobacilli. Res. Microbiol. 2014, 165, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Oellerich, M.; Jacobi, C.; Freund, S.; Niedung, K.; Bach, A.; Heesemann, J.; Trülzsch, K. Yersinia enterocolitica infection of mice reveals clonal invasion and abscess formation. Infect. Immun. 2007, 75, 3802–3811. [Google Scholar] [CrossRef] [PubMed]
- Barthel, M.; Hapfelmeier, S.; Quintanilla-Martínez, L.; Kremer, M.; Rohde, M.; Hogardt, M.; Pfeffer, K.; Rüssmann, H.; Hardt, W.D. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect. Immun. 2003, 71, 2839–2858. [Google Scholar] [CrossRef] [PubMed]
- Autenrieth, I.B.; Heesemann, J. In vivo neutralization of tumor necrosis factor-alpha and interferon-gamma abrogates resistance to Yersinia enterocolitica infection in mice. Med. Microbiol. Immunol. 1992, 181, 333–338. [Google Scholar] [CrossRef]
- Autenrieth, I.B.; Firsching, R. Penetration of M cells and destruction of Peyer’s patches by Yersinia enterocolitica: An ultrastructural and histological study. J. Med. Microbiol. 1996, 44, 285–294. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | L. casei |
|---|---|
| Bile tolerance | |
| Carao concentration | 0.0432 |
| Time (Hours) | <0.0001 |
| Carao concentration × time | 0.2587 |
| Acid Tolerance | |
| Carao concentration | 0.0254 |
| Time (Minutes) | <0.0001 |
| Carao concentration × time | 0.1543 |
| Microbial growth on TSB | |
| Carao concentration | 0.0201 |
| Time (Hours) | <0.0001 |
| Carao concentration × time | 0.1349 |
| Microbial growth on LPSM | |
| Carao concentration | 0.0765 |
| Time (Hours) | <0.0001 |
| Carao concentration × time | 0.1674 |
| Microbial growth under gastrointestinal digestion | |
| Carao concentration | 0.0980 |
| Time (Hours) | <0.0001 |
| Carao concentration × time | 0.1245 |
| Tolerance to NaCl | |
| Carao concentration | 0.0284 |
| NaCl concentration | <0.0001 |
| Carao concentration × time | 0.0723 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcia, J.; Zumbado, H.M.; Gil, M.Á.; Martín-Vertedor, D.; Montero-Fernández, I.; Yadav, A.; Aleman, R.S. Impact of Carao (Cassia grandis) on Lactobacillus plantarum Immunomodulatory and Probiotic Capacity. Appl. Microbiol. 2024, 4, 704-719. https://doi.org/10.3390/applmicrobiol4020048
Marcia J, Zumbado HM, Gil MÁ, Martín-Vertedor D, Montero-Fernández I, Yadav A, Aleman RS. Impact of Carao (Cassia grandis) on Lactobacillus plantarum Immunomodulatory and Probiotic Capacity. Applied Microbiology. 2024; 4(2):704-719. https://doi.org/10.3390/applmicrobiol4020048
Chicago/Turabian StyleMarcia, Jhunior, Hector Manuel Zumbado, Manuel Álvarez Gil, Daniel Martín-Vertedor, Ismael Montero-Fernández, Ajitesh Yadav, and Ricardo S. Aleman. 2024. "Impact of Carao (Cassia grandis) on Lactobacillus plantarum Immunomodulatory and Probiotic Capacity" Applied Microbiology 4, no. 2: 704-719. https://doi.org/10.3390/applmicrobiol4020048