



Biomolecules 2023, 13(4), 664; https://doi.org/10.3390/biom13040664 - 11 Apr 2023
Cited by 36 | Viewed by 4918
Abstract
►
Show Figures
Methyltransferase-like 3 (METTL3) is a typical component of N6-methyladenosine writers that exhibits methyltransferase activity and deposits methyl groups on RNA. Currently, accumulating studies have demonstrated the involvement of METTL3 in the regulation of neuro-physiological and pathological events. However, no reviews have comprehensively summarized
[...] Read more.
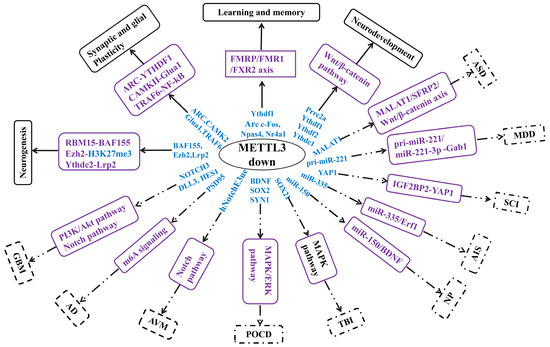
Methyltransferase-like 3 (METTL3) is a typical component of N6-methyladenosine writers that exhibits methyltransferase activity and deposits methyl groups on RNA. Currently, accumulating studies have demonstrated the involvement of METTL3 in the regulation of neuro-physiological and pathological events. However, no reviews have comprehensively summarized and analyzed the roles and mechanisms of METTL3 in these events. Herein, we are focused on reviewing the roles of METTL3 in regulating normal neurophysiological (Neurogenesis, Synaptic Plasticity and Glial Plasticity, Neurodevelopment, Learning and Memory,) and neuropathological (Autism Spectrum Disorder, Major Depressive Disorder, Neurodegenerative disorders, Brain Tumors, Brain Injuries, and Other Brain Disorders) events. Our review found that although the down-regulated levels of METTL3 function through different roles and mechanisms in the nervous system, it primarily inactivates neuro-physiological events and triggers or worsens neuropathological events. In addition, our review suggests that METTL3 could be used as a diagnostic biomarker and therapeutic target in the nervous system. Collectively, our review has provided an up-to-date research outline of METTL3 in the nervous system. In addition, the regulatory network for METTL3 in the nervous system has been mapped, which could provide directions for future research, biomarkers for clinical diagnosis, and targets for disease treatment. Furthermore, this review has provided a comprehensive view, which could improve our understanding of METTL3 functions in the nervous system.
Full article
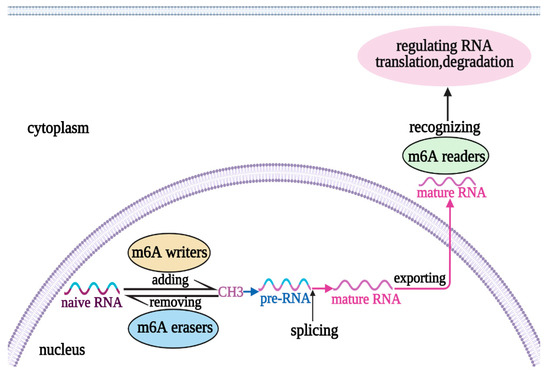
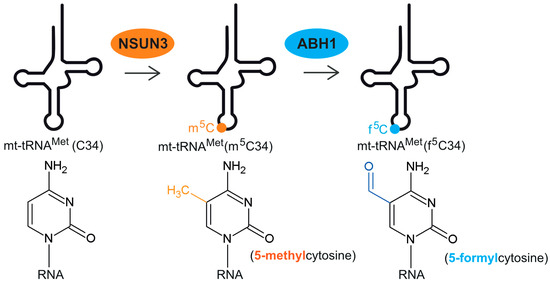
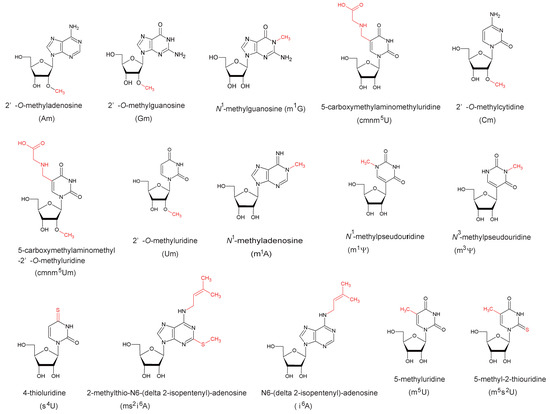
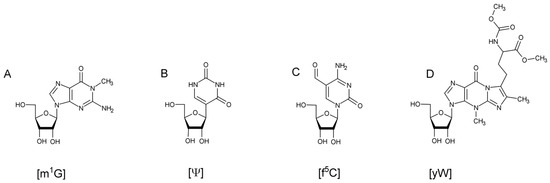
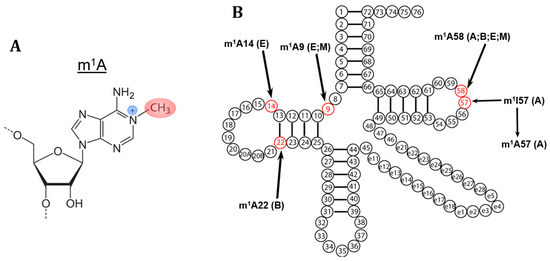
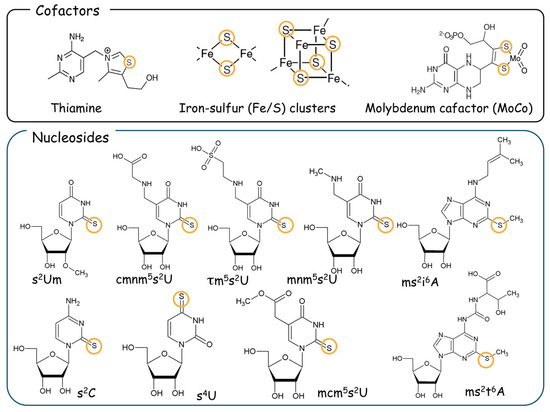
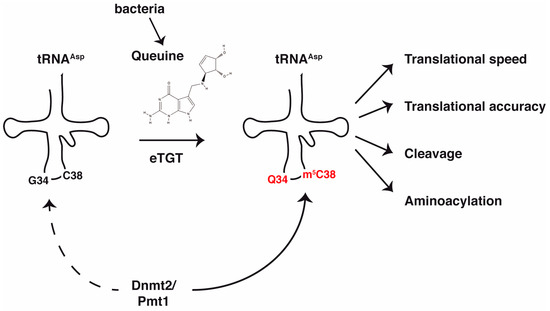
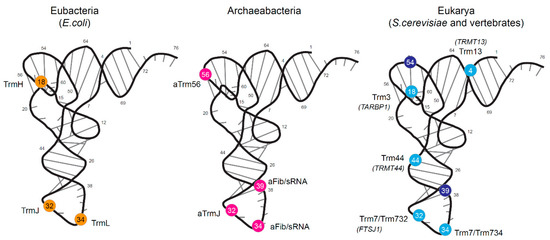
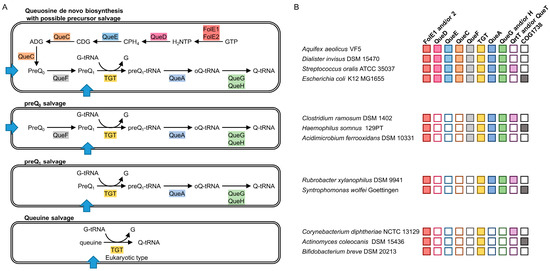
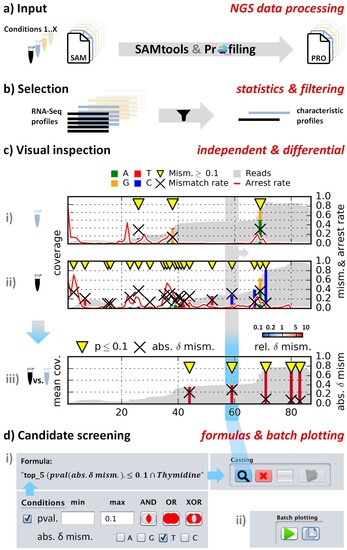
Figure 1
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}