Perturbed and Unperturbed: Analyzing the Conservatively Perturbed Equilibrium (Linear Case)



Abstract
:1. Introduction
- The equilibrium concentration values of all species are determined.
- Some of the species, at least two, are chosen to have their concentration perturbed from the equilibrium value.
- At least one species is not chosen, with its concentration value being kept at the equilibrium value.
- The perturbations mentioned in point 2 are required to satisfy all conservation laws applicable to the system reactions.
- The evolutions of all species concentrations are observed as they tend towards equilibrium.
- (a)
- on the detailed mechanism
- (b)
- on the values of the kinetic parameters
- (c)
- on the possibility of new regimes with an improved yield and selectivity.
1.1. Previous Study Review
1.2. Achieving Momentary Equilibrium (ME) at Some Extrema
- What is the influence of the mechanism structure on the CPE properties?
- Which differences exist between noncyclic reactions and cyclic ones? Or four cycles with a diagonal step?
- What is the influence of the strategy of perturbation—i.e., the distribution of perturbed and unperturbed species, their vicinity and interconnectivity within the mechanism?
- Is it possible to observe more complex dynamic behavior, such as the evolution of events, two extrema, overshooting the equilibrium value, etc.?
2. Materials and Methods
3. Results
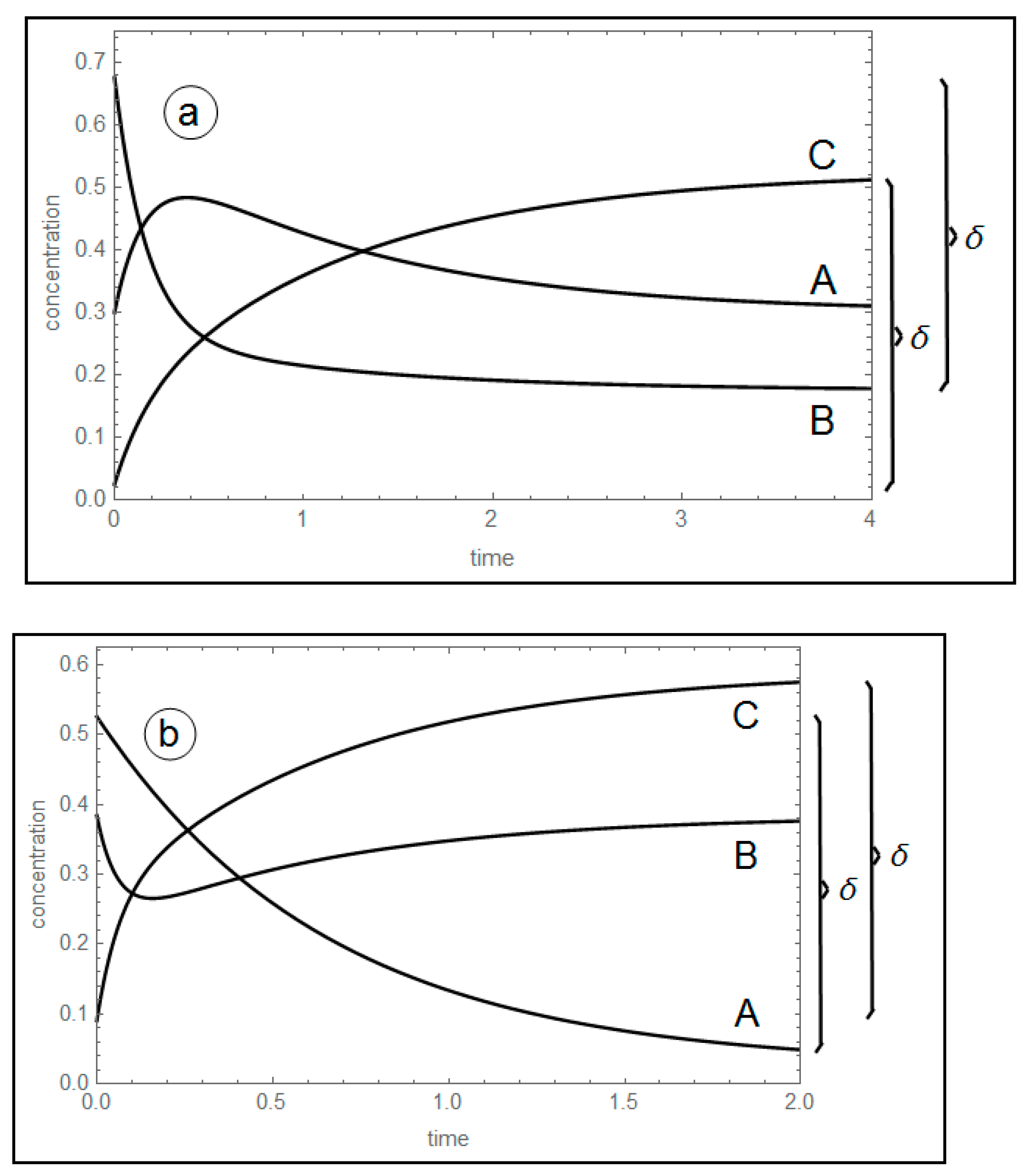
3.1. Analysis of Perturbed Species in a Three-Species Acyclic Mechanism (Two-Step Mechanism)
New Findings—Perturbed Species May Experience Either Monotone Relaxation or Behavior with one Extremum Peak
3.2. Three-Species Cyclic Mechanism
3.2.1. New Findings on the Three-Species Cyclic Mechanism—The Extremum Time for the Cyclic Mechanism Has the Same Analytical Expression as for the Acyclic
3.2.2. The Cyclic Mechanism’s Extremum Time is Shorter than that of the Acyclic Mechanism
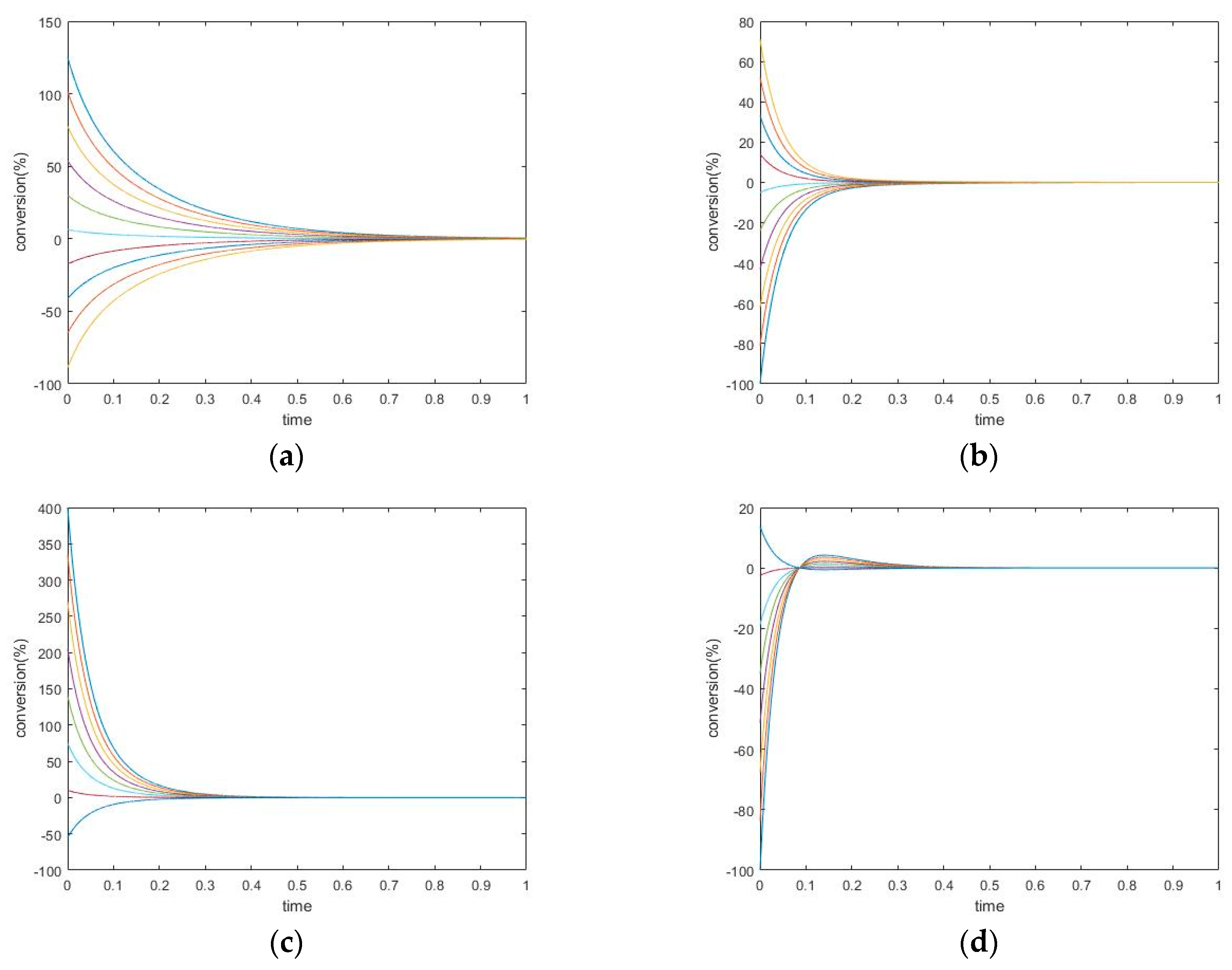
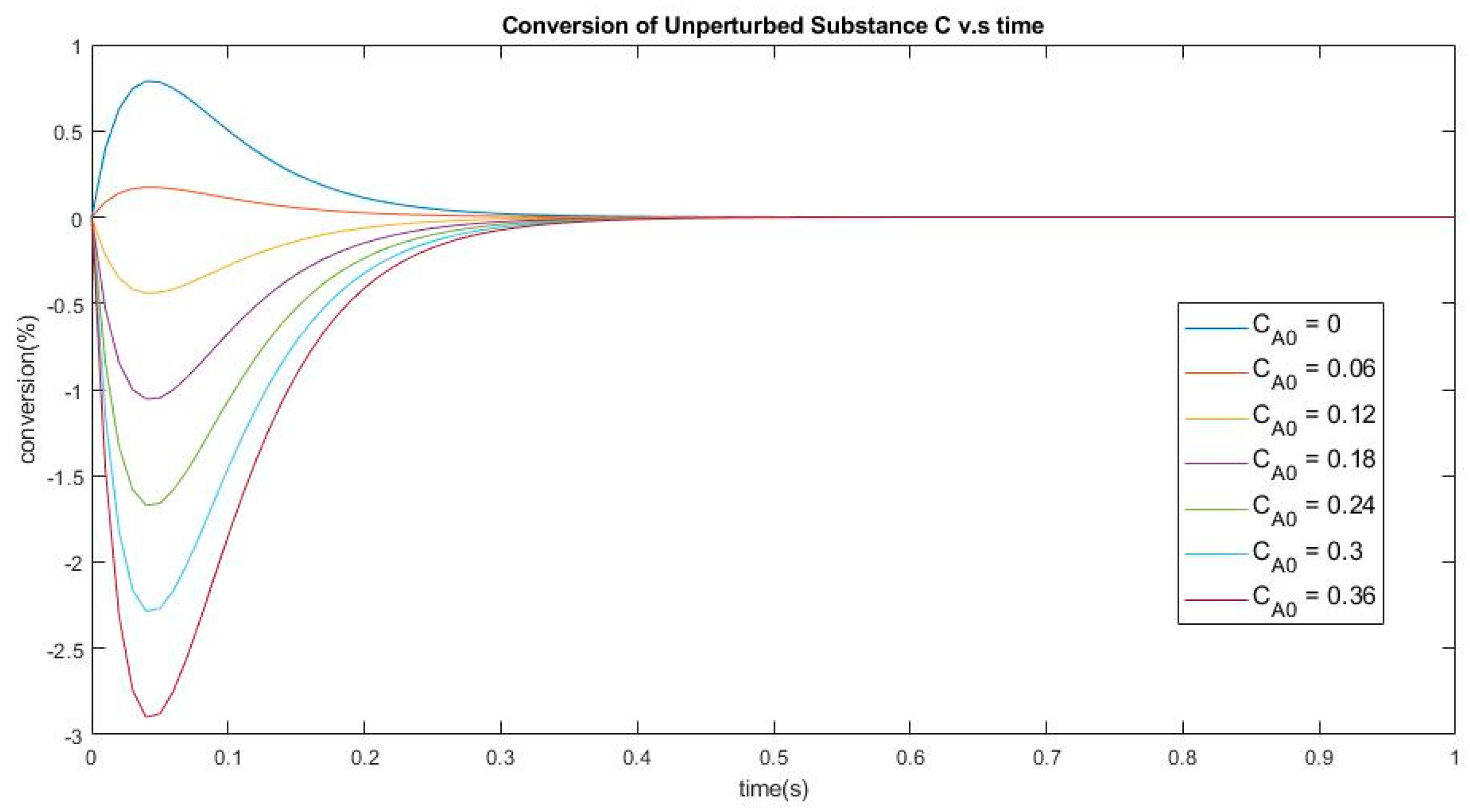
3.3. Four-Species Acyclic Mechanism
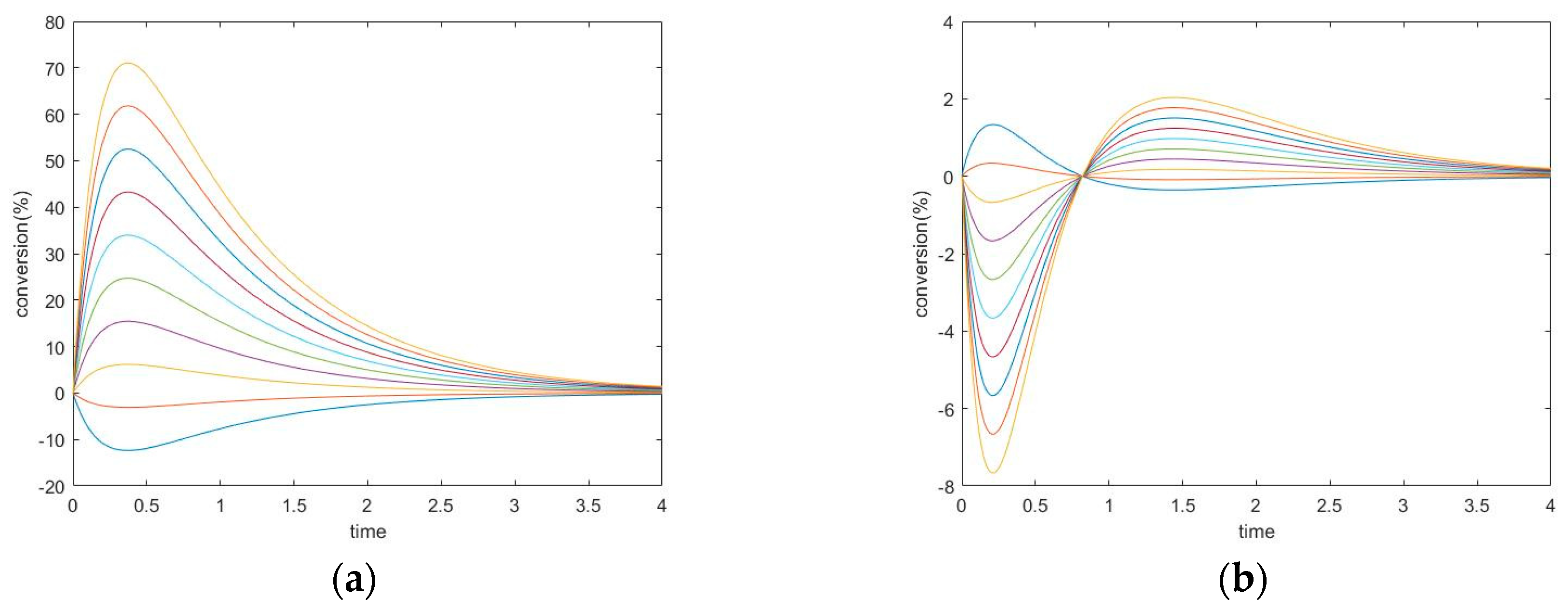
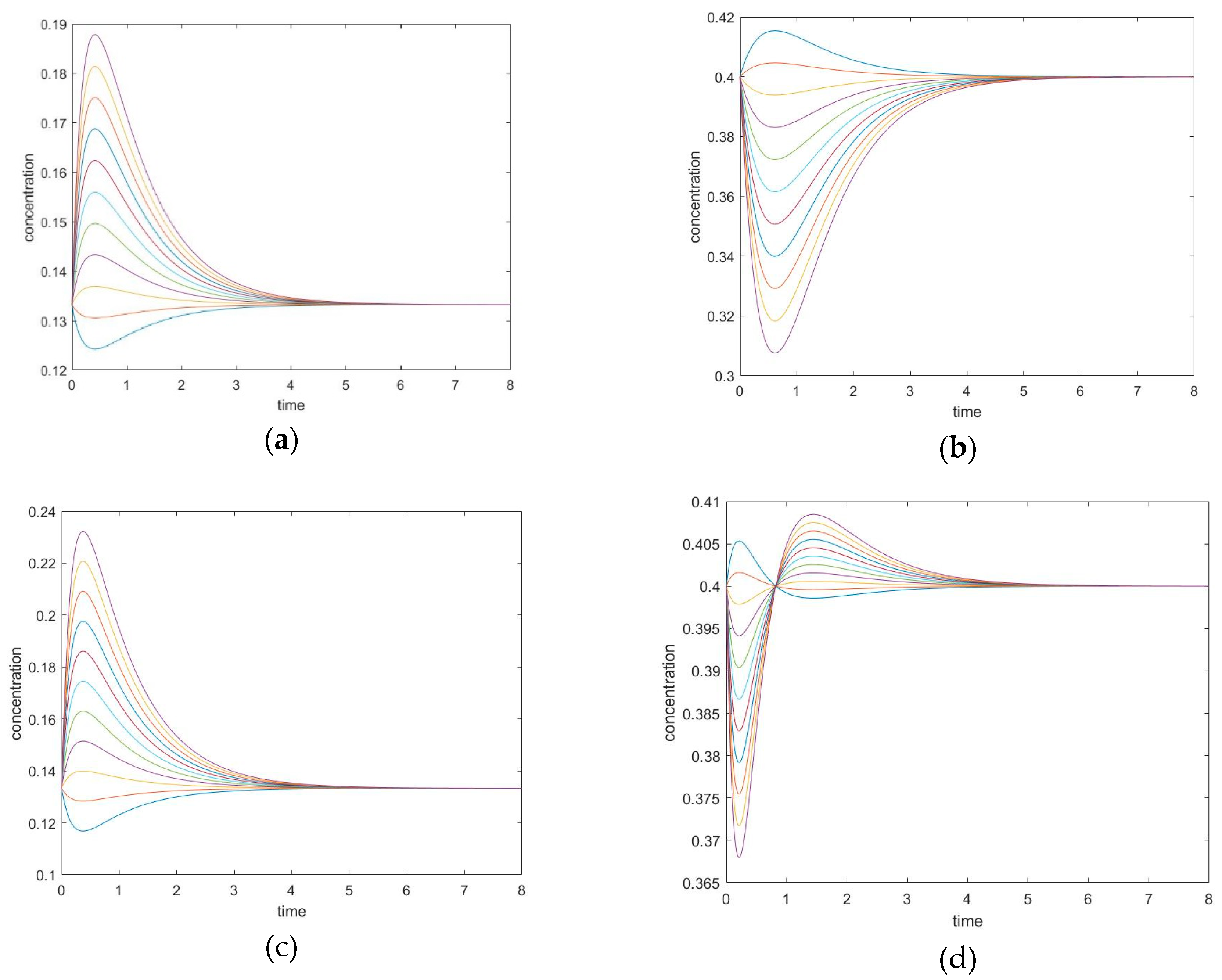
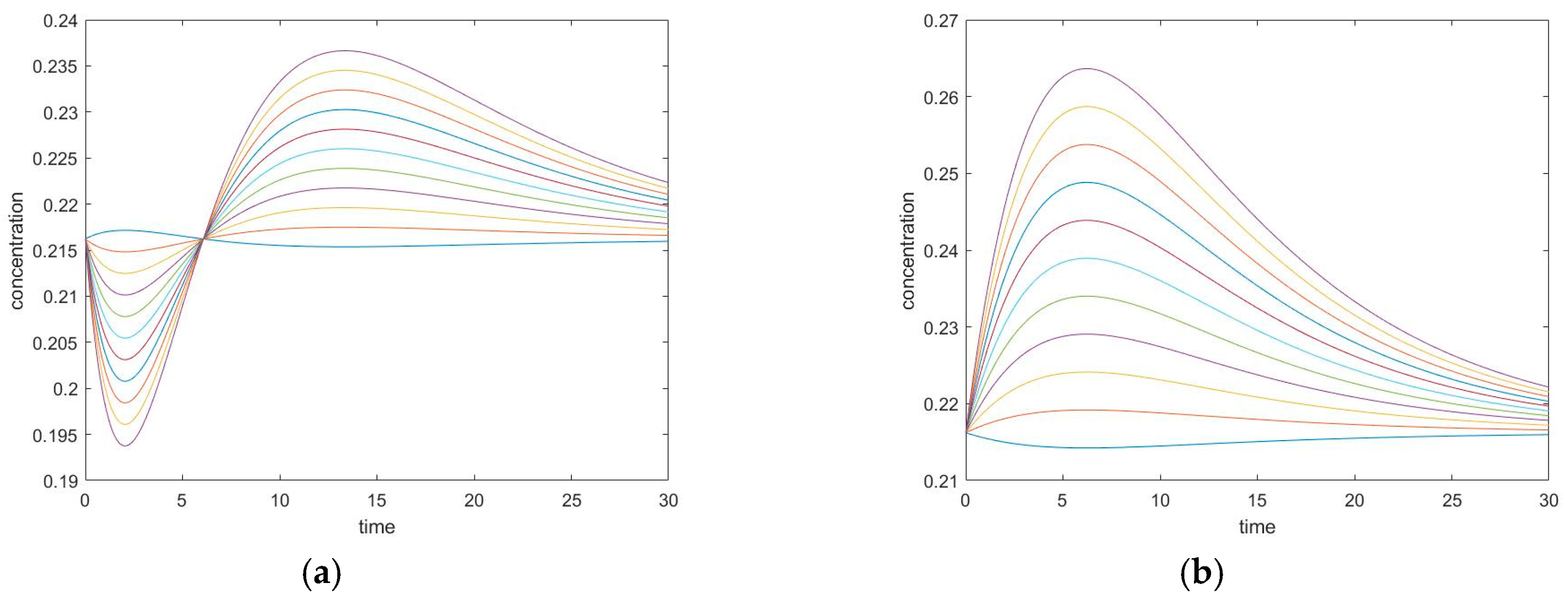
3.3.1. New Findings—Possibility of Two Extrema and an Inflection Point
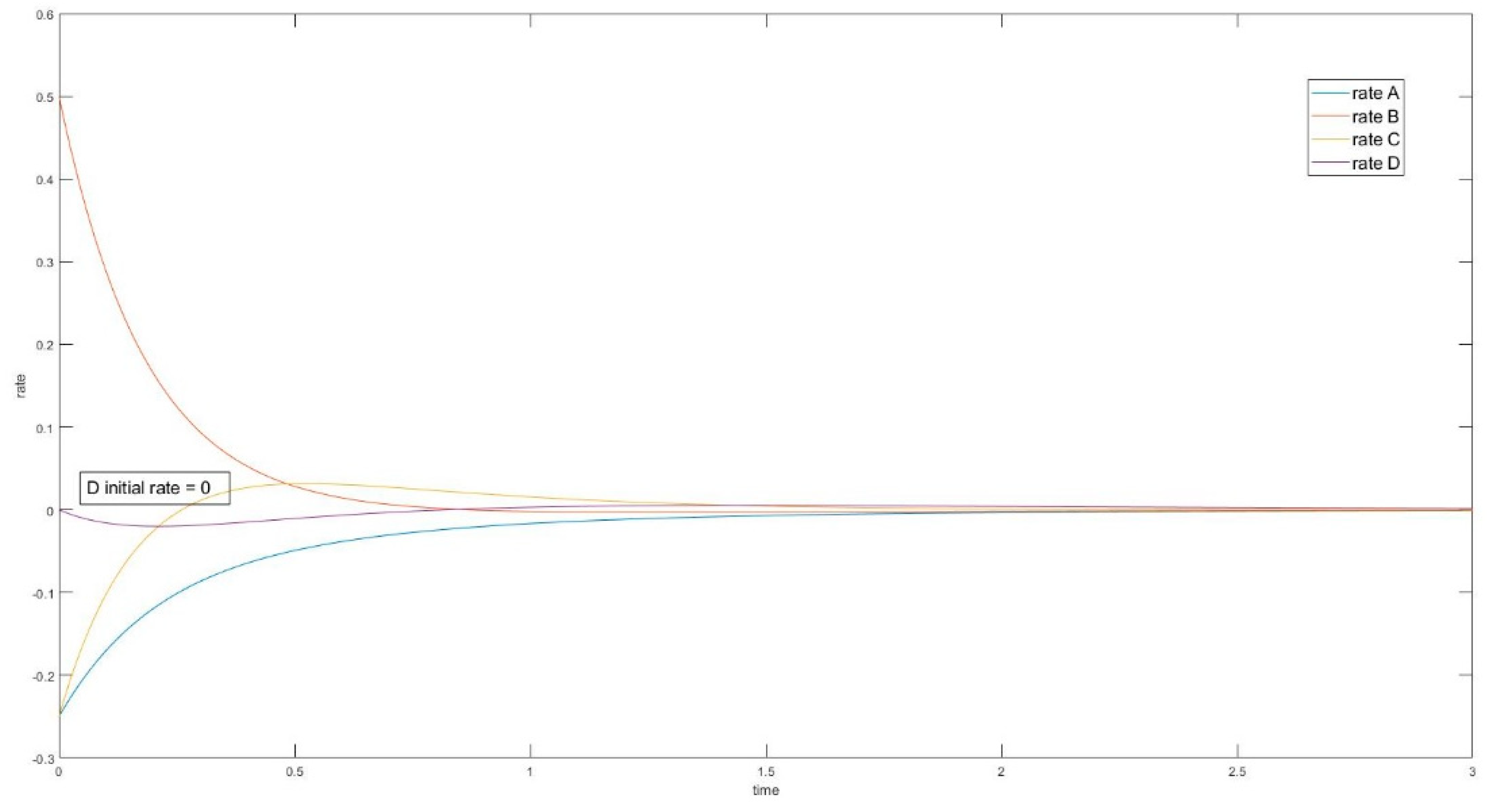
3.3.2. The Initial Rate is Zero for Unperturbed Species that are Connected only with Other Unperturbed Species
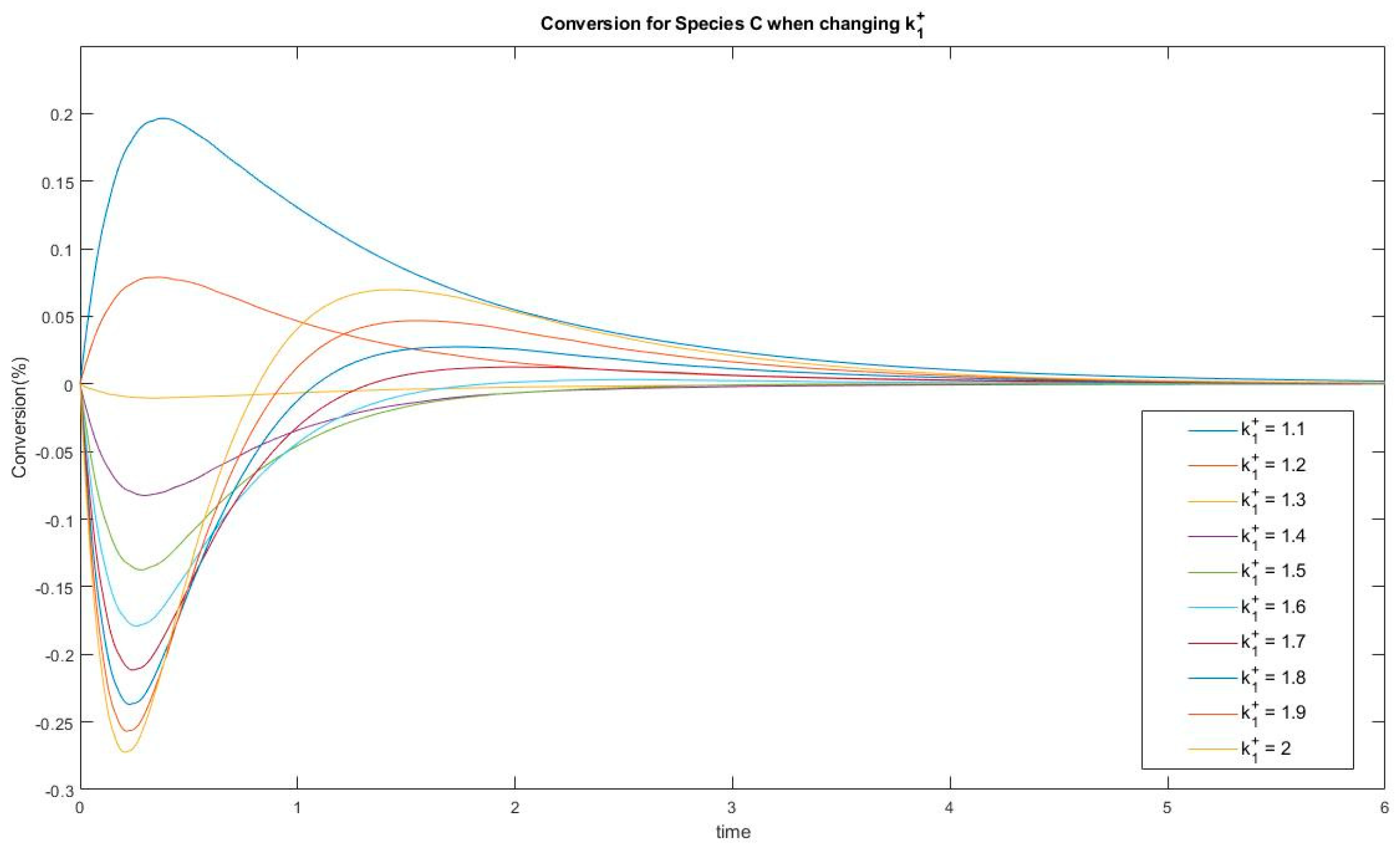
3.3.3. Evolution of Events: Change of the Number of Extrema due to Change in Kinetic Parameter Values
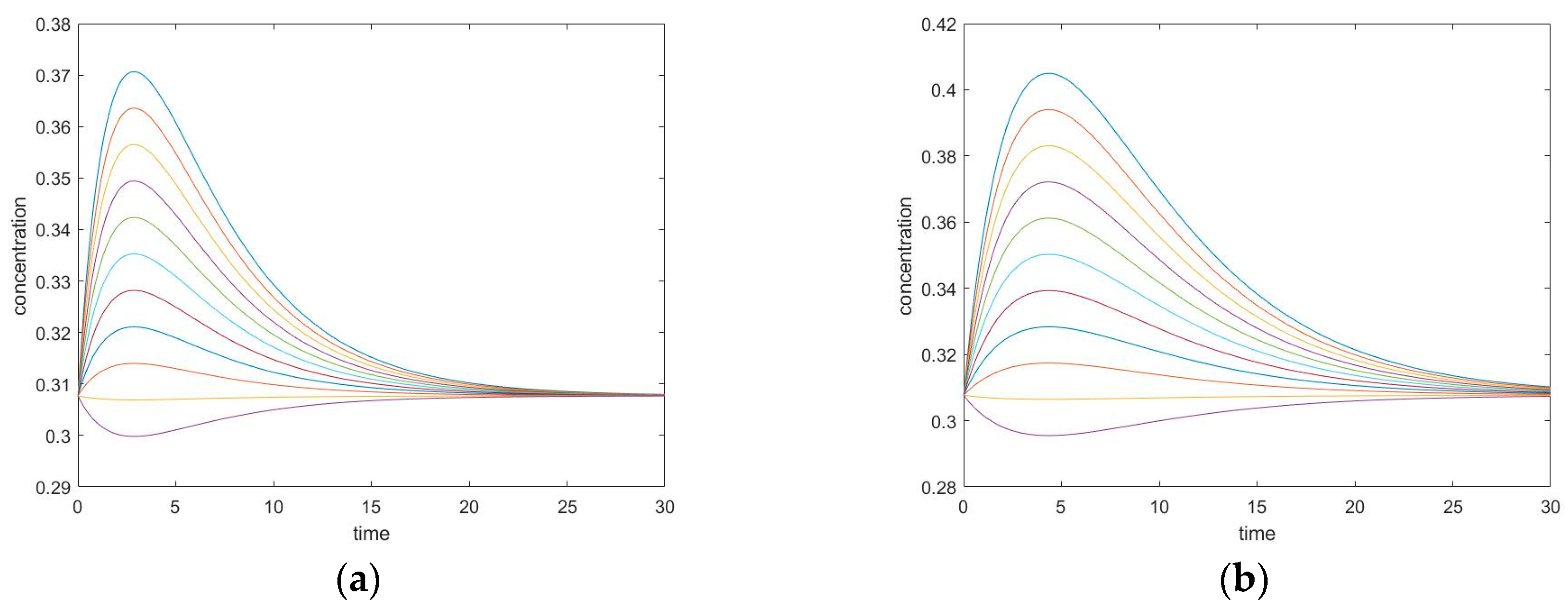
3.4. Four-Species Cyclic Mechanism
3.4.1. New Findings—Similarity with Four-Species Acyclic Mechanism: Occurrence of Two Extrema and an Inflection Point
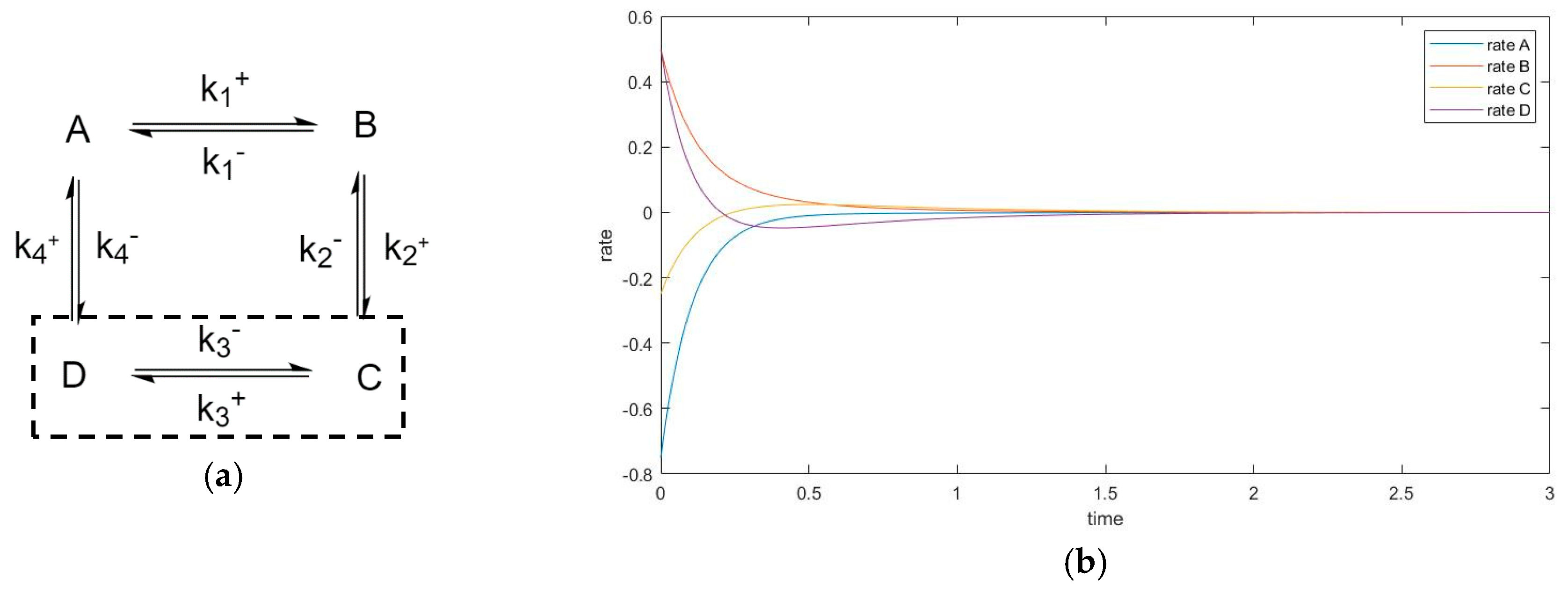
3.4.2. Zero Initial Rate Behavior for Unperturbed Species Does not Occur
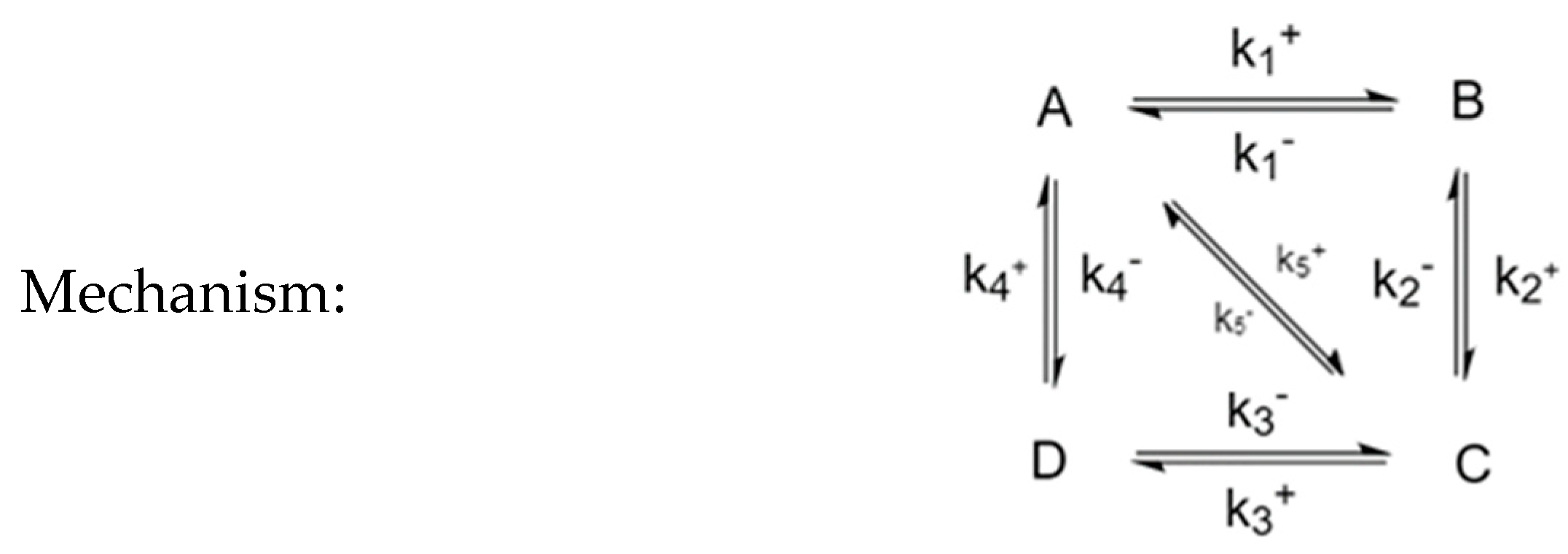
3.5. Four-Species Cyclic Mechanism with Additional Diagonal Connectivity
4. Discussion
4.1. Comparing Structural Differences: Number of Species in Mechanism
4.2. Evolution of Events: Effects of Kinetic Parameters on Complexity
5. Conclusions and Future Applications of CPE
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- Yablonsky, G.S.; Branco, P.D.; Marin, G.B.; Constales, D. Conservatively Perturbed Equilibrium (CPE) in Chemical Kinetics. Chem. Eng. Sci. 2019, 196, 384–390. [Google Scholar] [CrossRef]
- Yablonsky, G.S.; Branco, P.D.; Marin, G.B.; Constales, D. New Invariant Expressions in Chemical Kinetics. Entropy 2020, 22, 373. [Google Scholar] [CrossRef] [Green Version]
- Yablonsky, G.S.; Constales, D.; Marin, G.B. Equilibrium relationships for non-equilibrium chemical dependencies. Chem. Eng. Sci. 2011, 66, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Yablonsky, G.S.; Gorban, A.N.; Constales, D.; Galvita, V.; Marin, G.B. Reciprocal Relations Between Kinetic Curves. Europhys. Lett. 2011, 93, 20004–20007. [Google Scholar] [CrossRef]
- Yablonsky, G.S.; Constales, D.; Marin, G.B. New Types of Complexity in Chemical Kinetics: Intersections, Coincidences and Special Symmetric Relationships. Adv. Chem. Phys. 2014, 157, 69–73. [Google Scholar]
- Branco Pinto, D.; Yablonsky, G.S.; Marin, G.B.; Constales, D. New Patterns in Steady-State Chemical Kinetics: Intersections, Coincidences, Map of Events (Two-Step Mechanism). Entropy 2015, 17, 6783–6800. [Google Scholar] [CrossRef] [Green Version]
- Hankins, M.J.; Yablonsky, G.S.; Kiss, I.Z. Dual kinetic curves in reversible electrochemical systems. PLoS ONE 2017, 12, e0173786. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Yablonsky, G.S.; Constales, D.; Marin, G.B.; Muhler, M. Experimental confirmation of a new invariant for a non-linear chemical reaction. Chem. Eng. Sci. 2018, 191, 262–267. [Google Scholar] [CrossRef]
- Yablonsky, G.S.; Constales, D.; Marin, G.B. Joint kinetics: A new paradigm for chemical kinetics and chemical engineering. Curr. Opin. Chem. Eng. 2020, 29, 83–88. [Google Scholar] [CrossRef]
- Zeldovich, Y.B. Proof of the uniqueness of the solution of mass-action law equations. Zh. Fiz. Khim. 1938, 11, 685–687. (In Russian) [Google Scholar]
- Zeldovich, Y.B. Selected Works of Yakov Borisovich Zeldovich; Volume I: Chemical and Hydrodynanics, Chapter “Proof of the Uniqueness of the Solution of the Equations of the Law of Mass Action”; Princeton University Press: Princeton, NJ, USA, 2014; pp. 144–147. [Google Scholar]
- Shapiro, N.Z.; Shapley, L.S. Mass action laws and the Gibbs free energy function. J. Soc. Ind. Appl. Math. 1965, 13, 353–375. [Google Scholar] [CrossRef]
- Aris, R. Prolegomena to the rational analysis of systems of chemical reactions. Arch. Ration. Mech. Anal. 1965, 19, 81–99. [Google Scholar] [CrossRef]
- Aris, R. Prolegomena to the rational analysis of systems of chemical reactions II. Some addenda. Arch. Ration. Mech. Anal. 1968, 27, 356–364. [Google Scholar] [CrossRef]
- Horn, F.; Jackson, R. General mass action kinetics. Arch. Ration. Mech. Anal. 1972, 47, 81–116. [Google Scholar] [CrossRef]
- Vol’pert, A.I. Differential equations on graphs. Math. USSR Sbornik. 1972, 17, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Vol’pert, A.I.; Khudyaev, S.I. Analysis in Classes of Discontinuous Functions and Equations of Mathematical Physics; Martinus Nijhoff: Dordrecht, The Netherland, 1985; 704p. [Google Scholar]
- Gorban, A.N. On the problem of boundary equilibrium points. React. Kinet. Catal. Lett. 1980, 15, 315–319. [Google Scholar]
- Gorban, A.N.; Mirkes, E.M.; Yablonsky, G.S. Thermodynamics in the limit of irreversible reactions. Phys. A Stat. Mech. Its Appl. 2013, 392, 1318–1335. [Google Scholar] [CrossRef] [Green Version]
- Gorban, A.N.; Yablonsky, G.S. Extended detailed balance for systems with irreversible reactions. Chem. Eng. Sci. 2011, 63, 5388–5399. [Google Scholar] [CrossRef] [Green Version]
- Yablonskii, G.S.; Bykov, V.I.; Gorban, A.N.; Elokhin, V.I. Kinetic Models of Catalytic Reactions. In Comprehensive Chemical Kinetics; Compton, R.G., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; Volume 32, 396p. [Google Scholar]
- Marin, G.B.; Yablonsky, G.S.; Constales, D. Kinetics of Chemical Reactions: Decoding Complexity, 2nd ed.; John Wiley–VCH: Weinheim, Germany, 2019; p. 13. [Google Scholar]
- Gorban, A.N.; Yablonsky, G.S. Three Waves of Chemical Dynamics. Math. Model. Nat. Phenom. 2015, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.; Zhu, X.; Constales, D.; Yablonsky, G.S. Experimental verification of conservatively perturbed equilibrium for a complex non-linear chemical reaction. Chem. Eng. Sci. 2020, 229, 116008. [Google Scholar] [CrossRef]
- Eigen, M. Nobel Prize Lecture, “Immeasurably Fast Reactions”. 11 December 1967. Available online: https://www.nobelprize.org/uploads/2018/06/eigen-lecture.pdf (accessed on 28 September 2020).
- Bernasconi, C.F. Relaxation Kinetics; Academic Press: Cambridge, MA, USA, 1976; 288p. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
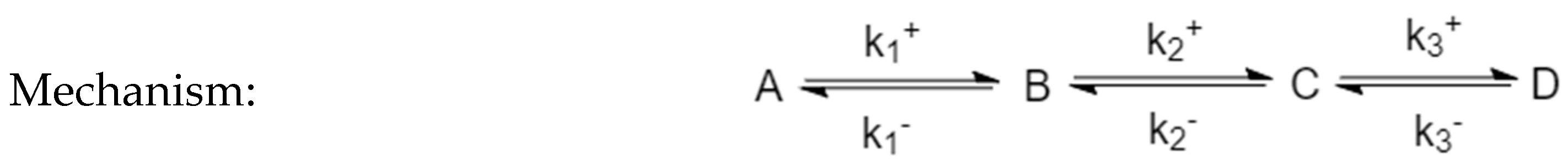{kind=link}
{kind=link}
{kind=link}
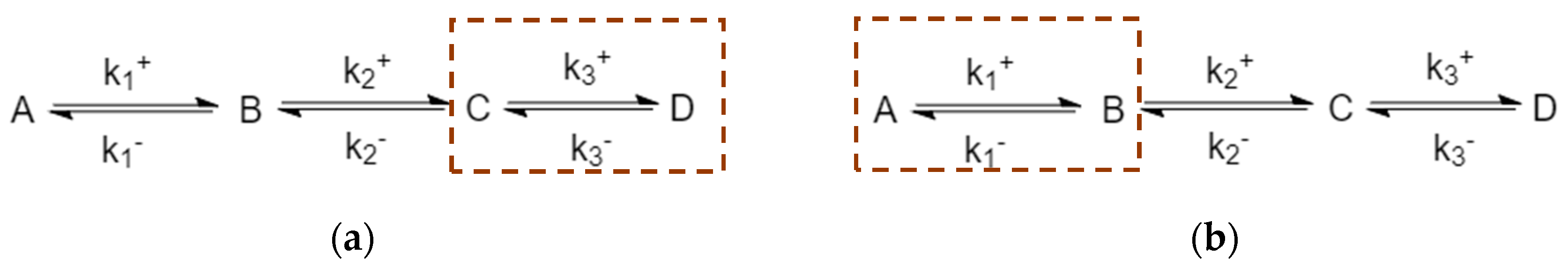{kind=link}
{kind=link}
{kind=link}
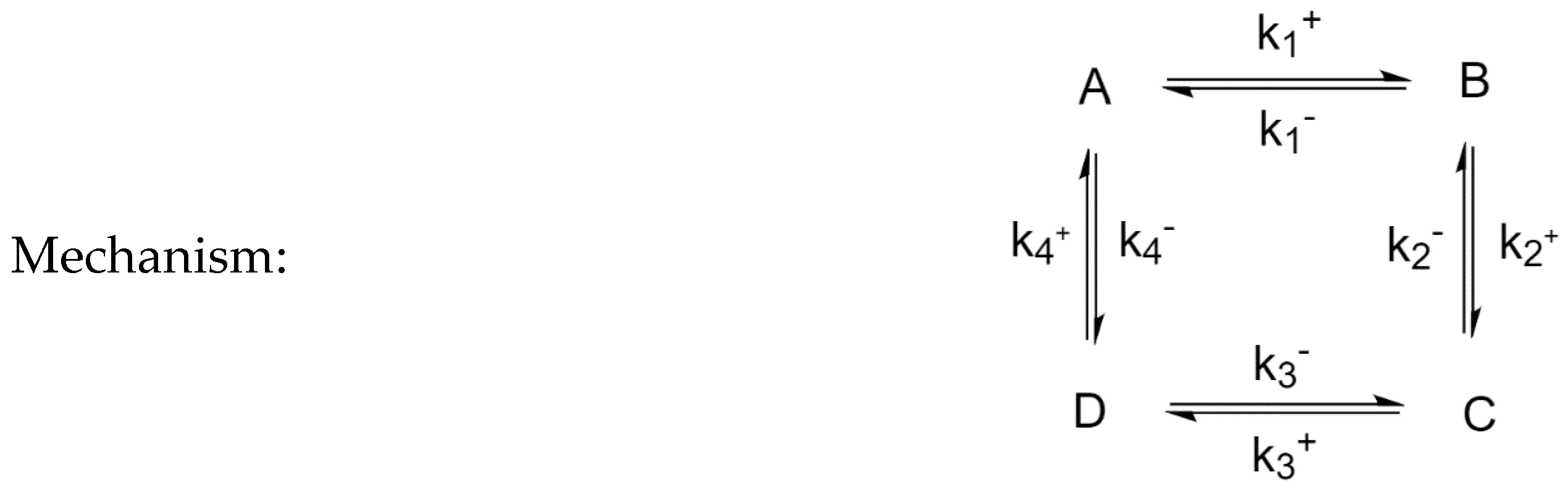{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Settings | Experiment #1 | Experiment #2 |
|---|---|---|
| Kinetic Parameters (s−1): | k1+ = 5, k1− = 4 k2+ = 12, k2− = 6 | k1+ = 16, k1− = 4 k2+ = 12, k2− = 6 |
| Perturbed species: | A, B | A, B |
| Unperturbed species: | C | C |
| Experiment Settings | Value | |
|---|---|---|
| Kinetic Parameters (s−1): | k1+ = 16 | k1− = 4 |
| k2+ = 12 | k2− = 6 | |
| k3+ = 8 | k3− = 1 | |
| Perturbed species: | A, B | |
| Unperturbed species: | C | |
| Experimental Settings | Value | |
|---|---|---|
| Kinetic parameters (s−1): | k1+ = 2 | k1− = 1 |
| k2+ = 3 | k2− = 1 | |
| k3+ = 1 | k3− = 1 | |
| Perturbed species: | A, D | |
| Unperturbed species: | B, C | |
| Experimental Settings | Values | |
|---|---|---|
| Kinetic parameters (s−1): | k1+ = 2 | k1− = 1 |
| k2+ = 3 | k2− = 1 | |
| k3+ = 1 | k3− = 1 | |
| Experiment | Perturbed Species | Unperturbed Species | Behavior |
|---|---|---|---|
| 1 | A, B | C, D | 2 extrema of [C], 1 of [D] |
| 2 | A, C | B, D | 1 extremum of [B], 1 of [D] |
| 3 | A, D | B, C | 2 extrema of [C], 1 of [B] |
| 4 | B, C | A, D | 1 extremum of [A], 1 of [D] |
| 5 | B, D | A, C | 1 extremum of [A], 1 of [C] |
| 6 | C, D | A, B | 1 extremum of [A], 1 of [B] |
| Experiment Settings | Value | |
|---|---|---|
| Kinetic parameters (s−1): | k1+ = 2 | k1− = 1 |
| k2+ = 3 | k2− = 1 | |
| k3+ = 1 | k3− = 1 | |
| k4+ = 1 | k4− = 6 | |
| Perturbed species: | A, D | |
| Unperturbed species: | B, C | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, Y.; Liu, X.; Constales, D.; Yablonsky, G.S. Perturbed and Unperturbed: Analyzing the Conservatively Perturbed Equilibrium (Linear Case). Entropy 2020, 22, 1160. https://doi.org/10.3390/e22101160
Xi Y, Liu X, Constales D, Yablonsky GS. Perturbed and Unperturbed: Analyzing the Conservatively Perturbed Equilibrium (Linear Case). Entropy. 2020; 22(10):1160. https://doi.org/10.3390/e22101160
Chicago/Turabian StyleXi, Yiming, Xinquan Liu, Denis Constales, and Gregory S. Yablonsky. 2020. "Perturbed and Unperturbed: Analyzing the Conservatively Perturbed Equilibrium (Linear Case)" Entropy 22, no. 10: 1160. https://doi.org/10.3390/e22101160
APA StyleXi, Y., Liu, X., Constales, D., & Yablonsky, G. S. (2020). Perturbed and Unperturbed: Analyzing the Conservatively Perturbed Equilibrium (Linear Case). Entropy, 22(10), 1160. https://doi.org/10.3390/e22101160