
Entropy as a Geometrical Source of Information in Biological Organizations

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
Collecting Samples
2.2. Methods
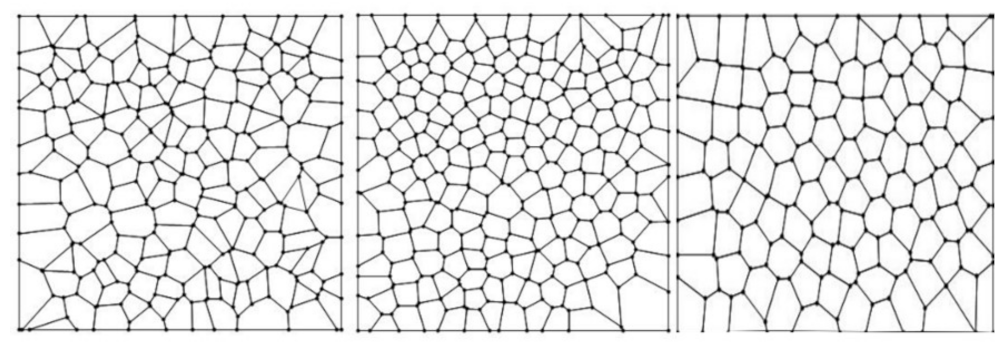
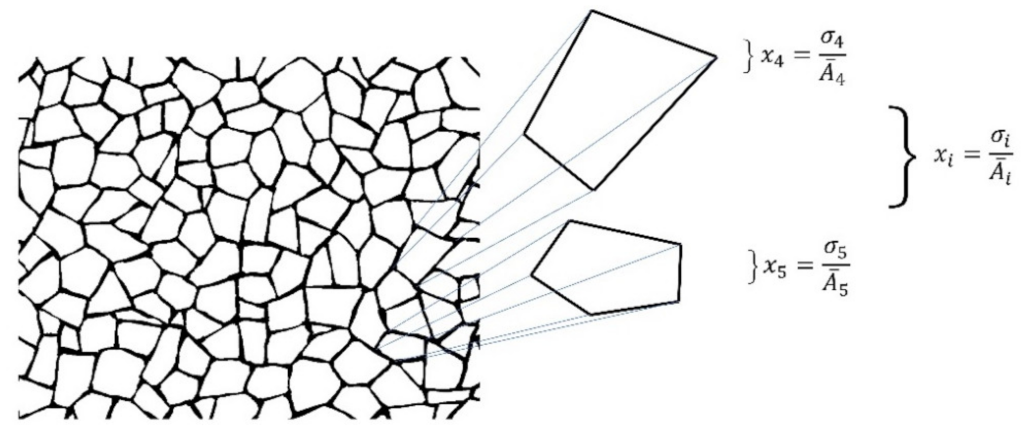
2.2.1. Mathematical Description of Shapes and Heterogeneity of Spatial Organization

3. Results
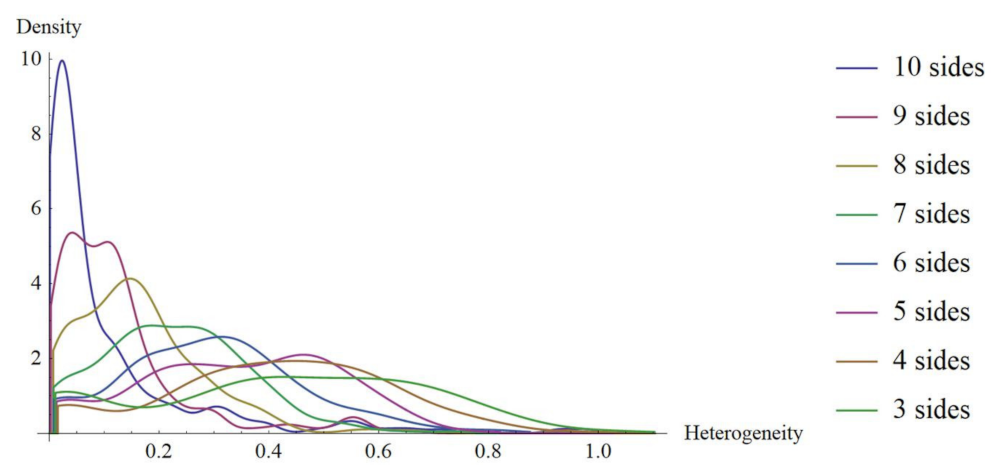
3.1. Continuous Distribution of Heterogeneity for Shapes -PDA
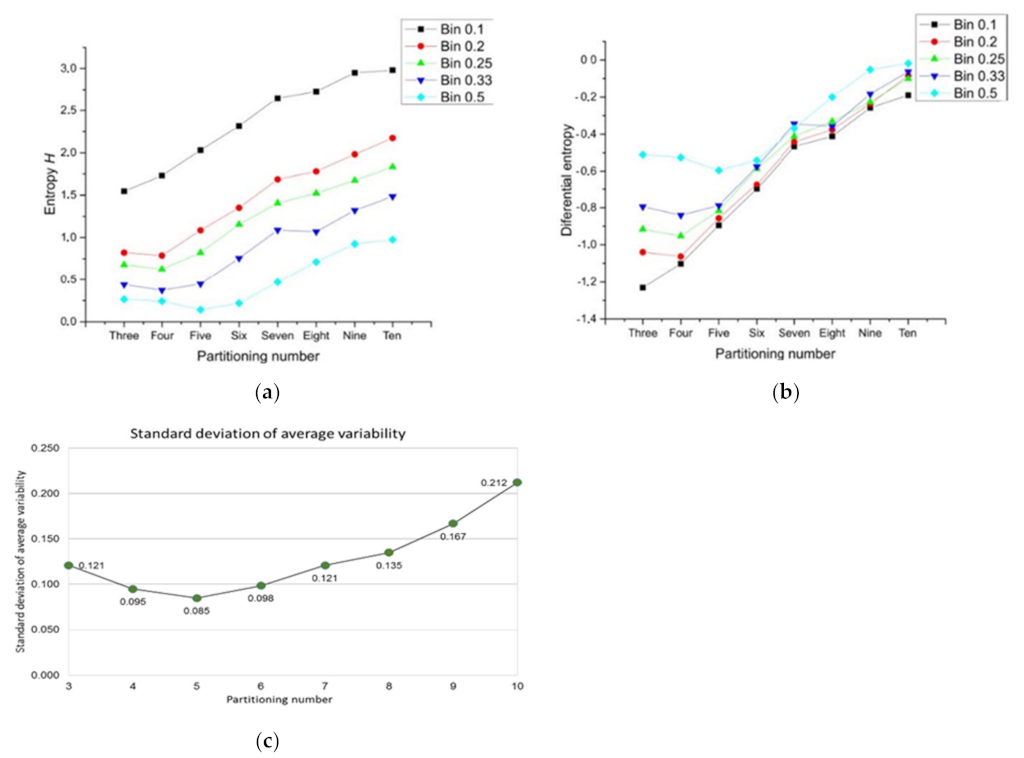
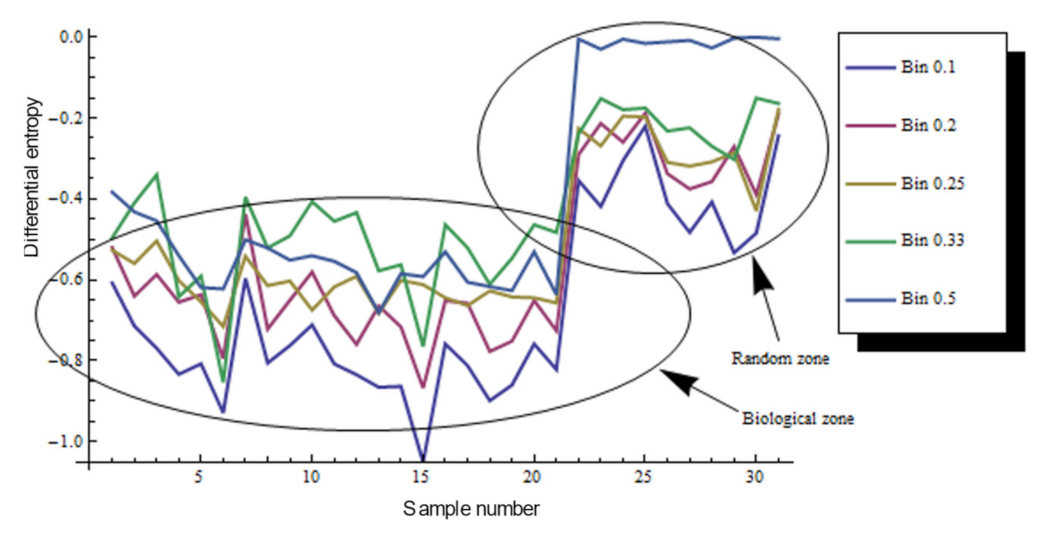
3.2. Bin Categorizations for Measuring Discrete and Continuous Entropy Using Polygons
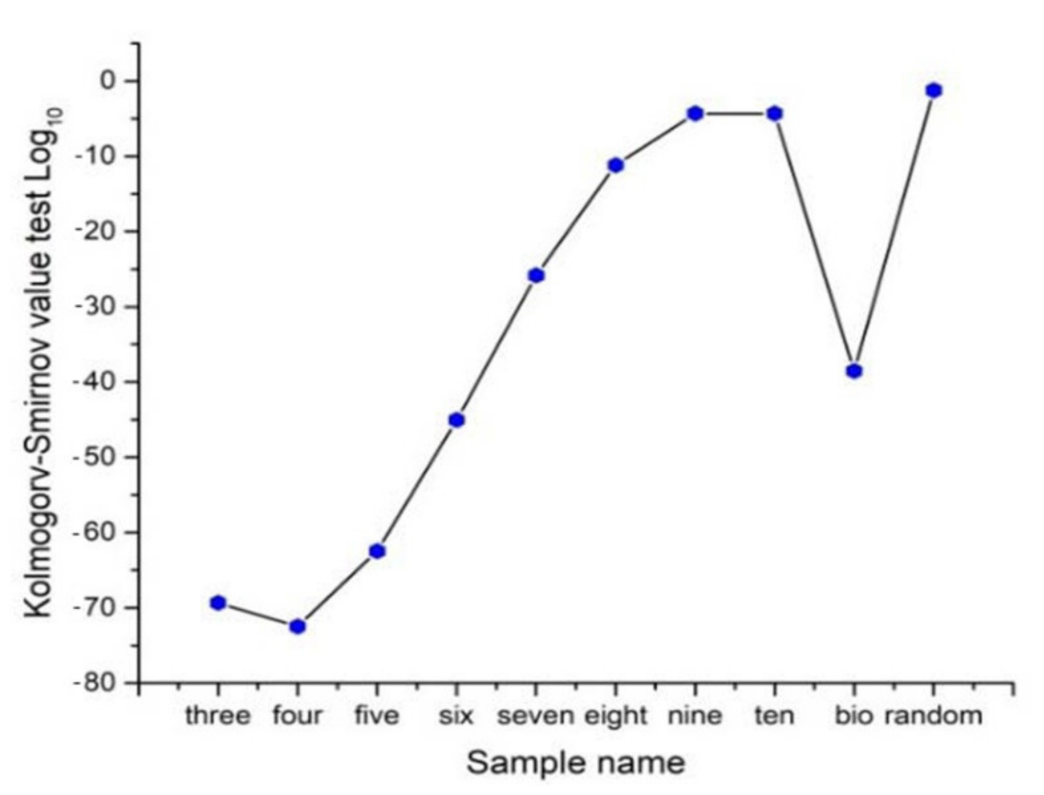
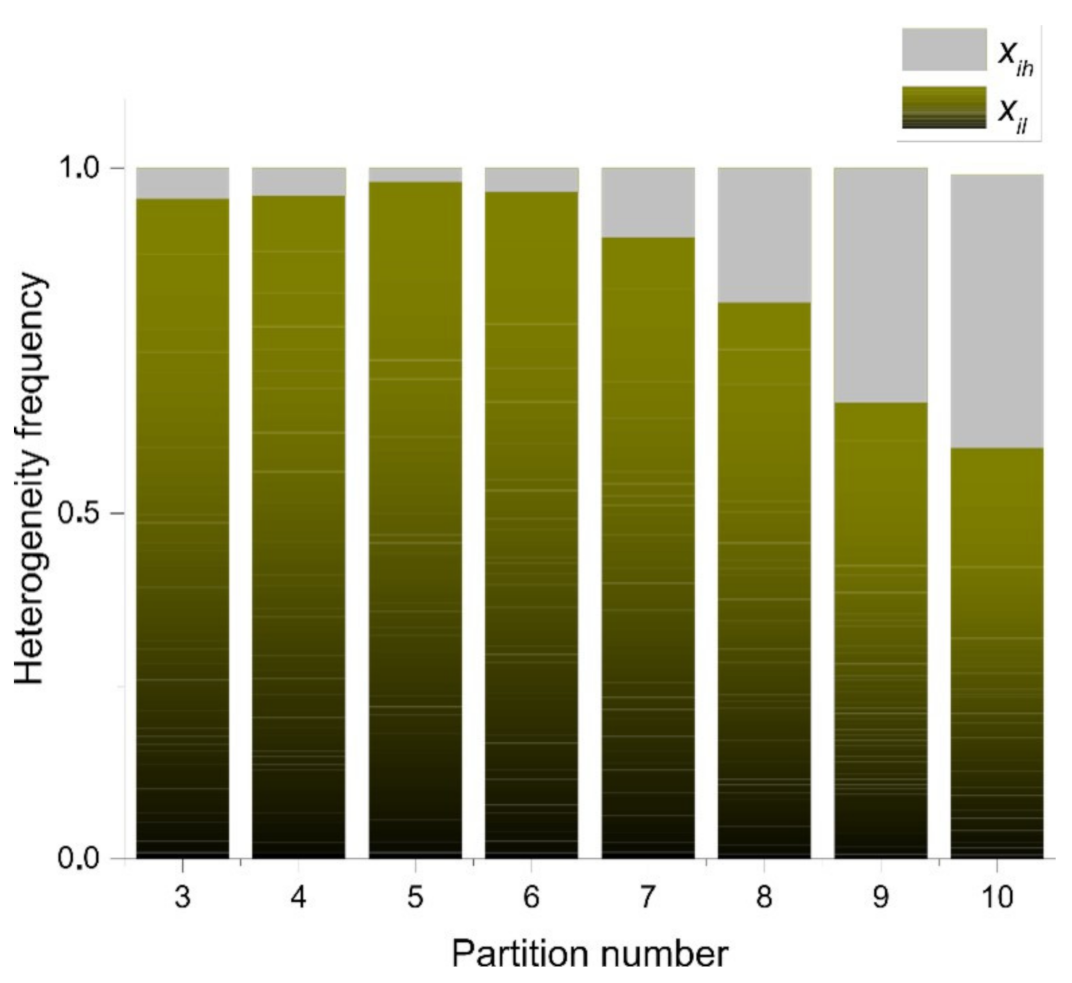
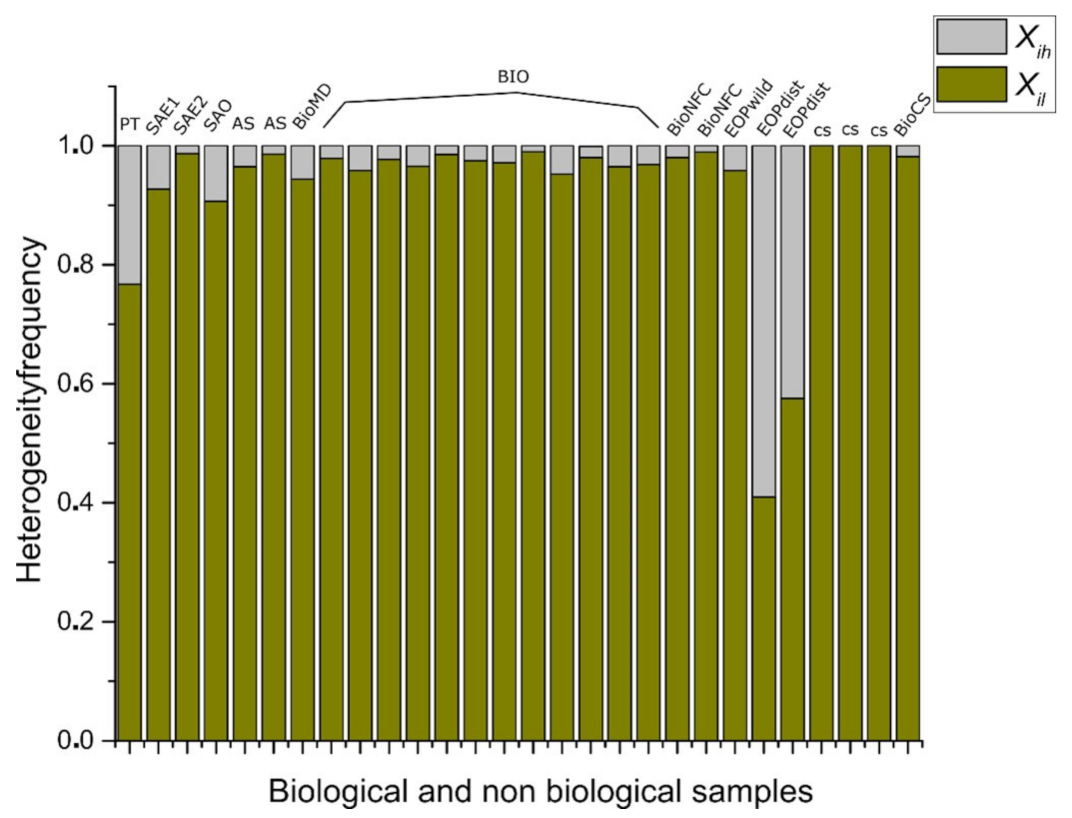
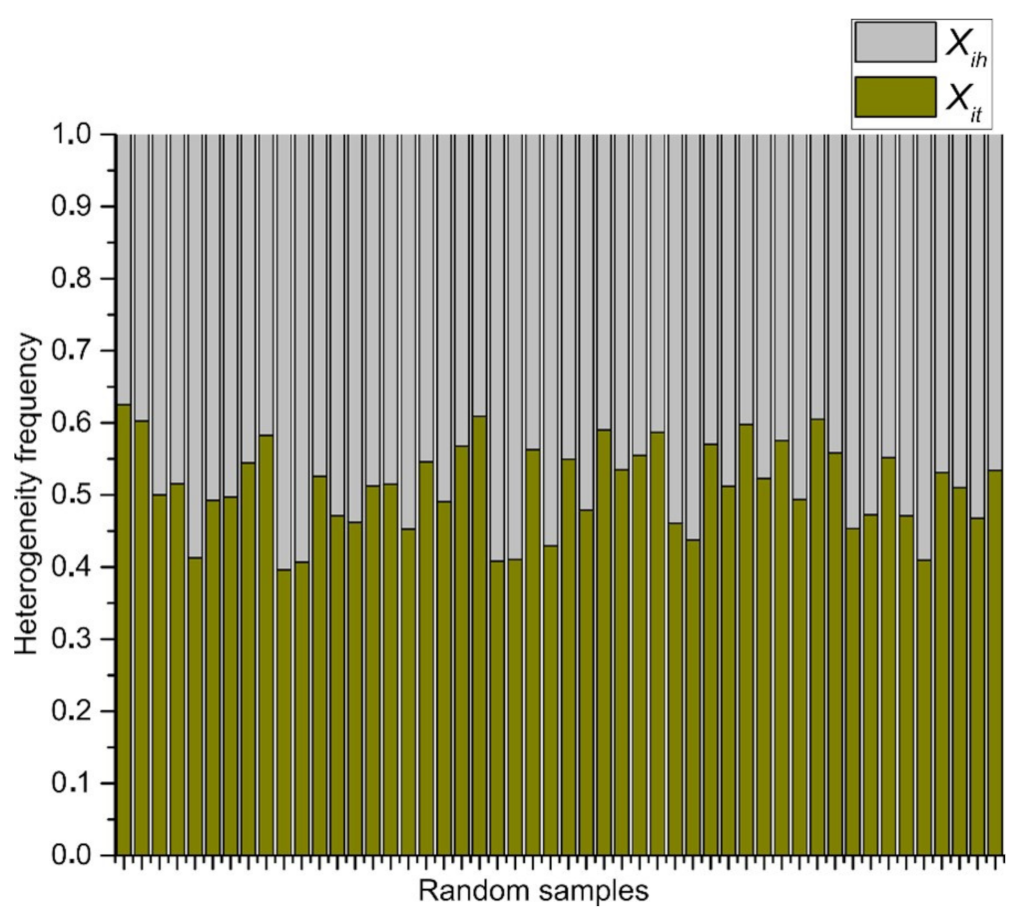
3.3. Statistical Frequency Distributions of Internal Partition in -PDA and Binary Localities in Bio, Non-Bio, and RA Samples
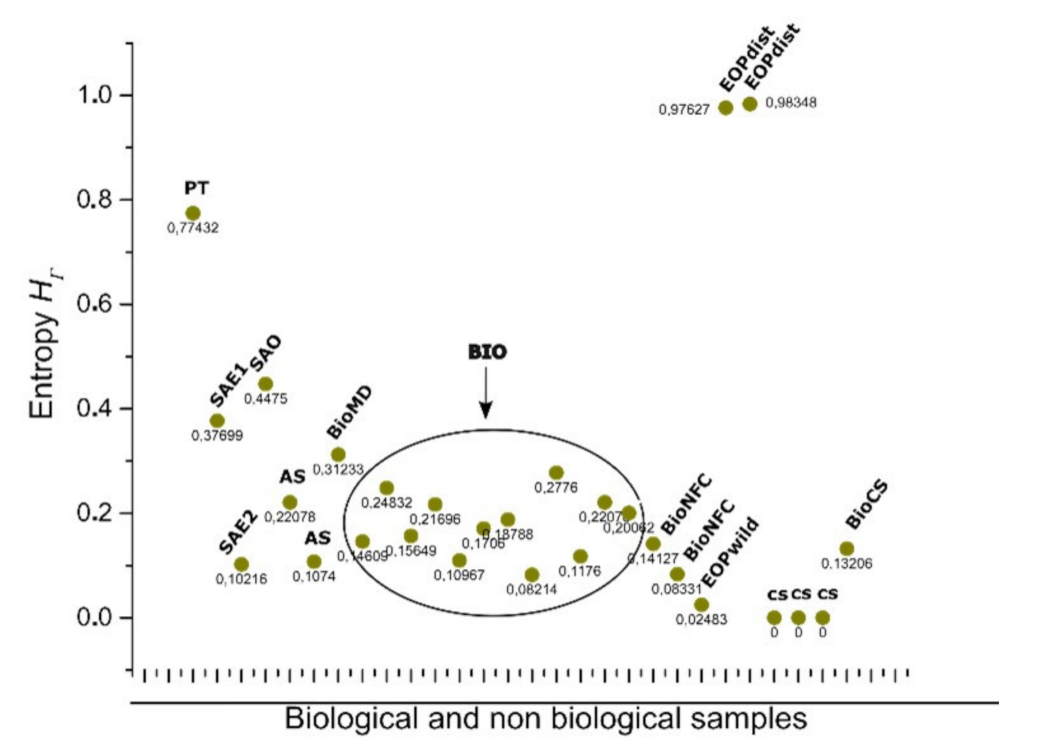
3.4. Discrete Entropy for Shapes from Bio, Non-Bio, and RA Samples Using Binarization
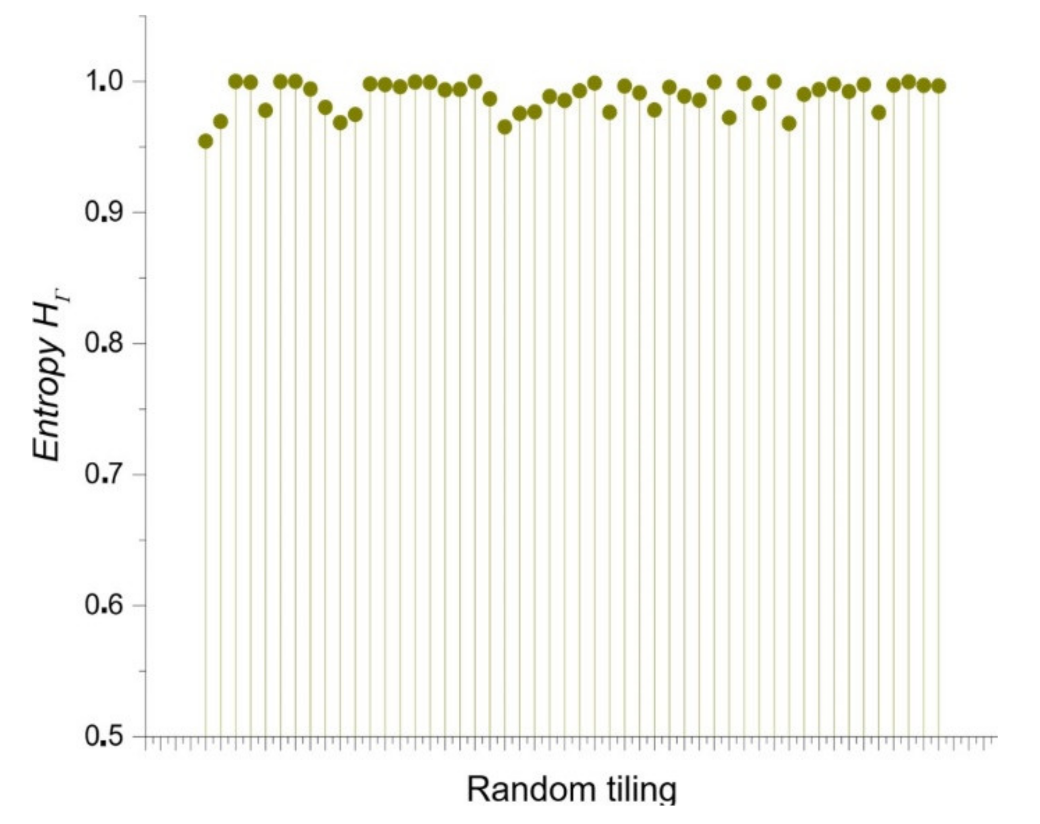
3.5. Continuous Entropy for Shapes from Bio, Non-Bio, and RA Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
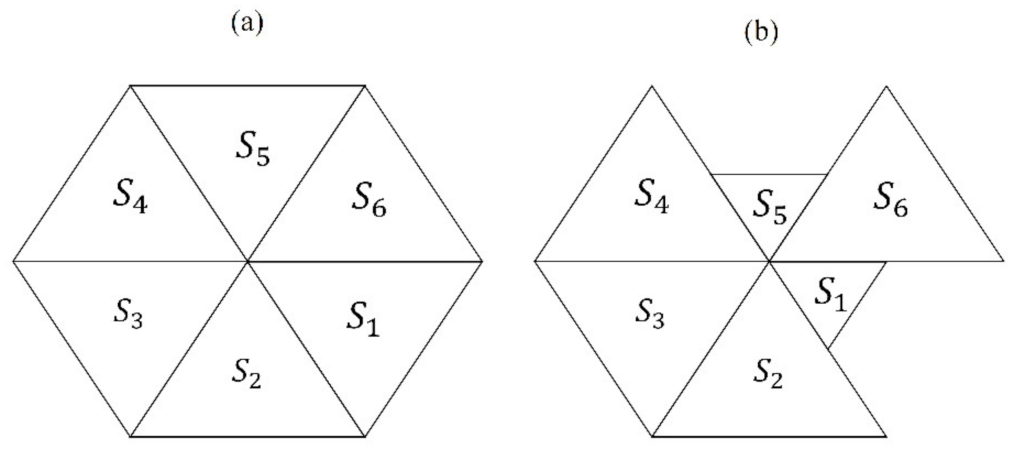
Appendix A. A Numerical Approach Using Partitions of Shapes -PDA (Planar Discrete Areas)
- a.
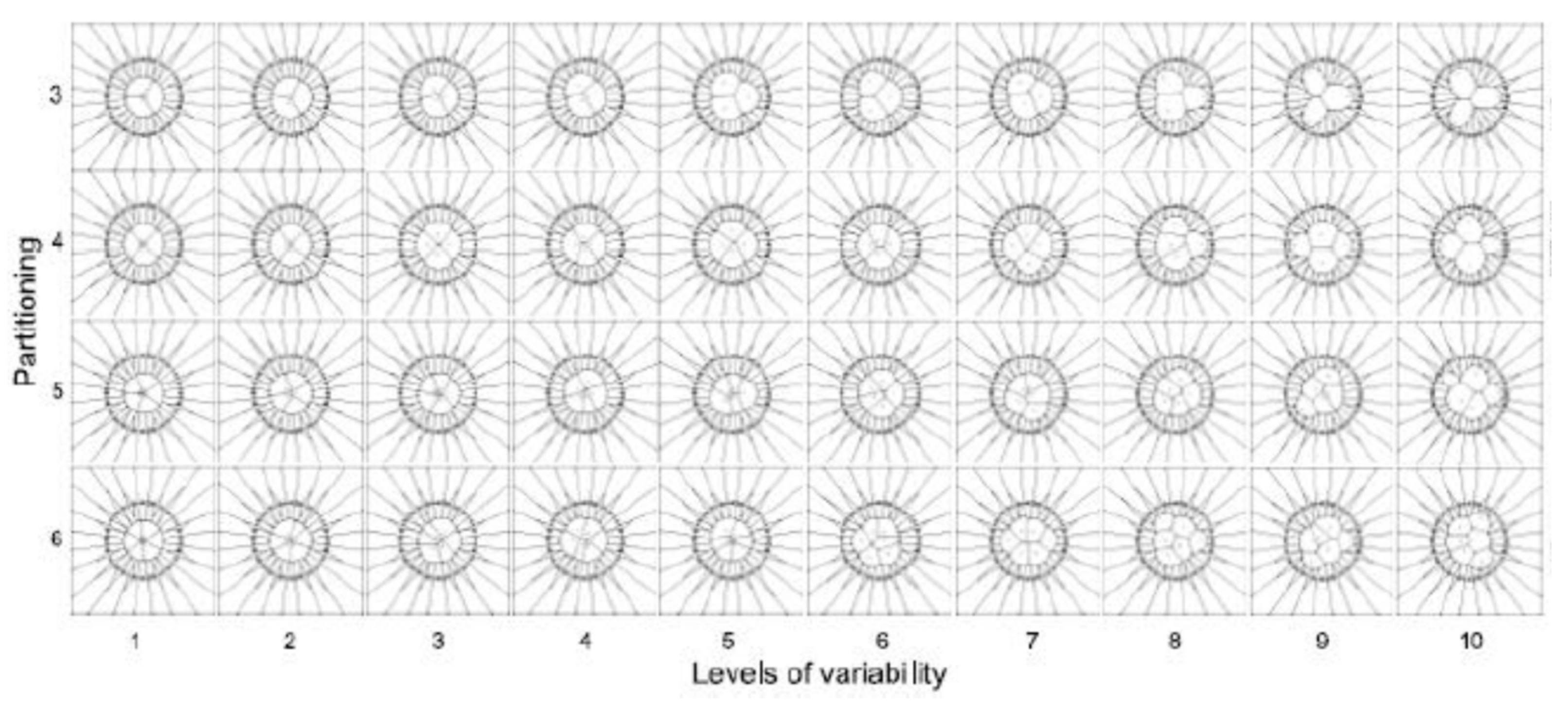
- The partitioning number (pn) defines the number of partitions inside a disc (ranging from 3 to 10): Each partition is constituted by a subset of a given number of sub-localities, such that , where is a spatial region which could be any -PDA in .
- b.
- Partition variability (pv) determines multiple levels of variability (10) inside each pn by using random points, which in turn will define the Voronoi diagrams.
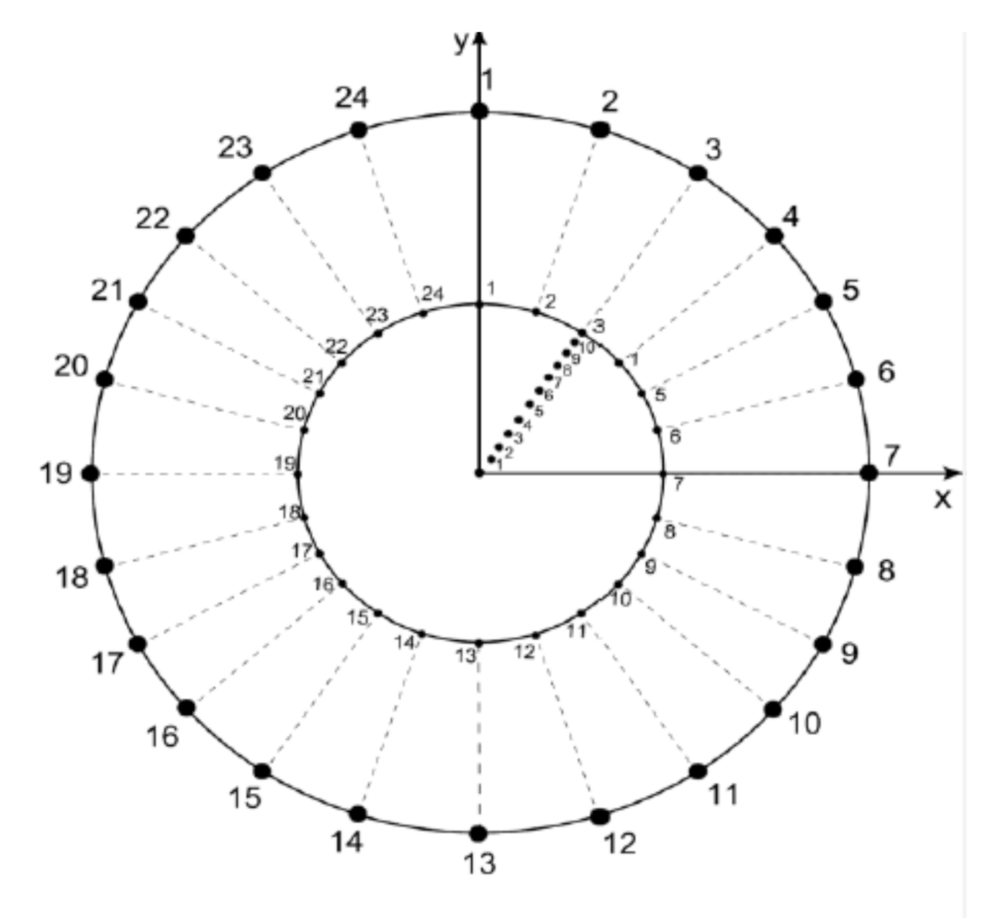
- Features of the external disc: the boundaries of the external limit are defined by 24 fixed points generated as follows: The radius of the external disk is set to r = 1 and consecutive points are separated by an angle θ/24 (where θ corresponds to 2π). Point 1 is aligned with axis y (Figure A1).
- Features of the internal disc: the boundaries of the internal limit are defined by 24 fixed points generated as follows: The radius of the internal disk is initially set to r = 0.53 ± 0.4 with 24 points consecutively separated by an angle θ/24. Point 1 is aligned with axis y. (Figure A1).
- Partitioning number (pn): once the number of partitions is defined, say n (where 3 ≤ n ≤ 10 and ), points are located in the disk at angles 2π/n ± 0.069 radians but at different radius. These radius values will define the pv, as described in the next item.
- Partition variability (pv). For each angular region defined above, 10 points are located at radius (between r = 0 and r = 10) at different positions to define different degrees of variability (diagonal points of internal disc at Figure A1). The first point (first level of variability) is at r = 1. After the second point, all of them are located at random radius between 1 to 10. Hence, each level of variability (10) is given by radii ranges except 1 which is fixed at 1 (diagonal points of internal disc); (a) 0 to 1, (b) 0 to 2, (c) 0 to 3, (d) 0 to 4, (e) 0 to 5, (f) 0 to 6, (g) 0 to 7, (h) 0 to 8, (i) 0 to 9 and (j) 0 to 10.
- Voronoi tessellations: the partition variability will define the broad spectrum of possibilities for area distribution inside discs without losing partitioning number using Voronoi tessellations.
- Area average: according to Equation (1), the average of areas requires a summation of sub-localities areas which were derived from pn with a changing variability pv.
- Data mining: once the partition areas inside discs were obtained and (1) was solved, (2) is used to obtain standard deviations of variability for each disc. In order to normalize the level of variability for each pn, an index dividing the standard deviation of partitions and the particular area average of each partition was obtained (variability average; Figure A2). There are eight particular area averages of partitions since we have a sample of 8 discs with different pn (from 3 to 10). These particular area averages are derived from a value n/(≈108.5 ± 1.5) which are n values obtained from the first level of variability (pv) at r = 1. It is important to say that the radius of the external disc (1) and the radius of the internal disc (r = 0.53 ± 0.4) was modified in order to get the particular area averages. However, despite the modification, the index between external discs and the internal ones remains constant. A sample of 20 discs to get 20 standard deviations was generated for each pn, and for each level of pv (10) giving a sample of 200 discs for each pn. An average of standard deviations (; variability average) was derived for each level of variability.
- Standard deviation. Finally, a standard deviation of all variability averages is obtained for each pn.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Partition Number | Area at Internal Disc (Level of Variability Pv1) | Particular Area Average |
|---|---|---|
| 3 | 107.2 | 35.7354 |
| 4 | 108.7 | 27.1963 |
| 5 | 109.5 | 21.9155 |
| 6 | 109.9 | 18.3248 |
| 7 | 110.1 | 15.74 |
| 8 | 110.32 | 13.7959 |
| 9 | 110.51 | 12.2794 |
| 10 | 110.605 | 11.0605 |
References
- Busiello, D.M.; Suweis, S.; Hidalgo, J.; Maritan, A. Explorability and the Origin of Network Sparsity in Living Systems. Sci. Rep. 2017, 7, 12323. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Jelassi, M.; Hazgui, H.; Ben Miled, S.; Bellamine Ben Saoud, N.; Taramasco, C. Biological Networks Entropies: Examples in Neural Memory Networks, Genetic Regulation Networks and Social Epidemic Networks. Entropy 2018, 20, 36. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, G. The Entropy of Randomized Network Ensembles. Eur. Lett. 2007, 81, 28005. [Google Scholar] [CrossRef]
- Demetrius, L.; Manke, T. Robustness and Network Evolution—an Entropic Principle. Phys. A Stat. Mech. Appl. 2005, 346, 682–696. [Google Scholar] [CrossRef]
- Cushman, S.A. Thermodynamics in Landscape Ecology: The Importance of Integrating Measurement and Modeling of Landscape Entropy. Landsc. Ecol. 2015, 30, 7–10. [Google Scholar] [CrossRef]
- Vranken, I.; Baudry, J.; Aubinet, M.; Visser, M.; Bogaert, J. A Review on the Use of Entropy in Landscape Ecology: Heterogeneity, Unpredictability, Scale Dependence and Their Links with Thermodynamics. Landsc. Ecol. 2015, 30, 51–65. [Google Scholar] [CrossRef]
- Parrott, L. Measuring Ecological Complexity. Ecol. Indic. 2010, 10, 1069–1076. [Google Scholar] [CrossRef]
- Proulx, R.; Parrott, L. Measures of Structural Complexity in Digital Images for Monitoring the Ecological Signature of an Old-Growth Forest Ecosystem. Ecol. Indic. 2008, 8, 270–284. [Google Scholar] [CrossRef]
- Frost, N.J.; Burrows, M.T.; Johnson, M.P.; Hanley, M.E.; Hawkins, S.J. Measuring Surface Complexity in Ecological Studies. Limnol. Oceanogr. Methods 2005, 3, 203–210. [Google Scholar] [CrossRef]
- Davies, P.C.W.; Rieper, E.; Tuszynski, J.A. Self-Organization and Entropy Reduction in a Living Cell. Biosystems 2013, 111, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Buskermolen, A.B.C.; Suresh, H.; Shishvan, S.S.; Vigliotti, A.; DeSimone, A.; Kurniawan, N.A.; Bouten, C.V.C.; Deshpande, V.S. Entropic Forces Drive Cellular Contact Guidance. Biophys. J. 2019, 116, 1994–2008. [Google Scholar] [CrossRef] [PubMed]
- Cabral, P.; Augusto, G.; Tewolde, M.; Araya, Y. Entropy in Urban Systems. Entropy 2013, 15, 5223–5236. [Google Scholar] [CrossRef]
- Gershenson, C.; Fernández, N. Complexity and Information: Measuring Emergence, Self-Organization, and Homeostasis at Multiple Scales. Complexity 2012, 18, 29–44. [Google Scholar] [CrossRef]
- Martínez-Berumen, H.A.; López-Torres, G.C.; Romo-Rojas, L. Developing a Method to Evaluate Entropy in Organizational Systems. Procedia Comput. Sci. 2014, 28, 389–397. [Google Scholar] [CrossRef]
- Alexander, C. The Nature of Order: An Essay on the Art of Building and the Nature of the Universe. Book 3, A Vision of a Living World; Center for Environmental Structure: Berkeley, CA, USA, 2005. [Google Scholar]
- López-Sauceda, J.; López-Ortega, J.; Laguna Sánchez, G.A.; Sandoval Gutiérrez, J.; Rojas Meza, A.P.; Aragón, J.L. Spatial Organization of Five-Fold Morphology as a Source of Geometrical Constraint in Biology. Entropy 2018, 20, 705. [Google Scholar] [CrossRef]
- Gómez-Gálvez, P.; Vicente-Munuera, P.; Tagua, A.; Forja, C.; Castro, A.M.; Letrán, M.; Valencia-Expósito, A.; Grima, C.; Bermúdez-Gallardo, M.; Serrano-Pérez-Higueras, Ó.; et al. Scutoids are a Geometrical Solution to Three-Dimensional Packing of Epithelia. Nat. Commun. 2018, 9, 2960. [Google Scholar] [CrossRef]
- Klatt, M.A.; Lovrić, J.; Chen, D.; Kapfer, S.C.; Schaller, F.M.; Schönhöfer, P.W.A.; Gardiner, B.S.; Smith, A.-S.; Schröder-Turk, G.E.; Torquato, S. Universal Hidden Order in Amorphous Cellular Geometries. Nat. Commun. 2019, 10, 811. [Google Scholar] [CrossRef]
- Rejniak, K.A.; Wang, S.E.; Bryce, N.S.; Chang, H.; Parvin, B.; Jourquin, J.; Estrada, L.; Gray, J.W.; Arteaga, C.L.; Weaver, A.M.; et al. Linking Changes in Epithelial Morphogenesis to Cancer Mutations Using Computational Modeling. PLoS Comput. Biol. 2010, 6, e1000900. [Google Scholar] [CrossRef]
- Sánchez-Gutiérrez, D.; Tozluoglu, M.; Barry, J.D.; Pascual, A.; Mao, Y.; Escudero, L.M. Fundamental Physical Cellular Constraints Drive Self-Organization of Tissues. EMBO J. 2016, 35, 77–88. [Google Scholar] [CrossRef]
- Sandersius, S.A.; Chuai, M.; Weijer, C.J.; Newman, T.J. Correlating Cell Behavior with Tissue Topology in Embryonic Epithelia. PLoS ONE 2011, 6, e18081. [Google Scholar] [CrossRef]
- Stooke-Vaughan, G.A.; Campàs, O. Physical Control of Tissue Morphogenesis across Scales. Curr. Opin. Genet. Dev. 2018, 51, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Bi, D. Multicellular Rosettes Drive Fluid-Solid Transition in Epithelial Tissues. Phys. Rev. X 2019, 9, 11029. [Google Scholar] [CrossRef]
- Bormashenko, E.; Frenkel, M.; Vilk, A.; Legchenkova, I.; Fedorets, A.A.; Aktaev, N.E.; Dombrovsky, L.A.; Nosonovsky, M. Characterization of Self-Assembled 2D Patterns with Voronoi Entropy. Entropy 2018, 20, 956. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, H. Spatial Heterogeneity Analysis: Introducing a New Form of Spatial Entropy. Entropy 2018, 20, 398. [Google Scholar] [CrossRef]
- Van Anders, G.; Klotsa, D.; Ahmed, N.K.; Engel, M.; Glotzer, S.C. Understanding Shape Entropy through Local Dense Packing. Proc. Natl. Acad. Sci. USA 2014, 111, E4812-21. [Google Scholar] [CrossRef]
- Tsuboi, A.; Ohsawa, S.; Umetsu, D.; Sando, Y.; Kuranaga, E.; Igaki, T.; Fujimoto, K. Competition for Space Is Controlled by Apoptosis-Induced Change of Local Epithelial Topology. Curr. Biol. 2018, 28, 2115–2128.e5. [Google Scholar] [CrossRef]
- Boghaert, E.; Gleghorn, J.P.; Lee, K.; Gjorevski, N.; Radisky, D.C.; Nelson, C.M. Host Epithelial Geometry Regulates Breast Cancer Cell Invasiveness. Proc. Natl. Acad. Sci. USA 2012, 109, 19632–19637. [Google Scholar] [CrossRef]
- Nicolis, G.; Prigogine, I. Self-Organization in Nonequilibrium Systems: From Dissipative Structures to Order Through Fluctuations; Wiley: Hoboken, NJ, USA, 1977; pp. 339–426. [Google Scholar]
- Klimontovich, Y.L. Turbulent Motion. The Structure of Chaos. In Turbulent Motion and the Structure of Chaos; Springer: Berlin/Heidelberg, Germany, 1991; Fundamental Theories of Physics; Volume 42, pp. 329–371. [Google Scholar] [CrossRef]
- González Valerio, M.A. Agenciamientos Materiales y Formales: Variaciones Sobre Morfologías. Agenciamientos Mater. Y Form. Var. Sobre Morfol. 2017, 19, 63–89. [Google Scholar] [CrossRef]
- Drag, M.I. Epithelium: The Lightweight, Customizable Epithelial Tissue Simulator. Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2015. [Google Scholar]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Continuous Symmetry Measures. J. Am. Chem. Soc. 1992, 114, 7843–7851. [Google Scholar] [CrossRef]
- Alemany, P.; Casanova, D.; Alvarez, S.; Dryzun, C.; Avnir, D. Continuous Symmetry Measures: A New Tool in Quantum Chemistry. Rev. Comput. Chem. 2017, 30, 289–352. [Google Scholar]
- Zabrodsky, H.; Avnir, D. Continuous Symmetry Measures. 4. Chirality. J. Am. Chem. Soc. 1995, 117, 462–473. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Symmetry as a Continuous Feature. IEEE Trans. Pattern Anal. Mach. Intell. 1995, 17, 1154–1166. [Google Scholar] [CrossRef]
- Bonjack, M.; Avnir, D. The Near-Symmetry of Protein Oligomers: NMR-Derived Structures. Sci. Rep. 2020, 10, 8367. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, M.; Fedorets, A.A.; Dombrovsky, L.A.; Nosonovsky, M.; Legchenkova, I.; Bormashenko, E. Continuous Symmetry Measure vs Voronoi Entropy of Droplet Clusters. J. Phys. Chem. C 2021, 125, 2431–2436. [Google Scholar] [CrossRef]
- Atia, L.; Bi, D.; Sharma, Y.; Mitchel, J.A.; Gweon, B.; Koehler, A.S.; DeCamp, S.J.; Lan, B.; Kim, J.H.; Hirsch, R.; et al. Geometric Constraints during Epithelial Jamming. Nat. Phys. 2018, 14, 613–620. [Google Scholar] [CrossRef]
- Gibson, W.T.; Gibson, M.C. Cell Topology, Geometry, and Morphogenesis in Proliferating Epithelia. Curr. Top. Dev. Biol. 2009, 89, 87–114. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gutiérrez, D.; Sáez, A.; Pascual, A.; Escudero, L.M. Topological Progression in Proliferating Epithelia Is Driven by a Unique Variation in Polygon Distribution. PLoS ONE 2013, 8, e79227. [Google Scholar] [CrossRef] [PubMed]
- Sáez, A.; Rivas, E.; Montero-Sánchez, A.; Paradas, C.; Acha, B.; Pascual, A.; Serrano, C.; Escudero, L.M. Quantifiable Diagnosis of Muscular Dystrophies and Neurogenic Atrophies through Network Analysis. BMC Med. 2013, 11, 77. [Google Scholar] [CrossRef]
- Escudero, L.M.; Costa, L.D.F.; Kicheva, A.; Briscoe, J.; Freeman, M.; Babu, M.M. Epithelial Organisation Revealed by a Network of Cellular Contacts. Nat. Commun. 2011, 2, 526. [Google Scholar] [CrossRef]
- Pilot, F.; Lecuit, T. Compartmentalized Morphogenesis in Epithelia: From Cell to Tissue Shape. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2005, 232, 685–694. [Google Scholar] [CrossRef]
- López-Sauceda, J.; Rueda-Contreras, M.D. A Method to Categorize 2-Dimensional Patterns Using Statistics of Spatial Organization. Evol. Bioinforma. 2017, 13, 1176934317697978. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sinclair, R. Namibian Fairy Circles and Epithelial Cells Share Emergent Geometric Order. Ecol. Complex. 2015, 22, 32–35. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Wiegand, T.; Yizhaq, H.; Hardenberg, J.; Meron, E. Adopting a Spatially Explicit Perspective to Study the Mysterious Fairy Circles of Namibia. Ecography 2015, 38, 1–11. [Google Scholar] [CrossRef]
- Contreras-Figueroa, G.; Hernandez-Sandoval, L.; Aragon-Vera, J.L. A measure of regularity for polygonal mosaics in biological systems. Theor. Biol. Med. Model. 2015, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.C.; Patel, A.B.; Nagpal, R.; Perrimon, N. The Emergence of Geometric Order in Proliferating Metazoan Epithelia. Nature 2006, 442, 1038–1041. [Google Scholar] [CrossRef]
- Nagpal, R.; Patel, A.; Gibson, M.C. Epithelial Topology. Bioessays 2008, 30, 260–266. [Google Scholar] [CrossRef]
- Patel, A.B.; Gibson, W.T.; Gibson, M.C.; Nagpal, R. Modeling and Inferring Cleavage Patterns in Proliferating Epithelia. PLOS Comput. Biol. 2009, 5, e1000412. [Google Scholar] [CrossRef]
- Stone, J.V. Information Theory: A Tutorial Introduction; Sebtel Press: LaVergne, TN, USA, 2015. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Jiao, Y.; Lau, T.; Hatzikirou, H.; Meyer-Hermann, M.; Corbo, J.C.; Torquato, S. Avian Photoreceptor Patterns Represent a Disordered Hyperuniform Solution to a Multiscale Packing Problem. Phys. Rev. E 2014, 89, 22721. [Google Scholar] [CrossRef]
- Cafaro, C.; Ali, S.A. Information Geometric Measures of Complexity with Applications to Classical and Quantum Physical Settings. Foundations 2021, 1, 45–62. [Google Scholar] [CrossRef]
- Summers, R.L. An Action Principle for Biological Systems. In Proceedings of the 10th International Conference on Mathematical Modeling in Physical Sciences (IC-MSQUARE 2021), Journal of Physics: Conference Series. Virtual, 6–9 September 2021. [Google Scholar]












| Mesh Categories | Abbreviation | Name and Number of Samples |
|---|---|---|
| - | PSP | Polygonal shape pattern (total number of samples 38) |
| - | -PDA | Planar discrete areas (8) |
| Bio | dWP | Drosophila prepupal wing discs (3) |
| Bio | dWL | Middle third instar wing discs (4) |
| Bio | BCA | Normal human biceps (2) |
| Bio | MD | Muscular dystrophy from skeletal muscles (1) |
| Bio | PSD | Pseudo stratified Drosophila disk epithelium (4) |
| Bio | NFC | Namibia fairy circles (2) |
| Bio | EOP | Ecological Oak Patterns (3) |
| Non-Bio | CS | Control simulations (5) |
| Non-Bio | SOE | Simulation out of equilibrium (1) |
| Non-Bio | SAE | Simulation at equilibrium (2) |
| Non-Bio | AS | Atrophy simulation (2) |
| Non-Bio | PT | Poisson–Voronoi tessellation (1) |
| RA | RA | Random arrangements (50) |
| Bin Width | r between Dis_E and STD_HRD | r between Dif_E and STD_HRD |
|---|---|---|
| 0.1 | 0.7215 | 0.7405 |
| 0.2 | 0.8129 | 0.8191 |
| 0.25 | 0.8161 | 0.8221 |
| 0.333 | 0.8642 | 0.8667 |
| 0.5 | 0.9311 | 0.9308 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Sauceda, J.; von Bülow, P.; Ortega-Laurel, C.; Perez-Martinez, F.; Miranda-Perkins, K.; Carrillo-González, J.G. Entropy as a Geometrical Source of Information in Biological Organizations. Entropy 2022, 24, 1390. https://doi.org/10.3390/e24101390
Lopez-Sauceda J, von Bülow P, Ortega-Laurel C, Perez-Martinez F, Miranda-Perkins K, Carrillo-González JG. Entropy as a Geometrical Source of Information in Biological Organizations. Entropy. 2022; 24(10):1390. https://doi.org/10.3390/e24101390
Chicago/Turabian StyleLopez-Sauceda, Juan, Philipp von Bülow, Carlos Ortega-Laurel, Francisco Perez-Martinez, Kalina Miranda-Perkins, and José Gerardo Carrillo-González. 2022. "Entropy as a Geometrical Source of Information in Biological Organizations" Entropy 24, no. 10: 1390. https://doi.org/10.3390/e24101390
APA StyleLopez-Sauceda, J., von Bülow, P., Ortega-Laurel, C., Perez-Martinez, F., Miranda-Perkins, K., & Carrillo-González, J. G. (2022). Entropy as a Geometrical Source of Information in Biological Organizations. Entropy, 24(10), 1390. https://doi.org/10.3390/e24101390