Effects of the Total Saponins from Rosa laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
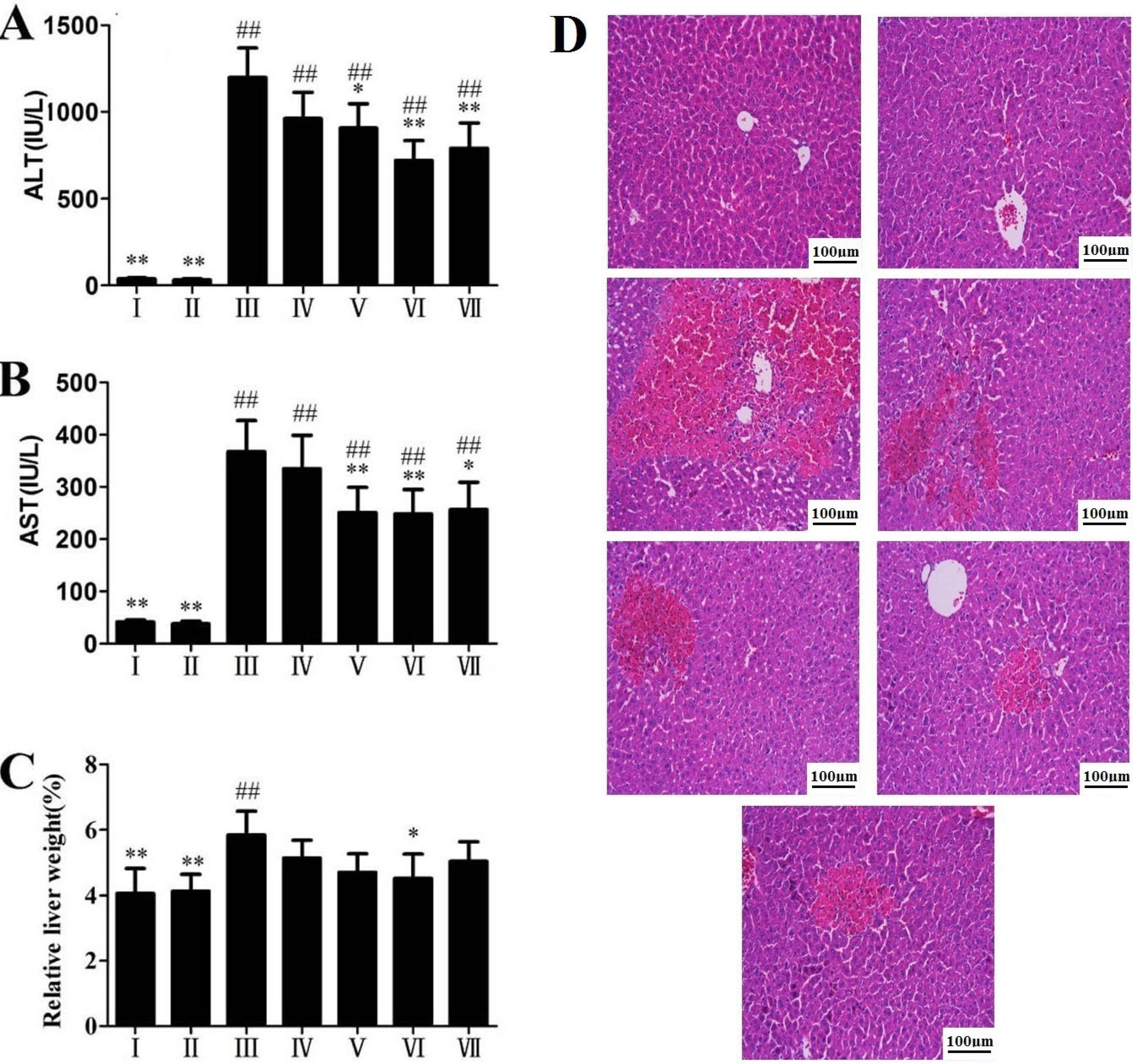
2.1. Effects of RLTS on Serum ALT, AST Activities and Liver Histopathology

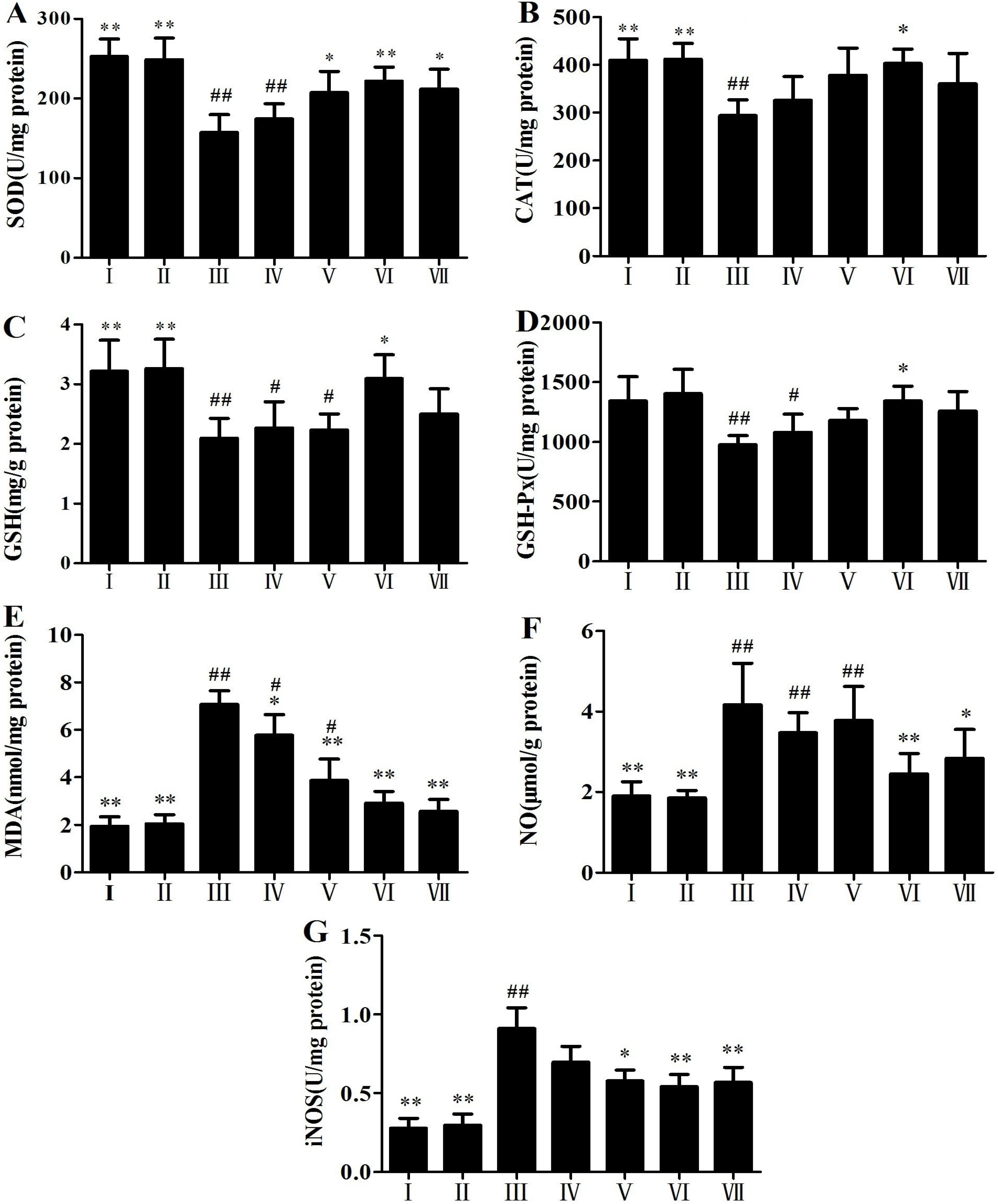
2.2. Antioxidant Activity of RLTS

2.3. Effects of RLTS on Nuclei and DNA Fragmentation Caused by APAP

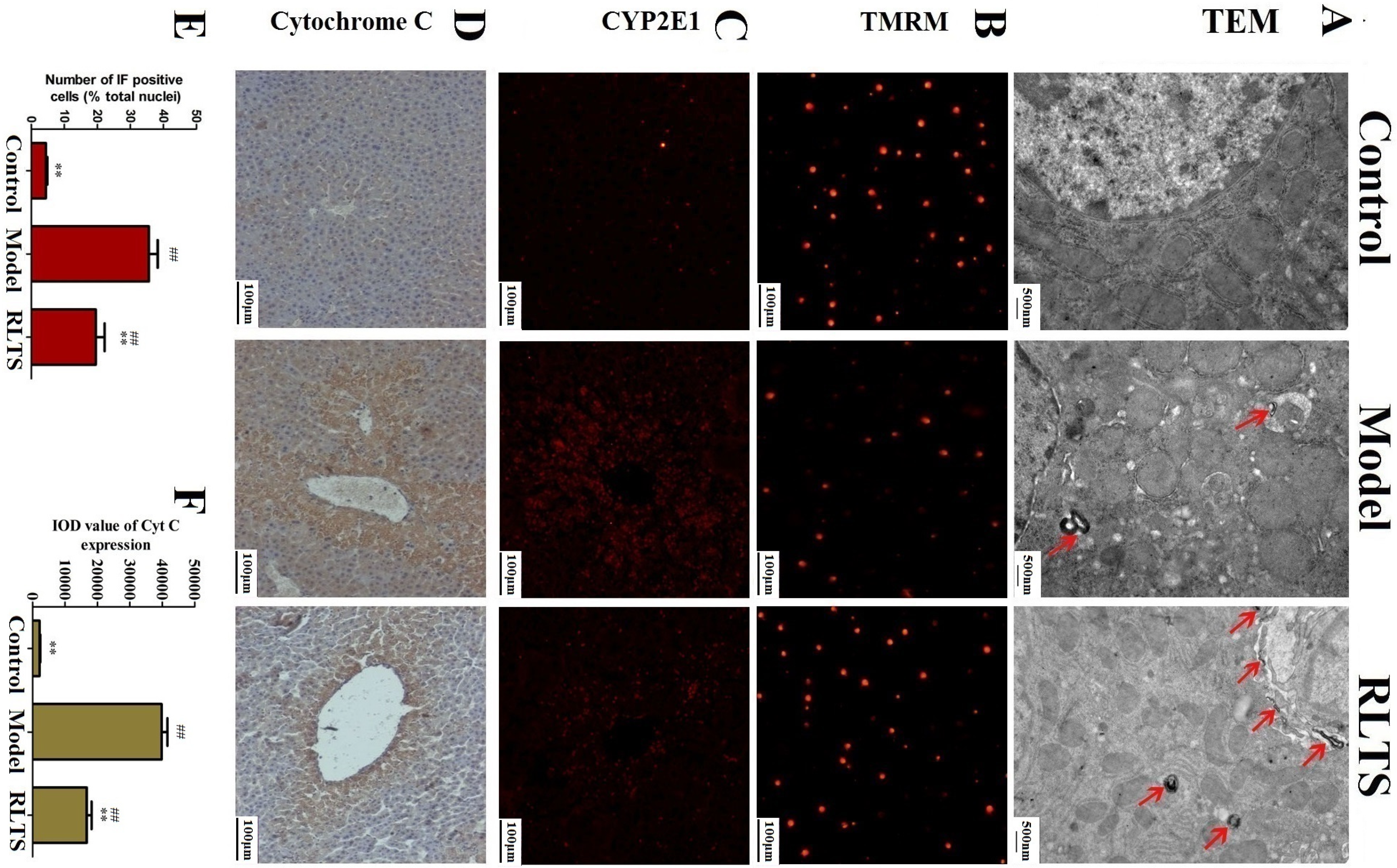
2.4. Effects of RLTS on Cellular Ultrastructure and Mitochondrial Injury
2.5. Effect of RLTS on Expressions of CYP2E1 and Cytochrome C

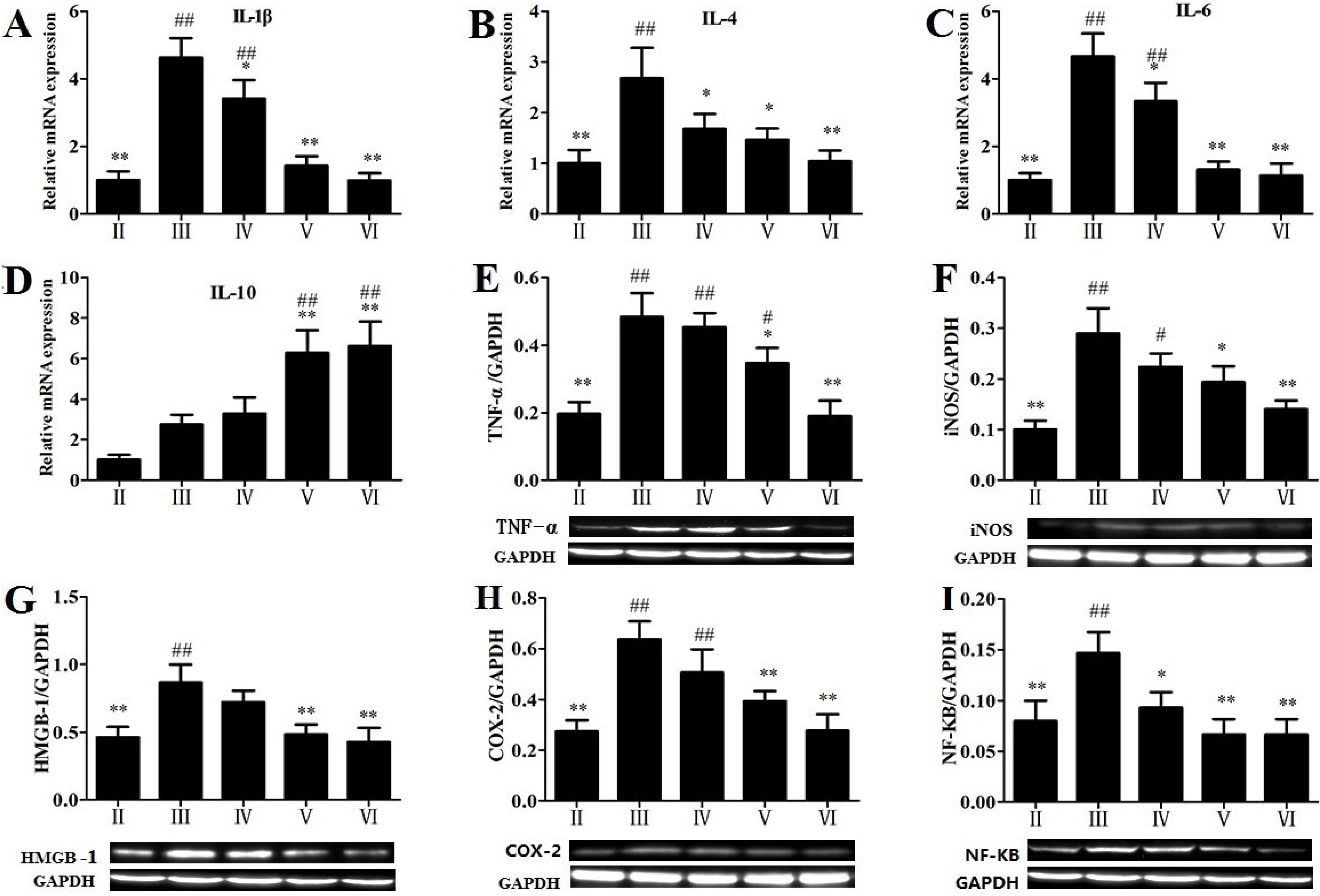
2.6. Effects of RLTS on some Molecular Expressions in Inflammatory Pathway

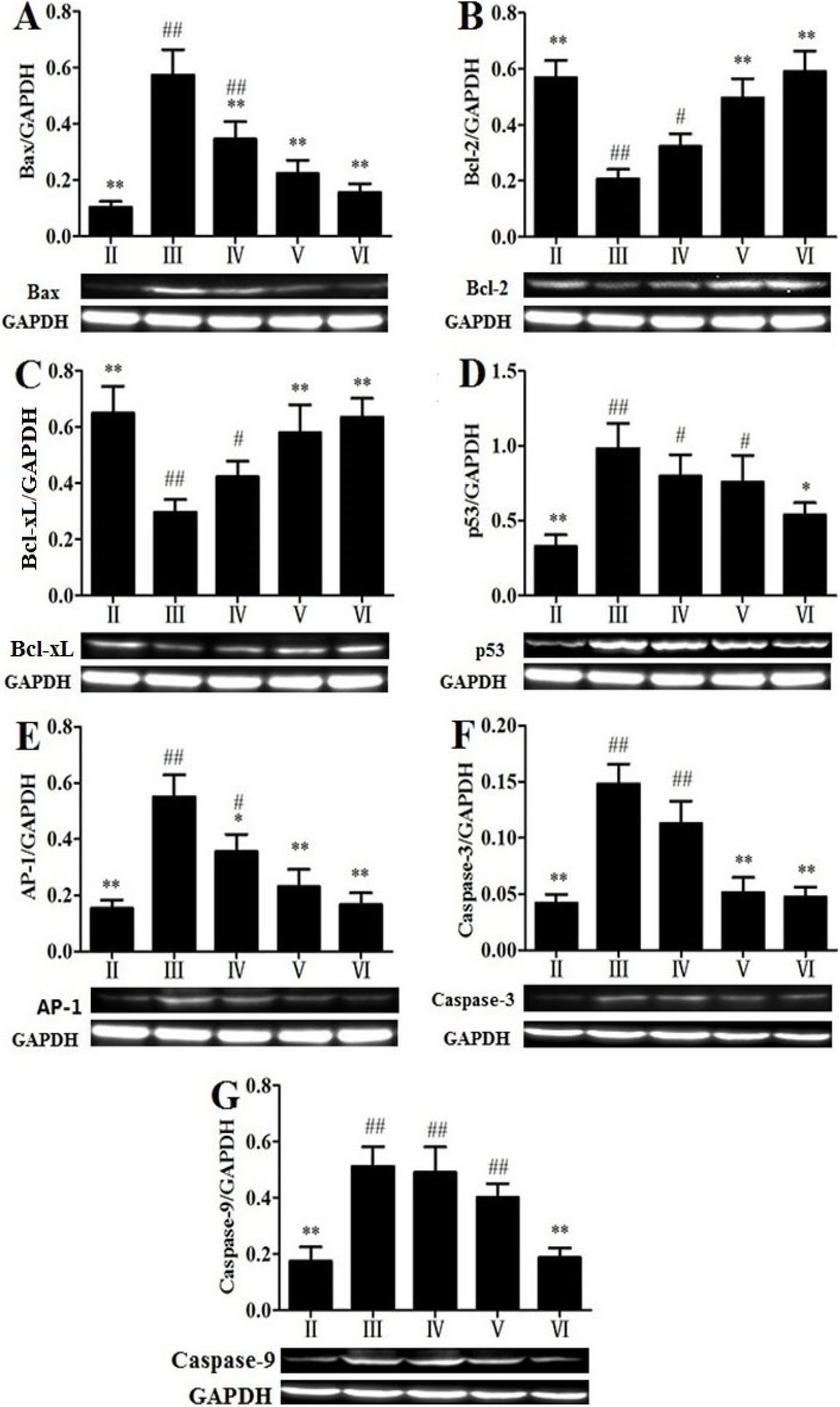
2.7. Effects of RLTS on Some Molecular Expressions Associated with Apoptosis

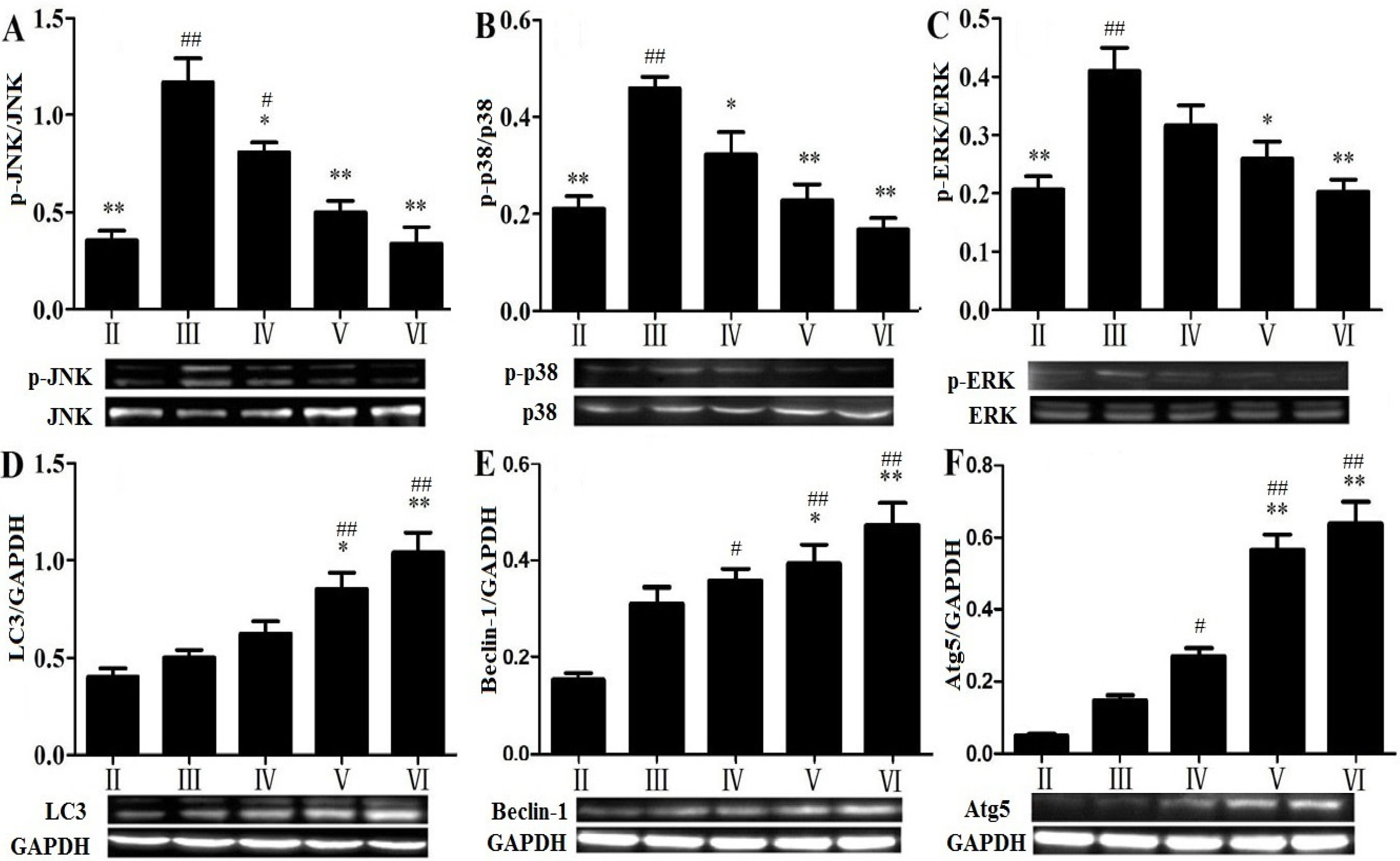
2.8. Effects of RLTS on Some Molecular Expressions in Autophagic Pathway

2.9. Discussion
3. Experimental
3.1. Plant Material and Preparation of RLTS
3.2. Chemical and Reagents
3.3. Animals
3.4. Experimental Design
3.5. Serum Biochemistry
3.6. Histopathological Examination
3.7. Determinating the Levels of Antioxidant Markers in Liver Tissue
3.8. Assay for Lipid Peroxidation Product
3.9. DAPI Staining
3.10. TUNEL Assay
3.11. Transmission Electron Microscopy (TEM) Assay
3.12. Determination of Mitochondria Membrane Potential (ΔΨm)
3.13. Immunofluorescence and Immunohistochemical Assays for CYP2E1 and Cytochrome C
3.14. Quantitative Real-time PCR
3.15. Western Blotting Assay
3.16. Statistical Analysis
4. Conclusions
Abbreviations
| RLTS | Rosa laevigata Michx fruit |
| APAP | acetaminophen |
| ALT | alanine transaminase |
| AST | aspartate transaminase |
| MDA | malondialdehyde |
| NO | nitric oxide |
| iNOS | inducible nitric oxide synthase |
| SOD | superoxide dismutase |
| CAT | catalase |
| GSH | glutathione |
| GSH-Px | glutathione peroxidase |
| TUNEL | TdT-mediated dUTP nick end labeling |
| TEM | transmission electron microscopy |
| IL-1β | interleukin 1β |
| IL-4 | interleukin |
| IL-6 | interleukin 6 |
| TNF-α | tumor necrosis factor α |
| Bax | Bcl-2 Assaciated X protein |
| HMGB-1 | High-mobility group 1 |
| COX-2 | cyclooxygenase-2 |
| NF-κB | nuclear factor kappa B |
| AP-1 | activator protein-1 |
| IL-10 | interleukin 10 |
| Bcl-2 | B-cell lymphoma-2 |
| Bcl-xL | B-cell lymphoma-extra large |
| LC3 | light chain 3 |
| Atg5 | autophagy protein 5 |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abraham, P. Oxidative stress in paracetamol-induced pathogenesis: (I)-Renal Damage. Indian J. Biochem. Biophys. 2005, 42, 59–62. [Google Scholar]
- Slitt, A.M.; Dominick, P.K.; Roberts, J.C.; Cohen, S.D. Effect of ribose cysteine pretreatment on hepatic and renal acetaminophen metabolite formation and glutathione depletion. Basic Clin. Pharmacol. Toxicol. 2005, 96, 487–494. [Google Scholar] [CrossRef]
- Jaeschke, H.; Knight, T.R.; Bajt, M.L. The role of oxidant stress and reactive nitrogen species in acetaminophen hepatotoxicity. Toxicol. Lett. 2003, 144, 279–288. [Google Scholar] [CrossRef]
- James, L.P.; Mayeux, P.R.; Hinson, J.A. Acetaminophen-induced hepatotoxicity. Drug Metab. Dispos. 2003, 31, 1499–1506. [Google Scholar] [CrossRef]
- Latchoumycandane, C.; Goh, C.W.; Ong, M.M.; Boelsterli, U.A. Mitochondrial Protection by the JNK Inhibitor Leflunomide Rescues Mice from Acetaminophen-induced Liver Injury. Hepatology 2007, 45, 412–421. [Google Scholar] [CrossRef]
- Burcham, P.C.; Harman, A.W. Acetaminophen toxicity results in site-specific mitochondrial damage in isolated mouse hepatocytes. J. Biol. Chem. 1991, 266, 5049–5054. [Google Scholar]
- Harman, A.W.; Kyle, M.E.; Serroni, A.; Farber, J.L. The killing of cultured hepatocytes by N-acetyl-p-benzoquinone imine (NAPQI) as a model of the cytotoxicity of acetaminophen. Biochem. Pharmacol. 1991, 41, 1111–1117. [Google Scholar] [CrossRef]
- Jaeschke, H.; Farhood, A.; Smith, C.W. Neutrophils contribute to ischemia/reperfusion injury in rat liver in vivo. FASEB J. 1990, 4, 3355–3339. [Google Scholar]
- Masubuchi, Y.; Suda, C.; Horie, T. Involvement of mitochondrial permeability transition in acetaminophen-induced liver injury in mice. J. Hepatol. 2005, 42, 110–116. [Google Scholar] [CrossRef]
- Adamson, G.M.; Harman, A.W. Oxidative stress in cultured hepatocytes exposed to acetaminophen. Biochem. Pharmacol. 1993, 45, 2289–2294. [Google Scholar] [CrossRef]
- Gardner, C.R.; Heck, D.E.; Yang, C.S.; Thomas, P.E.; Zhang, X.J.; DeGeorge, G.L.; Laskin, J.D.; Laskin, D.L. Role of nitric oxide in acetaminophen-induced hepatotoxicity in the rat. Hepatology 1998, 26, 748–754. [Google Scholar]
- Lemasters, J.J.; Nieminen, A.L.; Qian, T.; Trost, L.C.; Elmore, S.P.; Nishimura, Y.; Crowe, R.A.; Cascio, W.E.; Bradham, C.A.; Brenner, D.A.; et al. The mitochondrial permeability transition in cell death: A common mechanism in necrosis, apoptosis and autophagy. Biochim. Biophys. Acta 1998, 1366, 177–196. [Google Scholar] [CrossRef]
- Blazka, M.E.; Elwell, M.R.; Holladay, S.D.; Wilson, R.E.; Luster, M.I. Histopathology of acetaminophen-induced liver changes: Role of interleukin 1 alpha and tumor necrosis factor alpha. Toxicol. Pathol. 1996, 24, 181–189. [Google Scholar] [CrossRef]
- Levine, B. Eating oneself and uninvited guests: Autophagy-related pathways in cellular defense. Cell 2005, 120, 159–162. [Google Scholar]
- Sabiba, E.P.; Rasool, M.; Vedi, M.; Navaneethan, D.; Ravichander, M.; Parthasarathy, P.; Thella, S.R. Hepatoprotective and antioxidant potential of Withania somnifera against paracetamol-induced liver damage in rats. Int. J. Pharm. Pharm. Sci. 2013, 5, 648–651. [Google Scholar]
- Bajt, M.L.; Knight, T.R.; Lemasters, J.J.; Jaeschke, H. Acetaminophen-induced oxidant stress and cell injury in cultured mouse hepatocytes: Protection by N-acetyl cysteine. Toxicol. Sci. 2004, 80, 343–349. [Google Scholar] [CrossRef]
- Smilkstein, M.J.; Knapp, G.L.; Kulig, K.W.; Rumack, B.H. Efficacy of oral N– acetylcysteine in the treatment of acetaminophen overdose. Analysis of the national multicenter study (1976 to 1985). N. Engl. J. Med. 1988, 319, 1557–1562. [Google Scholar] [CrossRef]
- Zhao, X.; Cong, X.; Zheng, L.; Xu, L.; Yin, L.; Peng, J. Dioscin, a natural steroid saponin, shows remarkable protective effect against acetaminophen-induced liver damage in vitro and in vivo. Toxicol. Lett. 2012, 214, 69–80. [Google Scholar]
- Muriel, P.; Garciapina, T.; Perez-Alvarez, V.; Mourelle, M. Silymarin protects against paracetamol-induced lipid peroxidation and liver damage. J. Appl. Toxicol. 1992, 12, 439–442. [Google Scholar] [CrossRef]
- Kim, Y.W.; Ki, S.H.; Lee, J.R.; Lee, S.J.; Kim, C.W.; Kim, S.C.; Kim, S.G. Liquiritigen -in, an aglycone of liquiritin in Glycyrrhizae radix, prevents acute liver injuries in rats induced by acetaminophen with or without buthionine sulfoximine. Chem. Biol. Interact. 2006, 161, 125–138. [Google Scholar] [CrossRef]
- Ghosh, J.; Das, J.; Manna, P.; Sil, P.C. Arjunolic acid, a triterpenoid saponin, prevents acetaminophen (APAP)-induced liver and hepatocyte injury via the inhibition of APAP bioactivation and JNK-mediated mitochondrial protection. Free Radic. Biol. Med. 2010, 48, 535–553. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Nie, L.W.; Wu, B.J.; Yang, Y.; Zhao, S.S.; Jin, T. Hypolipedemic activity of the polysaccharose from Rosa laevigata Michx fruit. Chin. J. Public. Health 2004, 20, 829–830. [Google Scholar]
- Zou, H.T.; Yang, Y.; Chen, S.J. The resource values and prospective of Rosa laevigata Michx fruits as beverage. Technol. Food Ind. 2006, 27, 193–195. [Google Scholar]
- Zhao, Y.T.; Guo, X.M.; Li, F.Z. Antioxidative activity of polysaccharide from Rosa laevigata Michx. J. Biol. 2003, 20, 23–24. [Google Scholar]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becke, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Dong, D.S.; Zhang, S.; Yin, L.H.; Tang, X.Q.; Xu, Y.W.; Han, X.; Qi, Y.; Peng, J.Y. Protective effects of the total saponins from Rosa laevigata Michx fruit against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 62, 120–130. [Google Scholar] [CrossRef]
- Thomas, S.H. Paracetamol (acetaminophen) poisoning. Pharmacol. Ther. 1993, 60, 91–120. [Google Scholar] [CrossRef]
- Kass, G.E. Mitochondrial involvement in drug-induced hepatic injury. Chem. Biol. Interact. 2006, 163, 145–159. [Google Scholar] [CrossRef]
- Hinson, J.A.; Roberts, D.W.; James, L.P. Mechanisms of Acetaminophen-Induced Liver Necrosis. Handb. Exp. Pharmacol. 2010, 196, 369–405. [Google Scholar] [CrossRef]
- Raucy, J.L.; Lasker, J.M.; Lieber, C.S.; Black, M. Acetaminophen activation by human liver cytochromes P450IIE1 and P450IA2. Arch. Biochem. Biophys. 1989, 271, 270–283. [Google Scholar] [CrossRef]
- Ishida, Y.; Kondo, T.; Tsuneyama, K.; Lu, P.; Takayasu, T.; Mukaida, N. The pathogenic roles of tumor necrosis factor receptor p55 in acetaminophen-induced liver injury in mice. J. Leukoc. Biol. 2004, 75, 59–67. [Google Scholar]
- Aldaba-Muruato, L.R.; Moreno, M.G.; Shibayama, S.; Tsutsumi, V.; Muriel, P. Protective effects of allopurinol against acute liver damage and cirrhosis induced by carbon tetrachloride: Modulation of NF-κB, cytokine production and oxidative stress. Biochim. Biophys. Acta 2012, 1820, 65–75. [Google Scholar] [CrossRef]
- Taki-Eldin, A.; Zhou, L.; Xie, H.Y.; Chen, K.J.; Yong He, D.Y.; Zheng, S.S. Triiodothyronine attenuates hepatic ischemia/reperfusion injury in a partial hepatectomy model through inhibition of proinflammatory cytokines, transcription factors, and adhesion molecules. J. Surg. Res. 2012, 178, 646–656. [Google Scholar] [CrossRef]
- Watanabe, T.; Kubota, S.; Nagaya, M.; Ozaki, S.; Nagafuchi, H.; Akashi, K.; Taira, Y.; Tsukikawa, S.; Oowada, S.; Nakano, S. The Role of HMGB-1 on the Development of Necrosis During Hepatic Ischemia and Hepatic Ischemia/Reperfusion Injury in Mice. J. Surg. Res. 2005, 124, 59–66. [Google Scholar] [CrossRef]
- Hu, K.Q. Cyclooxygenase 2 (COX2)-prostanoid pathway and liver diseases. Prostaglandins Leukot. Essent. Fatty Acids 2003, 69, 329–337. [Google Scholar] [CrossRef]
- Bourdi, M.; Masubuchi, Y.; Reilly, T.P.; Amouzadeh, H.R.; Martin, J.L.; George, J.W.; Shah, A.G.; Pohl, L.R. Protection against acetaminophen-induced liver injury and lethality by interleukin 10: Role of inducible nitric oxide synthase. Hepatology 2002, 35, 289–298. [Google Scholar] [CrossRef]
- Dhingra, S.; Sharma, A.K.; Arora, R.C.; Slezak, J.; Singal, P.K. IL-10 attenuates TNF-alpha-induced NF kappaB pathway activation and cardiomyocyte apoptosis. Cardiovasc. Res. 2009, 82, 59–66. [Google Scholar] [CrossRef]
- Ray, S.D.; Mumaw, V.R.; Raje, R.R.; Fariss, M.W. Protection of acetaminophen-induced hepatocellular apoptosis and necrosis by cholesteryl hemisuccinate pretreatment. J. Pharmacol. Exp. Ther. 1996, 279, 1470–1483. [Google Scholar]
- Ferret, P.J.; Hammoud, R.; Tulliez, M.; Tran, A.; Trebeden, H.; Jaffray, P.; Malassagne, B.; Calmus, Y.; Weill, B.; Batteux, F. Detoxification of reactive oxygen species by a nonpeptidyl mimic of superoxide dismutase cures acetaminophen-induced acute liver failure in the mouse. Hepatology 2001, 33, 1173–1180. [Google Scholar] [CrossRef]
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta 1998, 1366, 139–149. [Google Scholar] [CrossRef]
- Ding, W.X.; Nam, O.C. Role of oxidative stress and mitochondrial changes in cyanobacterial- induced apoptosis and hepatotoxicity. FEMS Microbiol. Lett. 2003, 220, 1–7. [Google Scholar] [CrossRef]
- Weng, S.Y.; Yang, C.Y.; Li, C.C.; Sun, T.P.; Tung, S.Y.; Yen, J.Y. Synergism between p53 and Mcl-1 in protecting from hepatic injury, fibrosis and cancer. J. Hepatol. 2011, 54, 685–694. [Google Scholar] [CrossRef]
- Kim, H.Y.; Park, J.; Lee, K.H.; Lee, D.U.; Kwak, J.H.; Kim, Y.S.; Lee, S.M. Ferulic acid protects against carbon tetrachloride-induced liver injury in mice. Toxicology 2011, 282, 104–111. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Pal, P.B.; Sil, P.C. A 35 kD Phyllanthus niruri protein modulates iron mediated oxidative impairment to hepatocytes via the inhibition of ERKs, p38 MAPKs and activation of PI3k/Akt pathway. Food Chem. Toxicol. 2013, 56, 119–130. [Google Scholar] [CrossRef]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 Antiapoptotic Proteins Inhibit Beclin 1-Dependent Autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery for self-eating. Cell Death Differ. 2005, 12, 1542–1552. [Google Scholar] [CrossRef]
- Ohsumi, Y. Molecular dissection of autophagy: Two ubiquitin-like systems. Nat. Rev. Mol. Cell Biol. 2001, 2, 211–216. [Google Scholar] [CrossRef]
- Lavallard, V.J.; Meijer, A.J.; Codogno, P.; Gual, P. Autophagy, signaling and obesity. Pharmacol. Res. 2012, 66, 513–525. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, B.N.; Han, X.; Xu, L.N.; Qi, Y.; Yin, L.H.; Xu, Y.W.; Zhao, Y.Y.; Liu, K.X.; Peng, J.Y. Protection of the flavonoid fraction from Rosa laevigata Michx fruit against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 55, 60–69. [Google Scholar] [CrossRef]
- Jia, Y.N.; Ji, L.; Zhang, S.; Xu, L.N.; Yin, L.H.; Li, L.; Zhao, Y.Y.; Peng, J.Y. Total flavonoids from Rosa Laevigata Michx fruit attenuates hydrogen peroxide induced injury in human umbilical vein endothelial cells. Food Chem. Toxicol. 2012, 50, 3133–3141. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dong, D.; Xu, L.; Han, X.; Qi, Y.; Xu, Y.; Yin, L.; Liu, K.; Peng, J. Effects of the Total Saponins from Rosa laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis. Molecules 2014, 19, 7189-7206. https://doi.org/10.3390/molecules19067189
Dong D, Xu L, Han X, Qi Y, Xu Y, Yin L, Liu K, Peng J. Effects of the Total Saponins from Rosa laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis. Molecules. 2014; 19(6):7189-7206. https://doi.org/10.3390/molecules19067189
Chicago/Turabian StyleDong, Deshi, Lina Xu, Xu Han, Yan Qi, Youwei Xu, Lianhong Yin, Kexin Liu, and Jinyong Peng. 2014. "Effects of the Total Saponins from Rosa laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis" Molecules 19, no. 6: 7189-7206. https://doi.org/10.3390/molecules19067189
APA StyleDong, D., Xu, L., Han, X., Qi, Y., Xu, Y., Yin, L., Liu, K., & Peng, J. (2014). Effects of the Total Saponins from Rosa laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis. Molecules, 19(6), 7189-7206. https://doi.org/10.3390/molecules19067189