Partial Characterization of an Enzymatic Extract from Bentong Ginger (Zingiber officinale var. Bentong)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction of Ginger Protease (BGP) and Effect of Stabilizers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day of Storage | Enzyme Activity (unit/mL) of Different Stabilizers at 4 °C | |||
|---|---|---|---|---|
| Control | 0.2% Ascorbic Acid | 0.2% Ascorbic Acid + 5 mM EDTA | 10 mM Cysteine + 5 mM EDTA | |
| 0 | 11.7 ± 0.2 Aa | 25.9 ± 0.4 Ba | 30.7 ± 0.5 Ca | 32.6 ± 0.4 Ca |
| 1 | 5.0 ± 0.2 Ab | 23.8 ± 0.2 Ba | 29.0 ± 0.1 Cb | 31.9 ± 0.5 Da |
| 2 | 1.6 ± 0.2 Ac | 20.8 ± 0.4 Bb | 28.8 ± 0.3 Cb | 31.4 ± 0.2 Db |
| 3 | 0.7 ± 0.2 Ac | 19.4 ± 0.3 Bc | 28.4 ± 0.3 Cb | 30.9 ± 0.4 Db |
| 4 | 0.3 ± 0.1 Ac | 17.4 ± 0.3 Bd | 27.1 ± 0.5 Cc | 29.3 ± 0.2 Dc |

2.2. Characterization of Ginger Protease
2.2.1. Determination of Protease Type
2.2.2. Optimum Temperature

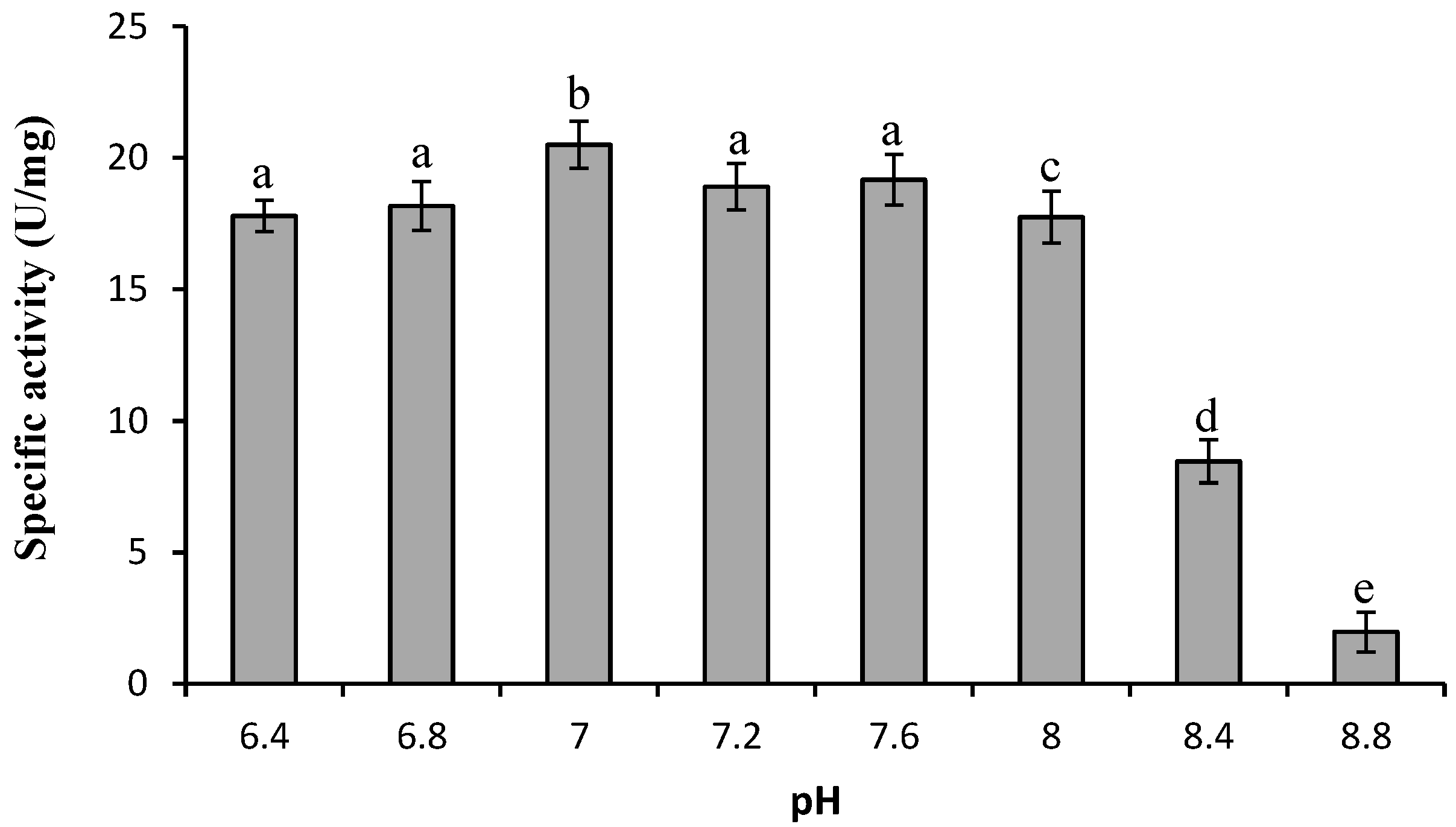
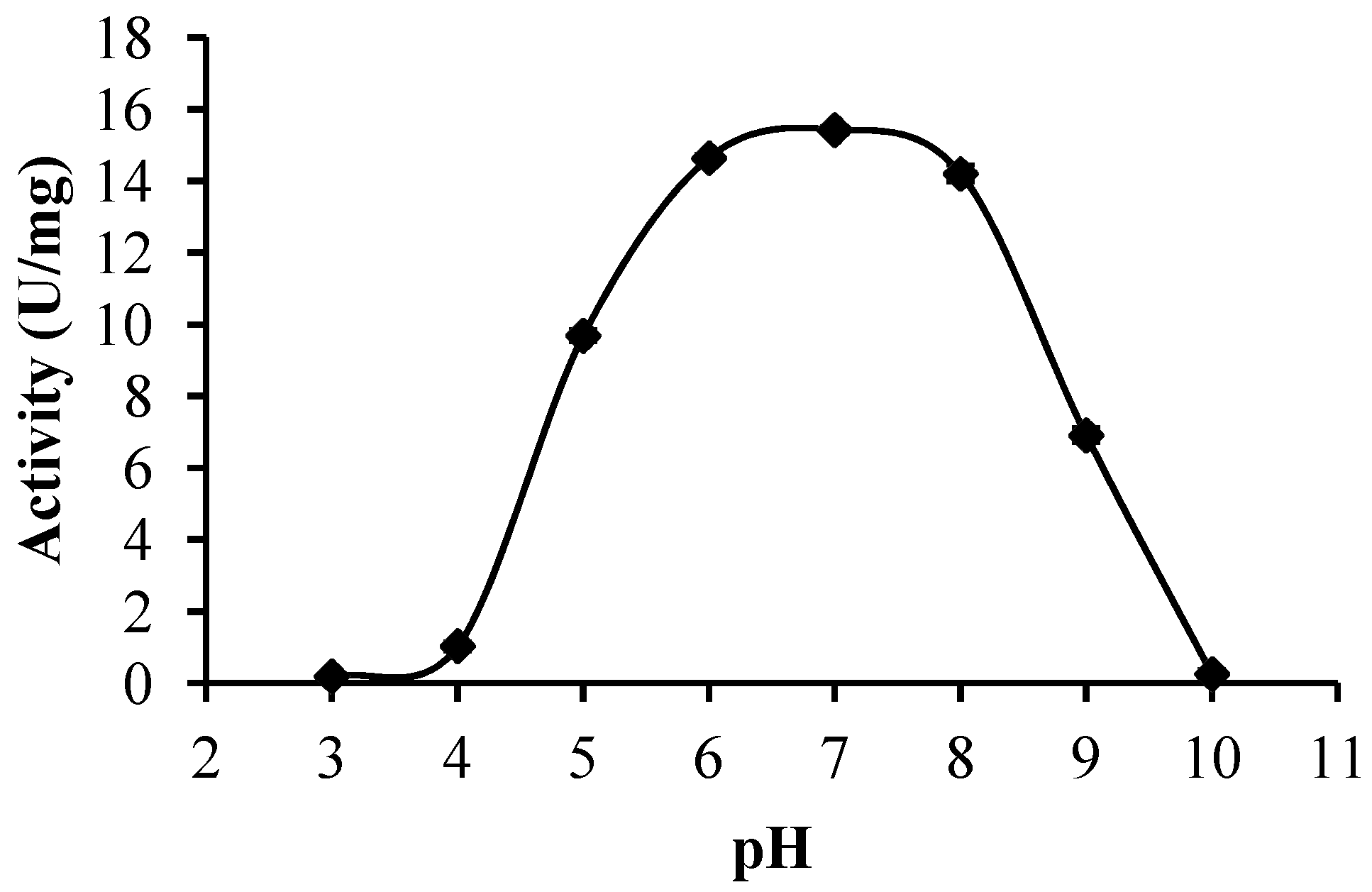
2.2.3. Optimum pH

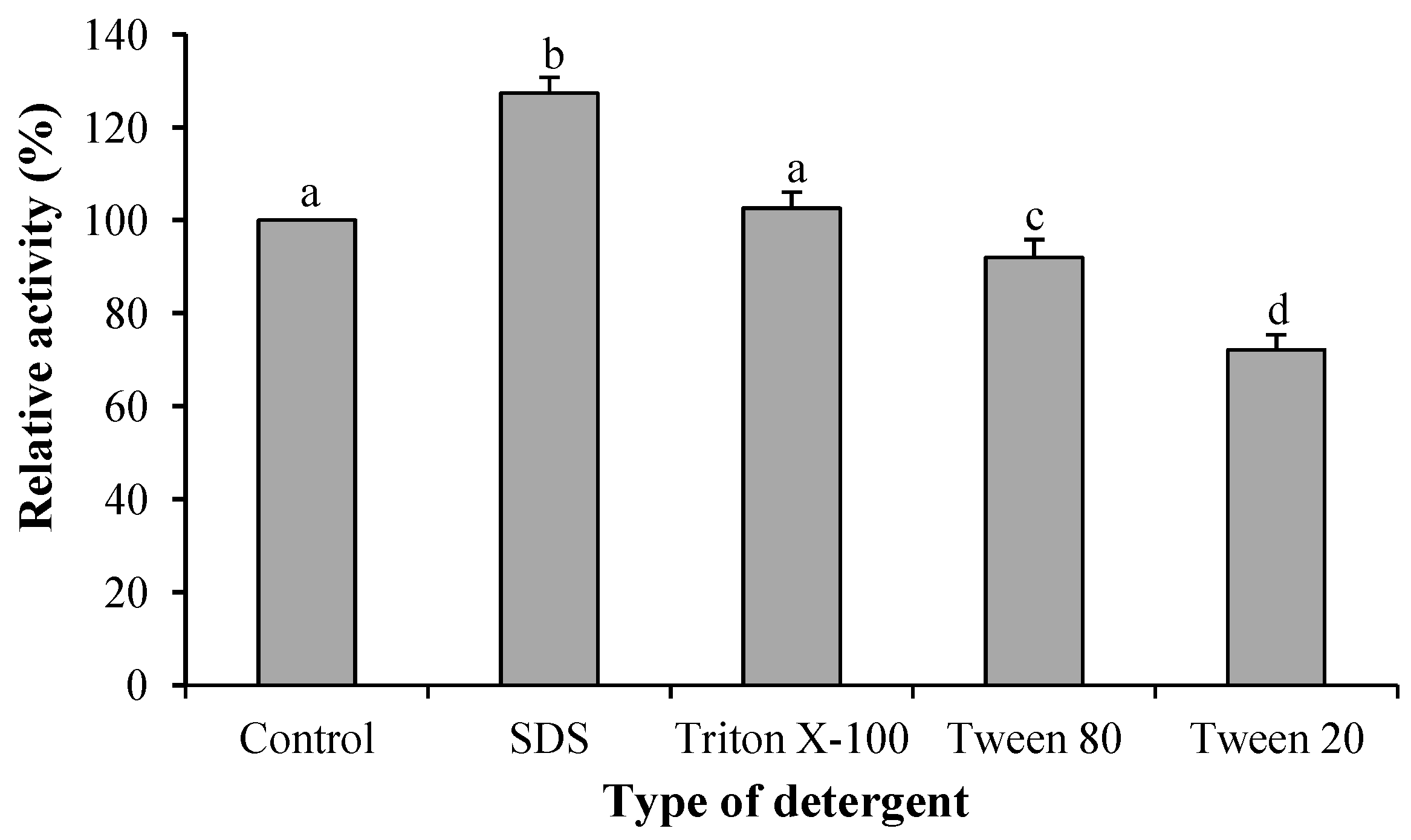
2.2.4. Effect of Cations and Detergents
| Cations | Relative Activity (%) |
|---|---|
| Control | 100.0 ± 0.0 a |
| K+ | 96.3 ± 3.4 a |
| Na+ | 100.7 ± 1.1 a |
| Li+ | 50.6 ± 3.2 b |
| Mg2+ | 89.0 ± 1.3 c |
| Ba2+ | 79.9 ± 2.2 d |
| Ca2+ | 76.4 ± 0.6 e |
| Co2+ | 59.5 ± 2.7 f |
| Zn2+ | 33.5 ± 2.2 g |
| Cu2+ | 4.7 ± 2.6 h |
| Hg2+ | 0.3 ± 0.1 i |
| Al3+ | 92.6 ± 3.4 c |


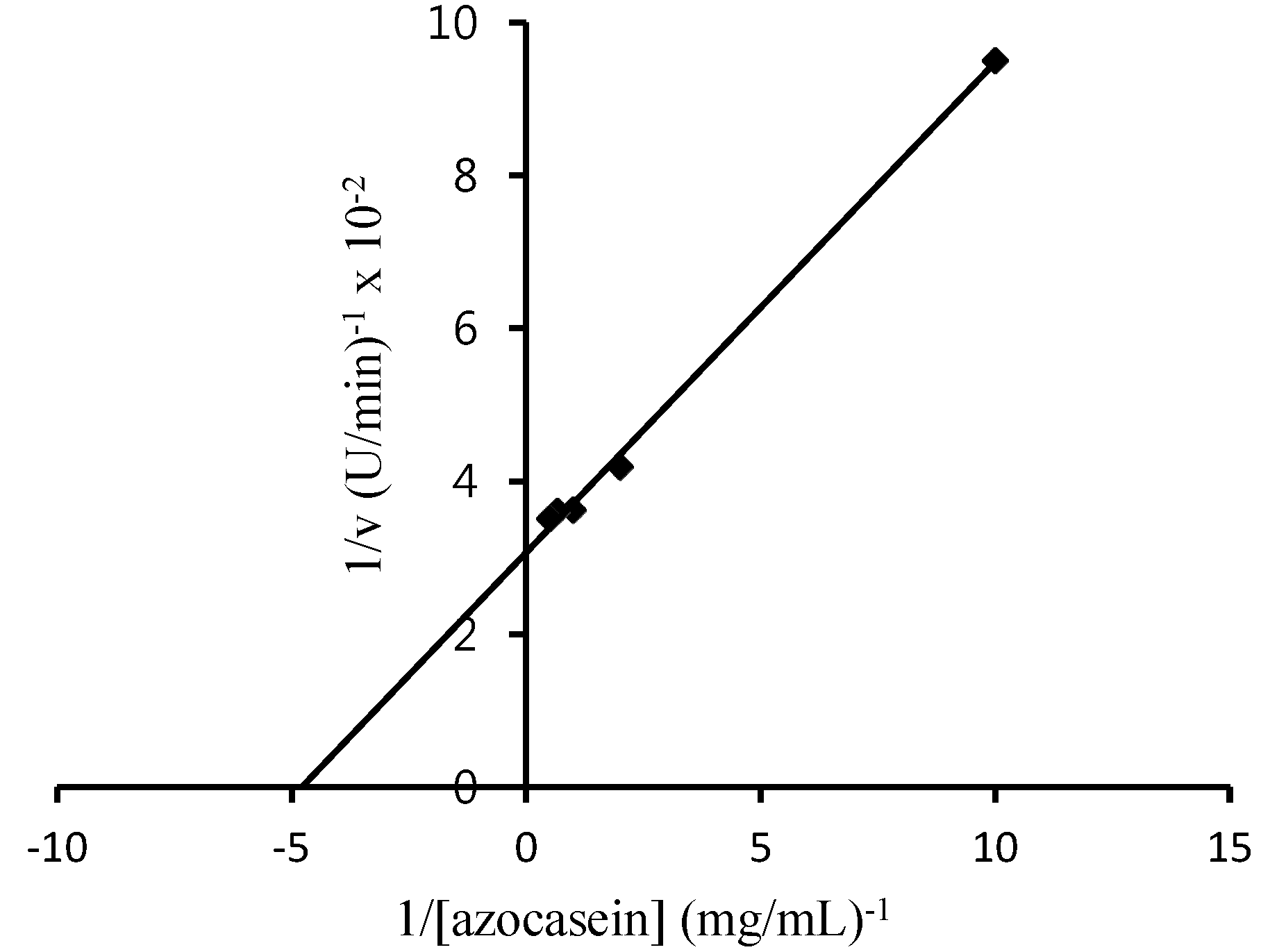
2.2.5. Km and Vmax Value of BGP
2.2.6. Storage Stability of BGP
| Time (day) | Relative Activity (%) at Different Storage Temperature | ||
|---|---|---|---|
| RT * | 4 ± 1 °C | −20 ± °C | |
| 0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 |
| 1 | 67.7 ± 1.1 | 87.1 ± 0.4 | 99.3 ± 0.1 |
| 2 | 47.1 ± 1.1 | 85.3 ± 0.2 | 98.5 ± 0.1 |
| 3 | 52.7 ± 0.7 | 83.2 ± 0.1 | 98.5 ± 0.1 |
| 4 | 37.0 ± 1.1 | 81.8 ± 0.7 | 98.1 ± 0.3 |
| 5 | 22.7 ± 2.3 | 78.8 ± 0.2 | 97.6 ± 0.3 |
| 10 | 16.3 ± 2.1 | 76.6 ± 0.3 | 96.9 ± 0.3 |
| 20 | 12.9 ± 2.3 | 74.0 ± 0.7 | 95.2 ± 0.1 |
| 30 | 9.9 ± 2.3 | 66.6 ± 0.2 | 94.4 ± 0.4 |
| 60 | 8.2 ± 3.3 | 62.1 ± 0.3 | 93.2 ± 0.4 |
| 90 | 3.4 ± 1.9 | 56.1 ± 0.8 | 93.6 ± 0.2 |
3. Experimental Section
3.1. Materials and Methods
3.2. Extraction of Bentong Ginger Protease (BGP)
3.3. Effect of pH on Extraction of BGP
3.4. Effect of Stabilizers on Extraction of Ginger Protease
3.5. Proteolytic Activity Assay
3.6. Protein Concentration Determination
3.7. Production of BGP Powder
3.8. Characterization of Bentong Ginger Protease
3.8.1. Determination of Protease Type
3.8.2. Optimum Temperature
3.8.3. Optimum pH
3.8.4. Effect of Cations and Detergents
3.8.5. Determination of Km and Vmax
3.8.6. Storage Stability
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ravindran, P.N.; Babu, N.B. Ginger: The Genus Zingiber; CRC Press: New York, NY, USA, 2005; pp. 1–15. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z.; Rahmat, A. Antioxidant activities, total phenolics and flavonoids content in two varieties of Malaysia young ginger (Zingiber. officinale Roscoe). Molecules 2010, 12, 4324–4333. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemical. Nat. Rev. Can. 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Jose, S. Industrial enzymes: A global strategic business report. Available online: http://www.prweb.com/releases/industrial_enzymes/proteases_carbohydrases/prweb8121185.htm (accessed on 2 July 2014).
- Rathina, R.K.; Mahendrakar, N.S. Effect of ensiling and organic solvents treatment on proteolytic enzymes of layer chicken intestine. J. Food Sci. Technol. 2010, 47, 320–324. [Google Scholar] [CrossRef]
- Su, H.P.; Huang, M.J.; Wang, H.T. Characterization of ginger proteases and their potential as a rennin replacement. J. Sci. Food Agric. 2009, 89, 1178–1185. [Google Scholar] [CrossRef]
- Ravikumar, G.; Gomathi, D.; Kalaiselvi, M.; Uma, C. A protease from the medicinal mushroom Pleurotus. sajor-caju; production, purification and partial characterization. Asian Pac. J. Trop. Biomed. 2012, 2, S411–S417. [Google Scholar] [CrossRef]
- Adulyatham, P.; Owusu-Apenten, R. Stabilization and partial purification of a protease from ginger rhizome (Zingiber. officinale Roscoe). J. Food Sci. 2005, 70, 231–234. [Google Scholar]
- Thompson, E.H.; Wolf, I.D.; Allen, C.E. Ginger rhizome: A new source of proteolytic enzyme. J. Food Sci. 1973, 38, 652–655. [Google Scholar] [CrossRef]
- Choi, K.H.; Laursen, R.A. Amino-acid sequence and glycan structures of cysteine protease with proline specifity from ginger rhizome (Zingiber. officinale). Eur. J. Biochem. 2000, 267, 1516–1526. [Google Scholar] [CrossRef]
- Bhaskar, N.; Sachindra, N.; Modi, V.; Sakhare, P.; Mahendrakar, N. Preparation of proteolytic activity rich ginger powder and evaluation of its tenderizing effect on spent-hen muscles. J. Musc. Food 2006, 17, 174–184. [Google Scholar] [CrossRef]
- Kim, M.; Hamilton, S.E.; Guddat, L.W.; Overall, C.M. Plant collagenase: Unique collagenolytic activity of cysteine proteases from ginger. Bba-Gen. Subjects 2007, 1770, 1627–1635. [Google Scholar] [CrossRef]
- Nafi, A.; Foo, H.L.; Jamilah, B.; Ghazali, H.M. Properties of proteolytic enzyme from ginger (Zingiber officinale Roscoe). Int. Food Res. J. 2013, 20, 2133–2138. [Google Scholar]
- Factfish. Malaysia: Ginger, production quantity (tons). Available online: http://www.factfish.com/statistic-country/malaysia/ginger,+production+quantity (accessed on 2 July 2014).
- Suhaimi, M.Y.; Mohamad, A.M.; Mahamud, S.; Khadzir, D. Effects of substrates on growth and yield of ginger cultivated using soilless culture. J. Trop. Agric. Fd. Sc 2012, 40, 159–168. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A. Elevated carbon dioxide increases contents of flavonoids and phenolic compounds, and antioxidant activities in Malaysian young ginger (Zingiber officinale Roscoe.) varieties. Molecules 2010, 15, 7907–7922. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E. Interactive effect of salicylic acid on some physiological features and antioxidant enzymes activity in ginger (Zingiber officinale Roscoe). Molecules 2013, 18, 5965–5979. [Google Scholar] [CrossRef]
- Aluko, R.E. The extraction and purification of proteins: An introduction. In Proteins in Food Processing, 1st ed.; Yada, R.Y., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 323–351. [Google Scholar]
- Qiao, Y.; Tong, J.; Wei, S.; Du, X.; Tang, X. Computer and computing technologies in agriculture II. In IFIP International Federation for Information Processing; Li, D., Chunjiang, Z., Eds.; Springer: Boston, MA, USA, 2009; pp. 1619–1628. [Google Scholar]
- Parisi, M.; Morena, S.; Fernandez, G. Characterization of a novel cysteine peptidase from tissue culture of garlic (Allium sativum L.). In Vitro Cell. Dev. Biol.-Plant 2002, 38, 608–612. [Google Scholar]
- Demir, Y.; Güngör, A.; Duran, E.D.; Demir, N. Cysteine protease (capparin) from capsules of caper (Capparisspinosa). Food Technol. Biotechnol. 2008, 46, 286–291. [Google Scholar]
- Frey, P.A.; Hegeman, A.D. Enzymatic Reaction Mechanisms; Oxford University Press, Inc.: New York, NY, USA, 2007; pp. 297–332. [Google Scholar]
- Huang, X.W.; Chen, L.J.; Luo, Y.B.; Guo, H.Y.; Ren, F.Z. Purification, characterization, and milk coagulating properties of ginger proteases. J. Dairy Sci. 2011, 94, 2259–2269. [Google Scholar] [CrossRef]
- Dubey, V.K.; Jagannadham, M.V. Procerain, a stable cysteine protease from the latex of Calotropis procera. Phytochemistry 2003, 62, 1057–1071. [Google Scholar] [CrossRef]
- Mehrnoush, A.; Mustafa, S.; Sarker, M.Z.I.; Yazid, A.M.M. Optimization of the conditions for extraction of serine protease from kesinai plant (Streblus. asper) leaves using response surface methodology. Molecules 2011, 16, 9245–9260. [Google Scholar]
- Voet, D.; Voet, J.G. Biochemistry, 4th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2011; pp. 129–162. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurenment with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar]
- Sample Availability: Samples of the compounds Bentong ginger are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nafi', A.; Ling, F.H.; Bakar, J.; Ghazali, H.M. Partial Characterization of an Enzymatic Extract from Bentong Ginger (Zingiber officinale var. Bentong). Molecules 2014, 19, 12336-12348. https://doi.org/10.3390/molecules190812336
Nafi' A, Ling FH, Bakar J, Ghazali HM. Partial Characterization of an Enzymatic Extract from Bentong Ginger (Zingiber officinale var. Bentong). Molecules. 2014; 19(8):12336-12348. https://doi.org/10.3390/molecules190812336
Chicago/Turabian StyleNafi', Ahmad, Foo Hooi Ling, Jamilah Bakar, and Hasanah M. Ghazali. 2014. "Partial Characterization of an Enzymatic Extract from Bentong Ginger (Zingiber officinale var. Bentong)" Molecules 19, no. 8: 12336-12348. https://doi.org/10.3390/molecules190812336
APA StyleNafi', A., Ling, F. H., Bakar, J., & Ghazali, H. M. (2014). Partial Characterization of an Enzymatic Extract from Bentong Ginger (Zingiber officinale var. Bentong). Molecules, 19(8), 12336-12348. https://doi.org/10.3390/molecules190812336