
Lectin Engineering, a Molecular Evolutionary Approach to Expanding the Lectin Utilities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
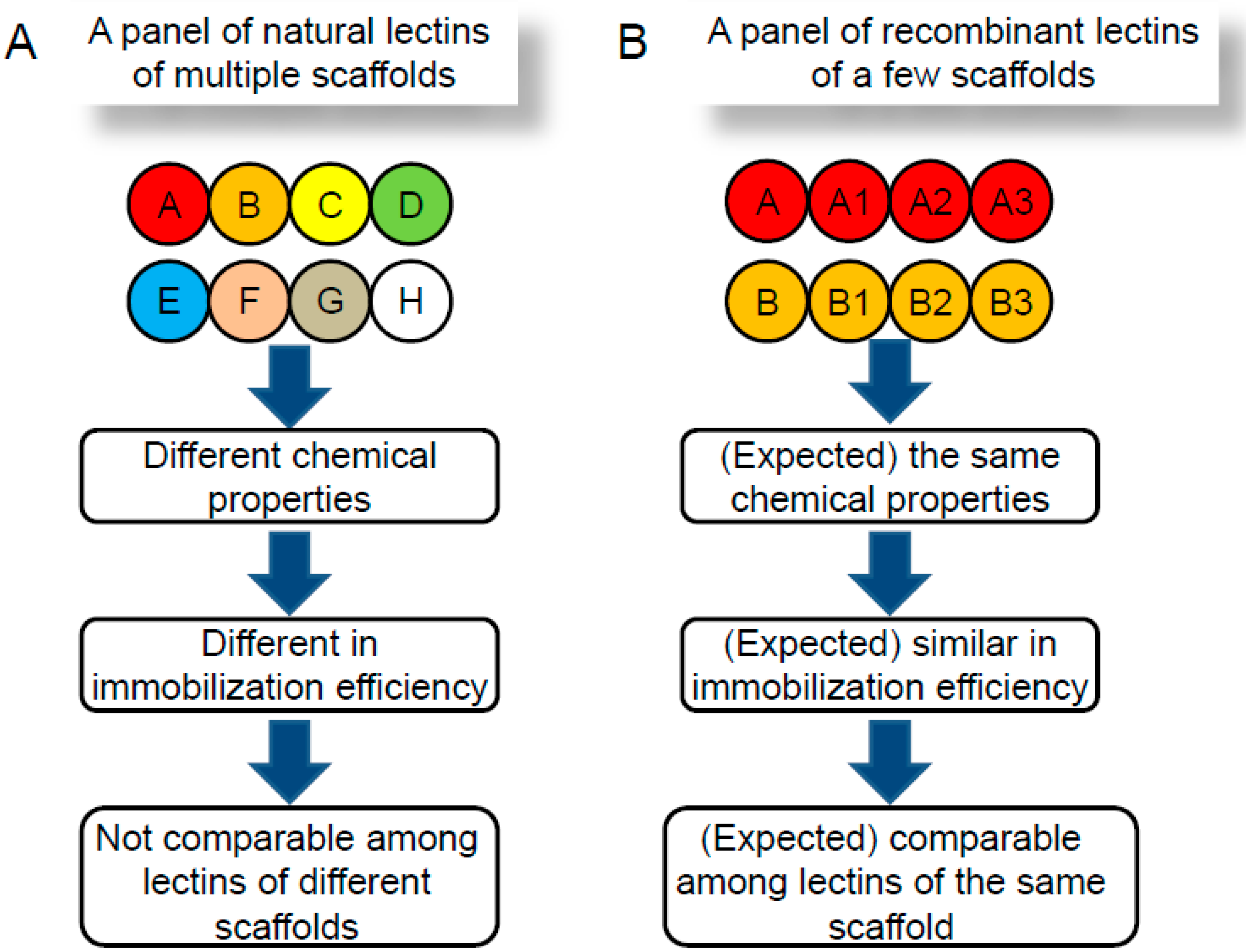
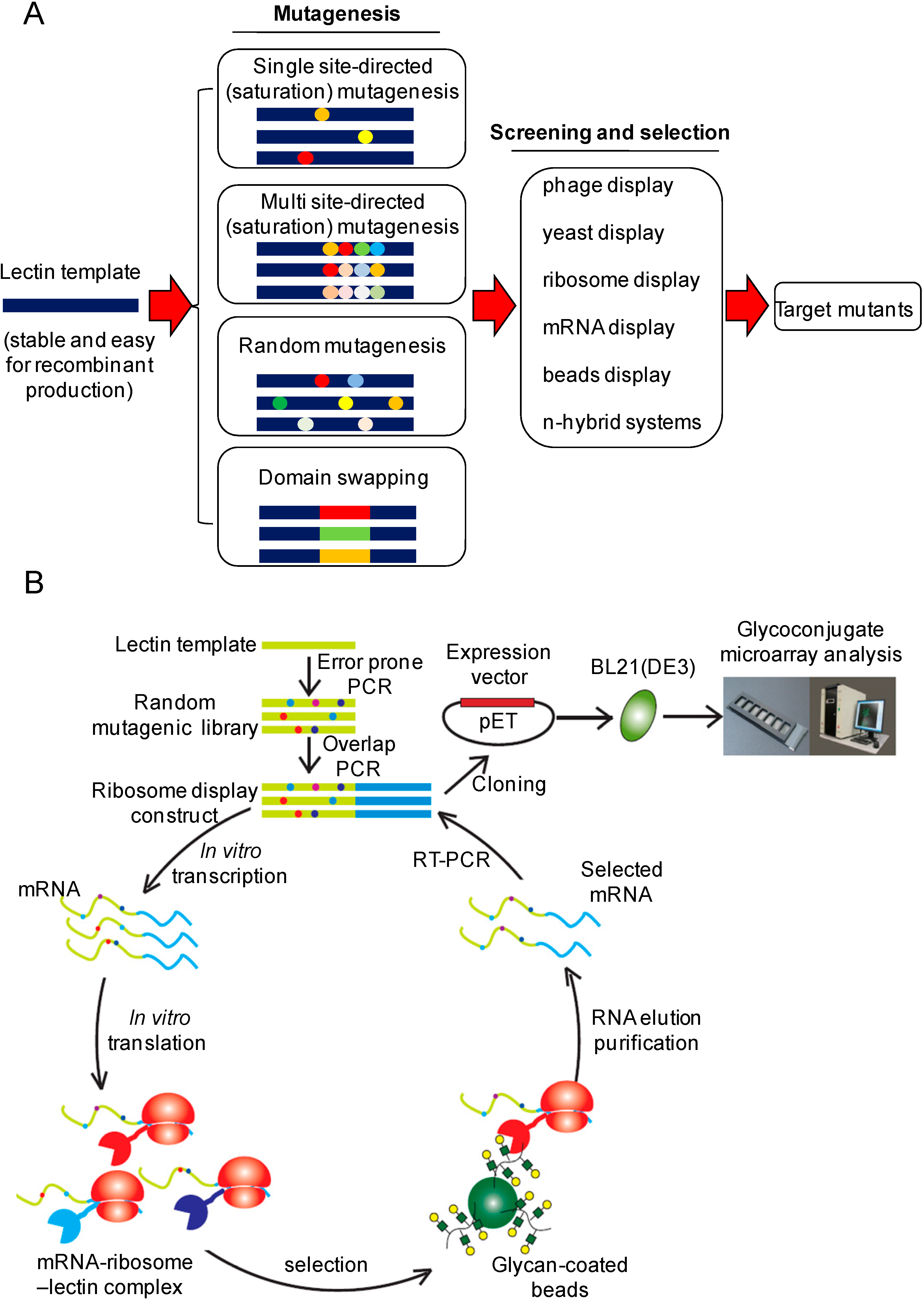
:1. Introduction
2. Current State of the Art of Lectin Engineering
2.1. Engineering of L-Type Lectins

2.2. Engineering of Galectins

2.3. Engineering of R-Type Lectins

2.4. Engineering of C-Type Lectins
2.5. Engineering of β-Propeller Lectins
3. Key Points Associated with the Strategy of Lectin Engineering
3.1. Scaffold (Template)
3.2. Methods for Mutagenesis
3.3. Methods for Screening

4. Future Perspectives
Acknowledgments
Author Contributions
Abbreviations
| Gal | galactose |
| GalNAc | N-acetylgalactosamine |
| Glc | glucose |
| GlcNAc | N-acetylglucosamine |
| Man | mannose |
| Fuc | fucose |
| Neu5Ac | N-Acetyl neuraminic acid |
Conflicts of Interest
References
- Hedlund, M.; Ng, E.; Varki, A.; Varki, N.M. alpha 2–6-Linked sialic acids on N-glycans modulate carcinoma differentiation in vivo. Cancer Res. 2008, 68, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Toyota, M.; Saito, S.; Onuma, Y.; Ito, Y.; Hiemori, K.; Fukumura, M.; Matsushima, A.; Nakanishi, M.; Ohnuma, K.; et al. Glycome diagnosis of human induced pluripotent stem cells using lectin microarray. J. Biol. Chem. 2011, 286, 20345–20353. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J. Lectin-based structural glycomics: Glycoproteomics and glycan profiling. Glycoconj. J. 2004, 21, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J.; Kuno, A.; Tateno, H. Lectin-based structural glycomics: A practical approach to complex glycans. Electrophoresis 2011, 32, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, L.; Mahal, L.K. Glycomic analysis: An array of technologies. ACS Chem. Biol. 2009, 4, 715–732. [Google Scholar] [CrossRef] [PubMed]
- Zaia, J. Mass spectrometry and the emerging field of glycomics. Chem. Biol. 2008, 15, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Hasehira, K.; Tateno, H.; Onuma, Y.; Ito, Y.; Asashima, M.; Hirabayashi, J. Structural and quantitative evidence for dynamic glycome shift on production of induced pluripotent stem cells. Mol. Cell. Proteomics 2012, 11, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.P.; Royle, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M. GlycoBase and autoGU: Tools for HPLC-based glycan analysis. Bioinformatics 2008, 24, 1214–1216. [Google Scholar] [CrossRef] [PubMed]
- Kuno, A.; Uchiyama, N.; Koseki-Kuno, S.; Ebe, Y.; Takashima, S.; Yamada, M.; Hirabayashi, J. Evanescent-field fluorescence-assisted lectin microarray: A new strategy for glycan profiling. Nat. Methods 2005, 2, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Pilobello, K.T.; Slawek, D.E.; Mahal, L.K. A ratiometric lectin microarray approach to analysis of the dynamic mammalian glycome. Proc. Natl. Acad. Sci. USA 2007, 104, 11534–11539. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Nakamura-Tsuruta, S.; Hirabayashi, J. Frontal affinity chromatography: Sugar-protein interactions. Nat. Protoc. 2007, 2, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.; Blixt, O.; Glaser, L.; Taubenberger, J.K.; Palese, P.; Paulson, J.C.; Wilson, I.A. Glycan microarray analysis of the hemagglutinins from modern and pandemic influenza viruses reveals different receptor specificities. J. Mol. Biol. 2006, 355, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Kamiya, Y.; Totani, K.; Kamiya, D.; Kawasaki, N.; Yamaguchi, D.; Matsuo, I.; Matsumoto, N.; Ito, Y.; Kato, K.; et al. Sugar-binding activity of the MRH domain in the ER alpha-glucosidase II beta subunit is important for efficient glucose trimming. Glycobiology 2009, 19, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, N.; Ichikawa, Y.; Matsuo, I.; Totani, K.; Matsumoto, N.; Ito, Y.; Yamamoto, K. The sugar-binding ability of ERGIC-53 is enhanced by its interaction with MCFD2. Blood 2008, 111, 1972–1979. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, N.; Matsuo, I.; Totani, K.; Nawa, D.; Suzuki, N.; Yamaguchi, D.; Matsumoto, N.; Ito, Y.; Yamamoto, K. Detection of weak sugar binding activity of VIP36 using VIP36-streptavidin complex and membrane-based sugar chains. J. Biochem. 2007, 141, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K.; Yamaguchi, D.; Tateno, H.; Hu, D.; Qin, S.Y.; Kawasaki, N.; Yamada, M.; Matsumoto, N.; Hirabayashi, J.; Ito, Y.; et al. The sugar-binding ability of human OS-9 and its involvement in ER-associated degradation. Glycobiology 2010, 20, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, D.; Kawasaki, N.; Matsuo, I.; Totani, K.; Tozawa, H.; Matsumoto, N.; Ito, Y.; Yamamoto, K. VIPL has sugar-binding activity specific for high-mannose-type N-glycans, and glucosylation of the alpha1,2 mannotriosyl branch blocks its binding. Glycobiology 2007, 17, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Onuma, Y.; Tateno, H.; Hirabayashi, J.; Ito, Y.; Asashima, M. rBC2LCN, a new probe for live cell imaging of human pluripotent stem cells. Biochem. Biophys. Res. Commun. 2013, 431, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Onuma, Y.; Ito, Y. Live-cell imaging of human pluripotent stem cells by a novel lectin probe rBC2LCN. Methods Mol. Biol. 2014, 1200, 313–318. [Google Scholar] [PubMed]
- Kuno, A.; Ikehara, Y.; Tanaka, Y.; Angata, T.; Unno, S.; Sogabe, M.; Ozaki, H.; Ito, K.; Hirabayashi, J.; Mizokami, M.; et al. Multilectin assay for detecting fibrosis-specific glyco-alteration by means of lectin microarray. Clin. Chem. 2011, 57, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.L.; Gildersleeve, J.C.; Mahal, L.K. A simple strategy for the creation of a recombinant lectin microarray. Mol. Biosyst. 2008, 4, 654–662. [Google Scholar] [CrossRef] [PubMed]
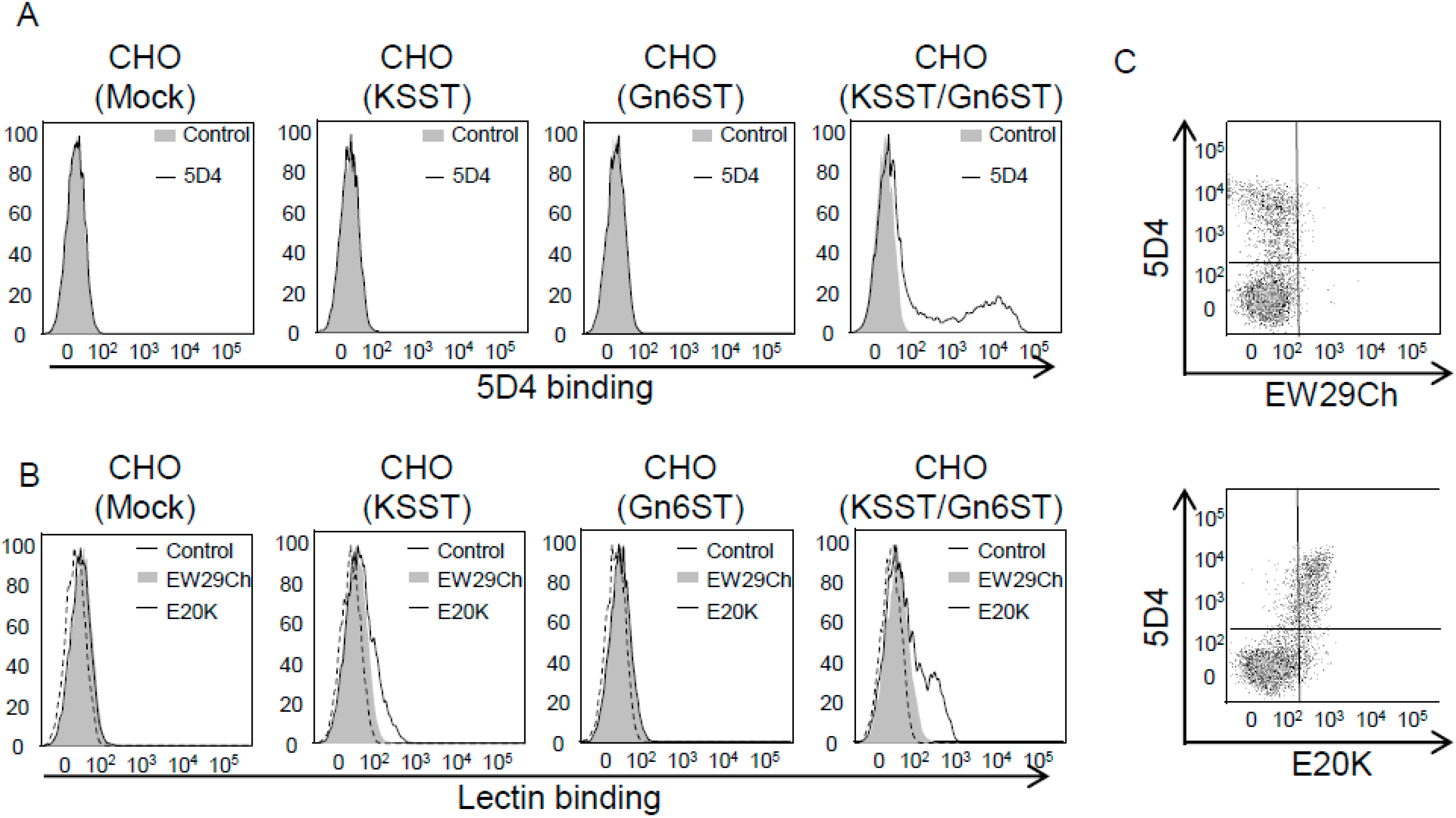
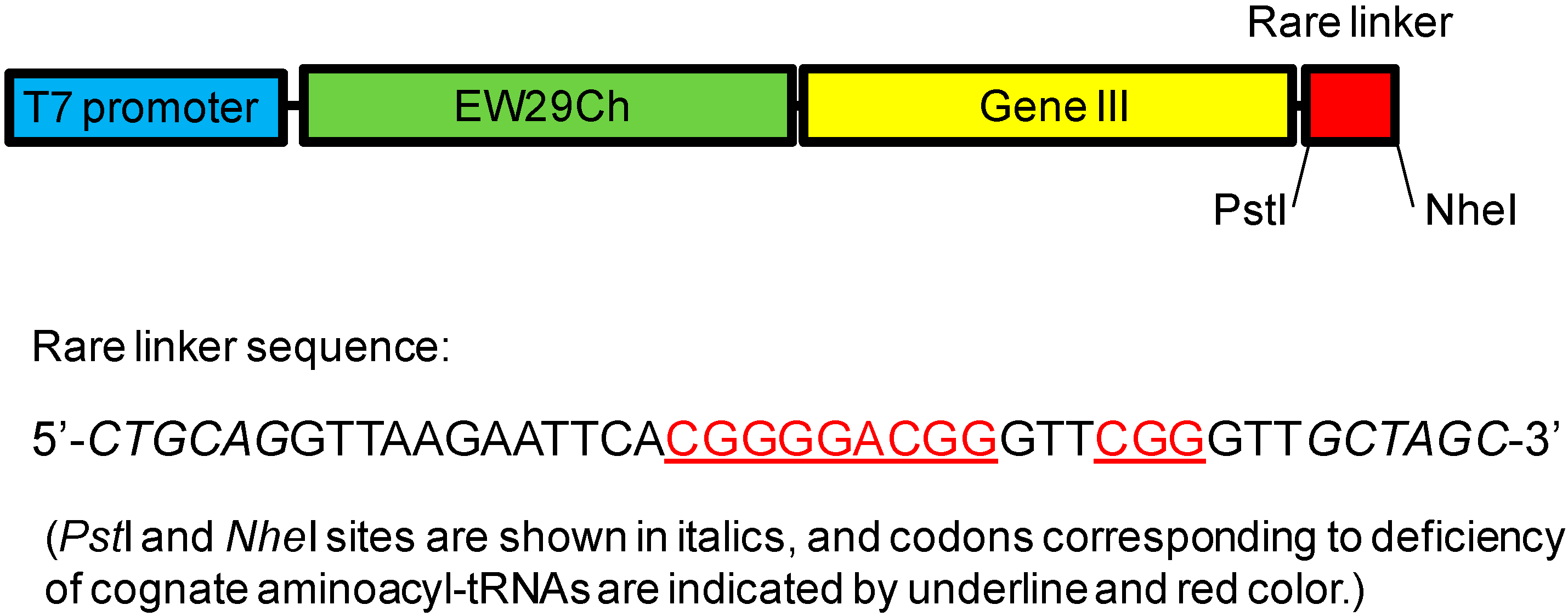
- Hu, D.; Tateno, H.; Kuno, A.; Yabe, R.; Hirabayashi, J. Directed evolution of lectins with sugar-binding specificity for 6-sulfo-galactose. J. Biol. Chem. 2012, 287, 20313–20320. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Lis, H. Legume lectins—A large family of homologous proteins. FASEB J. 1990, 4, 3198–3208. [Google Scholar] [PubMed]
- Arango, R.; Rodriguez-Arango, E.; Adar, R.; Belenky, D.; Loontiens, F.G.; Rozenblatt, S.; Sharon, N. Modification by site-directed mutagenesis of the specificity of Erythrina corallodendron lectin for galactose derivatives with bulky substituents at C-2. FEBS Lett. 1993, 330, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Jordan, E.T.; Goldstein, I.J. Site-directed mutagenesis studies on the lima bean lectin. Altered carbohydrate-binding specificities result from single amino acid substitutions. Eur. J. Biochem. 1995, 230, 958–964. [Google Scholar] [CrossRef] [PubMed]
- Thamotharan, S.; Karthikeyan, T.; Kulkarni, K.A.; Shetty, K.N.; Surolia, A.; Vijayan, M.; Suguna, K. Modification of the sugar specificity of a plant lectin: Structural studies on a point mutant of Erythrina corallodendron lectin. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 218–227. [Google Scholar] [CrossRef]
- Adhikari, P.; Bachhawat-Sikder, K.; Thomas, C.J.; Ravishankar, R.; Jeyaprakash, A.A.; Sharma, V.; Vijayan, M.; Surolia, A. Mutational analysis at Asn-41 in peanut agglutinin—A residue critical for the binding of the tumor-associated Thomsen-Friedenreich antigen. J. Biol. Chem. 2001, 276, 40734–40739. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Konami, Y.; Osawa, T.; Irimura, T. Alteration of the carbohydrate-binding specificity of the Bauhinia purpurea lectin through the preparation of a chimeric lectin. J. Biochem. 1992, 111, 87–90. [Google Scholar] [PubMed]
- Yamamoto, K.; Konami, Y.; Osawa, T. A chimeric lectin formed from Bauhinia purpurea lectin and Lens culinaris lectin recognizes a unique carbohydrate structure. J. Biochem. 2000, 127, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Nishiguchi, M.; Yoshida, K.; Sumizono, T.; Tazaki, K. Studies by site-directed mutagenesis of the carbohydrate-binding properties of a bark lectin from Robinia pseudoacacia. FEBS Lett. 1997, 403, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Maruyama, I.N.; Osawa, T. Cyborg lectins: Novel leguminous lectins with unique specificities. J. Biochem. 2000, 127, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Maenuma, K.; Yim, M.; Komatsu, K.; Hoshino, M.; Tachiki-Fujioka, A.; Takahashi, K.; Hiki, Y.; Bovin, N.; Irimura, T. A library of mutated Maackia amurensis hemagglutinin distinguishes putative glycoforms of immunoglobulin A1 from IgA nephropathy patients. J. Proteome Res. 2009, 8, 3617–3624. [Google Scholar] [CrossRef] [PubMed]
- Maenuma, K.; Yim, M.; Komatsu, K.; Hoshino, M.; Takahashi, Y.; Bovin, N.; Irimura, T. Use of a library of mutated Maackia amurensis hemagglutinin for profiling the cell lineage and differentiation. Proteomics 2008, 8, 3274–3283. [Google Scholar] [CrossRef] [PubMed]
- Streicher, H.; Sharon, N. Recombinant plant lectins and their mutants. Methods Enzymol. 2003, 363, 47–77. [Google Scholar] [PubMed]
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [PubMed]
- Leffler, H.; Carlsson, S.; Hedlund, M.; Qian, Y.; Poirier, F. Introduction to galectins. Glycoconj. J. 2004, 19, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Klyosov, A.A.; Traber, P.G. Galectins in Disease and Potential Therapeutic Approaches. ACS Symp. Ser. 2012, 1115, 3–43. [Google Scholar]
- Nishi, N.; Itoh, A.; Fujiyama, A.; Yoshida, N.; Araya, S.; Hirashima, M.; Shoji, H.; Nakamura, T. Development of highly stable galectins: Truncation of the linker peptide confers protease-resistance on tandem-repeat type galectins. FEBS Lett. 2005, 579, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Itoh, A.; Fukata, Y.; Miyanaka, H.; Nonaka, Y.; Ogawa, T.; Nakamura, T.; Nishi, N. Optimization of the inter-domain structure of galectin-9 for recombinant production. Glycobiology 2013, 23, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, L.; Lensch, M.; Gabius, H.J.; Fee, C.J.; Middelberg, A.P. Single-site Cys-substituting mutation of human lectin galectin-2: Modulating solubility in recombinant production, reducing long-term aggregation, and enabling site-specific monoPEGylation. Biomacromolecules 2008, 9, 3223–3230. [Google Scholar] [CrossRef] [PubMed]
- Kopitz, J.; Fik, Z.; Andre, S.; Smetana, K.; Gabius, H.J. Single-site mutational engineering and following monoPEGylation of the human lectin galectin-2: Effects on ligand binding, functional aspects, and clearance from serum. Mol. Pharm. 2013, 10, 2054–2061. [Google Scholar] [CrossRef] [PubMed]
- Ban, M.; Yoon, H.J.; Demirkan, E.; Utsumi, S.; Mikami, B.; Yagi, F. Structural basis of a fungal galectin from Agrocybe cylindracea for recognizing sialoconjugate. J. Mol. Biol. 2005, 351, 695–706. [Google Scholar] [CrossRef] [PubMed]
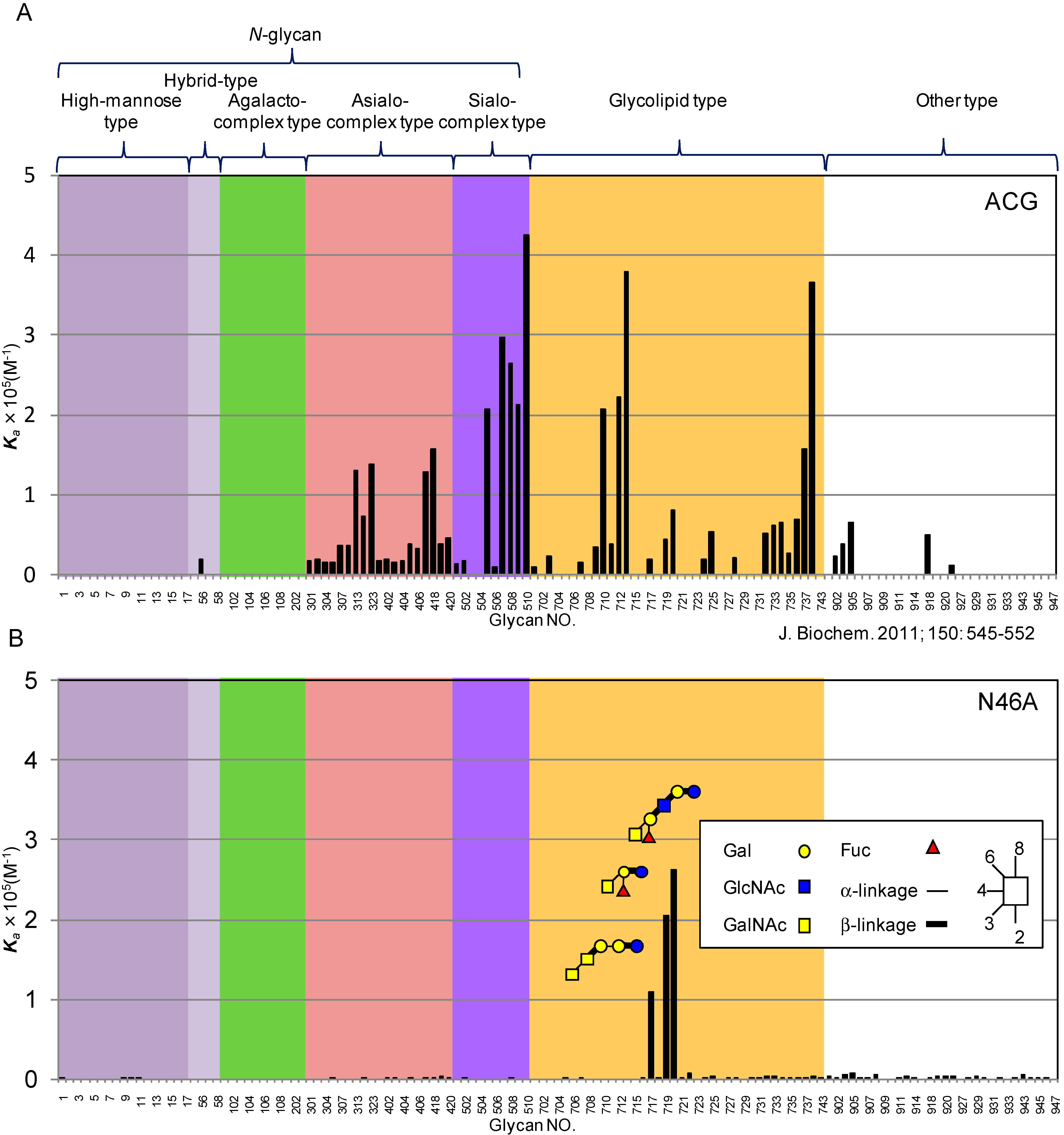
- Imamura, K.; Takeuchi, H.; Yabe, R.; Tateno, H.; Hirabayashi, J. Engineering of the glycan-binding specificity of Agrocybe cylindracea galectin towards alpha(2,3)-linked sialic acid by saturation mutagenesis. J. Biochem. 2011, 150, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Yagi, F.; Hiroyama, H.; Kodama, S. Agrocybe cylindracea lectin is a member of the galectin family. Glycoconj. J. 2001, 18, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Tateno, H.; Sato, T.; Narimatsu, H.; Hirabayashi, J. Tailoring GalNAc alpha 1–3Gal beta-specific lectins from a multi-specific fungal galectin: Dramatic change of carbohydrate specificity by a single amino-acid substitution. Biochem. J. 2013, 453, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, N.; Hu, D.; Tateno, H.; Makyio, H.; Hirabayashi, J.; Kato, R. Conformational change of a unique sequence in a fungal galectin from Agrocybe cylindracea controls glycan ligand-binding specificity. FEBS Lett. 2013, 587, 3620–3625. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Huang, H.; Tateno, H.; Nakakita, S.; Sato, T.; Narimatsu, H.; Yao, X.; Hirabayashi, J. Engineering of a 3'-sulpho-Galbeta1–4GlcNAc-specific probe by a single amino acid substitution of a fungal galectin. J. Biochem. 2015, 157, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Ryckaert, S.; Callewaert, N.; Jacobs, P.P.; Dewaele, S.; Dewerte, I.; Contreras, R. Fishing for lectins from diverse sequence libraries by yeast surface display—An exploratory study. Glycobiology 2008, 18, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Kuno, A.; Ito, S.; Suzuki, R.; Hasegawa, T.; Hirabayashi, J. NMR studies on the interaction of sugars with the C-terminal domain of an R-type lectin from the earthworm Lumbricus terrestris. FEBS J. 2009, 276, 2095–2105. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, N.; Goldstein, I.J.; Broekaert, W.F.; Nsimba-Lubaki, M.; Peeters, B.; Peumans, W.J. The elderberry (Sambucus nigra L.) bark lectin recognizes the Neu5Ac(alpha 2–6)Gal/GalNAc sequence. J. Biol. Chem. 1987, 262, 1596–1601. [Google Scholar] [PubMed]
- Kaku, H.; Tanaka, Y.; Tazaki, K.; Minami, E.; Mizuno, H.; Shibuya, N. Sialylated oligosaccharide-specific plant lectin from Japanese elderberry (Sambucus sieboldiana) bark tissue has a homologous structure to type II ribosome-inactivating proteins, ricin and abrin. cDNA cloning and molecular modeling study. J. Biol. Chem. 1996, 271, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Mori, A.; Uchiyama, N.; Yabe, R.; Iwaki, J.; Shikanai, T.; Angata, T.; Narimatsu, H.; Hirabayashi, J. Glycoconjugate microarray based on an evanescent-field fluorescence-assisted detection principle for investigation of glycan-binding proteins. Glycobiology 2008, 18, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Fiete, D.J.; Beranek, M.C.; Baenziger, J.U. A cysteine-rich domain of the “mannose” receptor mediates GalNAc-4-SO4 binding. Proc. Natl. Acad. Sci. USA 1998, 95, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J.; Dutta, S.K.; Kasai, K. Novel galactose-binding proteins in Annelida. Characterization of 29-kDa tandem repeat-type lectins from the earthworm Lumbricus terrestris. J. Biol. Chem. 1998, 273, 14450–14460. [Google Scholar] [CrossRef] [PubMed]
- Yabe, R.; Suzuki, R.; Kuno, A.; Fujimoto, Z.; Jigami, Y.; Hirabayashi, J. Tailoring a novel sialic acid-binding lectin from a ricin-B chain-like galactose-binding protein by natural evolution-mimicry. J. Biochem. 2007, 141, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Yabe, R.; Itakura, Y.; Nakamura-Tsuruta, S.; Iwaki, J.; Kuno, A.; Hirabayashi, J. Engineering a versatile tandem repeat-type alpha 2-6sialic acid-binding lectin. Biochem. Biophys. Res. Commun. 2009, 384, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Michelow, I.C.; Dong, M.; Mungall, B.A.; Yantosca, L.M.; Lear, C.; Ji, X.; Karpel, M.; Rootes, C.L.; Brudner, M.; Houen, G.; et al. A novel L-ficolin/mannose-binding lectin chimeric molecule with enhanced activity against Ebola virus. J. Biol. Chem. 2010, 285, 24729–24739. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Hartshorn, K.L.; White, M.R.; Moyo, P.; Michelow, I.C.; Koziel, H.; Kinane, B.T.; Schmidt, E.V.; Fujita, T.; Takahashi, K. Recombinant chimeric lectins consisting of mannose-binding lectin and L-ficolin are potent inhibitors of influenza A virus compared with mannose-binding lectin. Biochem. Pharmacol. 2011, 81, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, I.; Soding, J.; Lupas, A.N. Evolution of the beta-propeller fold. Proteins 2008, 71, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Olausson, J.; Astrom, E.; Jonsson, B.H.; Tibell, L.A.E.; Pahlsson, P. Production and characterization of a monomeric form and a single-site form of Aleuria aurantia lectin. Glycobiology 2011, 21, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Yadid, I.; Tawfik, D.S. Functional beta-propeller lectins by tandem duplications of repetitive units. Protein Eng. Des. Sel. 2011, 24, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Raemaekers, R.J.; de Muro, L.; Gatehouse, J.A.; Fordham-Skelton, A.P. Functional phytohemagglutinin (PHA) and Galanthus nivalis agglutinin (GNA) expressed in Pichia pastoris correct N-terminal processing and secretion of heterologous proteins expressed using the PHA-E signal peptide. Eur. J. Biochem. 1999, 265, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Propheter, D.C.; Hsu, K.L.; Mahal, L.K. Recombinant lectin microarrays for glycomic analysis. Methods Mol. Biol. 2011, 723, 67–77. [Google Scholar] [PubMed]
- Topin, J.; Arnaud, J.; Sarkar, A.; Audfray, A.; Gillon, E.; Perez, S.; Jamet, H.; Varrot, A.; Imberty, A.; Thomas, A. Deciphering the glycan preference of bacterial lectins by glycan array and molecular docking with validation by microcalorimetry and crystallography. PLoS ONE 2013, 8, e71149. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Percival Zhang, Y.H. Easy preparation of a large-size random gene mutagenesis library in Escherichia coli. Anal. Biochem. 2012, 428, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Krauss, U.; Jaeger, K.E.; Eggert, T. Rapid sequence scanning mutagenesis using in silico oligo design and the Megaprimer PCR of whole plasmid method (MegaWHOP). Methods Mol. Biol. 2010, 634, 127–135. [Google Scholar] [PubMed]
- Nannemann, D.P.; Birmingham, W.R.; Scism, R.A.; Bachmann, B.O. Assessing directed evolution methods for the generation of biosynthetic enzymes with potential in drug biosynthesis. Future Med. Chem. 2011, 3, 809–819. [Google Scholar] [CrossRef] [PubMed]
- McCullum, E.O.; Williams, B.A.; Zhang, J.; Chaput, J.C. Random mutagenesis by error-prone PCR. Methods Mol. Biol. 2010, 634, 103–109. [Google Scholar] [PubMed]
- Baldwin, A.J.; Busse, K.; Simm, A.M.; Jones, D.D. Expanded molecular diversity generation during directed evolution by trinucleotide exchange (TriNEx). Nucleic Acids Res. 2008, 36, e77. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Cornish, V.W. Screening and selection methods for large-scale analysis of protein function. Angew. Chem. Int. Ed. 2002, 41, 4402–4425. [Google Scholar] [CrossRef]
- He, M.; Khan, F. Ribosome display: Next-generation display technologies for production of antibodies in vitro. Expert Rev. Proteomics 2005, 2, 421–430. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Tateno, H.; Hirabayashi, J. Lectin Engineering, a Molecular Evolutionary Approach to Expanding the Lectin Utilities. Molecules 2015, 20, 7637-7656. https://doi.org/10.3390/molecules20057637
Hu D, Tateno H, Hirabayashi J. Lectin Engineering, a Molecular Evolutionary Approach to Expanding the Lectin Utilities. Molecules. 2015; 20(5):7637-7656. https://doi.org/10.3390/molecules20057637
Chicago/Turabian StyleHu, Dan, Hiroaki Tateno, and Jun Hirabayashi. 2015. "Lectin Engineering, a Molecular Evolutionary Approach to Expanding the Lectin Utilities" Molecules 20, no. 5: 7637-7656. https://doi.org/10.3390/molecules20057637