
A Herbal Formula HT048, Citrus unshiu and Crataegus pinnatifida, Prevents Obesity by Inhibiting Adipogenesis and Lipogenesis in 3T3-L1 Preadipocytes and HFD-Induced Obese Rats

Abstract
:1. Introduction
2. Results and Discussion
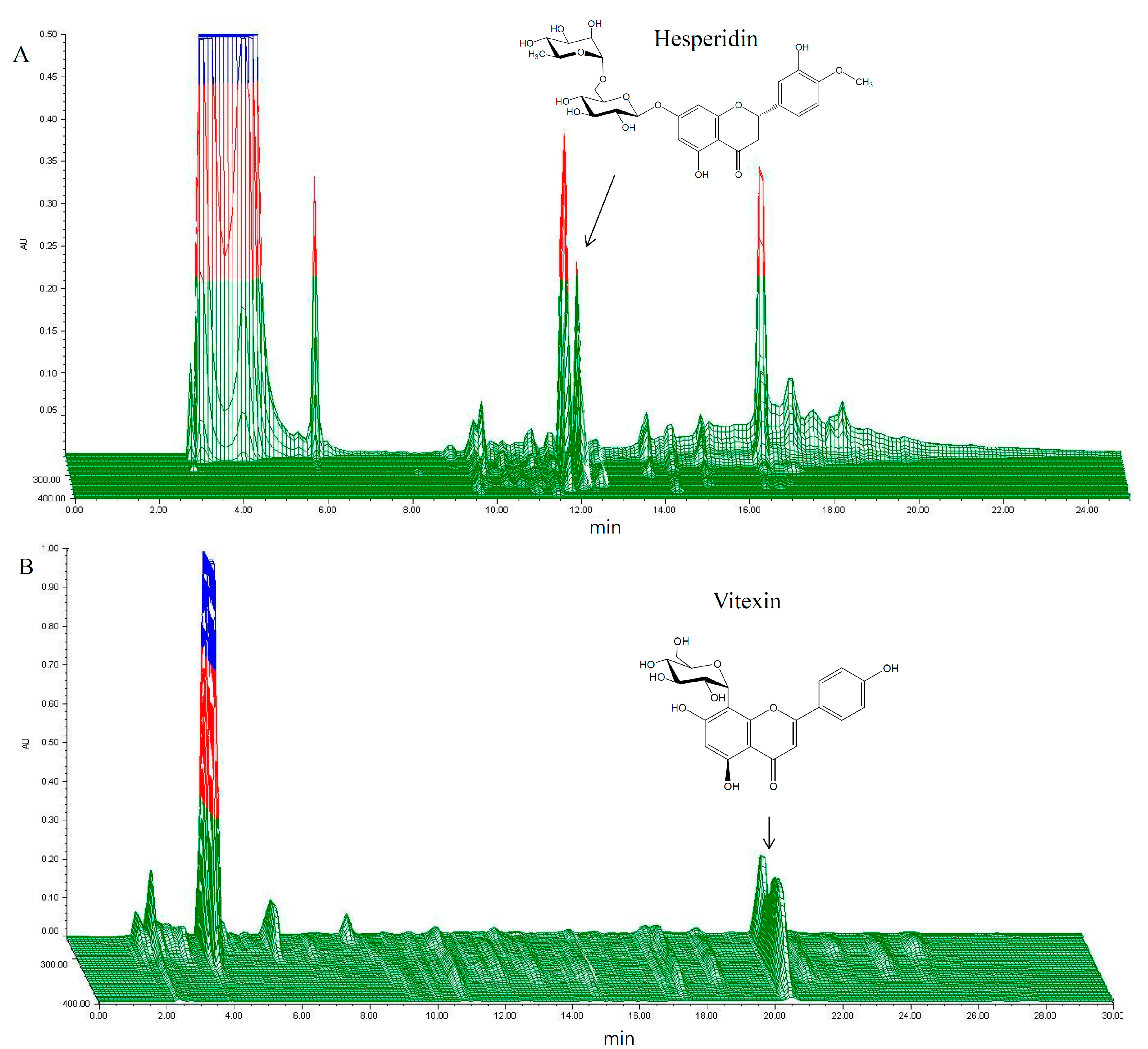
2.1. HPLC Analysis of HT048

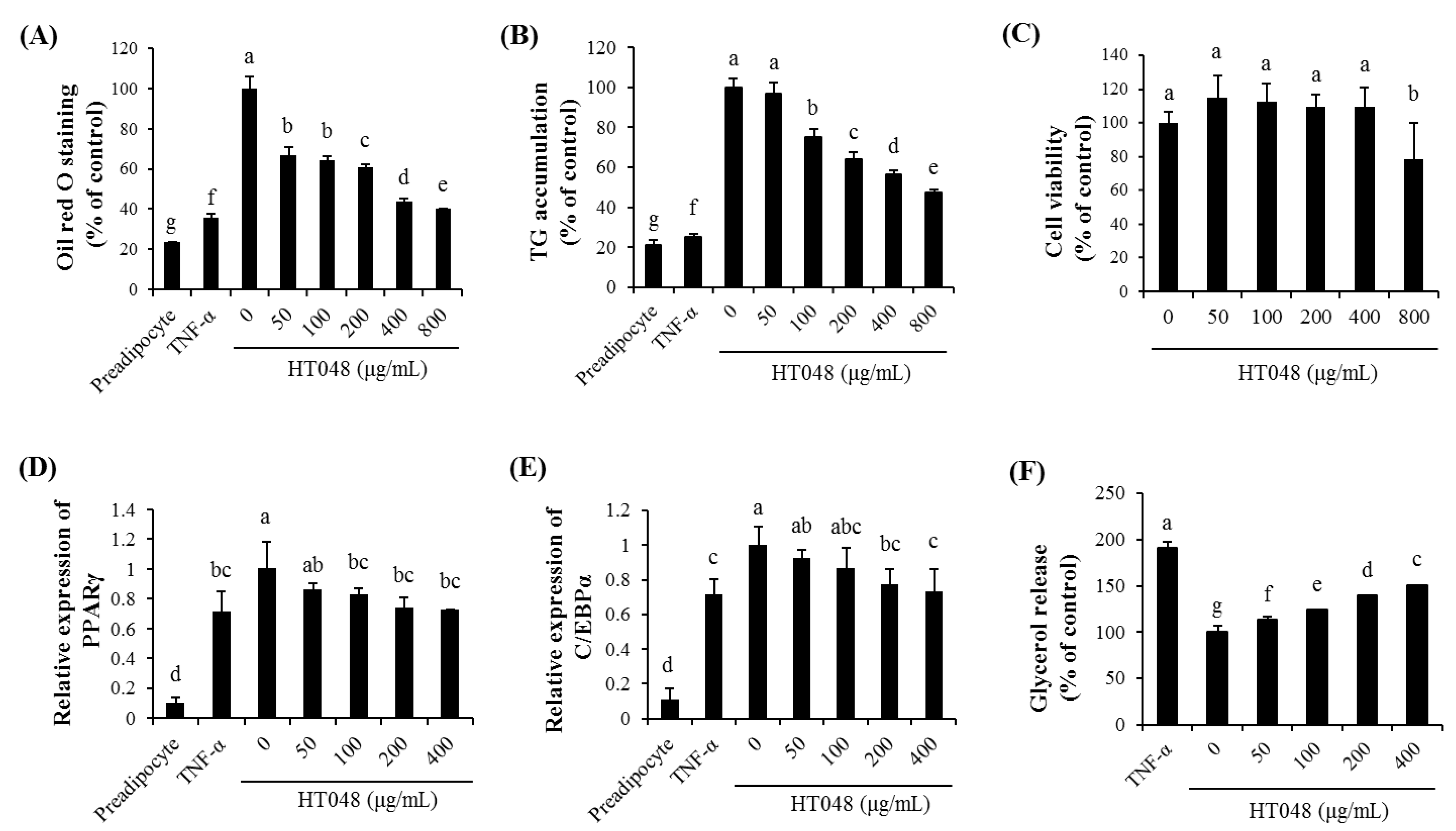
2.2. Effect of HT048 on Adipocyte Differentiation in 3T3-L1 Adipocytes

2.3. Effect of HT048 on the Expression of the Adipogenic Transcription Genes in 3T3-L1 Adipocytes
2.4. Effect of HT048 on Lipolysis in 3T3-L1 Adipocytes
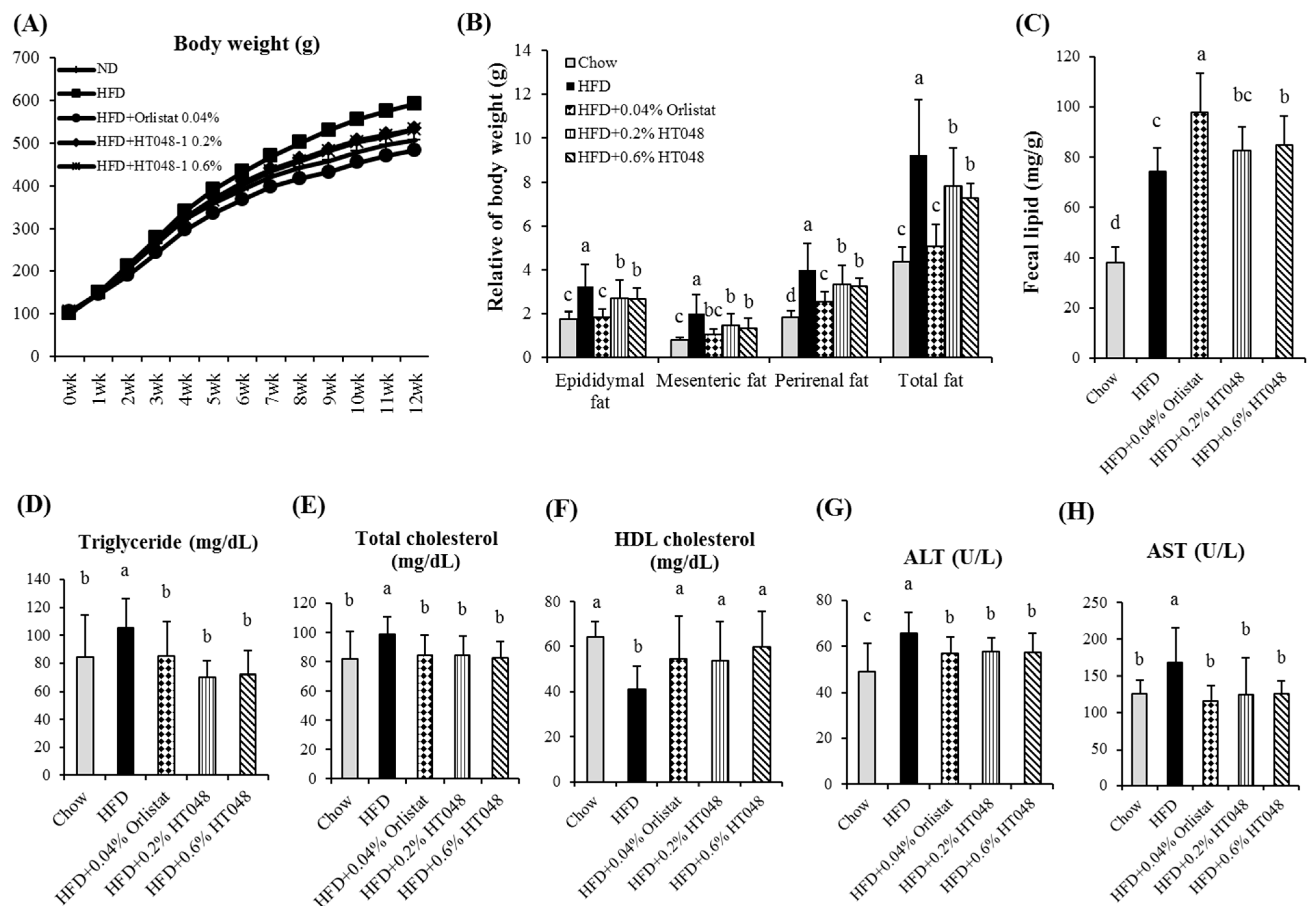
2.5. Effect of HT048 on Body and Organ Weights and Fecal Lipid Content in HFD-Induced Obese Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chow | HFD | HFD0.04% Orlistat | HFD0.2% HT048 | HFD0.6% HT048 | |
|---|---|---|---|---|---|
| Food intake (g/day) | 22.07 ± 0.68 a | 18.90 ± 0.21 b | 20.80 ± 1.97 a | 18.18 ± 0.36 b | 18.17 ± 1.26 b |
| Initial body weight (g) | 91.83 ± 5.37 | 91.92 ± 6.52 | 91.67 ± 4.48 | 91.92 ± 6.64 | 91.75 ± 6.92 |
| Final body weight (g) | 506.92 ± 26.37 bc | 592.42 ± 61.47 a | 483.83 ± 42.45 c | 533.75 ± 63.21 b | 532.00 ± 47.21 b |
2.6. Effect of HT048 on Serum Measurements in HFD-Induced Obese Rats

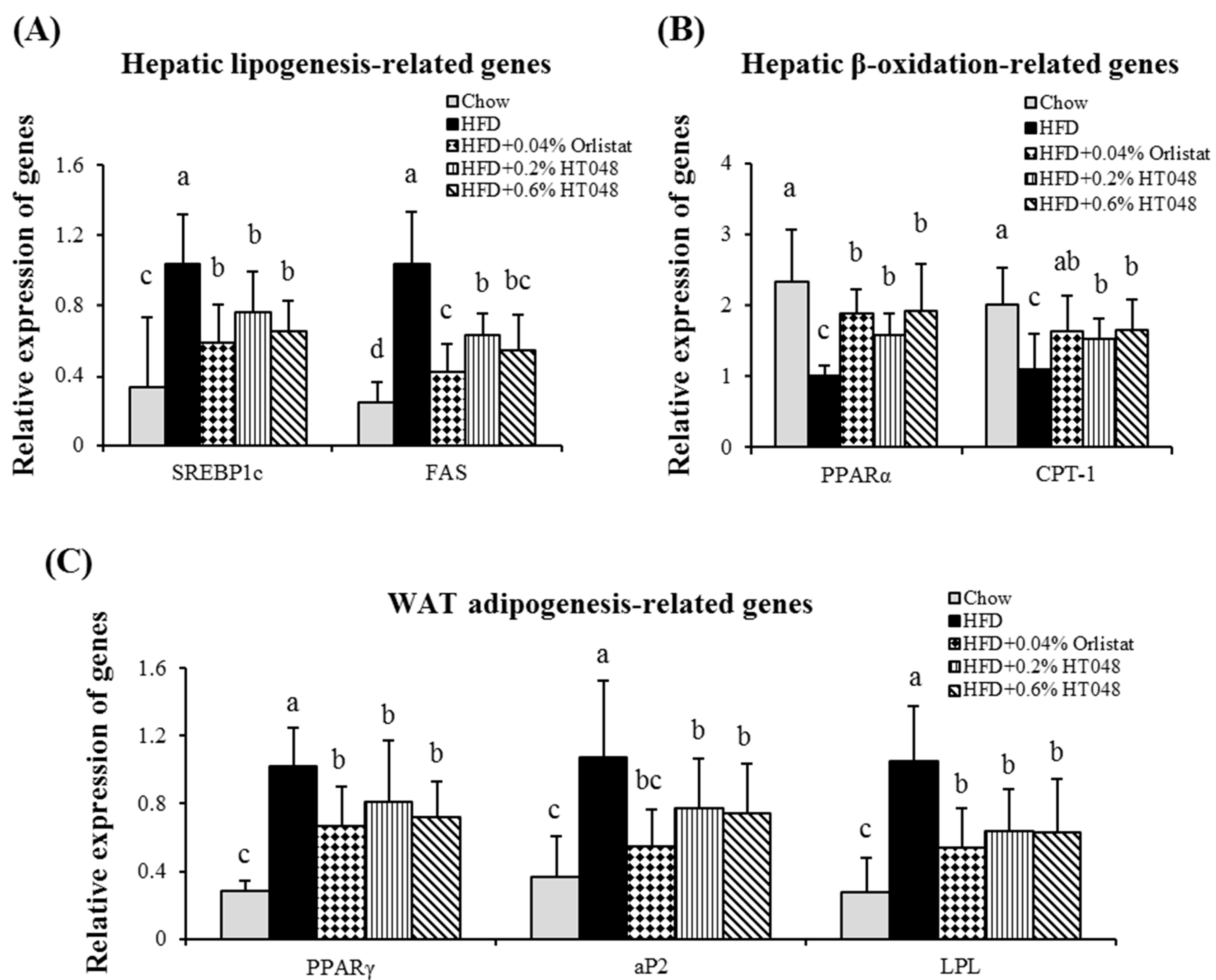
2.7. Effect of HT048 on Hepatic Expression of Lipogenesis-Related Genes in HFD-Induced Obese Rats
2.8. Effect of HT048 on Adipose Tissue Expression of Adipogenesis-Related Genes in HFD-Induced Obese Rats

2.9. Discussion
3. Experimental Section
3.1. Preparation of HT048
3.2. HPLC Analysis of HT048
3.3. Cell Culture and Adipocyte Differentiation
3.4. Cell Viability Assay
3.5. Measurement of TG Accumulation and Glycerol in 3T3-L1
3.6. Oil Red O Staining
3.7. Animal Experiments
3.8. Serum Measurements
3.9. Analysis of Fecal Lipids
3.10. Real-Time Quantitative PCR Analysis
3.11. Statistical Analysis
4. Conclusions
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Bray, G.A.; Bellanger, T. Epidemiology, trends, and morbidities of obesity and the metabolic syndrome. Endocrine 2006, 29, 109–117. [Google Scholar] [CrossRef]
- Gurevich-Panigrahi, T.; Panigrahi, S.; Wiechec, E.; Los, M. Obesity: Pathophysiology and clinical management. Curr. Med. Chem. 2009, 16, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Clasen, M.E.; Donnelly, J.E. Management of dyslipidemia in adults. Am. Fam. Physician 1998, 57, 2192–2204. [Google Scholar] [PubMed]
- Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Hasty, A.H.; Osuga, J.; Tamura, Y.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; et al. Sterol regulatory element-binding protein-1 as a key transcription factor for nutritional induction of lipogenic enzyme genes. J. Biol. Chem. 1999, 274, 35832–35839. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Molecular regulation of adipogenesis. Ann. Rev. Cell. Dev. Biol. 2000, 16, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Heck, A.M.; Yanovski, J.A.; Calis, K.A. Orlistat, a new lipase inhibitor for the management of obesity. Pharmacotherapy 2000, 20, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A. Drug treatment of obesity. Rev. Endocr. Metab. Disord. 2001, 2, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Zuo, Z.; Harrison, F.; Chow, M.S. Hawthorn. J. Clin. Pharmacol. 2002, 42, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Kuo, D.H.; Yeh, C.H.; Shieh, P.C.; Cheng, K.C.; Chen, F.A.; Cheng, J.T. Effect of shanzha, a chinese herbal product, on obesity and dyslipidemia in hamsters receiving high-fat diet. J. Ethnopharmacol. 2009, 124, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Assini, J.M.; Mulvihill, E.E.; Huff, M.W. Citrus flavonoids and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Cha, B.Y.; Choi, S.S.; Choi, B.K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Nobiletin improves obesity and insulin resistance in high-fat diet-induced obese mice. J. Nutr. Biochem. 2013, 24, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Song, M.; Park, J.; Park, S.; Kim, N.; Gaire, B.; Choi, H.-Y.; Kim, H. Anti-obesity effect of HT048, a herbal combination, in high fat diet-induced obese rats. Molecules 2012, 17, 14765–14777. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D’Alessio, D.; Tso, P. A controlled high-fat diet induces an obese syndrome in rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [PubMed]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell. Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-W.; Kim, S. Ginsenoside rc promotes anti-adipogenic activity on 3T3-L1 adipocytes by down-regulating C/EBPα and PPARγ. Molecules 2015, 20, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, D.; Kim, H.; Kim, J. Anti-lipase and lipolytic activities of ursolic acid isolated from the roots of actinidia arguta. Arch. Pharm. Res. 2009, 32, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Fève, B. Adipogenesis: Cellular and molecular aspects. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell. Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Moller, D.E.; Flier, J.S. Insulin resistance—Mechanisms, syndromes, and implications. New Engl. J. Med. 1991, 325, 938–948. [Google Scholar] [PubMed]
- Paccaud, F.; Schlüter-Fasmeyer, V.; Wietlisbach, V.; Bovet, P. Dyslipidemia and abdominal obesity: An assessment in three general populations. J. Clin. Epidemiol. 2000, 53, 393–400. [Google Scholar] [CrossRef]
- Modan, M.; Halkin, H.; Almog, S.; Lusky, A.; Eshkol, A.; Shefi, M.; Shitrit, A.; Fuchs, Z. Hyperinsulinemia. A link between hypertension obesity and glucose intolerance. J. Clin Invest. 1985, 75, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, H.; Zechner, R.; Zimmermann, R. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 2006, 281, 40236–40241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Huang, B.; Choi, S.K.; Seo, J.S. Anti-obesity effect of resveratrol-amplified grape skin extracts on 3T3-L1 adipocytes differentiation. Nutr. Res. Pract. 2012, 6, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. Srebps: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A. Control of macrophage activation and function by ppars. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Schoonjans, K.; Staels, B.; Auwerx, J. The peroxisome proliferator activated receptors (PPARs) and their effects on lipid metabolism and adipocyte differentiation. Biochim. Biophys. Acta 1996, 26, 93–109. [Google Scholar] [CrossRef]
- Son, Y.; Nam, J.S.; Jang, M.K.; Jung, I.A.; Cho, S.I.; Jung, M.H. Antiobesity activity of vigna nakashimae extract in high-fat diet-induced obesity. Biosci. Biotechnol. Biochem. 2013, 77, 332–338. [Google Scholar] [PubMed]
- Zhang, Y.; Fan, S.; Hu, N.; Gu, M.; Chu, C.; Li, Y.; Lu, X.; Huang, C. Rhein reduces fat weight in db/db mouse and prevents diet-induced obesity in c57bl/6 mouse through the inhibition of ppargamma signaling. PPAR Res. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, L.; Cai, W.; Fan, S.; Feng, L.; Ji, G.; Huang, C. Protopanaxatriol, a novel ppargamma antagonist from panax ginseng, alleviates steatosis in mice. Sci. Rep. 2014, 9. [Google Scholar] [CrossRef]
- Foster, D.W. The role of the carnitine system in human metabolism. Ann. N.Y. Acad. Sci. 2004, 1033, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fielding, B.A.; Frayn, K.N. Lipoprotein lipase and the disposition of dietary fatty acids. Br. J. Nutr. 1998, 80, 495–502. [Google Scholar] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Samples Availability: Milligram quantities of HT048 is available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.H.; Kim, Y.-S.; Song, M.; Lee, M.; Park, J.; Kim, H. A Herbal Formula HT048, Citrus unshiu and Crataegus pinnatifida, Prevents Obesity by Inhibiting Adipogenesis and Lipogenesis in 3T3-L1 Preadipocytes and HFD-Induced Obese Rats. Molecules 2015, 20, 9656-9670. https://doi.org/10.3390/molecules20069656
Lee YH, Kim Y-S, Song M, Lee M, Park J, Kim H. A Herbal Formula HT048, Citrus unshiu and Crataegus pinnatifida, Prevents Obesity by Inhibiting Adipogenesis and Lipogenesis in 3T3-L1 Preadipocytes and HFD-Induced Obese Rats. Molecules. 2015; 20(6):9656-9670. https://doi.org/10.3390/molecules20069656
Chicago/Turabian StyleLee, Yoon Hee, Young-Sik Kim, Mikyung Song, Minsu Lee, Juyeon Park, and Hocheol Kim. 2015. "A Herbal Formula HT048, Citrus unshiu and Crataegus pinnatifida, Prevents Obesity by Inhibiting Adipogenesis and Lipogenesis in 3T3-L1 Preadipocytes and HFD-Induced Obese Rats" Molecules 20, no. 6: 9656-9670. https://doi.org/10.3390/molecules20069656