2.1. Total Phenol Content and Phenolic Profile of Extracts
Hexane extraction was performed prior to extraction with 80% methanol to remove waxes and other highly lipophilic substances from the berry materials. The yield of the hexane extracts gave a median value of 1.0% of the dried berries. Then 80% aqueous methanol was used since aqueous alcohol is a good solvent for the extraction of phenolics [
11].
Table 1 shows the phenolic content of all berry extracts.
Crowberries, chokeberries and bog whortleberries were found to have the highest content of phenolics both in the methanol ASE extract and in the fresh berries. Purification by SPE resulted in extracts that contained from 5 to 25 times (average 8.8) more total phenolics compared to the ASE extracts. In addition to crowberry, chokeberry and bog whortleberry, the cloudberry methanol SPE extract also showed a high phenolic content. The results are comparable with literature values [
12]. The Folin–Ciocalteu reagent was used for quantifying the phenolic content in the berry extracts. The presence of other easily oxidizable substances, which are not phenolic compounds, such as sugars, ascorbic acid and vitamins may also react with the Folin–Ciocalteu reagent, causing an overestimation of total phenolic content [
13], particularly in the ASE extracts before SPE purification. This study shows a pronounced difference in phenolic content between wild bilberries (
Vaccinium myrtillus) (533 mg gallic acid equivalents (GAE)/100 g fresh weight) and the cultivated blueberries (
Vaccinium corymbosum) (211 mg GAE/100 g).
V. corymbosum do not have a long tradition in the Nordic diet but is gaining increased popularity among consumers.
NMR spectroscopy was employed to identify the main phenolic constituents of the extracts, together with data of the already known major phenolics of the berries. An advantage with
1H-NMR is that organic molecules containing hydrogen atoms will give signals in the spectrum at positions dependent on the surroundings of each hydrogen atom, and the area of the signals depends on the number of atoms giving rise to each signal and makes it possible to detect e.g., anthocyanins, flavonols, organic acids and sugars in the same spectrum. NMR spectroscopy also has the advantage of being a reproducible, non-destructive, robust and fast method, with no need for chromatographic separation ahead of the analysis [
14]. The methanol ASE extracts were dominated by signals from carbohydrates (δ 3–4 ppm, anomeric proton doublets δ 4.4–5.4 ppm) and signals from organic acids such as citric acid and malic acid (δ 2.6–2.9 ppm). The
1H-NMR spectra of the SPE methanol extracts showed that the intensity of the sugar signals was much reduced comparative to other signals in the spectra, an indication that SPE purification had removed free sugars, resulting in extracts enriched in aromatic compounds. The major phenolic constituents in each SPE extract were identified by comparison with NMR literature values. The presence of the phenolic compounds in each berry extract was further confirmed by assessment of the literature (
Table 2). A large number of published works describe the content of anthocyanins and other types of flavonoids and phenolics in the berries used [
15,
16,
17,
18,
19,
20,
21].
Anthocyanins were predominant in the dark coloured berry extracts and the aglycones were identified mainly based on the B-ring signals [
40,
41]. Cyanidins were identified by their H-6’ dd signals at δ 8.2–8.3 ppm and H-2’ doublet at δ 8.0–8.1 ppm, respectively, delphinidins by their H-2’ and H-6’ singlet at δ 7.7–7.8 ppm and malvidins with an H-2’ and H-6’ singlet at δ 7.9 ppm. The number of signals at δ ca. 9.0 ppm attributed to H-4 indicated the number of anthocyanins present in each extract, and the shift value at δ ca. 9.0 ppm indicated that anthocyanins were present in the flavylium form, as well. A few berry extracts (bog whortleberry, elderberry and sea buckthorn) were rich in flavonol glycosides. Here, too, the B-ring signals were used to distinguish between the aglycones [
42]. Quercetin, myricetin and isorhamnetin glycosides give meta-coupled doublets with the H-6 and H-8 signals (A-ring protons) at ca. δ 6.2 and 6.4 ppm and with a coupling constant of 2 Hz. The B-ring signals differed: quercetin (7.7–7.8 ppm, d,
J = 2 Hz, H-2’; 6.8 ppm, d,
J = 8 Hz, H-5’; 7.6 ppm, dd,
J = 8, 2 Hz, H-6’), myrcetin (7.3–7.4 ppm, s, H-2’ and H-6’) and isorhamnetin (7.9 ppm, d,
J = 2 Hz, H-2’; 6.9 ppm, d,
J = 8 Hz, H-5’; 7.6 ppm, dd,
J = 8, 2 Hz, H-6’; 3.95 ppm, s, -OCH3). Chlorogenic acids, characterized by the olefinic proton signals in the caffeoyl moiety at ca. 6.3 and ca. 7.6 ppm with a coupling constant of 15–16 Hz and quinic acid signals at 1.9–2.2 ppm [
43], were present in high amounts in the rowanberry extract, and somewhat less in black chokeberry, blueberry and elderberry extracts. Cloudberry and raspberry extracts contained high levels of ellagitannins, characterized by the singlets (δ 6.3–6.7 ppm) from protons on the hexahydroxydiphenoyl (HHDP) moieties and highly shielded anomeric proton signals [
44]. Benzoic acids were characterized by their deshielded doublet signals at δ 8.0–8.1 ppm (1H), and two sets of triplets at δ 7.5–7.6 (1H) and 7.4–7.5 ppm (2H) [
35], and were identified in the cloudberry and lingonberry SPE extracts. The
1H-NMR spectra of the SPE berry extracts are presented in the
supplementary materials.
Bilberry SPE Extract: Ten different signals from H-4 in anthocyanins were observed, four were major. This variety in content of anthocyanins gave rise to a complex spectrum with difficulties in identifying specific anthocyanins. However, signals from cyanidin and delphinidin dominated, followed by malvidin, in accordance with the literature [
22,
23].
Blackberry SPE Extract: There were two sets of signals from anthocyanins, with cyanidin 3-glucoside as the major one. One rhamnose methyl signal (δ 1.15,
J = 6 Hz) indicated the presence of cyanidin 3-rutinoside, which is in accordance with the literature [
24,
25].
Black chokeberry SPE Extract: Two major anthocyanins were present, cyanidin 3-galactoside and cyanidin 3-arabinoside, which were previously reported to be the major glycosides. In addition, signals from both chlorogenic acid and neochlorogenic acid were prominent [
19,
21,
26].
Black currant SPE Extract: Signals from two main anthocyanins were observed: delphinidin (major) and cyanidin types. Two sets of signals from the methyl group in rhamnose indicated the presence of delphinidin-3-rutinoside and cyaniding-3-rutinoside [
19,
21,
27].
Blueberry SPE Extract: Similar to the bilberry extract, blueberries gave rise to a crowded spectrum of anthocyanin signals, 15 signals from H-4 in the anthocyanin could be observed. There were no major signals from cyaniding or peonidin (H-6’, dd, δ 8.2–8.3 ppm), and malvidin and delphinidin type anthocyanins were the dominating ones. Chlorogenic acid was the major non-flavonoid present [
21,
27,
28].
Bog whortleberry SPE Extract: The spectrum was dominated by signals from glycosides of the four main aglycones: quercetin, myricetin, malvidin and delphinidin. The H-4 signal from anthocyanins indicated the presence of at least seven anthocyanins, from the B-ring signals maldivin seemed to be the major aglycone (δ 7.92 (s)), which could be due to malvidin-3-glucoside or malvidin-3-galactoside, previously reported to be the major aglycones [
27,
29,
30].
Cloudberry SPE Extract: There were signals from two major compounds, benzoic acid and an ellagitannin. The spectrum contained six characteristic singlets from protons in the HHDP units (δ 6.3–6.7 ppm) of the ellagitannin, a broad singlet at δ 7.02 ppm corresponding to protons in galloyl units of the molecule, and two shielded anomeric proton signals (δ 5.98, d,
J = 8.7 Hz and δ 6.40, d,
J = 4.0 Hz). This is consistent with the literature values for sanguiin H6, a major ellagitannin in cloudberries [
31,
32]. Benzoic acid signals were also observed.
Crowberry SPE Extract: Signals from three major anthocyanins were prominent, they were of delphinidin, malvidin and cyanidin type. Plausible identities are delphinidin-3-galactoside, cyanidin-3-galactoside and malvidin-3-galactoside, according to the literature [
21,
27,
33].
Elderberry SPE extract: The spectrum showed the presence of four major compounds, cyanidin-3-glucoside, cyanidin-3-sambubioside, rutin and chlorogenic acid, which is in accordance with the literature [
19,
27,
34].
Lingonberry SPE Extract: Signals from benzoic acids were the most prominent in the aromatic region, identified as 1-
O-benzoyl-glucose and 6-
O-benzoyl-glucose. Both α (
J = 3.5 Hz) and β (
J = 7.5 Hz) anomeric proton signals were present. Minor cyanidin signals were observed [
21,
27,
35].
Raspberry SPE Extract: The
1H-NMR signals matched with the ellagitannin signals from cloudberries, thus indicating that the major aromatic compound was sanguiin H-6, which is in accordance with a recent review. Minor cyanidin signals were observed [
31,
36,
37].
Red currant SPE Extract: Weak signals in the aromatic area were observed with no dominating phenolics. The aromatic signals were possibly glycosylated phenolic acids, which have previously been reported in these berries [
19]. Traces of cyanidin signals were observed [
19,
21].
Rowanberry SPE Extract: The major aromatic compounds were chlorogenic acid and neochlorogenic acid, present in an approximately 1:1 ratio [
21,
38].
Sea buckthorn SPE Extract: Two sets of isorhamnetin signals dominated the aromatic region of the NMR spectrum. The presence of signals from rhamnose methyl groups at δ 0.9–1.0 ppm indicate isorhamnetin rhamnosides which are common in this species [
21,
27,
39].
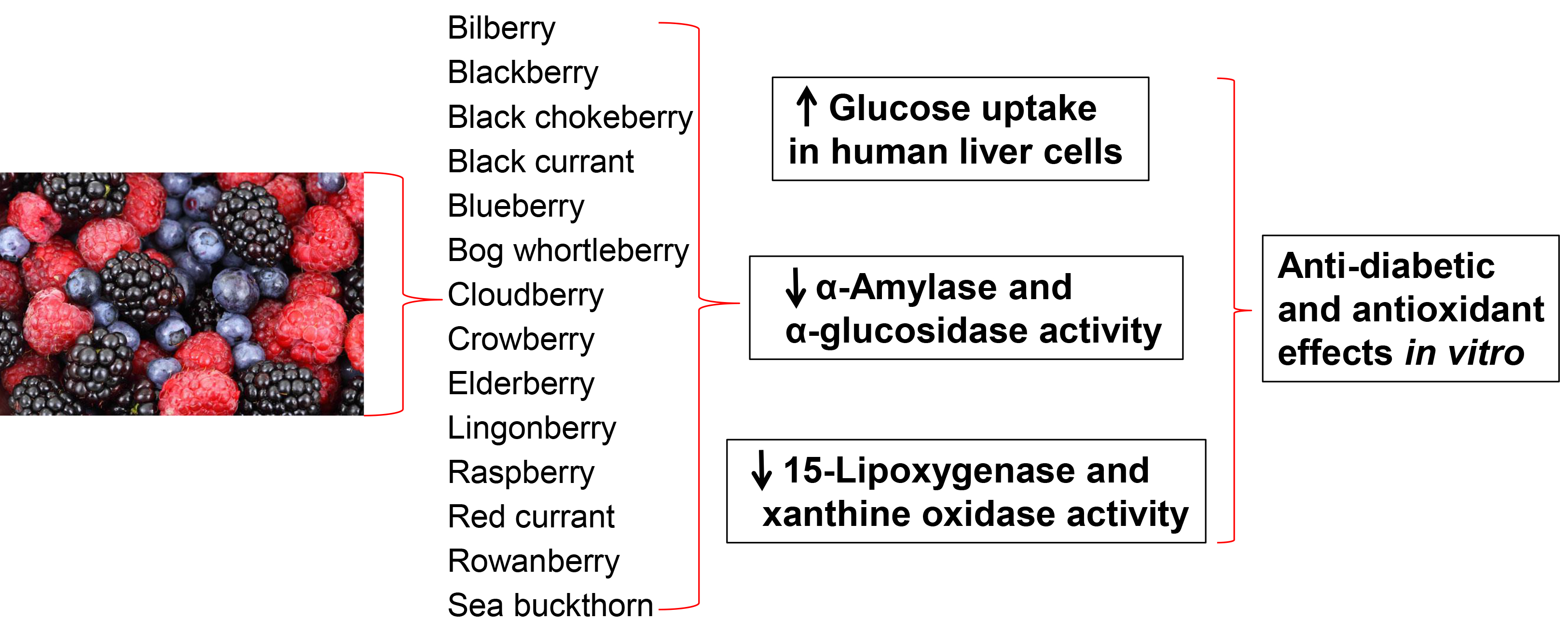
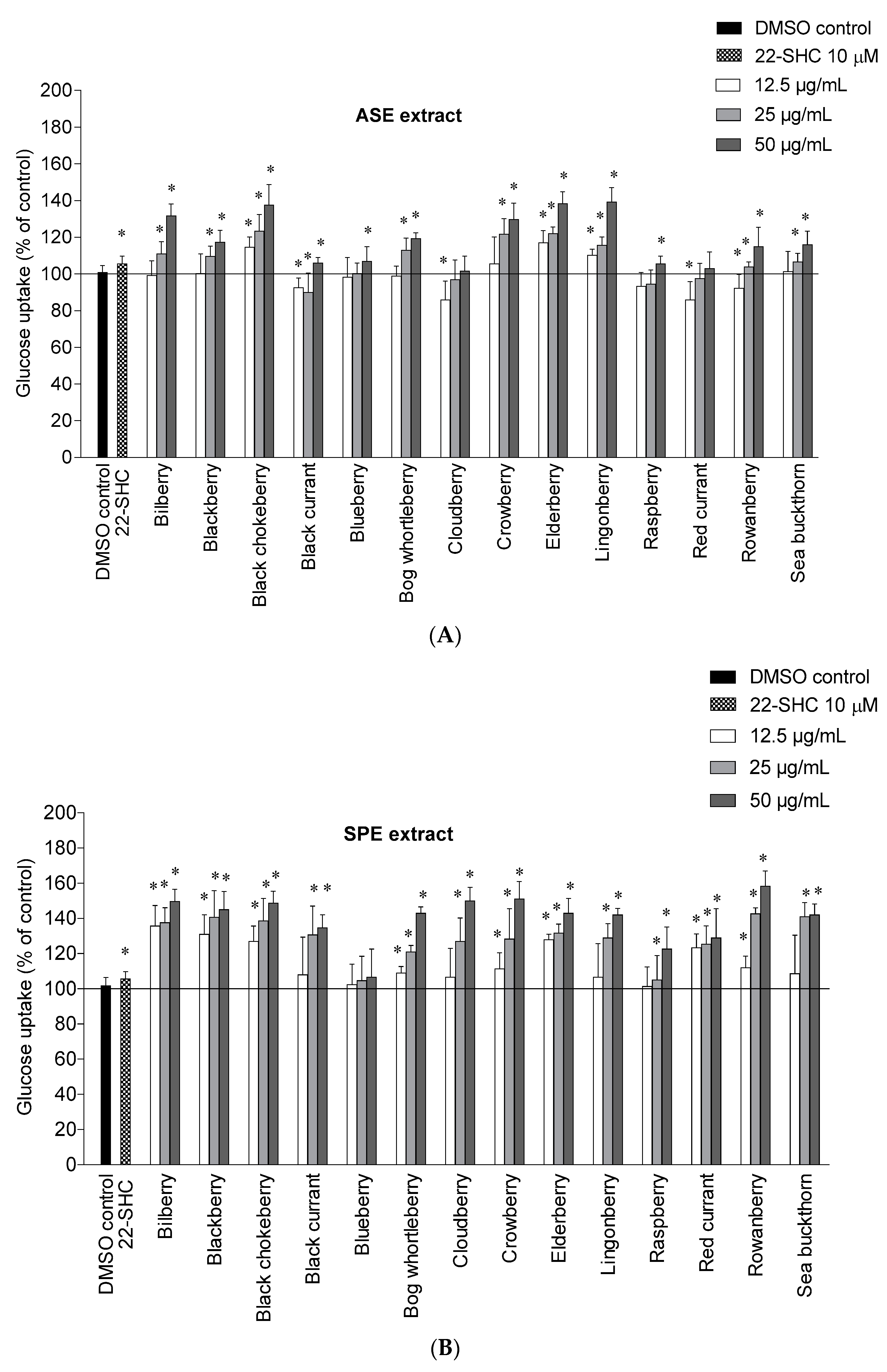
2.2. Glucose Uptake in HepG2-Cells
This study shows for the first time the potential of Nordic berry extracts to stimulate glucose uptake in human liver cells. The ASE and SPE extracts were tested at 12.5, 25 and 50 µg/mL and the results are shown in
Figure 1A,B. The cytotoxicity of the berry extracts on the liver cells was determined by the Bradford protein method [
45]. In this case none of the berry extracts showed a drastic change of protein levels in the cells and were therefore not cytotoxic to the liver cells.
Among the ASE extracts, lingonberry and elderberry showed the highest stimulation of uptake at 50 µg/mL, with an increase of 39.3 ± 7.6% and 38.3 ± 6.5% respectively, followed by black chokeberry (37.7 ± 11.0%), bilberry (31.7 ± 6.5%) and crowberry (29.7 ± 9.0%) compared to 0.1% DMSO control. The bilberry, blackberry, bog whortleberry, crowberry, rowanberry and sea buckthorn ASE extracts showed a significant increase only at 25 µg/mL and 50 µg/mL, while the black currant, blueberry and raspberry ASE extracts only showed a significant increase at 50 µg/mL. For all berries, the glucose uptake was higher with the SPE extracts compared to the ASE extracts. It seems that higher content of polyphenols and simpler phenolics are important for the mode of action.
A high content of anthocyanins seems to be important for glucose uptake in several of the berries. The anthocyanin rich extracts of bilberry, blackberry, black chokeberry, bog whortleberry, crowberry and elderberry significantly increased the glucose uptake at 25 and 50 µg/mL for the ASE extracts and at all tested concentrations for the SPE extracts. The importance of anthocyanins is supported by other studies showing that anthocyanins can stimulate the glucose uptake in liver and skeletal muscle cells [
46,
47]. The rowanberry SPE extract showed the highest maximal efficacy on glucose uptake at 50 µg/mL with an increase of 58.3 ± 8.5%, followed by the crowberry SPE extract (51.0 ± 9.8%), the cloudberry SPE extract (50.0 ± 7.5%), and the bilberry SPE extract (49.7 ± 6.8%). There does not seem to be one single constituent that contributes to the effect, rather a number of phenolic substances can stimulate the uptake as we have recently described [
46,
48]. Chlorogenic acids likely play an important role in the uptake of glucose observed for rowanberry, and might also be involved in the high activity observed for black chokeberry and elderberry in addition to cyanidin glycosides [
48]. Ellagitannins present in cloudberry and raspberry (
Table 2), and cyanidin glycosides present in crowberry might be important contributors in these berries.
Benzoic acid, which is a major constituent of the cloudberry SPE extract, has in a previous study shown a small increase of glucose uptake in human myotubes at 10 µM [
48]. The bilberry SPE extract contains a complex mixture of delphinidin-, cyanidin- and malvidin glycosides, and their individual contribution to the glucose uptake is difficult to predict. There is a need for more studies to investigate how the different anthocyanin aglycones contribute to this effect. The SPE extracts from blackberry, black chokeberry, bog whortleberry, elderberry, lingonberry and sea buckthorn showed a significant increase of glucose uptake in the range of 42.0–48.7% at the highest tested concentration (50 µg/mL) compared to DMSO control. At the lowest concentration (12.5 µg/mL) the anthocyanin rich extracts bilberry SPE extract, the blackberry SPE extract and the elderberry SPE extract were the most potent with increased glucose uptake of 35.7 ± 11.6%, 31.0 ± 11.0% and 28.0 ± 3.0%, respectively. Quercetin and isorhamnetin glycosides from the flavonol-rich extracts from elderberry, sea buckthorn and bog whortleberry might also contribute to the glucose stimulation [
48]. The high content of benzoyl glycosides in lingonberry and phenolic acids in red currant might also play a major role in the uptake of glucose in HepG2-cells. The sea buckthorn SPE extract showed the lowest phenolic content (
Table 1), but still showed a high increase of glucose uptake compared to the other SPE extracts. It seems that the type of phenolic is more important for the uptake of glucose than the total phenolic content. In view of the potent effect of cloudberry and lingonberry SPE extracts rich in benzoic acids or its derivatives, benzoic acid derivatives should be subjects for further studies, in addition to isolated anthocyanins and flavonols. The enhanced uptake of glucose observed for these berry extracts in the liver cells might be effective in the skeletal muscle cells as well. We have previously seen that the stimulation of glucose uptake by the phenolics is similar in both the liver cells and the skeletal muscle cells [
48].
2.3. Inhibition of α-Amylase and α-Glucosidase
Small intestinal α-glucosidase and pancreatic α-amylase are key enzymes of dietary carbohydrate digestion in humans. Inhibitors of these enzymes by phenolic rich extracts may offer a natural dietary approach to prevent T2D because it will be effective in retarding carbohydrate digestion and glucose absorption to suppress postprandial hyperglycemia. The ASE extracts of the 14 Nordic berries were all potent inhibitors of α-amylase with IC
50 values ranging from 6.3 to 21.4 µg/mL (
Table 3).
Among the ASE extracts, the crowberry (IC
50 6.3 ± 0.8 µg/mL) and the bog whortleberry (IC
50 9.1 ± 3.5 µg/mL) showed the highest α-amylase activity, followed by cloudberry (IC
50 9.6 ± 0.7 µg/mL). The SPE extracts appeared to have equal or somewhat higher α-amylase inhibitory activity compared to ASE extracts, with the anthocyanin-rich crowberry (5.3 ± 0.9 µg/mL) and black chokeberry (6.0 ± 1.0 µg/mL) extracts being the most active ones. Regarding the α-glucosidase inhibitory activity of the SPE extracts, the crowberry, the chokeberry and the cloudberry possessed the highest inhibitory activity, with IC
50 values of 10.9 ± 1.1, 12.0 ± 1.7 and 13.6 ± 1.3 µg/mL, respectively. All the tested ASE and SPE extracts were more potent than the acarbose used as control compound (IC
50 84.7 ± 3.8 µg/mL for α-glucosidase, IC
50 73.3 ± 4.3 µg/mL for α-amylase). Bilberry, black currant, blackberry, black chokeberry, red currant and lingonberry have been reported to be potent α-amylase inhibitors [
49,
50], while the inhibition of α-glucosidase has been reported for many of the berries, except for bog whortleberry, sea buckthorn and cloudberry [
49,
51,
52]. It has been reported that berry anthocyanins and ellagitannins play an important role in the inhibition of α-amylase and α-glucosidase, respectively [
8]. This study indicates that ellagitannins might contribute to a higher activity towards α-glucosidase since the raspberry and cloudberry were among the most active extracts against this enzyme. Since cloudberry and raspberry extracts contain the ellagitannin sanguiin-6, the possibility of the precipitation of enzyme–tannin complexes cannot be ruled out [
53]. However, no such precipitation was observed. The other extracts appear not to contain high amounts of tannins. Furthermore, precipitation at neutral or basic pH values would not be expected [
54,
55]. Anthocyanin rich extracts were most active as α-glucosidase inhibitors. Rowanberry, with its high content of chlorogenic acids, was also highly active in this assay. As we have recently shown [
46,
48], chlorogenic and neochlorogenic acids, anthocyanins and flavonoids are potent inhibitors of α-amylase and α-glucosidase and contribute to the observed effects.

 ,
, 


{kind=link}
{kind=link}