Exploratory Cortex Metabolic Profiling Revealed the Sedative Effect of Amber in Pentylenetetrazole-Induced Epilepsy-Like Mice

Abstract
:
1. Introduction
2. Results and Discussion
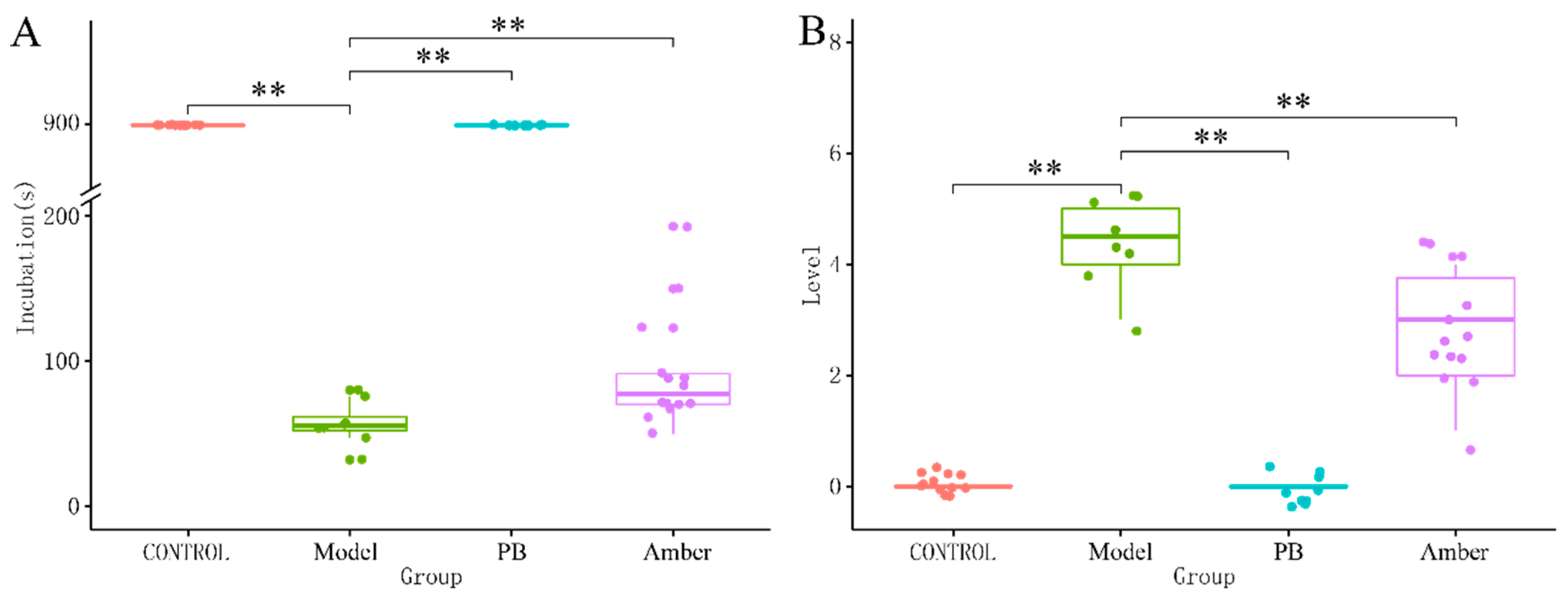
2.1. Behavioral Analysis
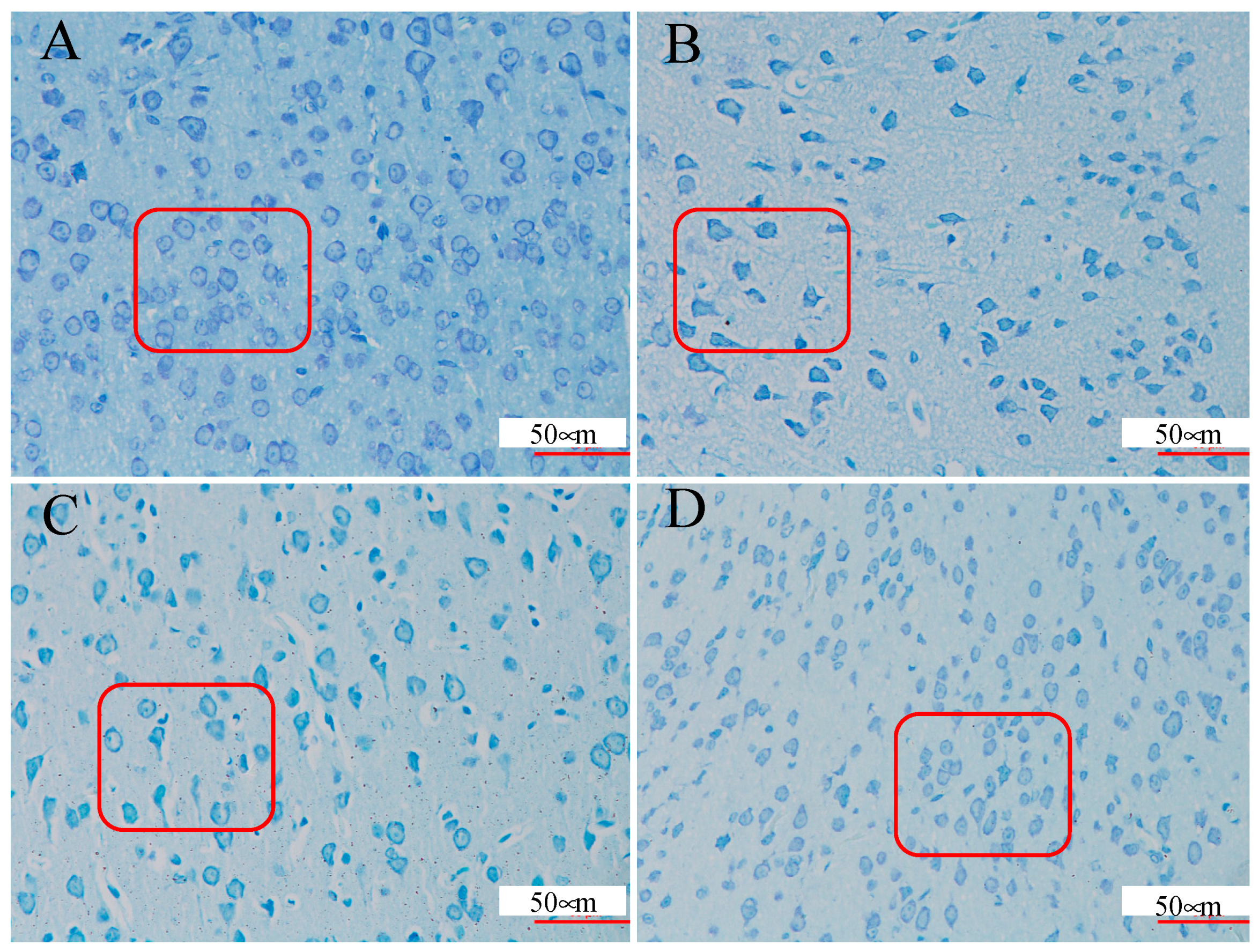
2.2. Brain Tissue Nissl Staining
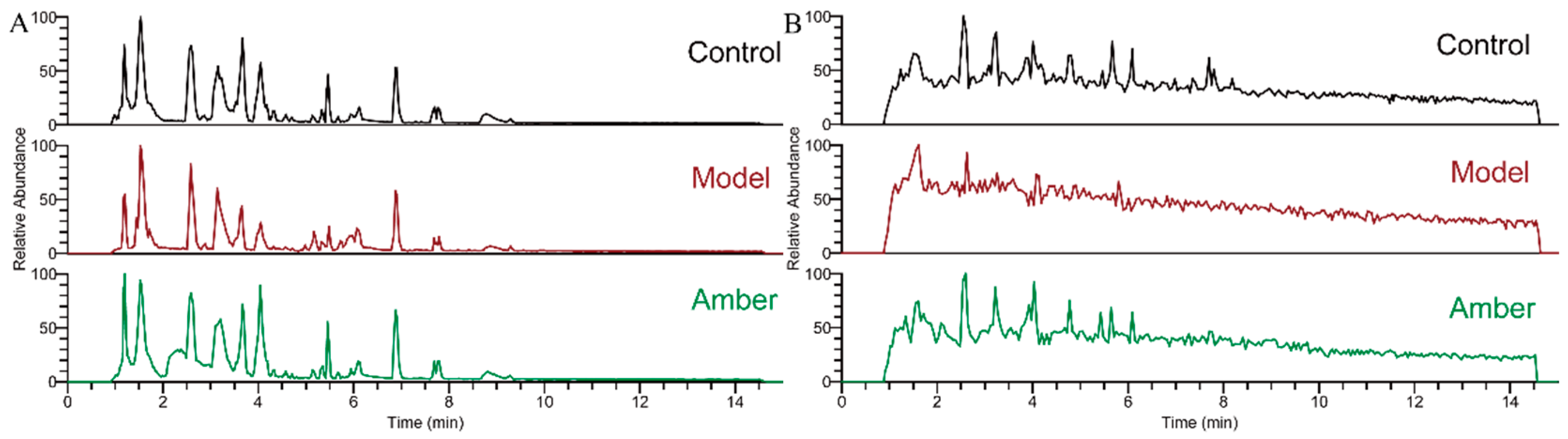
2.3. Analysis of Metabolite Profiling
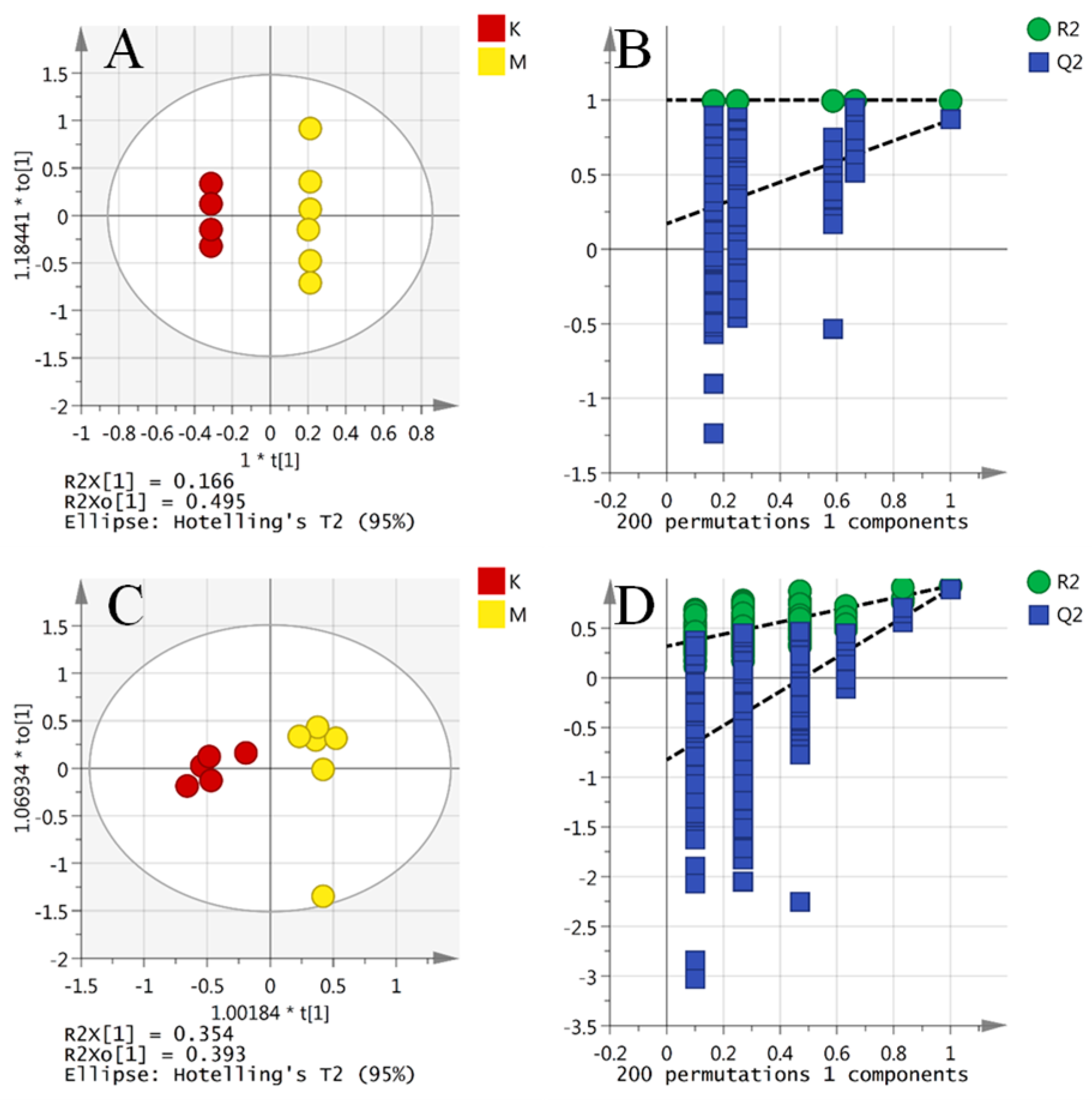
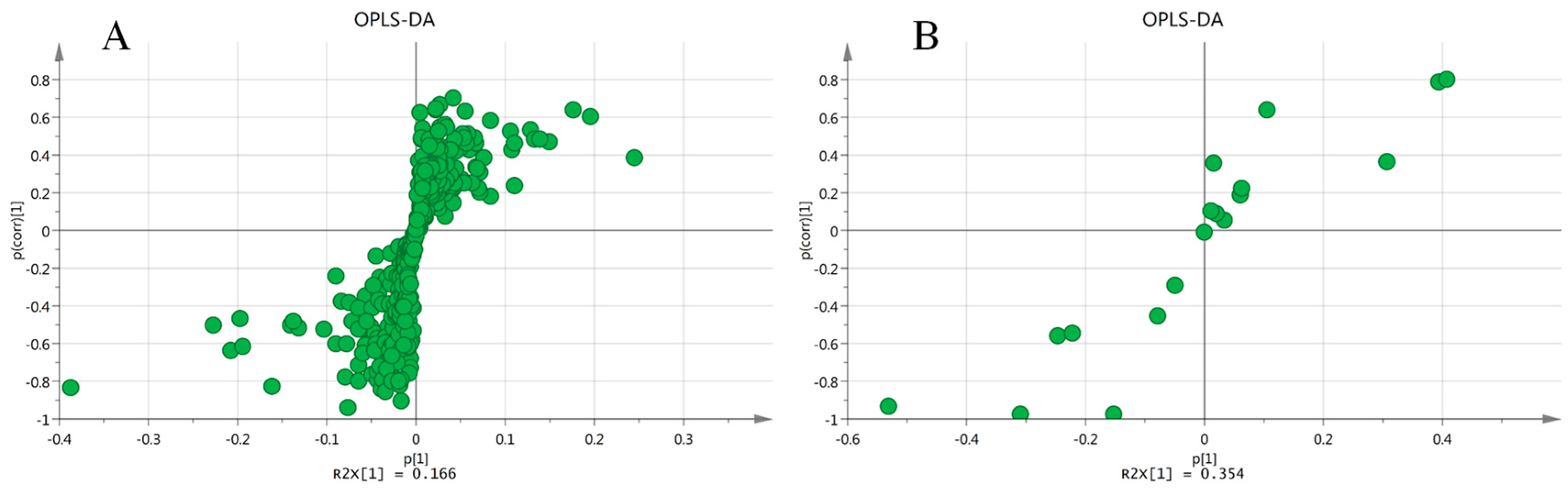
2.4. Multivariate Data Analysis
2.5. Comparison of Model Against Control Using OPLS-DA
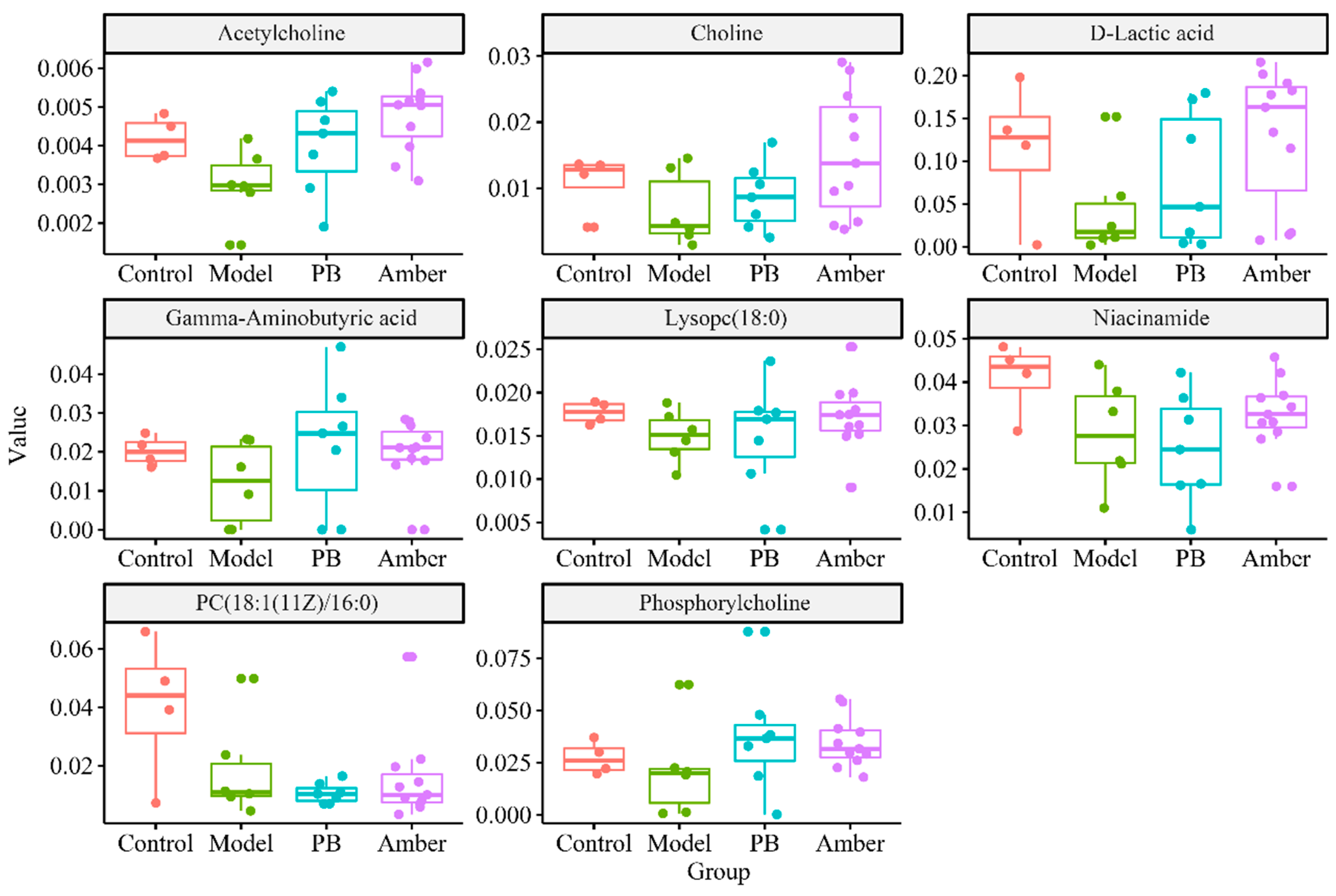
2.6. Screening and Identification of Metabolic Differences
2.7. Metabolic Pathway Analysis
3. Materials and Methods
3.1. Materials and Extract Preparation
3.2. In Vivo Experiments Protocol
3.3. UHPLC–LTQ–Orbitrap MS Analyses
3.4. Data Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wei, C.; Li, Y.; Yao, H.; Liu, H.; Zhang, X.; Guo, R. A metabonomics study of epilepsy in patients using gas chromatography coupled with mass spectrometry. Mol. Biosyst. 2012, 8, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Carpay, J.A.; Ferrari, M.D. The adverse effects of antiepileptic drugs differ in patients with migraine. Lancet Neurol. 2012, 11, 935. [Google Scholar] [CrossRef]
- Wolf, P. Acute drug administration in epilepsy: A review. CNS Neurosci. Ther. 2011, 17, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Tomson, T.; Battino, D.; Bonizzoni, E.; Craig, J.; Lindhout, D.; Sabers, A.; Perucca, E.; Vajda, F. Dose-dependent risk of malformations with antiepileptic drugs: An analysis of data from the EURAP epilepsy and pregnancy registry. Lancet Neurol. 2011, 10, 609–617. [Google Scholar] [CrossRef]
- Chen, B.; Choi, H.; Hirsch, L.J.; Katz, A.; Legge, A.; Buchsbaum, R.; Detyniecki, K. Psychiatric and behavioral side effects of antiepileptic drugs in adults with epilepsy. Epilepsy Behav. 2017, 76, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Gaitatzis, A.; Sander, J.W. The long-term safety of antiepileptic drugs. CNS Drugs 2013, 27, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Sada, N.; Lee, S.; Katsu, T.; Otsuki, T.; Inoue, T. Epilepsy treatment. Targeting LDH enzymes with a stiripentol analog to treat epilepsy. Science 2015, 347, 1362–1367. [Google Scholar] [CrossRef]
- Malkki, H. Epilepsy-burning questions and emerging therapies. Nat. Rev. Neurol. 2014, 10, 243. [Google Scholar] [CrossRef]
- Wu, L.; Liu, S.; Wu, D.; Yu, B.; Wang, Y.; Wang, R.; Yang, J. Intervention Study of Mineral Chinese Medicine Chloriti Lapis in PTZ-kindled Epileptic Rat. J. Chinese Med. Mater. 2016, 39, 155–159. [Google Scholar]
- Zhang, X. Observation on treating insomnia following orthopedics surgery with the Hu’po. Anshen Decoct. 2015, 7, 114–115. (In Chinese) [Google Scholar]
- Sun, X. Clinical observation of hupodingxian capsule in treatment of epilepsy in 94 cases. Xinjiang J. Trad. Chinese Med. 2007, 25, 39–40. (In Chinese) [Google Scholar]
- Zou, S. 57 cases of primary epilepsy treated by DianXianMi Pill. Jilin J. Chinese Med. 1993, 1, 21. (In Chinese) [Google Scholar]
- Lambert, J.B.; Poinar, G.O. Amber: The organic gemstone. Accounts Chem. Res. 2002, 35, 628–636. [Google Scholar] [CrossRef]
- Nohra, Y.A.; Perrichot, V.; Jeanneau, L.; Le Polles, L.; Azar, D. Chemical Characterization and Botanical Origin of French Ambers. J. Nat. Prod. 2015, 78, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Otto, A.; Krumbiegel, G.; Simoneit, B. The natural product biomarkers in succinite, glessite and stantienite ambers from Bitterfeld, Germany. Rev. Palaeobot. Palyno. 2006, 140, 27–49. [Google Scholar] [CrossRef]
- Anderson, K.B.; Winans, R.E.; Botto, R.E. The nature and fate of natural resins in the geosphere—II. Identification, classification and nomenclature of resinites. Org. Geochem. 1992, 18, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Koshino, H.; Takahashi, S.; Shimizu, E.; Takahashi, H.; Yoshida, J.; Shinden, H.; Tsujimura, M.; Kofujita, H.; Uesugi, S.; et al. Ca2+-Signal Transduction Inhibitors, Kujiol A and Kujigamberol B, Isolated from Kuji Amber Using a Mutant Yeast. J. Nat. Prod. 2018, 81, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kobayashi, M.; Okawa, Y.; Inui, T.; Yoshida, J.; Higashio, H.; Shinden, H.; Uesugi, S.; Koshino, H.; Kimura, K. Yeast Ca2+-signal transduction inhibitors isolated from Dominican amber prevent the degranulation of RBL-2H3 cells through the inhibition of Ca2+-influx. Fitoterapia 2016, 113, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Funaba, M.; Ikeda, T.; Abe, M. Degranulation in RBL-2H3 cells: Regulation by calmodulin pathway. Cell Biol. Int. 2003, 27, 879–885. [Google Scholar] [CrossRef]
- Maruyama, M.; Kobayashi, M.; Uchida, T.; Shimizu, E.; Higashio, H.; Ohno, M.; Uesugi, S.; Kimura, K. Anti-allergy activities of Kuji amber extract and kujigamberol. Fitoterapia 2018, 127, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Spagou, K.; Wilson, I.D.; Masson, P.; Theodoridis, G.; Raikos, N.; Coen, M.; Holmes, E.; Lindon, J.C.; Plumb, R.S.; Nicholson, J.K.; et al. HILIC-UPLC-MS for exploratory urinary metabolic profiling in toxicological studies. Anal. Chem. 2011, 83, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Cruz, T.; Gleizes, M.; Balayssac, S.; Mornet, E.; Marsal, G.; Millan, J.L.; Malet-Martino, M.; Nowak, L.G.; Gilard, V.; Fonta, C. Identification of altered brain metabolites associated with TNAP activity in a mouse model of hypophosphatasia using untargeted NMR-based metabolomics analysis. J. Neurochem. 2017, 140, 919–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Want, E.J.; O’Maille, G.; Smith, C.A.; Brandon, T.R.; Uritboonthai, W.; Qin, C.; Trauger, S.A.; Siuzdak, G. Solvent-dependent metabolite distribution, clustering, and protein extraction for serum profiling with mass spectrometry. Anal. Chem. 2006, 78, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Wang, G.; Ran, N.; Lin, H.; Wang, Z.; Guan, X.; Yuan, Y.; Fang, K.; Liu, J.; Wang, F. Inhibitory Effect of Methotrexate on Rheumatoid Arthritis Inflammation and Comprehensive Metabolomics Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole Time of Flight-Mass Spectrometry (UPLC-Q/TOF-MS). Int. J. Mol. Sci. 2018, 19, 2894. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, H.; Qu, C.; Tang, Y.; Shen, J.; Li, W.; Yue, S.; Kai, J.; Shang, G.; Zhu, Z.; et al. Urine and plasma metabonomics coupled with UHPLC-QTOF/MS and multivariate data analysis on potential biomarkers in anemia and hematinic effects of herb pair Gui-Hong. J. Ethnopharmacol. 2015, 170, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Razali, M.; Zainal, Z.A.; Maulidiani, M.; Shaari, K.; Zamri, Z.; Mohd, I.M.; Khatib, A.; Abas, F.; Ling, Y.S.; Rui, L.L.; et al. Classification of Raw Stingless Bee Honeys by Bee Species Origins Using the NMR- and LC-MS-Based Metabolomics Approach. Molecules 2018, 23, 2160. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Shi, B.; Koo, I.; Yin, X.; Lorkiewicz, P.; Suhail, H.; Rattan, R.; Giri, S.; McClain, C.J.; Zhang, X. Analysis of stable isotope assisted metabolomics data acquired by GC-MS. Anal. Chim. Acta 2017, 980, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Murugesu, S.; Ibrahim, Z.; Ahmed, Q.U.; Nik, Y.N.; Uzir, B.F.; Perumal, V.; Abas, F.; Saari, K.; El-Seedi, H.; Khatib, A. Characterization of alpha-Glucosidase Inhibitors from Clinacanthus nutans Lindau Leaves by Gas Chromatography-Mass Spectrometry-Based Metabolomics and Molecular Docking Simulation. Molecules 2018, 23, 2402. [Google Scholar] [CrossRef] [PubMed]
- Kuehnbaum, N.L.; Britz-McKibbin, P. New advances in separation science for metabolomics: Resolving chemical diversity in a post-genomic era. Chem. Rev. 2013, 113, 2437–2468. [Google Scholar] [CrossRef]
- Lüttjohann, A.; Fabene, P.F.; van Luijtelaar, G. A revised Racine’s scale for PTZ-induced seizures in rats. Physiol. Behav. 2009, 98, 579–586. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, R.; Miller, J.W. What is an epileptic seizure? Unifying definitions in clinical practice and animal research to develop novel treatments. Epilepsy Curr. 2010, 10, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, T.; Ali, A.M.; Al, W.M.; Pickard, B.; Watson, D.G. Metabolomic Profiling of Post-Mortem Brain Reveals Changes in Amino Acid and Glucose Metabolism in Mental Illness Compared with Controls. Comput. Struct. Biotechnol. J. 2016, 14, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triba, M.N.; Le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS models in metabolomics: The impact of permutation of dataset rows on the K-fold cross-validation quality parameters. Mol. Biosyst. 2015, 11, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Huang, Y.; Sun, J.; Wen, Y.; Qin, F.; Zhao, L.; Xiong, Z. A HILIC-UHPLC–MS/MS untargeted urinary metabonomics combined with quantitative analysis of five polar biomarkers on osteoporosis rats after oral administration of Gushudan. J. Chromatogr. B 2018, 1072, 40–49. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Flynn, C.J.; Wecker, L. Concomitant increases in the levels of choline and free fatty acids in rat brain: Evidence supporting the seizure-induced hydrolysis of phosphatidylcholine. J. Neurochem. 1987, 48, 1178–1184. [Google Scholar] [CrossRef]
- Sloviter, R.S.; Dempster, D.W. “Epileptic” brain damage is replicated qualitatively in the rat hippocampus by central injection of glutamate or aspartate but not by GABA or acetylcholine. Brain Res. Bull. 1985, 15, 39–60. [Google Scholar] [CrossRef]
- Gale, K. GABA and epilepsy: Basic concepts from preclinical research. Epilepsia 1992, 33 (Suppl. 5), S3–S12. [Google Scholar]
- Carmody, S.; Brennan, L. Effects of pentylenetetrazole-induced seizures on metabolomic profiles of rat brain. Neurochem. Int. 2010, 56, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Rankin-Gee, E.K.; McRae, P.A.; Baranov, E.; Rogers, S.; Wandrey, L.; Porter, B.E. Perineuronal net degradation in epilepsy. Epilepsia 2015, 56, 1124–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Amber samples are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | HMDB ID | Metabolites | Formula | m/z | ESI Mode | p Value | Ratio Control/Model |
|---|---|---|---|---|---|---|---|
| 1 | HMDB0010404 | Lysopc(22:6(4z,7z,10z,13z,16z,19z)) | C30H50NO7P | 568.3398 | + | <0.001 | 4.29 |
| 2 | HMDB0001406 | Niacinamide | C6H6N2O | 123.0553 | + | <0.001 | 2.64 |
| 3 | HMDB0002815 | Lysopc(18:1(9z)) | C26H52NO7P | 522.3554 | + | <0.001 | 2.95 |
| 4 | HMDB0000895 | Acetylcholine | C7H16NO2 | 146.1181 | + | <0.001 | 2.49 |
| 5 | HMDB0000157 | Hypoxanthine | C5H4N4O | 137.0458 | + | <0.001 | 2.56 |
| 6 | HMDB0062176 | N-lactoyl-leucine | C9H17NO4 | 204.1230 | + | <0.001 | 2.52 |
| 7 | HMDB0000201 | Acetylcarnitine | C9H17NO4 | 204.1230 | + | <0.001 | 2.26 |
| 8 | HMDB0010396 | Lysopc(20:4(8z,11z,14z,17z)) | C28H50NO7P | 544.3398 | + | <0.001 | 3.03 |
| 9 | HMDB0144632 | diethylamino-Acetaldehyde | C6H13NO | 116.1070 | + | <0.001 | 2.47 |
| 10 | HMDB0010382 | Lysopc(16:0) | C24H50NO7P | 496.3398 | + | <0.001 | 3.32 |
| 11 | HMDB0031779 | Isoprothiolane | C12H18O4S2 | 291.0719 | + | <0.001 | 2.28 |
| 12 | HMDB0000195 | Inosine | C10H12N4O5 | 269.0880 | + | <0.002 | 2.33 |
| 13 | HMDB0000562 | Creatinine | C4H7N3O | 114.0662 | + | <0.003 | 2.51 |
| 14 | HMDB0011496 | Lysope(0:0/22:6(4z,7z,10z,13z,16z,19z)) | C27H44NO7P | 526.2928 | + | <0.004 | 3.12 |
| 15 | HMDB0000062 | l-carnitine | C7H15NO3 | 162.1125 | + | 0.0015 | 3.17 |
| 16 | HMDB0010720 | butenoic acid | C4H6O2 | 87.04406 | + | 0.0017 | 3.06 |
| 17 | HMDB0000112 | Gamma-aminobutyric acid(GABA) | C4H9NO2 | 104.0706 | + | 0.0018 | 3.18 |
| 18 | HMDB0010384 | Lysopc(18:0) | C26H54NO7P | 524.3711 | + | 0.0018 | 3.05 |
| 19 | HMDB0038039 | Isovaleric acid amine | C5H13NO2 | 120.1019 | + | 0.0025 | 2.16 |
| 20 | HMDB0060348 | 2-Maleylacetate | C6H6O5 | 159.0288 | + | 0.0028 | 2.58 |
| 21 | HMDB0000064 | Creatine | C4H9N3O2 | 132.0768 | + | 0.0029 | 2.31 |
| 22 | HMDB0005065 | Oleoyl carnitine | C25H47NO4 | 426.3578 | + | 0.0041 | 1.59 |
| 23 | HMDB0001565 | Phosphorylcholine | C5H15NO4P | 184.0739 | + | 0.0105 | 2.96 |
| 24 | HMDB0010382 | Lysopc(16:0) | C24H50NO7P | 496.3398 | + | 0.0110 | 8.87 |
| 25 | HMDB0000097 | Choline | C5H14NO | 104.1075 | + | 0.0211 | 2.45 |
| 26 | HMDB0008067 | Pc(18:1(11z)/16:0) | C42H82NO8P | 760.5851 | + | 0.0228 | 4.07 |
| 27 | HMDB0008003 | Pc(16:1(9z)/18:0) | C42H82NO8P | 760.5851 | + | 0.0329 | 2.56 |
| 28 | HMDB0007911 | Pc(14:1(9z)/20:0) | C42H82NO8P | 760.5851 | + | 0.0405 | 4.29 |
| 29 | HMDB0007879 | Pc(14:0/20:1(11z)) | C42H82NO8P | 760.5851 | + | 0.0465 | 3.05 |
| 30 | HMDB0001406 | Niacinamide | C6H6N2O | 123.0553 | + | <0.001 | 4.29 |
| 31 | HMDB0001311 | d-lactic acid | C3H6O3 | 89.0244 | − | 0.0981 | 3.86 |
| 32 | HMDB0000805 | Pyrrolidonecarboxylic acid | C5H7NO3 | 128.0353 | − | 0.0529 | 2.43 |
| 33 | HMDB0035291 | Isoplumbagin | C11H8O3 | 187.0401 | − | 0.0103 | 3.62 |
| 34 | HMDB0014118 | Trifluoroacetic acid | C2HF3O2 | 112.9856 | − | 0.0327 | 1.82 |
| 35 | HMDB0000700 | Hydroxypropionic acid | C3H6O3 | 89.0244 | − | 0.0368 | 0.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Chen, C.; Zhu, Y.; Shang, E.; Zhao, M.; Guo, S.; Guo, J.; Qian, D.; Tang, Z.; Yan, H.; et al. Exploratory Cortex Metabolic Profiling Revealed the Sedative Effect of Amber in Pentylenetetrazole-Induced Epilepsy-Like Mice. Molecules 2019, 24, 460. https://doi.org/10.3390/molecules24030460
Zhu Z, Chen C, Zhu Y, Shang E, Zhao M, Guo S, Guo J, Qian D, Tang Z, Yan H, et al. Exploratory Cortex Metabolic Profiling Revealed the Sedative Effect of Amber in Pentylenetetrazole-Induced Epilepsy-Like Mice. Molecules. 2019; 24(3):460. https://doi.org/10.3390/molecules24030460
Chicago/Turabian StyleZhu, Zhenhua, Chenkai Chen, Yue Zhu, Erxin Shang, Ming Zhao, Sheng Guo, Jianming Guo, Dawei Qian, Zhishu Tang, Hui Yan, and et al. 2019. "Exploratory Cortex Metabolic Profiling Revealed the Sedative Effect of Amber in Pentylenetetrazole-Induced Epilepsy-Like Mice" Molecules 24, no. 3: 460. https://doi.org/10.3390/molecules24030460