
Generation of High Affinity Anti-Peptide Polyclonal Antibodies Recognizing Goat αs1-Casein

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
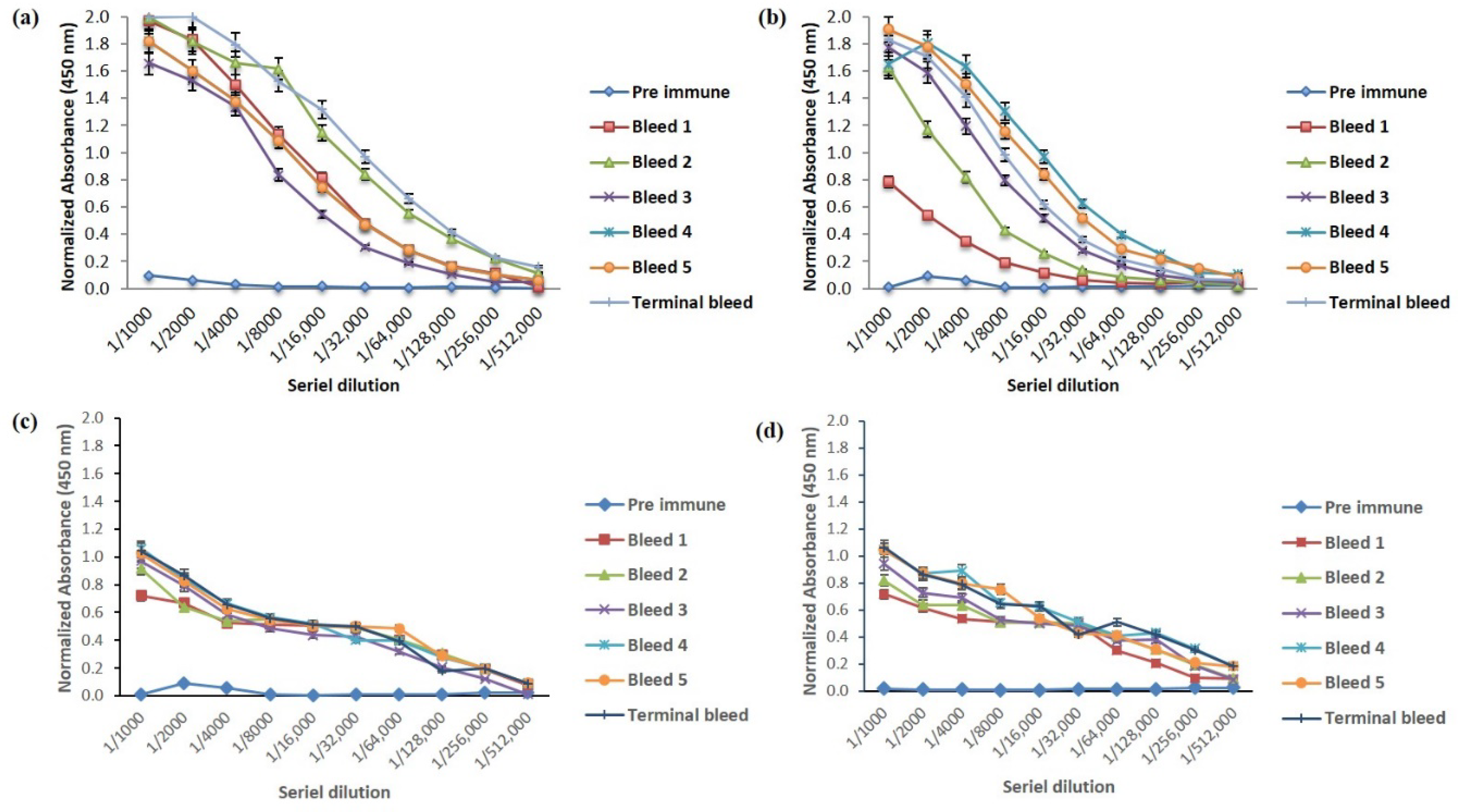
2.1. Humoral Response
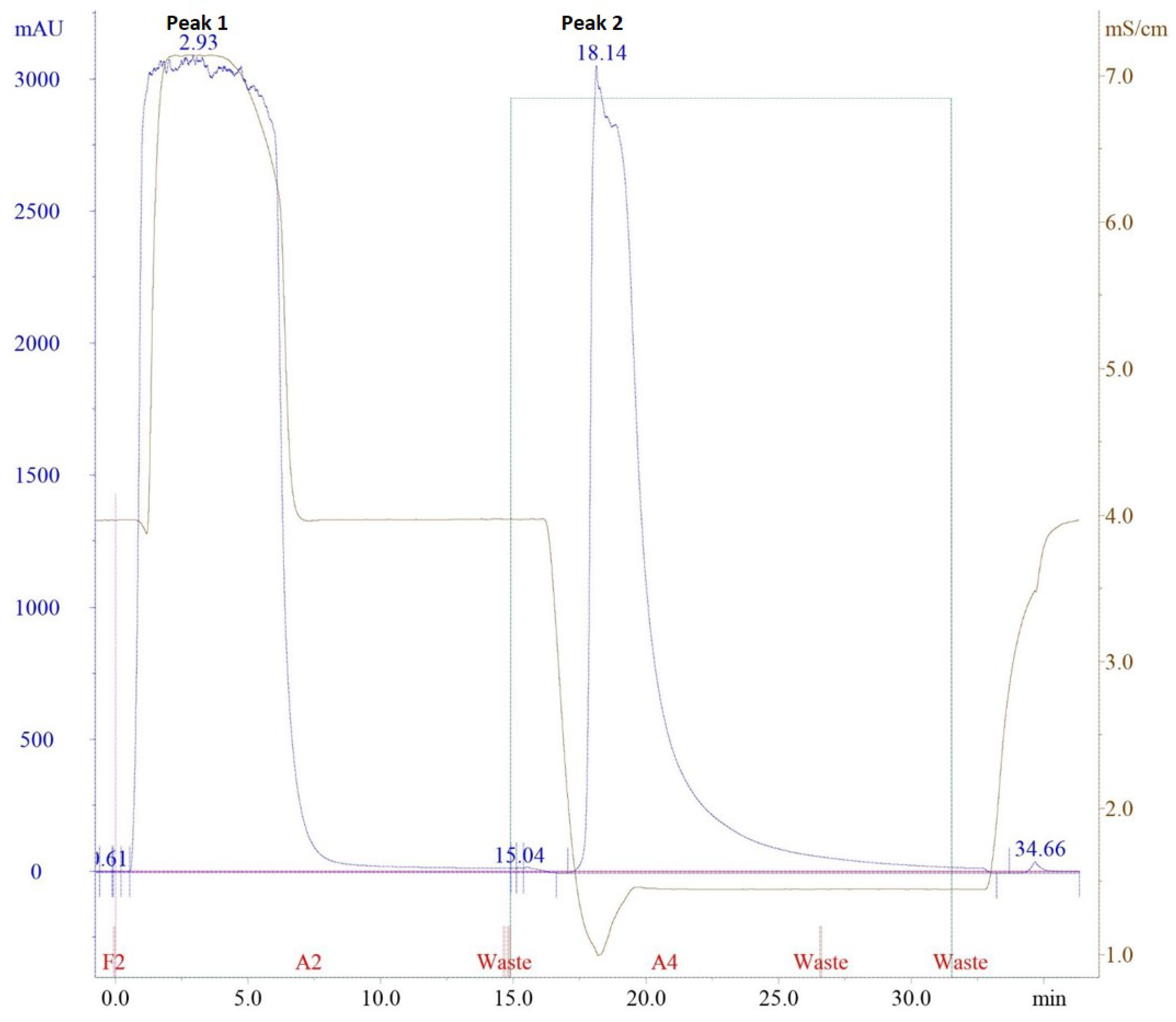
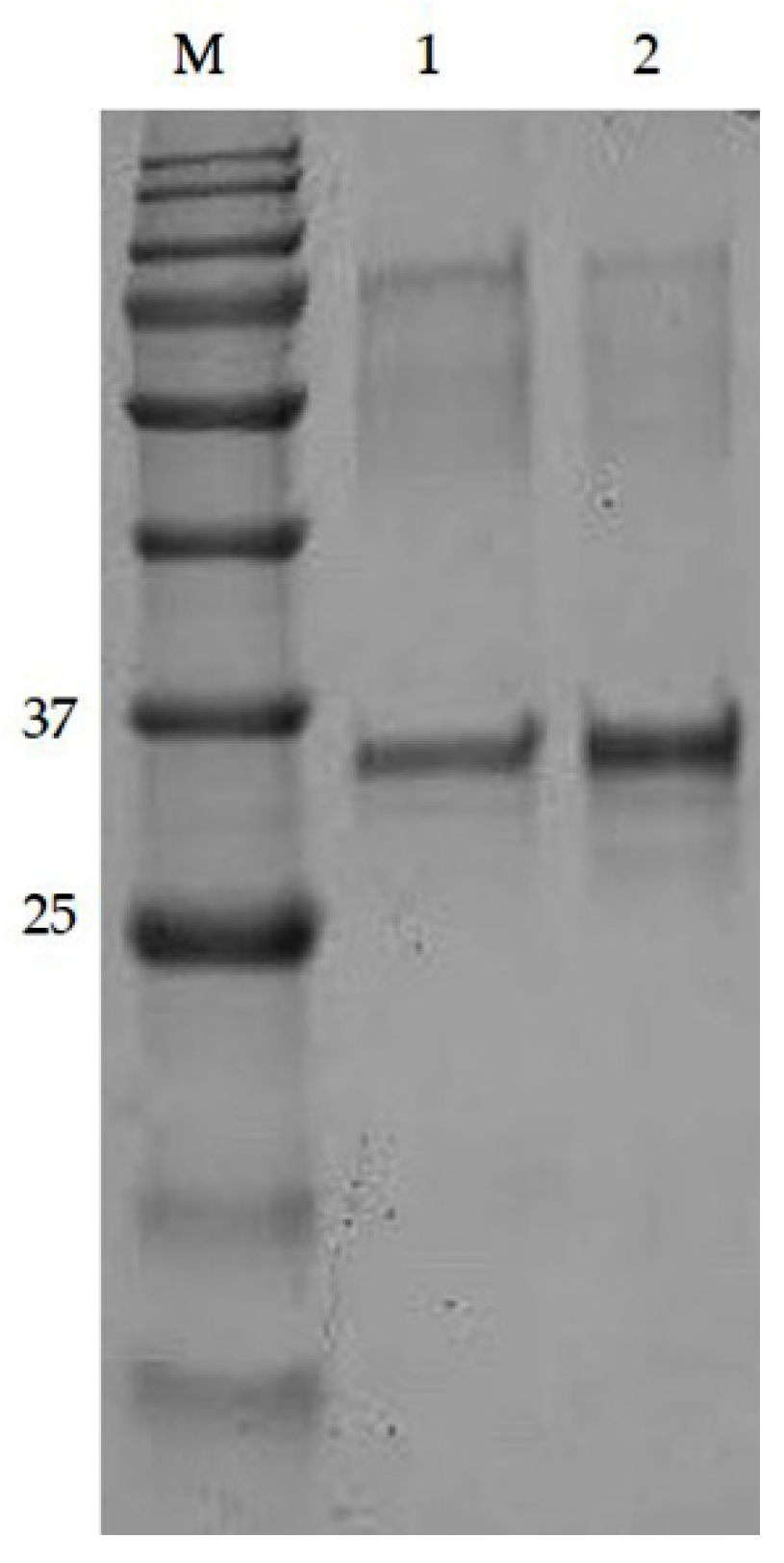
2.2. Purification of Polyclonal Antibody (pAb) Using Affinity Chromatography
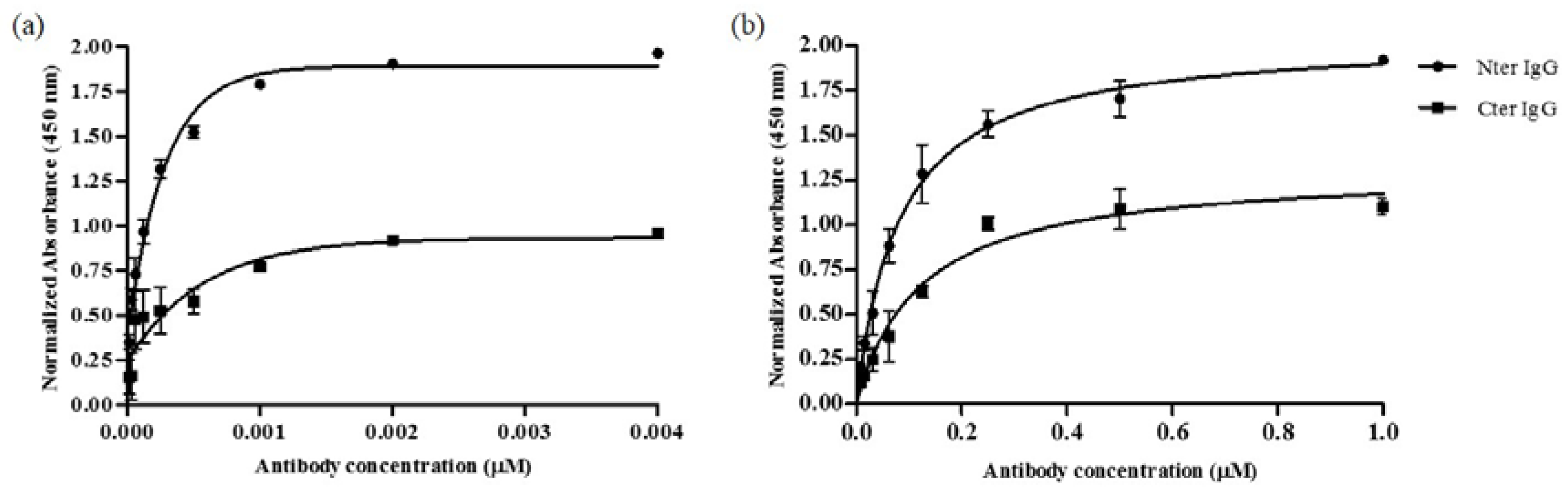
2.3. Binding of Purified IgGs Towards Peptides-BSA and Goat αs1-Casein Using Indirect ELISA
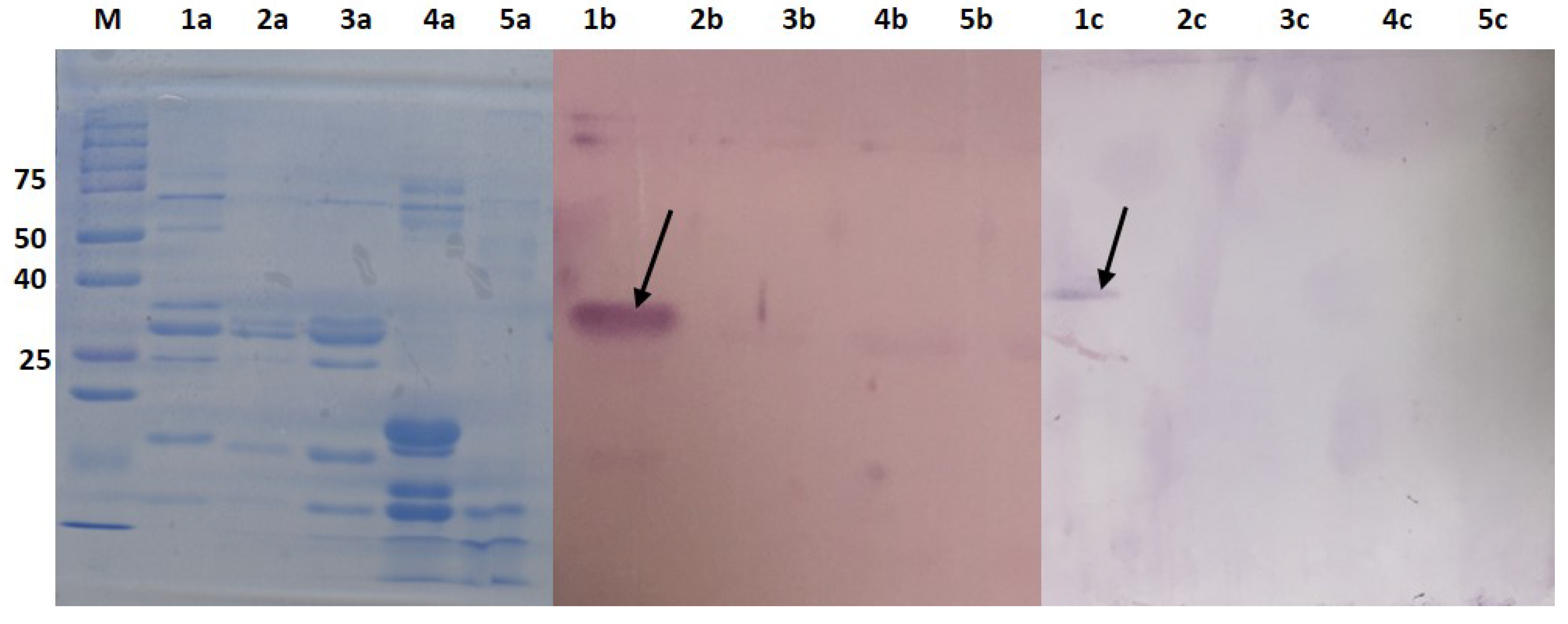
2.4. Cross Reactivity
3. Materials and Methods
3.1. Materials
3.2. Synthesis of Peptide
3.3. Immunization of Animals
3.4. Humoral Response ELISA
3.5. Affinity Chromatography
3.6. Milk Samples Preparation for Western Blot
3.7. SDS-PAGE and Western Blot Analysis
3.8. Purification of αs1-Casein from Goat’s Milk
3.9. Indirect ELISA
3.10. Data Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Najafi, M.; Rahimi Mianji, G.; Ansari Pirsaraie, Z. Cloning and comparative analysis of gene structure in promoter site of alpha-s1 casein gene in Naeinian goat and sheep. Meta Gene 2014, 2, 854–861. [Google Scholar] [CrossRef]
- Mangia, N.P.; Saliba, L.; Zoumpopoulou, G.; Chessa, S.; Anastasiou, R.; Karayiannis, I.; Sgouras, D.; Tsakalidou, E. Fermentation goat milk with different alpha-s1 casein genotype (csn1s1) fermented by selected lactobacillus paracasei as potential functional food. Fermentation 2019, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Cebo, C.; Lopez, C.; Henry, C.; Beauvallet, C.; Ménard, O.; Bevilacqua, C.; Bouvier, F.; Caillat, H.; Martin, P. Goat αs1-casein genotype affects milk fat globule physicochemical properties and the composition of the milk fat globule membrane. J. Dairy Sci. 2012, 95, 6215–6229. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Flores, F.; Montaldo, H.H.; Torres-Vázquez, J.A.; Alonso-Morales, R.A.; Gayosso-Vázquez, A.; Valencia-Posadas, M.; Castillo-Juárez, H. Additive and dominance effects of the alpha s1-casein locus on milk yield and composition traits in dairy goats. J. Dairy Res. 2012, 79, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, A.J.; Mcdonald, N.A.; Kivits, L.J.; Hurford, D.R.; Fahey, S.; Prosser, C. Allergic responses induced by goat milk as1-casein in a murine model of gastrointestinal atopy. J. Dairy Sci. 2012, 95, 83–90. [Google Scholar] [CrossRef]
- Bevilacqua, C.; Martin, P.; Candalh, C.; Fauquant, J.; Piot, M.; Roucayrol, A.M.; Pilla, F.; Heyman, M. Goats’ milk of defective αs1-casein genotype decreases intestinal and systemic sensitization to β-lactoglobulin in guinea pigs. J. Dairy Res. 2001, 68, 217–227. [Google Scholar] [CrossRef]
- Sela-Culang, I.; Kunik, V.; Ofran, Y. The structural basis of antibody-antigen recognition. Front. Immunol. 2013, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rudra, J.S.; Mishra, S.; Chong, A.S.; Mitchell, R.A.; Nardin, E.H.; Nussenzweig, V.; Collier, J.H. Self-assembled peptide nanofibers raising durable antibody responses against a malaria epitope. Biomaterials 2012, 33, 6476–6484. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Malhotra, R.; Peczuh, M.W.; Rusling, J.F. Electrochemical immunosensors for antibodies to peanut allergen Ara h2 using gold nanoparticle-peptide films. Anal. Chem. 2010, 82, 5865–5871. [Google Scholar] [CrossRef] [Green Version]
- Vermeersch, P.; Geboes, K.; Mariën, G.; Hoffman, I.; Hiele, M.; Bossuyt, X. Diagnostic performance of IgG anti-deamidated gliadin peptide antibody assays is comparable to IgA anti-tTG in celiac disease. Clin. Chim. Acta 2010, 411, 931–935. [Google Scholar] [CrossRef]
- Rudra, J.S.; Tian, Y.F.; Jung, J.P.; Collier, J.H. A self-assembling peptide acting as an immune adjuvant. Proc. Natl. Acad. Sci. USA 2009, 107, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Muller-Renaud, S.; Dupont, D.; Dulieu, P. Development of a biosensor immunoassay for the quantification of αs1-casein in milk. J. Dairy Res. 2005, 72, 57–64. [Google Scholar] [CrossRef]
- Ayyar, B.V.; Arora, S.; Murphy, C.; O’Kennedy, R. Affinity chromatography as a tool for antibody purification. Methods 2012, 56, 116–129. [Google Scholar] [CrossRef]
- Coelho, L.C.B.B.; Andrea, F.S.S.; Thiago, H.N.; Maria, T.S.C.; Patricia, M.G.P. Protein Purification by Affinity Chromatography. In Protein Purification; Rizwan, A., Ed.; Intech: Shanghai, China, 2012; pp. 53–72. ISBN 978-953-307-831-1. [Google Scholar] [CrossRef] [Green Version]
- Denizli, A. Purification of antibodies by affinity chromatography. J. Biol. Chem. 2011, 39, 1–18. [Google Scholar] [CrossRef]
- Szurdoki, F.; Székács, A.; Le, H.M.; Hammock, B.D. Synthesis of haptens and protein conjugates for the development of immunoassays for the insect growth regulator fenoxycarb. J. Agric. Food Chem. 2002, 50, 29–40. [Google Scholar] [CrossRef]
- Majidi, J.; Abdolalizadeh, J.; Amirkhiz, M.B.; Majidi, S. Production and purification of polyclonal antibody against bovine immunoglobulins in rabbits. Afr. J. Biotechnol. 2007, 6, 1369–1372. [Google Scholar] [CrossRef]
- Senocq, D.; Dupont, D.; Rolet-Répécaud, O.; Faurie, F.; Levieux, D. Antipeptide antibodies recognizing plasmin sensitive sites in bovine β-casein sequence. J. Agric. Food Chem. 2001, 49, 1571–1577. [Google Scholar] [CrossRef]
- Hancock, D.C.; OReilly, N.J. Synthetic Peptides as Antigens for Antibody Production. In Immunochemical Protocols; Burns, R., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2005; Volume 295, pp. 13–25. [Google Scholar] [CrossRef]
- Lee, B.; Huang, J.; Jayathilaka, L.P.; Lee, J.; Gupta, S. Antibody Production with Synthetic Peptides. In Methods in Molecular Biology; Steven, D.S., Ed.; Springer Science and Business Media: New York, NY, USA, 2016; Volume 1474, pp. 25–47. ISBN 9781493963522. [Google Scholar]
- Trier, N.H.; Hansen, P.R.; Houen, G. Production and characterization of peptide antibodies. Methods 2012, 56, 136–144. [Google Scholar] [CrossRef]
- Mimotopes Antipeptide Antibodies: Immunological Methods and Protocols. Available online: http://www.mimotopes.com/files/editor_upload/File/PeptidesAndAntibodies/PT2011-1-Antipeptide-Antibodies.PDF (accessed on 10 December 2019).
- Kapila, R.; Kavadi, P.K.; Kapila, S. Comparative evaluation of allergic sensitization to milk proteins of cow, buffalo and goat. Small Rumin. Res. 2013, 112, 191–198. [Google Scholar] [CrossRef]
- Cooper, M.; Lummas, S. Antibodies for immunochemistry. In Immunohistochemistry and Immunocytochemistry: Essential Methods; Renshaw, S., Ed.; John Wiley & Sons: Cambridge, UK, 2017; pp. 1–24. ISBN 9781118717776. [Google Scholar] [CrossRef]
- Muller-Renaud, S.; Dupont, D.; Dulieu, P. Quantification of k-casein in milk by optical immunosensor. Food Agric. Immunol. 2003, 15, 265–277. [Google Scholar] [CrossRef]
- Antibody Purification Handbook; GE Healthcare: Uppsala, Sweden, 2002; ISBN 18-1037-46.
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Popovic, P.; Diane, D.; Bruce, S.R.; Michael, R.S. Immunoglobulin Titers and Immunoglobulin Subtypes. In Measuring Immunity; Michael, T.L., Angus, W.T., Eds.; Elsevier: London, UK, 2005; pp. 159–171. ISBN 012455900X. [Google Scholar]
- Clark, A.; Befus, D.; O’Hashi, P.; Hart, F.; Schunk, M.; Fletch, A.; Griffin, G. Guidelines on: Antibody Production; Canadian Council on Animal Care: Ottawa, ON, Canada, 2002; ISBN 0-919087-37-X. [Google Scholar]
- He, S.; Li, X.; Gao, J.; Tong, P.; Chen, H. Development of sandwich ELISA for testing bovine -β-lactoglobulin allergenic residues by specific polyclonal antibody against human IgE binding epitopes. Food Chem. 2017, 227, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Vincent, D.; Ezernieks, V.; Elkins, A.; Nguyen, N.; Moate, P.J.; Cocks, B.G.; Rochfort, S. Milk bottom-up proteomics: Method optimization. Front. Genet. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ruiz, J.Á.; Miralles, B.; Agüera, P.; Amigo, L. Quantitative determination of αs2- and αs1-casein in goat’s milk with different genotypes by capillary electrophoresis. J. Chromatogr. A 2004, 1054, 279–284. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds (Nter and Cter polyclonal IgGs) are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Amino Acid | Exposed/Buried | Relative Surface Accessibility | Helix Probability | Sheet Probability | Coil Probability | Epitope Probability |
|---|---|---|---|---|---|---|---|
| 1 | R | B | 0.184 | 0.176 | 0.004 | 0.82 | 0.424 |
| 2 | P | B | 0.234 | 0.058 | 0.017 | 0.925 | 0.464 |
| 3 | K | E | 0.515 | 0.018 | 0.019 | 0.964 | 0.508 |
| 4 | H | B | 0.151 | 0.018 | 0.019 | 0.964 | 0.540 |
| 5 | P | E | 0.315 | 0.058 | 0.017 | 0.925 | 0.594 |
| 6 | I | B | 0.16 | 0.053 | 0.043 | 0.903 | 0.621 |
| 7 | N | E | 0.473 | 0.053 | 0.043 | 0.903 | 0.619 |
| 8 | H | B | 0.262 | 0.058 | 0.017 | 0.925 | 0.609 |
| 9 | R | E | 0.458 | 0.113 | 0.043 | 0.844 | 0.620 |
| 10 | G | E | 0.604 | 0.113 | 0.087 | 0.8 | 0.627 |
| 11 | L | B | 0.271 | 0.113 | 0.087 | 0.8 | 0.643 |
| 12 | S | B | 0.302 | 0.053 | 0.043 | 0.903 | 0.650 |
| 13 | P | E | 0.571 | 0.053 | 0.043 | 0.903 | 0.654 |
| 14 | E | E | 0.571 | 0.053 | 0.043 | 0.903 | 0.627 |
| 15 | V | E | 0.469 | 0.184 | 0.043 | 0.773 | 0.623 |
| 16 | P | E | 0.476 | 0.184 | 0.043 | 0.773 | 0.632 |
| 17 | N | E | 0.517 | 0.184 | 0.043 | 0.773 | 0.634 |
| 18 | E | E | 0.517 | 0.191 | 0.086 | 0.723 | 0.639 |
| 185 | P | B | 0.205 | 0.018 | 0.047 | 0.935 | 0.572 |
| 186 | I | B | 0.275 | 0.018 | 0.088 | 0.893 | 0.576 |
| 187 | G | B | 0.195 | 0.018 | 0.088 | 0.893 | 0.594 |
| 188 | S | B | 0.278 | 0.018 | 0.088 | 0.893 | 0.594 |
| 189 | E | E | 0.622 | 0.053 | 0.043 | 0.903 | 0.596 |
| 190 | N | E | 0.525 | 0.053 | 0.043 | 0.903 | 0.596 |
| 191 | S | B | 0.298 | 0.053 | 0.043 | 0.903 | 0.605 |
| 192 | G | E | 0.599 | 0.053 | 0.043 | 0.903 | 0.604 |
| 193 | K | E | 0.517 | 0.052 | 0.084 | 0.864 | 0.611 |
| 194 | T | B | 0.239 | 0.02 | 0.205 | 0.775 | 0.603 |
| 195 | T | E | 0.383 | 0.02 | 0.205 | 0.775 | 0.588 |
| 196 | M | B | 0.247 | 0.018 | 0.141 | 0.84 | 0.514 |
| 197 | P | E | 0.376 | 0.018 | 0.088 | 0.893 | 0.439 |
| 198 | L | E | 0.581 | 0.018 | 0.088 | 0.893 | 0.366 |
| 199 | W | E | 0.497 | 0.003 | 0.003 | 0.994 | 0.303 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohsin, A.Z.; Sukor, R.; Selamat, J.; Meor Hussin, A.S.; Ismail, I.H.; Jambari, N.N.; Mustaffa-Kamal, F. Generation of High Affinity Anti-Peptide Polyclonal Antibodies Recognizing Goat αs1-Casein. Molecules 2020, 25, 2622. https://doi.org/10.3390/molecules25112622
Mohsin AZ, Sukor R, Selamat J, Meor Hussin AS, Ismail IH, Jambari NN, Mustaffa-Kamal F. Generation of High Affinity Anti-Peptide Polyclonal Antibodies Recognizing Goat αs1-Casein. Molecules. 2020; 25(11):2622. https://doi.org/10.3390/molecules25112622
Chicago/Turabian StyleMohsin, Aliah Zannierah, Rashidah Sukor, Jinap Selamat, Anis Shobirin Meor Hussin, Intan Hakimah Ismail, Nuzul Noorahya Jambari, and Farina Mustaffa-Kamal. 2020. "Generation of High Affinity Anti-Peptide Polyclonal Antibodies Recognizing Goat αs1-Casein" Molecules 25, no. 11: 2622. https://doi.org/10.3390/molecules25112622
APA StyleMohsin, A. Z., Sukor, R., Selamat, J., Meor Hussin, A. S., Ismail, I. H., Jambari, N. N., & Mustaffa-Kamal, F. (2020). Generation of High Affinity Anti-Peptide Polyclonal Antibodies Recognizing Goat αs1-Casein. Molecules, 25(11), 2622. https://doi.org/10.3390/molecules25112622