
Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L.

 , ,
, ,  ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Phenolic Contents and Antioxidant Activities of C. maritimum Extract
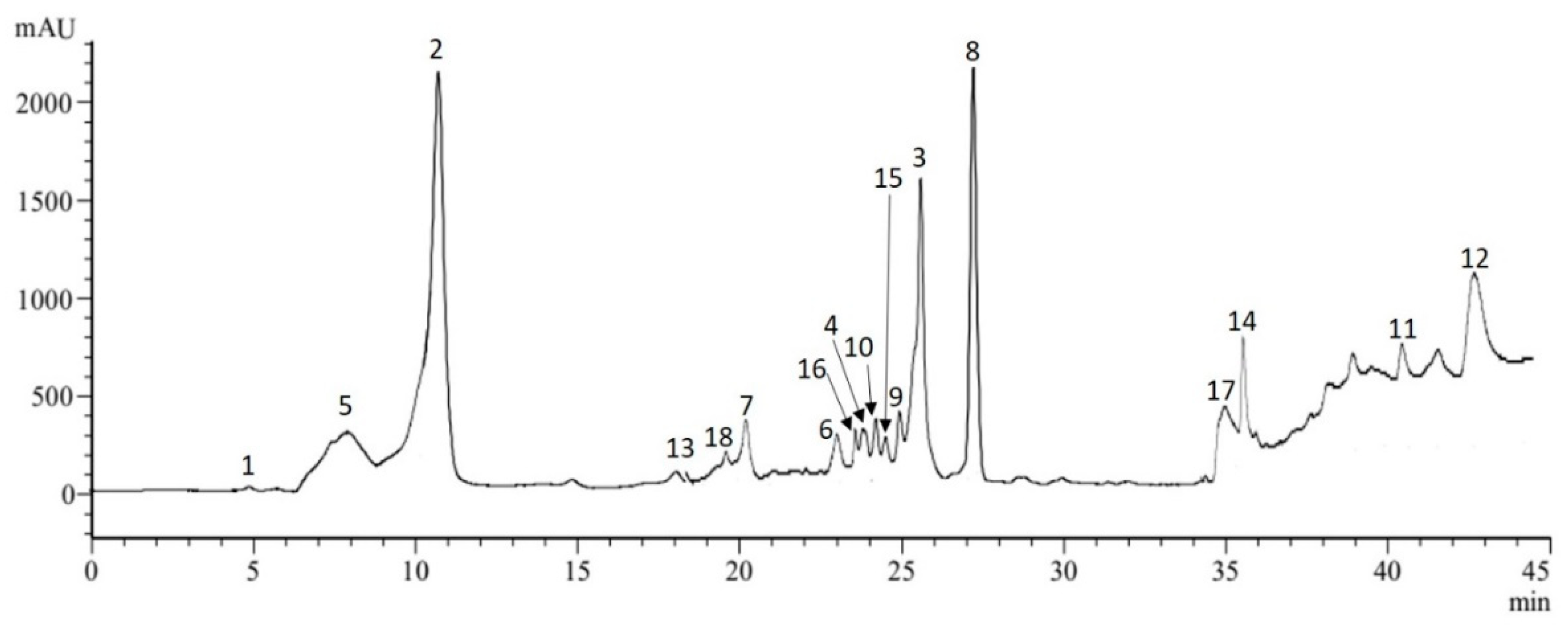
2.1.1. Phenolic Compounds and Phytochemical Profile
2.1.2. In Vitro Antioxidant Properties
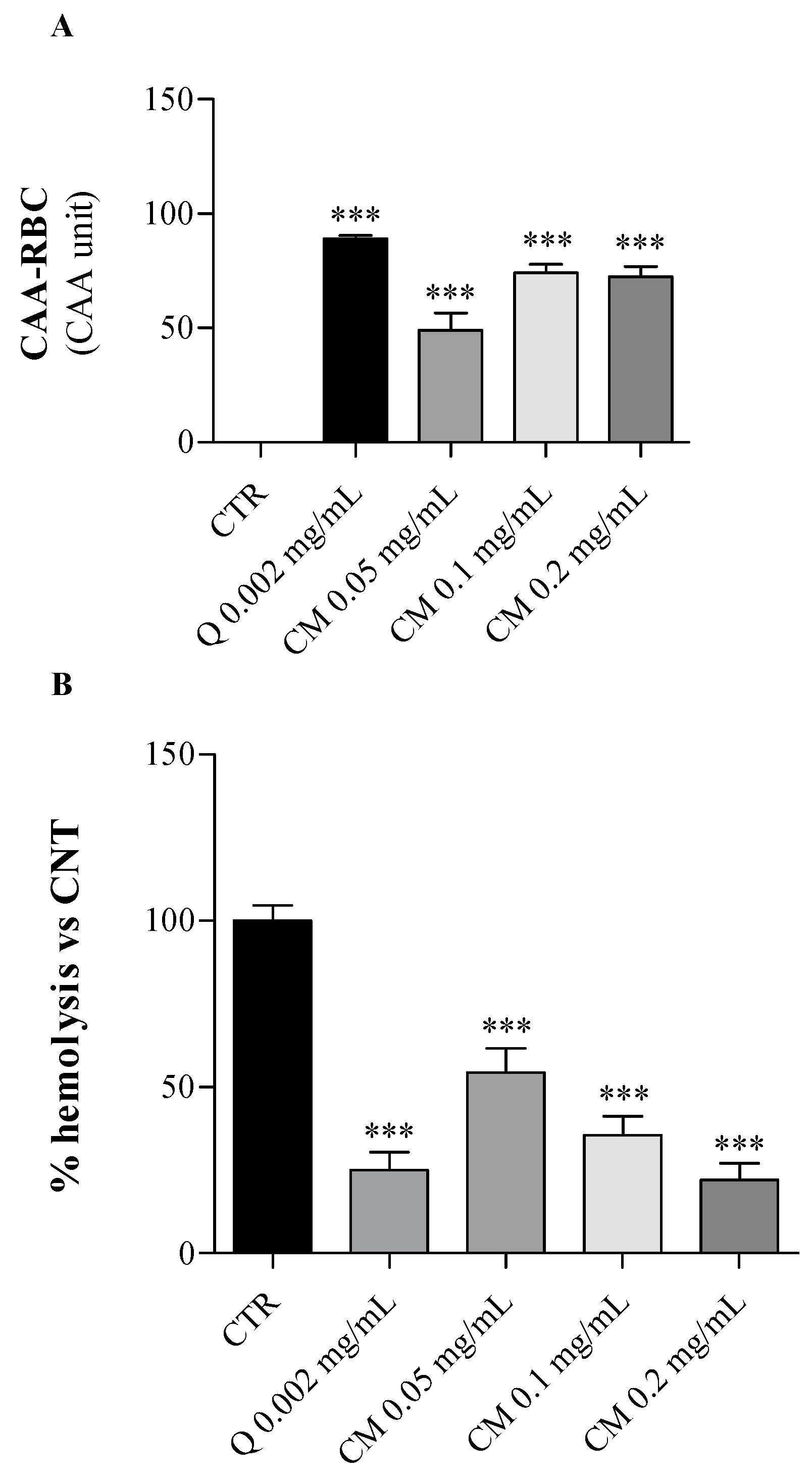
2.2. Biological Effects of C. maritimum on Human Erythrocytes
2.3. Antimicrobial Activity of C. maritimum L. Extract
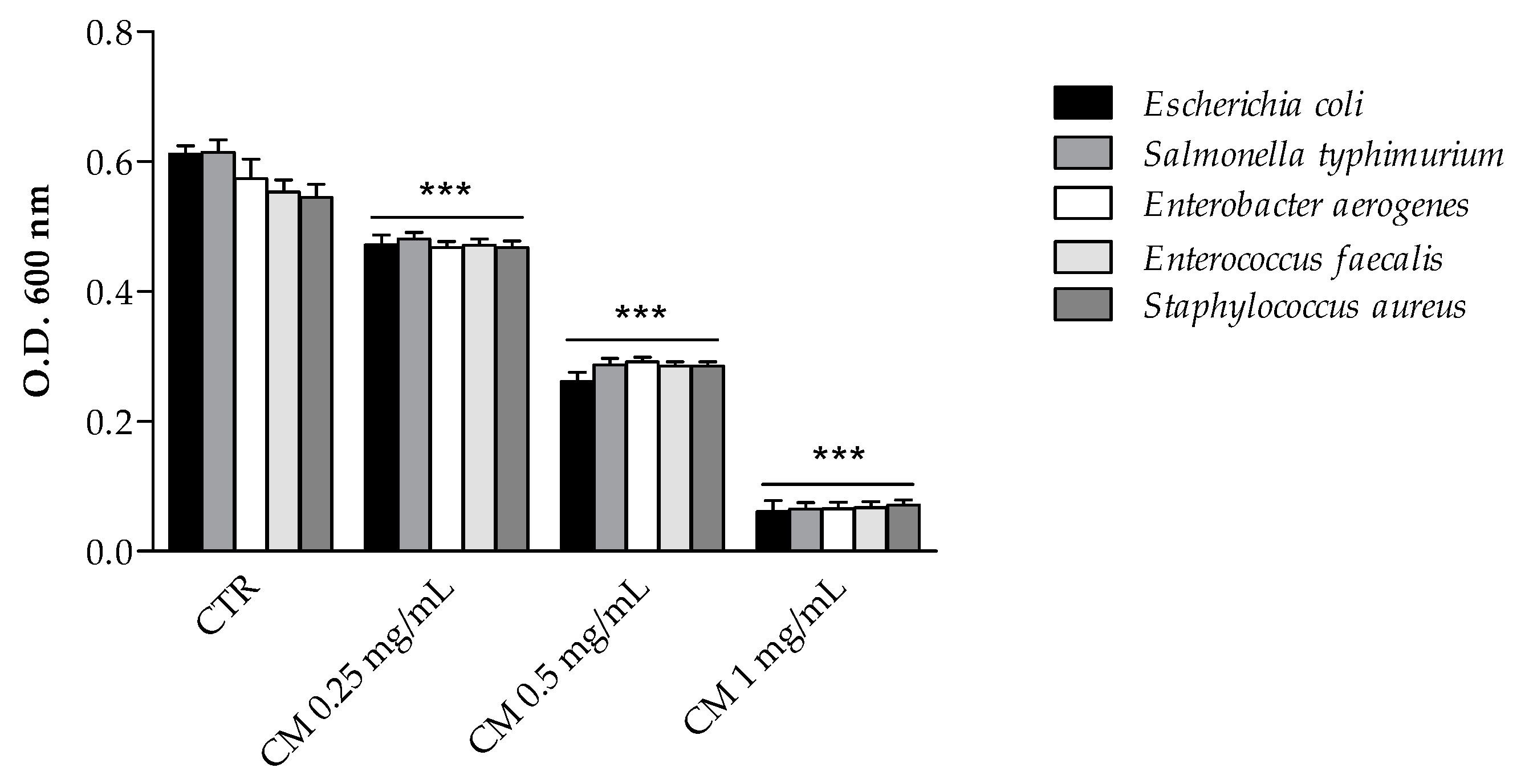
2.3.1. Inhibition of Bacterial Growth
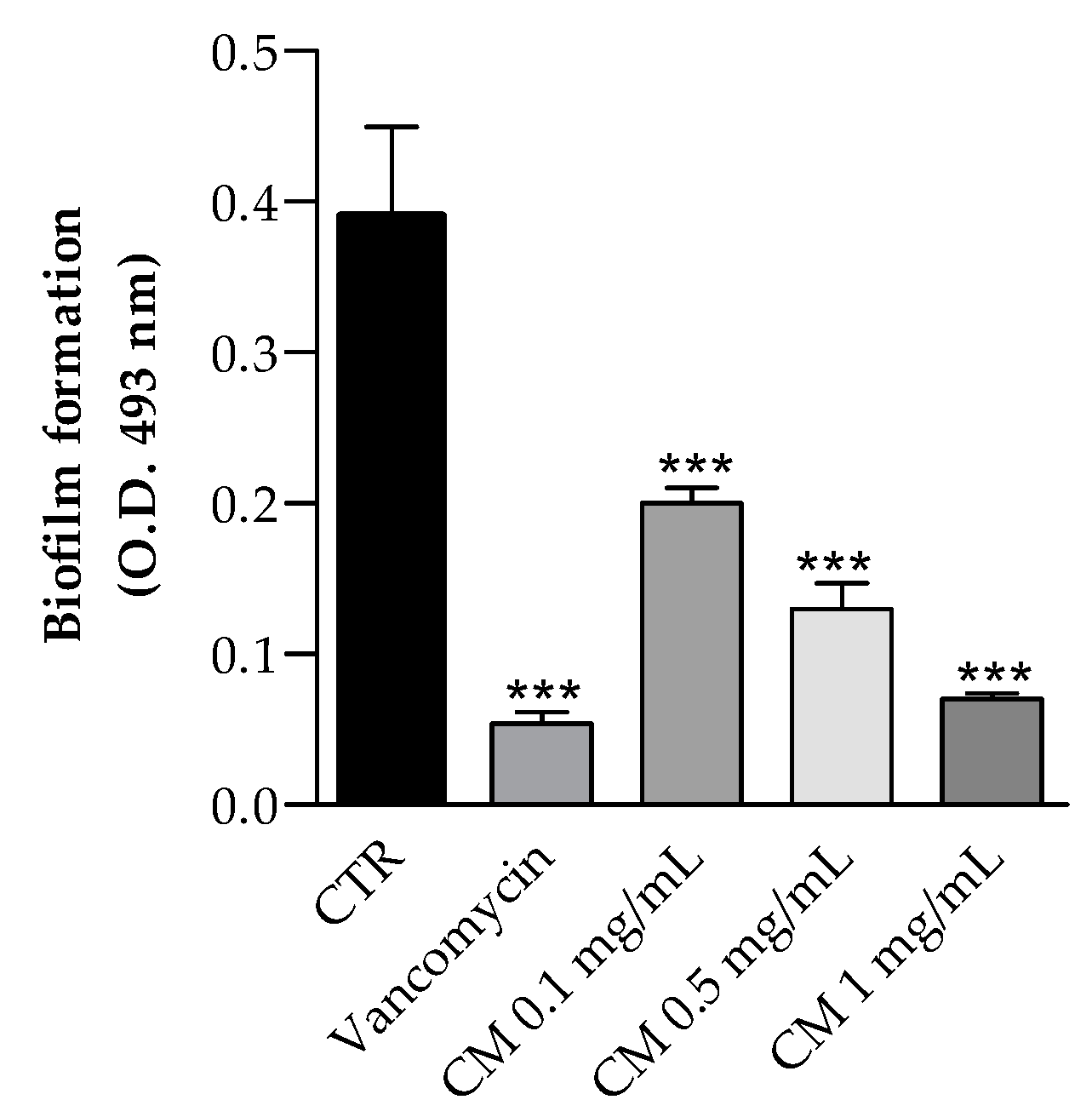
2.3.2. Biofilm Inhibition
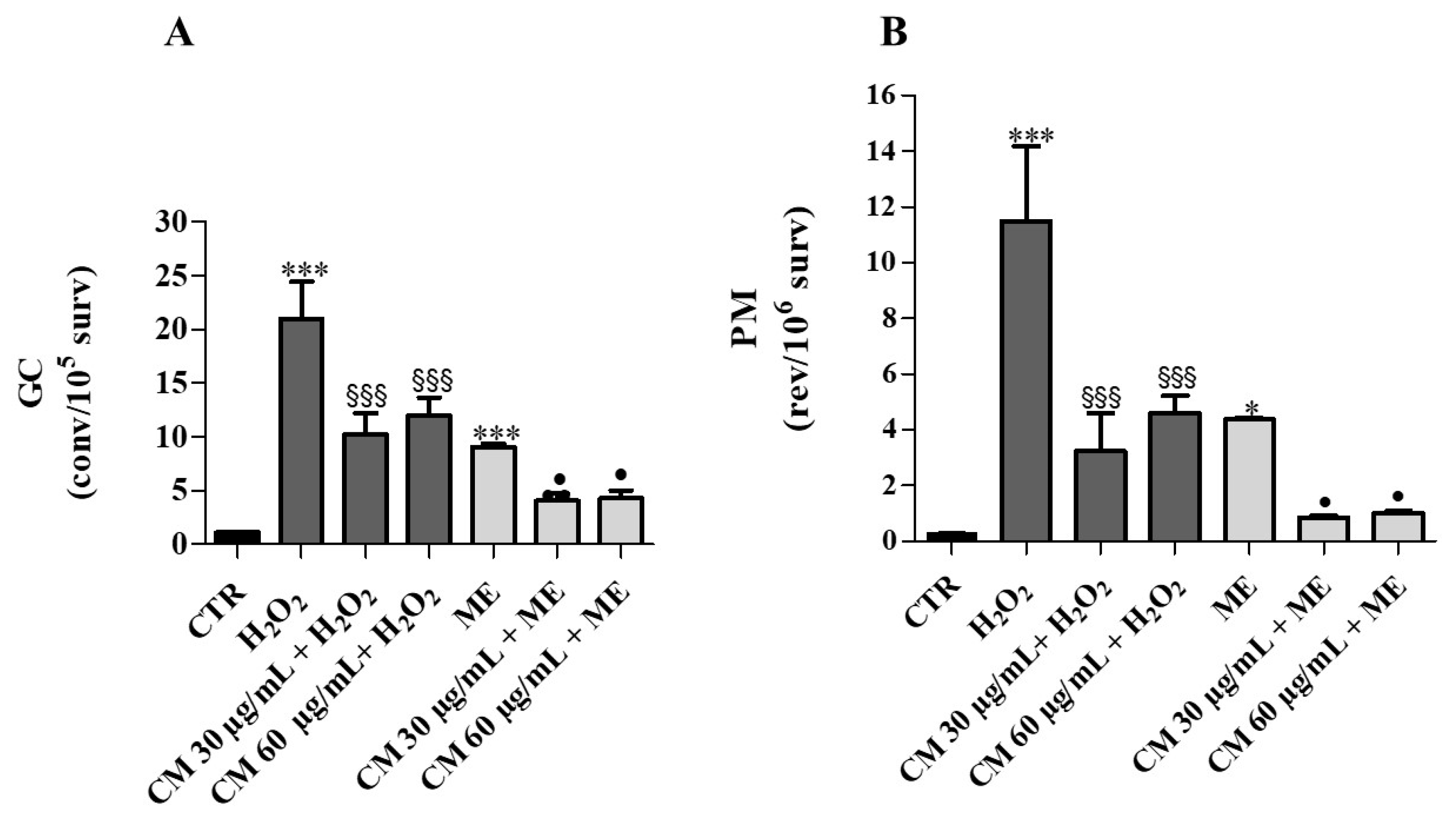
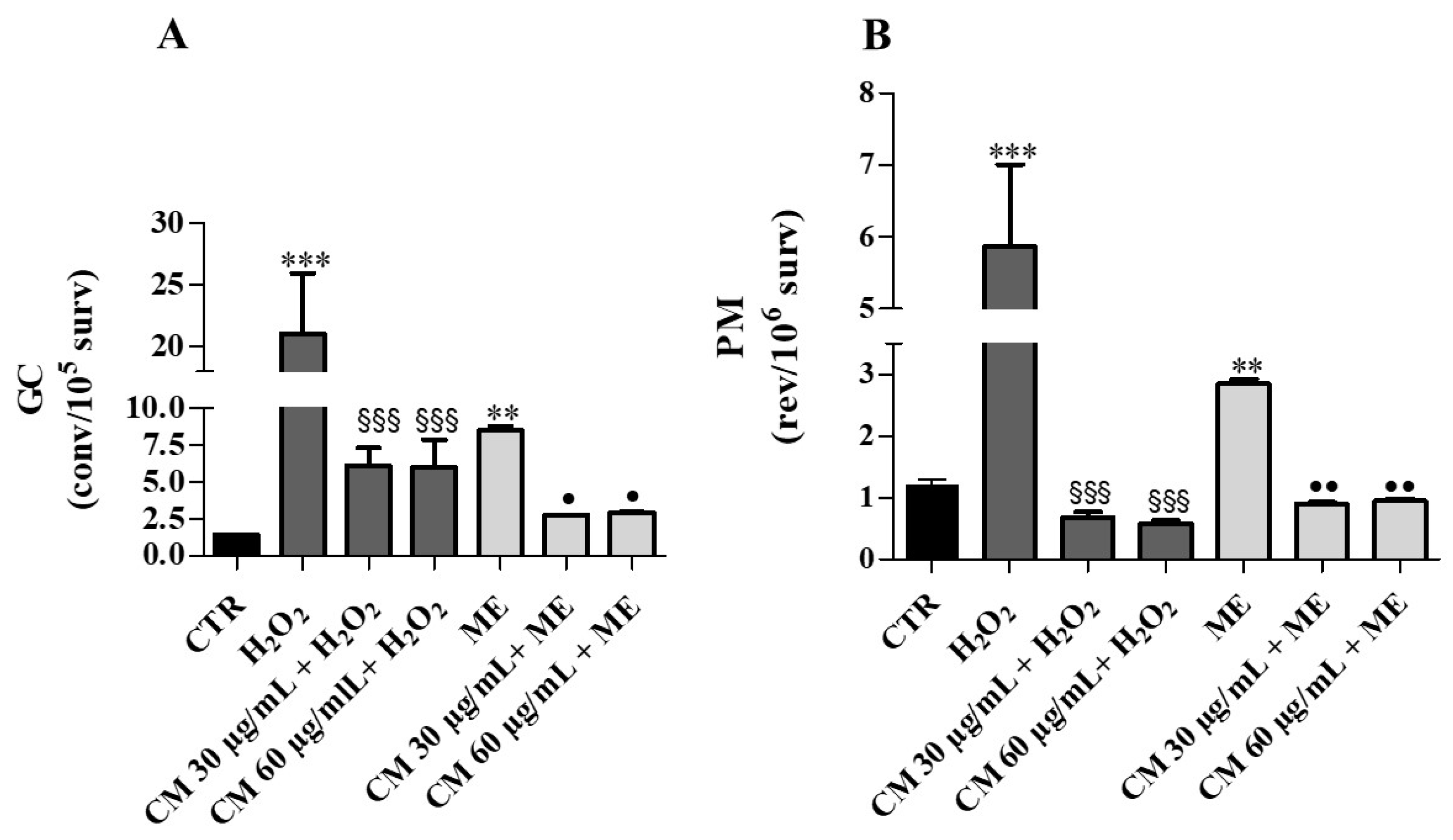
2.4. Antimutagenesis Assay in Yeast Cells
3. Materials and Methods
3.1. Plant Material
3.2. Hydro-Ethanolic Extraction
3.3. Phytochemical Characterization and Phenolic Compounds Profiling by LC-ESI-MS Analysis
3.3.1. Phenolic Contents of C. maritimum L. Extract
3.3.2. Phytochemical Profile of C. maritimum L. Extract
3.4. Antioxidant Activities (DPPH, ORAC and FRAP)
3.5. Preparation of Erythrocytes
3.5.1. Cellular Antioxidant Activity (CAA) in Human Erythrocytes
3.5.2. Oxidative Erythrocytes Hemolysis
3.6. Antimicrobial Activity
3.6.1. Bacterial Media
3.6.2. Growth Conditions of Pathogenic Bacteria
3.6.3. Antimicrobial Activity
3.6.4. Biofilm Production and Inhibition (Crystal Violet Assay)
3.7. Antimutagenesis Assay in Yeast Cells
- (1)
- Incubation assay: the C. maritimum extract (30 and 60 µg/mL) and oxidizing agents were incubated for 90 min, under shaking (30 °C) together with cells in logarithmic phase.
- (2)
- Growth assay: the oxidizing substances were added in the flasks where yeast cells were grown together with C. maritimum and incubated for 90 min under shaking (30 °C).
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Méot-Duros, L.; Magné, C. Antioxidant activity and phenol content of Crithmum maritimum L. leaves. Plant Physiol. Biochem. 2009, 47, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Jallali, I.; Zaouali, Y.; Missaoui, I.; Smaoui, A.; Abdelly, C.; Ksouri, R. Variability of antioxidant and antibacterial effects of essential oils and acetonic extracts of two edible halophytes: Crithmum maritimum L. and Inula crithmoïdes L. Food Chem. 2014, 145, 1031–1038. [Google Scholar] [CrossRef]
- Mekinić, I.G.; Šimat, V.; Ljubenkov, I.; Burčul, F.; Grga, M.; Mihajlovski, M.; Lončar, R.; Katalinić, V.; Skroza, D. Influence of the vegetation period on sea fennel, Crithmum maritimum L. (Apiaceae), phenolic composition, antioxidant and anticholinesterase activities. Ind. Crops Prod. 2018, 124, 947–953. [Google Scholar] [CrossRef]
- da Conceiçao Mendes Ferreira da Costa, M.; da Silva, A.G.; Sant’Anna da Silva, A.; de Menezes Lima, V.L.; Bezerra-Silva, P.C.; da Rocha, S.K.L.; do Amaral Ferraz Navarro, D.M.; Dos Santos Correia, M.T.; Napoleão, T.H.; da Silva, M.V.; et al. Essential oils from leaves of medicinal plants of Brazilian flora: Chemical composition and activity against Candida species. Medicines 2017, 4, 27. [Google Scholar]
- de Sousa, R.M.F.; Rosa, J.S.; Oliveira, L.; Cunha, A.; Fernandes-Ferreira, M. Activities of Apiaceae essential oils and volatile compounds on hatchability, development, reproduction and nutrition of Pseudaletia unipuncta (Lepidoptera: Noctuidae). Ind. Crops Prod. 2015, 63, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Souid, A.; Della Croce, C.M.; Pozzo, L.; Ciardi, M.; Giorgetti, L.; Gervasi, P.G.; Abdelly, C.; Magné, C.; Hamed, K.B.; Longo, V. Antioxidant properties and hepatoprotective effect of the edible halophyte Crithmum maritimum L. against carbon tetrachloride-induced liver injury in rats. Eur. Food Res. Technol. 2020, 246, 1393–1403. [Google Scholar] [CrossRef]
- Hamed, K.B.; Castagna, A.; Elkahoui, S.; Ranieri, A.; Abdelly, C. Sea fennel (Crithmum maritimum L.) under salinity conditions: A comparison of leaf and root antioxidant responses. Plant Growth Regul. 2007, 53, 185–194. [Google Scholar] [CrossRef]
- Pereira, C.G.; Barreira, L.; da Rosa Neng, N.; Nogueira, J.M.F.; Marques, C.; Santos, T.F.; Varela, J.; Custódio, L. Searching for new sources of innovative products for the food industry within halophyte aromatic plants: In vitro antioxidant activity and phenolic and mineral contents of infusions and decoctions of Crithmum maritimum L. Food Chem Toxicol. 2017, 107, 581–589. [Google Scholar] [CrossRef]
- Renna, M.; Gonnella, M.; Caretto, S.; Mita, G.; Serio, F. Sea fennel (Crithmum maritimum L.): From underutilized crop to new dried product for food use. Genet. Resour. Crop Evol. 2017, 64, 205–216. [Google Scholar] [CrossRef]
- Cornara, L.; La Rocca, A.; Marsili, S.; Mariotti, M.G. Traditional uses of plants in the eastern riviera (Liguria, Italy). J. Ethnopharmacol. 2009, 125, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, R.; Hayek, S.A.; Ibrahim, S.A. Plant extracts as antimicrobials. In Handbook of Natural Antimicrobials for Food Safety and Quality; Taylor, T.M., Ed.; Elsevier: Cambridge, UK, 2015; pp. 31–47. [Google Scholar]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Adámez, J.D.; Samino, E.G.; Sánchez, E.V.; González-Gómez, D. In vitro estimation of the antibacterial activity and antioxidant capacity of aqueous extracts from grape seeds (Vitis vinifera L.). Food Control 2012, 24, 136–141. [Google Scholar] [CrossRef]
- Frassinetti, S.; Della Croce, C.M.; Caltavuturo, L.; Longo, V. Antimutagenic and antioxidant activity of Lisosan G in Saccharomyces cerevisiae. Food Chem. 2012, 135, 2029–2034. [Google Scholar] [CrossRef]
- Bourgou, S.; Tammar, S.; Salem, N.; Mkadmini, K.; Msaada, K. Phenolic composition, essential oil, and antioxidant activity in the aerial part of Artemisia herba-alba from several provenances: A comparative study. Int. J. Food Prop. 2016, 19, 549–563. [Google Scholar] [CrossRef]
- Medini, F.; Ksouri, R.; Falleh, H.; Megdiche, W.; Trabelsi, N.; Abdelly, C. Effects of physiological stage and solvent on polyphenol composition, antioxidant and antimicrobial activities of Limonium densiflorum. J. Med. Plant Res. 2011, 5, 6719–6730. [Google Scholar]
- Jallali, I.; Megdiche, W.; M’Hamdi, B.; Oueslati, S.; Smaoui, A.; Abdelly, C.; Riadh, K. Changes in phenolic composition and antioxidant activities of the edible halophyte Crithmum maritimum L. with physiological stage and extraction method. Acta Physiol. Plant. 2012, 34, 1451–1459. [Google Scholar] [CrossRef]
- Nabet, N.; Boudries, H.; Chougui, N.; Loupassaki, S.; Souagui, S.; Burlo, F.; Hernandez, F.; Carbonell-Barrachina, A.A.; Madani, K.; Larbat, R. Biological activities and secondary compound composition from Crithmum maritimum aerial parts. Int. J. Food Prop. 2017, 20, 1843–1855. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Zheng, Q.; Sun, R.; Wu, J.; Li, X.; Liu, R. Recent progress in the study of Artemisiae Scopariae Herba (Yin Chen), a promising medicinal herb for liver diseases. Biomed. Pharmacother. 2020, 130, 110513. [Google Scholar] [CrossRef] [PubMed]
- Viola, H.; Wasowski, C.; Marder, M.; Wolfman, C.; Paladini, A.C.; Medina, J.H. Sedative and hypnotic properties of Salvia guaranitica St. Hil. and of its active principle, Cirsiliol. Phytomedicine 1997, 4, 47–52. [Google Scholar] [CrossRef]
- Yakoub, A.R.B.; Abdehedi, O.; Jridi, M.; Elfalleh, W.; Nasri, M.; Ferchichi, A. Flavonoids, phenols, antioxidant, and antimicrobial activities in various extracts from Tossa jute leave (Corchorus olitorus L.). Ind. Crops Prod. 2018, 118, 206–213. [Google Scholar] [CrossRef]
- Santana-Gàlvez, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Onakpoya, I.J.; Spencer, E.A.; Thompson, M.J.; Heneghan, C.J. The effect of chlorogenic acid on blood pressure: A systematic review and meta-analysis of randomized clinical trials. J. Hum. Hypertens. 2015, 29, 77–81. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Jallali, I.; Debez, A.; Magné, C.; Isoda, H.; Abdelly, C. Medicinal halophytes potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Caddeo, C.; Manca, M.L.; Peris, J.E.; Usach, I.; Diez-Sales, O.; Matos, M.; Manconi, M. Tocopherol-loaded transferosomes: In vitro antioxidant activity and efficacy in skin regeneration. Int. J. Pharm. 2018, 551, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Souid, A.; Bellani, L.; Gabriele, M.; Pucci, L.; Smaoui, A.; Abdelly, C.; Hamed, K.B.; Longo, V. Phytochemical and biological activities in Limonium species collected in different biotopes of Tunisia. Chem. Biodivers. 2019, 16, e1900216. [Google Scholar] [CrossRef]
- Adamczak, A.; Ozarowski, M.; Karpinski, T.M. Antibacterial activity of some flavonoids and organic acids widely distributed in plants. J. Clin. Med. 2020, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2014, 238, 589–596. [Google Scholar] [CrossRef]
- Luís, Â.; Silva, F.; Sousa, S.; Duarte, A.P.; Domingues, F. Antistaphylococcal and biofilm inhibitory activities of gallic, caffeic and chlorogenic acids. Biofouling 2014, 30, 69–79. [Google Scholar] [CrossRef]
- Fiamegos, Y.C.; Kastritis, P.L.; Exarchou, V.; Han, H.; Bonvin, A.M.J.J.; Vervoort, J.; Lewis, K.; Hamblin, M.R.; Tegos, G.P. Antimicrobial and efflux pump inhibitory activity of caffeoylquinic acids from Artemisia absinthium against Gram-positive pathogenic bacteria. PLoS ONE 2011, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houta, O.; Akrout, A.; Najja, H.; Neffati, M.; Amri, H. Chemical composition, antioxidant and antimicrobial activities of essential oil from Crithmum maritimum cultivated in Tunisia. J. Essent. Oil-Bear. Plants 2015, 18, 1459–1466. [Google Scholar] [CrossRef]
- Ruberto, G.; Baratta, M.T.; Deans, S.G.; Dorman, H.J.D. Antioxidant and antimicrobial activity of Foeniculum vulgare and Crithmum maritimum essential oils. Planta Med. 2000, 66, 687–693. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewsk, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Alves-Silva, J.M.; Guerra, I.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Figueirinha, A.; Salgueiro, L. Chemical composition of Crithmum maritimum L. essential oil and hydro-distillation residual water by GC-MS and HPLC-DAD-MS/MS, and their biological activities. Ind. Crops Prod. 2020, 149, 112329. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Ghasemian, A. An overview on anti-biofilm properties of quercetin against bacterial pathogens. World J. Microbiol. Biotechnol. 2019, 35, 143. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Khan, M.S.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug resistant Escherichia coli and Staphylococcus aureus. Food Control 2017, 79, 325–332. [Google Scholar] [CrossRef]
- Zimmermann, F.K.; Kern, R.; Rasenberg, H.A. Yeast strain for simultaneous detection of induced mitotic crossing-over, mitotic gene conversion and reverse mutation. Mutat. Res. 1975, 28, 381–388. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 2000, 299, 152–178. [Google Scholar]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef] [PubMed]
- Romani, A.; Mancini, P.; Tatti, S.; Vincieri, F. Polyphenols and polysaccharides in Tuscan grapes and wines. Ital. J. Food Sci. 1996, 8, 13–24. [Google Scholar]
- Sun, B.; da Silva, J.M.R.; Spranger, I. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Food Chem. 1998, 46, 4267–4274. [Google Scholar] [CrossRef]
- Sokmen, A.; Gulluce, M.; Akpultat, H.A.; Daferera, D.; Tepe, B.; Polissiou, M. The in vitro antimicrobial and antioxidant activities of the essential oils and methanol extracts of endemic Thymus spathulifolius. Food Control 2004, 15, 627–634. [Google Scholar] [CrossRef]
- Bacchiocca, M.; Biagiotti, E.; Ninfali, P. Nutritional and technological reasons for evaluating the antioxidant capacity of vegetable products. Ital. J. Food Sci. 2006, 18, 1–9. [Google Scholar]
- Gabriele, M.; Parri, E.; Felicioli, A.; Sagona, S.; Pozzo, L.; Biondi, C.; Domenici, V.; Pucci, L. Phytochemical composition and antioxidant activity of Tuscan bee pollen of different botanic origins. Ital. J. Food Sci. 2015, 27, 248–259. [Google Scholar]
- Rodrigues, M.J.; Soszynski, A.; Martins, A.; Rauter, A.P.; Neng, N.R.; Nogueira, J.M.F.; Varela, J.; Barreira, L.; Custodio, L. Unravelling the antioxidant potential and the phenolic composition of different anatomical organs of the marine halophyte Limonium algarvense. Ind. Crops Prod. 2015, 77, 315–322. [Google Scholar] [CrossRef]
- Di Ciccio, P.; Vergara, A.; Festino, A.R.; Paludi, D.; Zanardi, E.; Ghidini, S.; Ianieri, A. Biofilm formation by Staphylococcus aureus on food contact surfaces: Relationship with temperature and cell surface hydrophobicity. Food Control 2015, 50, 930–936. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and antioxidant activity of a selected lectin-free common bean (Phaseolus vulgaris L.) in two cell-based models. Plant Food Hum. Nutr. 2015, 70, 35–41. [Google Scholar] [CrossRef]
- Blando, F.; Russo, R.; Negro, C.; De Bellis, L.; Frassinetti, S. Antimicrobial and antibiofilm activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. cladode polyphenolic extracts. Antioxidants 2019, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, 2437. [Google Scholar] [CrossRef] [PubMed]
- Del Carratore, R.; Bronzetti, G.; Bauer, C.; Corsi, C.; Nieri, R.; Paolini, M.; Giagoni, P. Cytochrome P-450 factors determining synthesis in strain D7 Saccharomyces cerevisiae. An alternative system to microsomal assay. Mutat. Res. 1983, 121, 117–123. [Google Scholar] [CrossRef]
- Peláez-Soto, A.; Fernández-Espinar, M.T.; Roig, P.; Gil, J.V. Evaluation of the ability of polyphenol extracts of cocoa and red grape to promote the antioxidant response in yeast using a rapid multi-well assay. J. Food. Sci. 2017, 82, 324–332. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. maritimum Leaves Extract | |
|---|---|
| Total polyphenols (mg GAE g−1 DW) | 31.70 ± 0.09 |
| Total flavonoids (mg CE g−1 DW) | 25.61 ± 0.04 |
| Flavonols (mg QE g−1 DW) | 17.30 ± 0.06 |
| CTC (mg CE g−1 DW) | 0.97 ± 0.09 |
| DPPH (IC50, mg mL−1) | 0.22 ± 0.04 |
| ORAC (µmol TE g−1 DW) | 15,835 ± 71 |
| FRAP (EC50, mg mL−1) | 1.82 ± 0.02 |
| N° | Metabolic Class Subclass | RT (min) | Compound Identified | Content (mg g−1 DW) |
|---|---|---|---|---|
| 5 | Organic acid Phenolic acids Hydroxycinnamic acids | 8.2 | Quinic acid | 3.78 |
| 1 | 5.0 | Gallic acid | - | |
| 2 | 11.1 | Chlorogenic acid | 7.25 | |
| 3 | 25.8 | Neochlorogenic acid | 2.03 | |
| 4 | 23.9 | Cryptochlorogenic acid | 1.17 | |
| 6 | 23.0 | Caffeic acid | 0.67 | |
| 7 | 20.3 | p-Coumaric acid | 1.02 | |
| 8 | 27.7 | Trans ferulic acid | 1.41 | |
| 9 | 24.9 | 4,5-di-O-caffeoylquinic acid | 0.72 | |
| Flavonoids | ||||
| 10 | 24.4 | Naringin | 0.14 | |
| 11 | 40.5 | Naringenin | 0.09 | |
| 12 | 42.8 | Luteolin-7-O-glucoside | 0.35 | |
| 13 | 18.2 | Rutin | 1.75 | |
| 14 | 35.5 | Cirsiliol | 1.31 | |
| Flavonols | ||||
| 15 | 25.4 | Quercetin | 0.09 | |
| 16 | 23.7 | Kaempferol | 0.72 | |
| 17 | 35.0 | Hyperoside (quercetin-3-O-galactoside) | 1.12 | |
| 18 | 19.8 | Quercetrin (quercetin-3-O-rhamnoside) | 1.41 | |
| Total | 25.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souid, A.; Della Croce, C.M.; Frassinetti, S.; Gabriele, M.; Pozzo, L.; Ciardi, M.; Abdelly, C.; Hamed, K.B.; Magné, C.; Longo, V. Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L. Molecules 2021, 26, 5380. https://doi.org/10.3390/molecules26175380
Souid A, Della Croce CM, Frassinetti S, Gabriele M, Pozzo L, Ciardi M, Abdelly C, Hamed KB, Magné C, Longo V. Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L. Molecules. 2021; 26(17):5380. https://doi.org/10.3390/molecules26175380
Chicago/Turabian StyleSouid, Aymen, Clara Maria Della Croce, Stefania Frassinetti, Morena Gabriele, Luisa Pozzo, Marco Ciardi, Chedly Abdelly, Karim Ben Hamed, Christian Magné, and Vincenzo Longo. 2021. "Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L." Molecules 26, no. 17: 5380. https://doi.org/10.3390/molecules26175380
APA StyleSouid, A., Della Croce, C. M., Frassinetti, S., Gabriele, M., Pozzo, L., Ciardi, M., Abdelly, C., Hamed, K. B., Magné, C., & Longo, V. (2021). Nutraceutical Potential of Leaf Hydro-Ethanolic Extract of the Edible Halophyte Crithmum maritimum L. Molecules, 26(17), 5380. https://doi.org/10.3390/molecules26175380