
Phytochemical Analysis and Anti-Inflammatory and Anti-Osteoarthritic Bioactive Potential of Verbascum thapsus L. (Scrophulariaceae) Leaf Extract Evaluated in Two In Vitro Models of Inflammation and Osteoarthritis

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Phenylethanoid Glycosides Profile and Total Phenolic Content
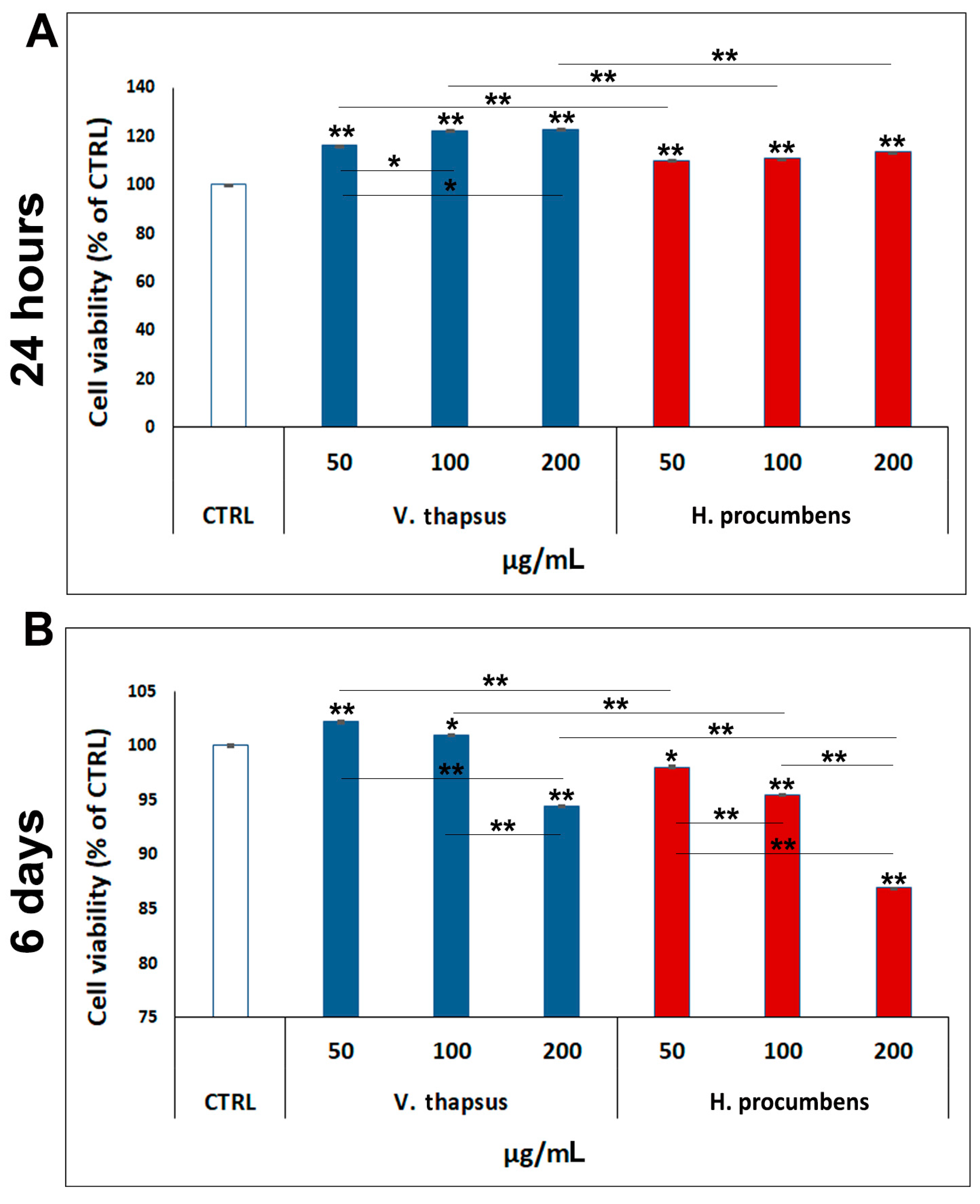
2.2. Evaluation of the Cytotoxic Effect of V. thapsus Extract on RAW 264.7 Cell Line
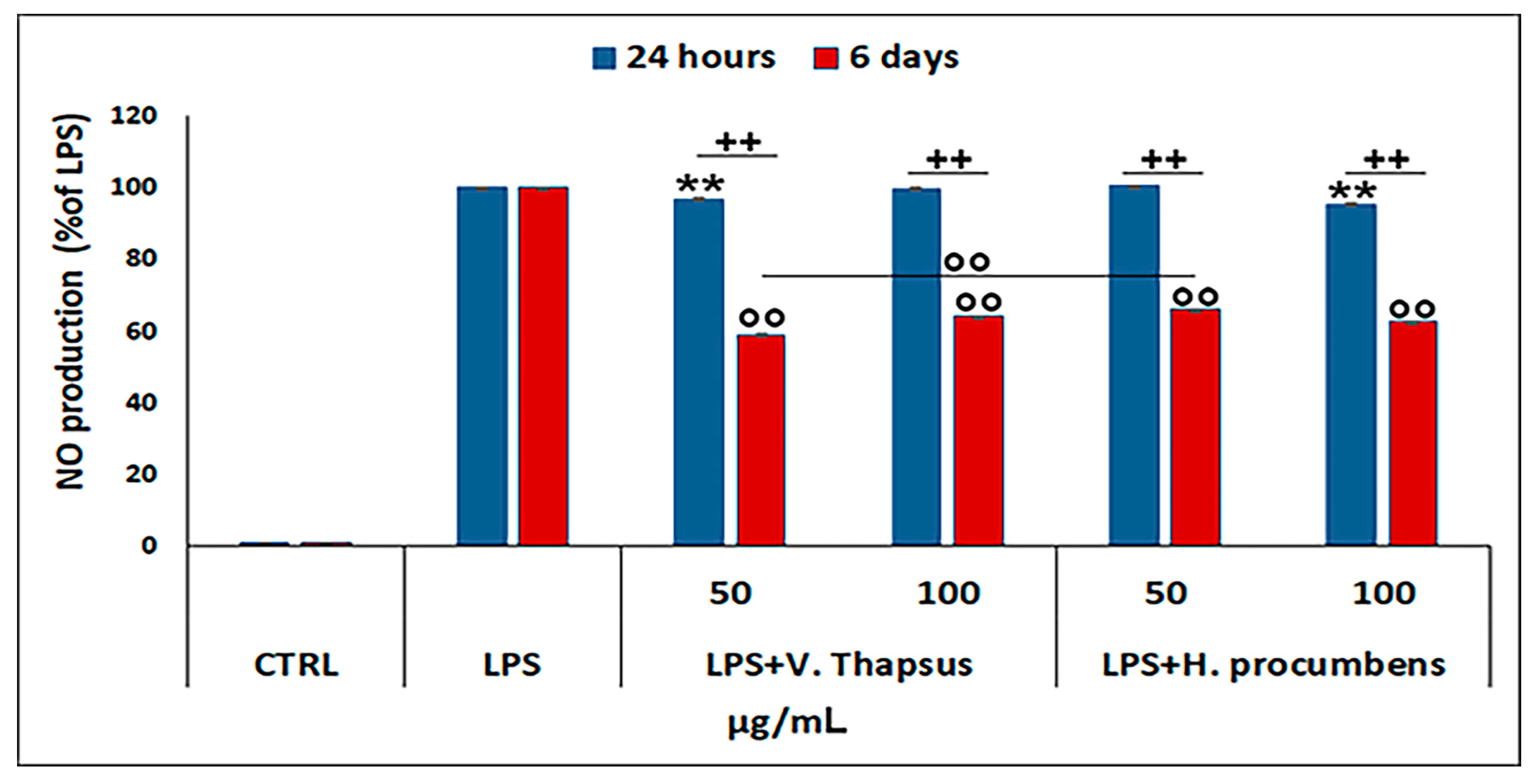
2.3. Evaluation of the Anti-Inflammatory Property of V. thapsus Extract on RAW 264.7 Cells
2.4. Evaluation of the Cytotoxic Effect of V. thapsus Extract on HC Cell Line
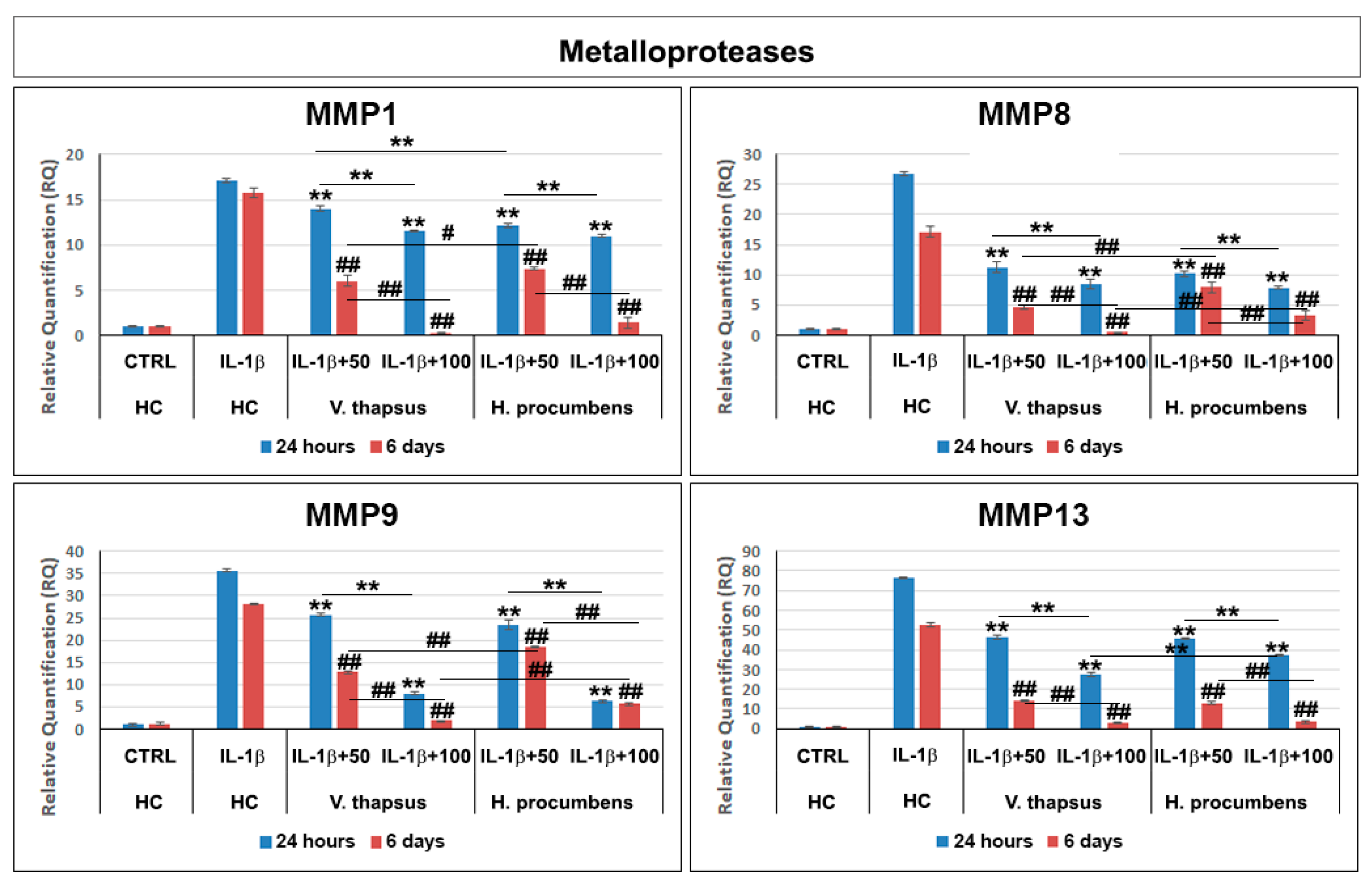
2.5. Anti-Osteoarthritis Effect of V. thapsus Extract
3. Discussion
4. Materials and Methods
4.1. Chemicals and Drugs
4.2. Plant Material and Extraction Procedure
4.3. Determination of Total Phenolic Content
4.4. Determination of Phenylethanoid Glycosides by HPLC-DAD
4.5. Cell Cultures
4.6. Cell Viability Assays
4.7. NO• Production
4.8. Quantitative Real-Time PCR (qRT-PCR)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chun, J.M.; Kim, H.S.; Lee, A.Y.; Kim, S.H.; Kim, H.K. Anti-inflammatory and antiosteoarthritis effects of Saposhnikovia divaricata ethanol extract: In vitro and in vivo studies. Evid.-Based Complement. Alternat. Med. 2016, 1984238. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Sofat, N.; Ejindu, V.; Kiely, P. What makes osteoarthritis painful? The evidence for local and central pain processing. Rheumatology 2011, 50, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Sgaglione, N.A. Biologic approaches to articular cartilage surgery: Future trends. Orthop. Clin. N. Am. 2005, 36, 485–495. [Google Scholar] [CrossRef]
- Yimam, M.; Lee, Y.C.; Kim, T.W.; Moore, B.; Jiao, P.; Hong, M.; Kim, H.J.; Nam, J.B.; Kim, M.R.; Oh, J.S.; et al. UP3005, a botanical composition containing two standardized extracts of Uncaria gambir and Morus alba, improves pain sensitivity and cartilage degradations in monosodium iodoacetate-induced rat on disease model. Evid.-Based Complement. Alternat. Med. 2015, 785638. [Google Scholar] [CrossRef] [Green Version]
- Henrotin, Y.; Lambert, C.; Couchourel, D.; Ripoll, C.; Chiotelli, E. Nutraceuticals: Do they represent a new era in the management of osteoarthritis?—A narrative review from the lessons taken with five products. Osteoarthr. Cartil. 2011, 19, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wegener, T.; Lüpke, N.P. Treatment of patients with arthrosis of hip or knee with an aqueous extract of devil’s claw (Harpagophytum procumbens DC.). Phytother. Res. 2003, 17, 1165–1172. [Google Scholar] [CrossRef]
- Menghini, L.; Recinella, L.; Leone, S.; Chiavaroli, A.; Cicala, C.; Brunetti, L.; Vladimir-Knežević, S.; Orlando, G.; Ferrante, C. Devil’s claw (Harpagophytum procumbens) and chronic inflammatory diseases: A concise overview on preclinical and clinical data. Phytother. Res. 2019, 33, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Denner, S.S. A review of the efficacy and safety of devil’s claw for pain associated with degenerative musculoskeletal diseases, rheumatoid, and osteoarthritis. Holist Nurs. Pract. 2007, 21, 203–207. [Google Scholar] [CrossRef]
- Chrubasik, S. Wirksamkeit pflanzlicher Schmerzmittel am Beispiel des Teufelskrallenwurzelextrakts [Devil’s claw extract as an example of the effectiveness of herbal analgesics]. Orthopade 2004, 33, 804–808. [Google Scholar] [CrossRef]
- Pignatti, S. Verbascum thapsus L. In Flora d’Italia; Edagricole: Milano, Italy, 2017; Volume 3, pp. 327–328. ISBN 8850652437. [Google Scholar]
- Turker, A.U.; Gurel, E. Common mullein (Verbascum thapsus L.): Recent advances in research. Phytother. Res. 2005, 19, 733–739. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Menghini, L.; Patruno, A.; Vinciguerra, I.; De Lutiis, M.A.; Felaco, M.; Felaco, P.; Grilli, A. Anti-inflammatory properties of the plant Verbascum mallophorum. J. Biol. Regul. Homeost. Agents 2009, 23, 189–195. [Google Scholar] [PubMed]
- Frezza, C.; Bianco, A.; Serafini, M.; Foddai, S.; Salustri, M.; Reverberi, M.; Gelardi, L.; Bonina, A.; Bonina, F.P. HPLC and NMR analysis of the phenyl-ethanoid glycosides pattern of Verbascum thapsus L. cultivated in the Etnean area. Nat. Prod. Res. 2019, 33, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Hu, X.P.; Zeng, Y.; Li, Y.; Wu, H.Q.; Qiu, R.Z.; Ma, W.J.; Li, T.; Li, C.Y.; He, Z.D. Advanced research on acteoside for chemistry and bioactivities. J. Asian Nat. Prod. Res. 2011, 13, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Pastore, S.; Lulli, D.; Fidanza, P.; Potapovich, A.I.; Kostyuk, V.A.; De Luca, C.; Mikhal’chik, E.; Korkina, L.G. Plant polyphenols regulate chemokine expression and tissue repair in human keratinocytes through interaction with cytoplasmic and nuclear components of epidermal growth factor receptor system. Antioxid. Redox Signal. 2012, 16, 314–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Z.; Yang, B. Phenylethanoid glycosides: Research advances in their phytochemistry, pharmacological activity and pharmacokinetics. Molecules 2016, 21, 991. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharmacother. 2020, 129, 110452. [Google Scholar] [CrossRef]
- Shen, C.L.; Smith, B.J.; Lo, D.F.; Chyu, M.C.; Dunn, D.M.; Chen, C.H.; Kwun, I.S. Dietary polyphenols and mechanisms of osteoarthritis. J. Nutr. Biochem. 2012, 23, 1367–1377. [Google Scholar] [CrossRef]
- Korkina, L.; Kostyuk, V.; De Luca, C.; Pastore, S. Plant phenylpropanoids as emerging anti-inflammatory agents. Mini Rev. Med. Chem. 2011, 11, 823–835. [Google Scholar] [CrossRef]
- Korkina, L.G.; Mikhal’chik, E.; Suprun, M.V.; Pastore, S.; Dal Toso, R. Molecular mechanisms underlying wound healing and anti-inflammatory properties of naturally occurring biotechnologically produced phenylpropanoid glycosides. Cell Mol. Biol. 2007, 53, 84–91. [Google Scholar] [PubMed]
- Wesa, A.; Galy, A. Increased production of pro-inflammatory cytokines and enhanced T cell responses after activation of human dendritic cells with IL-1 and CD40 ligand. BMC Immunol. 2002, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Papi, S.; Ahmadizar, F.; Hasanvand, A. The role of nitric oxide in inflammation and oxidative stress. Immunopathol. Persa 2019, 5, e08. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside--a review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef]
- Tian, X.Y.; Li, M.X.; Lin, T.; Qiu, Y.; Zhu, Y.T.; Li, X.L.; Tao, W.D.; Wang, P.; Ren, X.X.; Chen, L.P. A review on the structure and pharmacological activity of phenylethanoid glycosides. Eur. J. Med. Chem. 2021, 209, 112563. [Google Scholar] [CrossRef] [PubMed]
- Houard, X.; Goldring, M.B.; Berenbaum, F. Homeostatic mechanisms in articular cartilage and role of inflammation in osteoarthritis. Curr. Rheumatol. Rep. 2013, 15, 375. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Georgiev, M.I.; Cao, H.; Nahar, L.; El-Seedi, H.R.; Sarker, S.D.; Xiao, J.; Lu, B. Therapeutic potential of phenylethanoid glycosides: A systematic review. Med. Res. Rev. 2020, 40, 2605–2649. [Google Scholar] [CrossRef]
- Mehana, E.E.; Khafaga, A.F.; El-Blehi, S.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Yang, C.Y.; Chanalaris, A.; Troeberg, L. ADAMTS and ADAM metalloproteinases in osteoarthritis—Looking beyond the ‘usual suspects’. Osteoarthr. Cartil. 2017, 25, 1000–1009. [Google Scholar] [CrossRef] [Green Version]
- Genovese, C.; Addamo, A.; Malfa, G.A.; Acquaviva, R.; Di Giacomo, C.; Tomasello, B.; Ragusa, S.; Toscano, M.A.; Lupo, G.; Anfuso, C.D.; et al. Antioxidant and antimicrobial activities of Castanea sativa Mill. (Fagaceae) extract: New therapeutic perspectives. Plant Biosystem. 2021, 155, 1032–1040. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; La Mantia, A.; Pappalardo, F.; Ragusa, M.; Renis, M.; Di Giacomo, C. The antioxidant activities of Betula etnensis Rafin. ethanolic extract exert protective and anti-diabetic effects on streptozotocin-induced diabetes in rats. Antioxidants 2020, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Dolcimascolo, A.; Torrisi, F.; Zappalà, A.; Gulino, R.; Parenti, R. MiR-19a Overexpression in FTC-133 Cell Line Induces a More De-Differentiated and Aggressive Phenotype. Int. J. Mol. Sci. 2018, 19, 3944. [Google Scholar] [CrossRef] [Green Version]
- Tenuta, M.C.; Malfa, G.A.; Marco, B.; Rosaria, A.; Loizzo, M.R.; Dugay, A.; Bouzidi, C.; Tomasello, B.; Tundis, A.; Deguin, B. LC-ESI-QTOF-MS profiling, protective effects on oxidative damage, and inhibitory activity of enzymes linked to type 2 diabetes and nitric oxide production of Vaccinium corymbosum L. (Ericaceae) extracts. J. Berry Res. 2020, 10, 603–622. [Google Scholar] [CrossRef]
- Calabrese, G.; Ardizzone, A.; Campolo, M.; Conoci, S.; Esposito, E.; Paterniti, I. Beneficial effect of tempol, a membrane-permeable radical scavenger, on inflammation and osteoarthritis in in vitro models. Biomolecules 2021, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Vicari, L.; Calabrese, G.; Forte, S.; Giuffrida, R.; Colarossi, C.; Parrinello, N.L.; Memeo, L. Potential Role of Activating Transcription Factor 5 during Osteogenesis. Stem Cells Int. 2016, 5282185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, G.; Gulino, R.; Giuffrida, R.; Forte, S.; Figallo, E.; Fabbi, C.; Salvatorelli, L.; Memeo, L.; Gulisano, M.; Parenti, R. In vivo evaluation of biocompatibility and chondrogenic potential of a cell-free collagen-based scaffold. Front. Physiol. 2017, 8, 984. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peack | Compound | Wavelength (nm) | Ret. Time (min) | Content (mg/mL) |
|---|---|---|---|---|
| 1 | samioside | 330 | 30.37 | 52.3 |
| 2 | echinacoside | 330 | 33.80 | 42.1 |
| 3 | forsythoside B | 330 | 35.23 | 108.1 |
| 4 | verbascoside | 330 | 37.12 | 254.8 |
| 5 | iso-verbascoside | 330 | 40.61 | 142.9 |
| 6 | martynoside | 330 | 42.10 | 118.4 |
| Target | Forward | Reverse |
|---|---|---|
| Il-1ß | GGAGAATGACCTGAGCACCT | GGAGGTGGAGAGCTTTCAGT |
| Il-6 | AGTCCTGATCCAGTTCCTGC | CTACATTTGCCGAAGAGCCC |
| iNOS | ACAGCACATTCAGATCCCCA | GCCGAGATTTGAGCCTCATG |
| COX2 | TAGTACTCCCGGTTGAAGCC | ACGATGGGCATGAAACTGTG |
| MMP1 | CTGAAGGTGATGAAGCAGCC | AGTCCAAGAGAATGGCCGAG |
| MMP8 | ATGGACCAACACCTCCGCAA | GTCAATTGCTTGGACGCTGC |
| MMP9 | CGCAGACATCGTCATCCAGT | GGATTGGCCTTGGAAGATGA |
| MMP13 | CTATGGTCCAGGAGATGAAG | AGAGTCTTGCCTGTATCCTC |
| GAPDH | GGAAGGTGAAGGTCGGAGT | TGGGTGGAATCATATTGGAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calabrese, G.; Zappalà, A.; Dolcimascolo, A.; Acquaviva, R.; Parenti, R.; Malfa, G.A. Phytochemical Analysis and Anti-Inflammatory and Anti-Osteoarthritic Bioactive Potential of Verbascum thapsus L. (Scrophulariaceae) Leaf Extract Evaluated in Two In Vitro Models of Inflammation and Osteoarthritis. Molecules 2021, 26, 5392. https://doi.org/10.3390/molecules26175392
Calabrese G, Zappalà A, Dolcimascolo A, Acquaviva R, Parenti R, Malfa GA. Phytochemical Analysis and Anti-Inflammatory and Anti-Osteoarthritic Bioactive Potential of Verbascum thapsus L. (Scrophulariaceae) Leaf Extract Evaluated in Two In Vitro Models of Inflammation and Osteoarthritis. Molecules. 2021; 26(17):5392. https://doi.org/10.3390/molecules26175392
Chicago/Turabian StyleCalabrese, Giovanna, Agata Zappalà, Anna Dolcimascolo, Rosaria Acquaviva, Rosalba Parenti, and Giuseppe Antonio Malfa. 2021. "Phytochemical Analysis and Anti-Inflammatory and Anti-Osteoarthritic Bioactive Potential of Verbascum thapsus L. (Scrophulariaceae) Leaf Extract Evaluated in Two In Vitro Models of Inflammation and Osteoarthritis" Molecules 26, no. 17: 5392. https://doi.org/10.3390/molecules26175392