Abstract
Gynostemma pentaphyllum (Thunb.) Makino (GP), also named Jiaogulan in Chinese, was known to people for its function in both health care and disease treatment. Initially and traditionally, GP was a kind of tea consumed by people for its pleasant taste and weight loss efficacy. With the passing of the centuries, GP became well known as more than just a tea. Until now, numbers of bioactive compounds, including saponins (also named gypenosides, GPS), polysaccharides (GPP), flavonoids, and phytosterols were isolated and identified in GP, which implied the great medicinal worth of this unusual tea. Both in vivo and in vitro tests, ranging from different cell lines to animals, indicated that GP possessed various biological activities including anti-cancer, anti-atherogenic, anti-dementia, and anti-Parkinson’s diseases, and it also had lipid-regulating effects as well as neuroprotection, hepatoprotective, and hypoglycemic properties. With the further development and utilization of GP, the research on the chemical constituents and pharmacological properties of GP were deepening day by day and had made great progress. In this review, the recent research progress in the bioactive compounds, especially gypenosides, and the pharmacological activities of GP were summarized, which will be quite useful for practical applications of GP in the treatment of human diseases.
1. Introduction
Gynostemma pentaphyllum (Thunb.) Makino is a creeping perennial herb that belongs to the family Cucurbitaceae. As a well-known edible and medicinal plant, GP has a long history of application in oriental medicine since the Ming dynasty [1]. The famous classical book of Chinese material medica, Compendium of Materia Medica, first recorded the usage and curative effect of GP [2]. In 1986, GP was listed by the Ministry of Science and Technology as the first “precious Chinese medicine” to be developed in the “Spark Program”. Due to its extensive biological activities, GP was brought into the list of functional foods by Ministry of Public Health of China on 5 March 2002. Currently, products containing GP had been marketed in many Asian countries and US [3]. In recent years, GP has attracted great attention owing to its wide biological activities with minimal pharmacological toxicity [4]. GP had been used in treating various diseases, such as hyperlipidemia, hyperglycemia, cancer, and so on, as shown in Figure 1. To conduct a comprehensive review, a great deal of literature on GP was obtained by searching the PubMed and China National Knowledge Infrastructure (CNKI) databases. In this paper, the research progress on chemical constituents and pharmacological properties of GP were introduced, which could provide reference for the further development of GP.
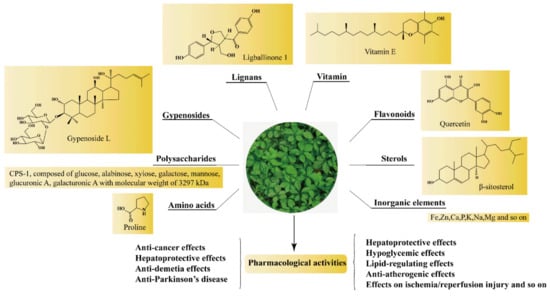
Figure 1.
The main active components and pharmacological activities of GP.
2. Chemical Constituents
Phytochemical studies revealed that the chemical constituents in GP included GPS, GPP, flavonoids, phytosterols, amino acids and inorganic elements, and so on (Figure 1). The detailed information of the main constituents of GP follows.
2.1. Gypenosides
Gypenosides isolated from GP were believed to be the major active constituents responsible for its various biological activities and reported clinical effects. In 1976, the dammarane-type saponins from GP were isolated by Japanese scholars for the first time [5]. By 2017, nearly 201 dammarane-type gypenosides had been reported in GP [6]. Since 2017, 47 new dammarane-type gypenosides had been isolated and identified, the structures are shown in Figure 2, eight of which were consistent with ginsenosides Rb1, Rb3, F2, Rg3, Rc, Rd, malonyl-Rb1, and malonyl-Rd, respectively. These ginsenosides made up around 25% of the total gypenosides in the plant, and GP was the first plant containing ginseng saponins outside of the Araliaceae family. Therefore, GP has been praised as “southern ginseng” [2]. In addition, two cucurbitane-type saponins were found in GP [7,8], and their structures are shown in Figure 3.
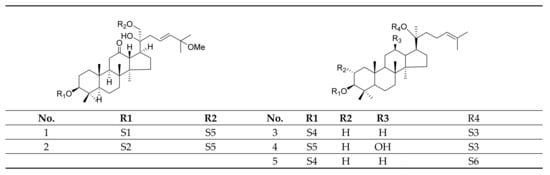
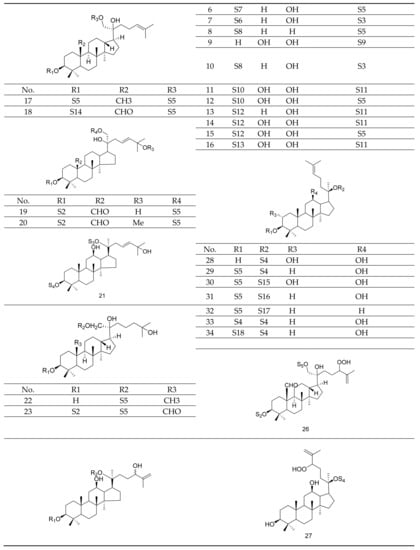
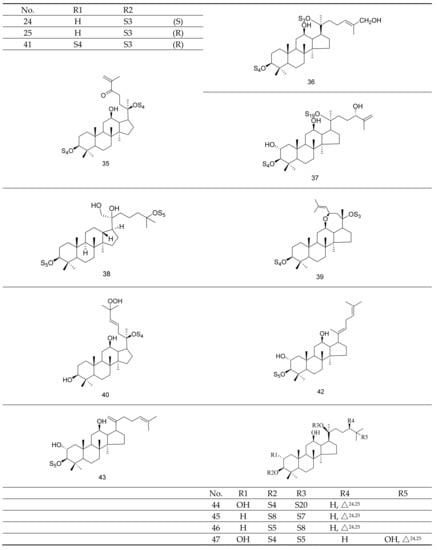
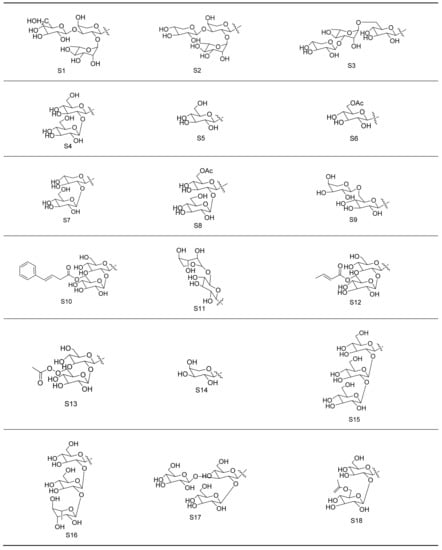
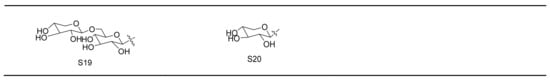
Figure 2.
Structures of new dammarane-type saponins.
Figure 3.
Structures of two cucurbitane-type saponins.
2.2. Polysaccharides, Flavonoids, Sterols, Amino Acids, and Inorganic Elements
So far, polysaccharides with various monosaccharide constituents and chemical structures have been isolated from GP. The molecular weight, monosaccharide composition, and chemical structures could be influenced by both different extraction/purification techniques [9]. Polysaccharides were mainly found in the stems and leaves of GP [10]. Ma et al. [11] determined the content of polysaccharide in different parts of GP, and the results showed that the content of polysaccharide in leaves (1.78 ± 0.60) % was higher than in stems (0.84 ± 0.23) %. GPP exhibited almost no toxic side effects on the human body but possessed broad application prospects in preventing and treating various diseases. Although flavonoids were secondary metabolites in the medicinal herb kingdom, there are few reports about flavonoids of GP. So far, more than 10 kinds of flavonoids have been reported. Ling et al. [12] investigated the comprehensive chemical profiles of GP for the first time using HPLC-ESI-QTOF-MS, and seven flavonoid glycosides were identified. From 1986 to 2020, about eighteen sterols were isolated from GP. Studies had shown that GP contained 18 kinds of amino acids, including eight kinds of essential amino acids [13]. However, the content of amino acids in GP varied from place to place. In addition, GP contained 23 kinds of inorganic elements, among which 13 kinds of trace elements and five kinds of major elements were essential to human body [13].
3. Pharmacological Properties of GP
3.1. Anti-Cancer Effect
To date, numerous studies have demonstrated the strong anti-cancer effect of the main components of GP, such as GPS, GPP, and flavonoids (as shown in Table 1). There are several mechanisms of GP’s anti-cancer effect, as shown in Figure 4.

Table 1.
Anti-cancer effect of GP.
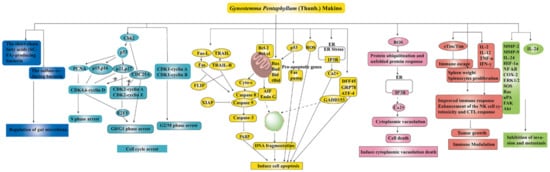
Figure 4.
The anti-cancer mechanisms of GP ↑ meant up-regulation, ⊥ meant down-regulation.
3.1.1. Immune Modulation
Tumors are closely related to the state of the body’s immune system, and strengthening immune function played an important role in the treatment of tumors. GPP (50 and 100 mg/kg for 12 days, i.g.) had an inhibitory effect on tumor growth in mouse forestomach carcinoma (MFC) gastric tumor-bearing mice, for which the tumor quality of the GPP group was significantly reduced. Furthermore, GPP could effectively prevent the immune organ atrophy and hypofunction of MFC gastric tumor-bearing mice, suggesting that the anti-tumor effect of GPP was closely associated with immune enhancement [38]. Liu et al. investigated the effect of GPS on the tumor growth in tumor-bearing mice, and the results showed that the growth of the tumor was inhibited by the intraperitoneal injection of 10–40 mg/kg GPS, and 20 mg/kg GPS exerted the strongest inhibitory effect. In addition, the number of splenic lymphocytes was increased, and the activity of natural killer (NK) cells was enhanced [39]. Studies had demonstrated that soluble Tim-3 played an important role in inhibiting the immune escape in a variety of cancer. Gypenoside XLIX (20 and 80 µg) could reduce the immune escape of non-small cell lung cancer A549 cells by promoting the stim-3/Tim-3 ratio [40].
3.1.2. Induction of Cell Apoptosis
Inducing apoptosis was one of the effective ways to treat tumor, which could improve the recovery of tumor patients. Yan et al. [30] evaluated the cytotoxicity of GPS in human colorectal cancer SW-480 cells, and the results showed that GPS (70–130 µg/mL) could decrease the percentage of viable cells and increase the plasma membrane permeability of SW-480 cells in a dose- and time-dependent manner. More importantly, reactive oxygen species (ROS) play an important role in GPS-induced cell toxicity and apoptosis. After the GPS treatment, the level of intracellular ROS level was increased, and DNA fragmentation and apoptotic morphology were observed. Liu et al. [41] investigated the effects of GPS on renal cell carcinoma (RCC) and its molecular mechanism for the first time. Studies concluded that the cell viability of 786-O and Caki-1 treated with gradient concentration of GPS (0, 75, 150, 300, 600 µg/mL) for 24 h showed a significant decrease. In addition, GPS induced the apoptosis of RCC cells through regulating the phosphoinositide 3-kinases/protein kinase B/mammalian target of rapamycin (PI3K/Akt/mTOR) signaling pathway.
3.1.3. Inhibition of Cell Migration
Metastasis was believed to be one of the major challenges for a successful cancer treatment and the prevention of cancer. GPS exerted an inhibitory effect on cell migration in vitro in a dose-dependent manner. GPS could induce the microfilament network collapse and injure the cell shape and migration ability [30]. In addition, damulin B was reported to inhibit the migration of human lung cancer A549 and H1299 cells by down-regulating the protein production of matrix metalloproteinase-2 and -9 (MMP-2 and -9) [35].
3.1.4. Regulation of Gut Microbiota
At present, accumulating evidence indicates that targeted intervention in the gut microbiota composition had shown encouraging results in cancer prevention and treatments. It was demonstrated that treatment with GPS significantly boosted the growth of short-chain fatty acids-producing bacteria that played an important role in maintaining the colonic health, and it down-regulated the relative abundance of sulfate-reducing bacteria, which were known to produce hydrogen sulfide and contribute to damage of the intestinal epithelium or even promote cancer progression in ApcMin/+ mice. Therefore, GPS exerted cancer-preventive effects and had the potential to be a new preventive medicine against colorectal cancer [42].
3.1.5. Cell Cycle Arrest
Gypenoside LI, a gypenoside monomer from GP, had a potent cytotoxic effect on melanoma cells by arresting the cell cycle at the S phase and inhibiting melanoma cell proliferation through the up-regulation of miR-128-3p [14]. Studies had shown that GPS could induce G0/G1 arrest in A549 cells. The molecular mechanism of cell cycle arrest was investigated by Western blotting, showing that cyclin-dependent protein kinase inhibitors such as p16, p21, p27, and p53 proteins were accordingly increased with the increasing time of incubation with GPS in the A549 cells [20]. Flavonoids extracted from GP were reported to induce cycle arrest at both S and G2/M phases with the concurrent modulated expression of the cellular proteins cyclin A, B, p53, and p2, and then inhibit the proliferation of lung cancer A549 cells [43].
3.1.6. Others
A growing amount of evidence had proven that senescence was an effective tumor-suppressive approach in cancer treatment. Gypenoside L could inhibit the proliferation of cancer cells by inducing senescence. Chronic administration of chemotherapeutic drugs resulted in the resistance of cancer cells due to protective autophagy. However, gypenoside L could render cancer cells more sensitive to chemotherapy such as cisplatin by impairing the protective autophagy. Based on this, gypenoside L could be explored as a promising agent for solving the problem of resistance [16].
3.1.7. Toxicity on Normal Cells and Cancer Cells
Studies had shown that GPS was capable of exerting different alternative cytotoxicity in cancer cells and normal cells. Gypensapogenin H isolated from the hydrolysate of total saponins of GP exhibited potent growth inhibitory effects on tumor cells. It significantly inhibited the growth of human breast cancer cells, whereas it showed lower toxicity to normal human breast epithelial cells than the commonly used anti-cancer drugs paclitaxel and 5-Fu [36]. The above result was consistent with a study conducted to explore the cytotoxicity of GPS on normal peripheral blood mononuclear cells [29]. In addition, different cancer cells exerted different sensitivity to the sham gypenoside. The cytotoxic activities of gypsapogenin A isolated from the acid hydrolysate of total saponins against HepG2 and A549 cells were evaluated, and the results showed that the IC50 values of gypsapogenin A against HepG2 and A549 cells were 24.12 and 59.81 µM, respectively [44].
3.1.8. Relationship between Anti-Cancer Activity and Structure of GPS
The anti-cancer activity of GPS was associated with its structure. Xing et al. [19] evaluated the cytotoxic activities of several dammarane saponins (gypenoside L, gypenoside LI, gypenoside LVI, and gypenoside XLVI) on A549 cells with ginsenoside as a positive control by CCK-8 assay and found that the loss of sugar might be related to the enhanced inhibition of cancer cell proliferation. Once the free hydroxyl in the C20 position was glycosylated, the anti-cancer activity was extremely decreased. As shown in Figure 5, gypenoside stereoisomers, gypenoside L (S configuration at C20), and gypenoside LI (R configuration at C20), both with free hydroxyls in the C20 position, showed stronger activity with IC50 values of 29.38 ± 2.52 µM and 21.36 ± 0.78 µM respectively against A549 cells compared with gypenoside LVI and gypenoside XLVI glycosylated in the C20 position. Cui et al. [45] evaluated the cytotoxic activities of damulin E and damulin F (their structures are shown in Figure 5) against A549 cells and showed that after being incubated for 24 h, compared with positive control (ginsenoside Rg3), damulin F showed stronger cytotoxic activity against A549 cells with the IC50 value of 19.8 ± 0.4 µM, whereas damulin E appeared to have weaker activity with the IC50 value of 38.9 ± 0.6 µM. Therefore, saponins with a double bond in C20 (21) exhibited stronger activities than saponins with a double bond in C20 (22). However, some studies [44,46,47,48] did not mention the IC50 value or positive control, and the cytotoxic activities could not compare, so the relationship of the structure and activities needed further investigation.
Figure 5.
Relationship between anti-cancer activity and structure of GPS.
3.2. Anti-Atherogenic Effect
Accumulating evidences had demonstrated that GP could be developed as potential and promising agents for atherosclerosis (AS) treatment. The detailed parameters of experiments including experimental models, dosage, and possible mechanisms were listed in Table 2. The anti-atherogenic mechanisms are shown in Figure 6, which were mainly related to the reduction of foam cell formation, inflammation, DNA damage, and endothelial cells (ECs) apoptosis and the increasement of autophagy and cholesterol flux.
Table 2.
Anti-atherogenic effect of GP.
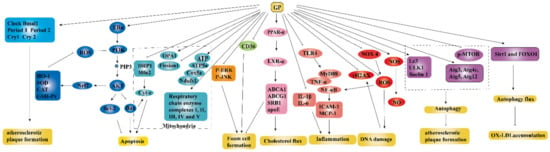
Figure 6.
Anti-atherogenic effect of GP ↓ meant up-regulation, ⊥ meant down-regulation.
Inflammation occurred at different stages of AS progression. Therefore, anti-inflammatory therapy had become an important intervention to delay the progression of the disease. Recent studies had shown that the expression of TLR4 (Toll-like receptor 4) and its downstream signaling molecules MyD88 (Myeloid differentiation factor 88), NF-κB, and TNF-α in aortic tissue were inhibited after GP intervention (80 and 160 mg/kg for 10 weeks, i.g.), thus alleviating vascular inflammation [61]. Studies had demonstrated that phytoestrogens, compounds with structural similarities to estradiol, could bind weakly to estrogen receptors and play a major role in the prevention and treatment of AS. Gypenoside XVII (GP-17), a novel phytoestrogen, had a similar structure to estradiol, and its anti-atherogenic effect of GP-17 had been verified in vivo and vitro. GP-17 treatment (50 mg/kg) significantly decreased blood lipid levels, increased the expression of antioxidant enzymes, and decreased atherosclerotic lesion size, thus effectively preventing the progression of atherosclerosis in ApoE-/- mice. Oxidized low-density lipoprotein (Ox-LDL) in the pathogenesis of atherosclerosis was an exceptionally important risk factor, which contributed to endothelium apoptosis. GP-17 (50 µg/mL) significantly inhibited Ox-LDL-induced ROS generation and apoptosis in human umbilical vein endothelial cells (HUVECs) by the estrogen receptor alpha (ERα)/PI3K/Akt pathway. Furthermore, GP-17 had been verified as a selective ERα modulator for atherosclerosis treatment [49].
With the deepened understanding of the pathogenesis of AS, the inhibition of the formation of macrophage foam cells and atherosclerotic plaque had been paid more attention for new drug development. The formation of macrophage foam cells was through scavenger receptor-mediated endocytosis of Ox-LDL. CD36 receptor was the most important scavenger receptor in the macrophage cell membrane to mediate the endocytosis of Ox-LDL. Studies had shown that the expression of the scavenger receptor of CD36 in macrophages was downed by GPS intervention (100 µg/mL), whereby the intake of Ox-LDL in macrophages was reduced [50]. In addition, Shen et al. confirmed the inhibitory effect of gypenoside LVI (GPLVI) on Ox-LDL-induced foam cell formation for the first time. Treatment with GPLVI (25, 50, and 100 µg/mL) significantly promoted cholesterol efflux by increasing the expression of ATP-binding cassette transporter G1 (ABCG1), scavenger receptor class B type I (SRB1), and liver X receptor (LXR) α, thus reducing the formation of foam cells [52]. Several studies had shown that the activation of LXR could inhibit the development of atherosclerosis in mice, induce ATP-binding cassette transporter A1 (ABCA1) and apolipoprotein E gene expression, and increase cholesterol efflux, which lead to a decreased cholesterol burden in the arterial wall, as well as enhanced high-density lipoprotein (HDL) levels [63]. Gynosaponin TR1 was reported to be an agonist of LXR with selectivity for LXR-α over LXR-β, and this selectivity of TR1 toward LXR-α might prevent the elevation of triglycerides (TG) [63]. Recent studies had found that autophagy was closely related to AS, and moderate autophagy could prevent macrophages from transforming into foam cells to delay the occurrence and development of AS. Studies had shown that GPS (2.973 g/kg for 4 weeks, i.g.) might enhance the level of autophagy of AS model mice by regulating the mTOR/unc-51-like kinase 1 (ULK1) autophagy signaling pathway, alleviate the formation of atherosclerotic plaque, and prevent atherosclerosis [53].
3.3. Lipid-Regulating Effect
3.3.1. Effect on Hyperlipidemia
A large number of studies had shown that GP had the effect of lowering blood lipid and was commonly used together with other traditional Chinese medicines or combined with Western medicines to treat hyperlipidemia. GPS was the major component that played an important role in lowering blood lipid levels. It was found that the nucleus structure of GPS was very similar to that of endogenous bile acids, which could activate the farnesoid X receptor in the liver, thus up-regulating the key enzymes CYP8B1 and CYP7A1 in bile acid synthesis, promoting the synthesis and secretion of bile acids, promoting lipid metabolism, and lowering blood lipid [64]. Mao [65] studied the lipid-lowering effect of gypenoside SL-1 and SL-2 isolated from GP on HepG2 cells in vitro. The results showed that SL-1 and SL-2 (10, 50, and 100 µg/mL) could decrease the content of TC, TG, and low-density lipoprotein (LDL) and increase the content of HDL, thus reducing lipid accumulation in HepG2 cells incubated with oleic acid. Currently, the first-line drug used for the treatment of hyperlipidemia was statins. However, its lipid-lowering effect could not reach the target proposed by the National Institutes of Health of the United States [66]. That was because the expression of proprotein convertase subtilisin/kexin 9 (PCSK9) while lowering lipids was enhanced, which further induced the increased low-density lipoprotein receptor (LDLR) degradation, thus inhibiting the lipid-lowering effect [67]. A number of clinical studies had reported that statin therapy might cause the increase in transaminase in the body while lowering lipid levels, thus affecting liver function [66]. Studies had demonstrated that GPS could significantly lower blood lipids, and when combined with simvastatin, the effect of lowering lipid levels was improved by suppressing the increased expression of PCSK9 and reducing the degradation of LDLR. In addition, the increase in serum transaminase induced by simvastatin was reversed, and liver function was improved [67,68]. Yin et al. isolated and identified ten novel saponins named yunnangypenosides (1–10) and a known one from GP. Furthermore, these compounds could significantly reduce lipid levels except for those of 2 [69].
3.3.2. Anti-Obesity Effect
More and more evidence has shown that GP also possessed a potent anti-obesity effect. GP extract (GPE) (300 mg/kg) with much higher contents of gypenoside, gypenoside LI, and ginsenoside Rg3 exhibited a stronger anti-obesity effect than the positive group (Orlistat, 30 mg/kg). When GPE was given, the increases in body weight, fat mass, white adipose tissue, and adipocyte hypertrophy were suppressed, and the levels of serum triglyceride, total cholesterol, and low-density lipoprotein-cholesterol were lower than the high-fat-diet group. The lipid-lowering effect was associated with AMP-activated protein kinase (AMPK) activation, which led to increased SIRT1 expression [70]. GPE (300 mg/kg) named actiponin containing large amounts of damulin A and damulin B were successfully delivered to the skeletal muscle and liver, and it prevented or improved obesity in ob/ob mice by stimulating fatty acid oxidation and activating AMPK in these organs. Actiponin (200 mg/kg) reduced body weight and plasma total cholesterol level without any effect on food intake [71]. In addition, GPS could prevent high-fat diet-induced obesity by promoting energy expenditure. When treated with GPS (300 mg/kg), brown adipocyte tissue activity and white adipose tissue browning were increased [72]. Inhibiting adipogenesis was effective and promising for the treatment of obesity as well. An isolated novel saponin JS (100 µM) could inhibit adipocyte differentiation and adipogenesis, which might be associated with Wnt/β-catenin signaling activation [73]. Recently, a double-blind, randomized, clinical trial with a treatment duration of 16 weeks was conducted to assess the efficacy and safety of a commercially available capsule-form herbal supplement containing GPE on improving body composition in overweight males and females. Studies showed that GPE (450 mg/d) was capable of altering fat mass and fat distribution in overweight and obese males and females compared to a placebo. Following 16 weeks of treatment, a significant reduction in total body weight and total fat mass was observed. There was a significant difference between males and females. Males had a significant reduction in visceral fat and females had a significant reduction in gynoid fat [74]. It was proved that a high-fat diet can lead to changes of gut microbiota, which might further contribute to obesity. High ratio of Firmicutes to Bacteroidetes had been found in obese mice and human adults when compared to lean mice and human adults. GPS treatment (100 and 300 mg/kg) significantly decreased the ratio of Firmicutes to Bacteroidetes by 20% and 58.6% respectively and increased Akkermansia muciniphila abundance in the gut microbiota, thereby preventing the development of obesity [72]. At present, the application of GPS was mainly for internal use. GPS had an obvious lipid-lowering effect for external use as well, which could meet different needs aimed at the uneven distribution of fat in the body [75].
When considering these findings together, GP is a promising traditional Chinese medicine (TCM) used for hyperlipidemia and obesity.
3.4. Neuroprotective Effect
3.4.1. Effect on Alzheimer’s Disease
Alzheimer’s disease (AD) is a common degenerative neurological disease, which is mainly manifested in the severe decline of cognitive function and memory. The characteristic pathological changes of AD include extracellular senile plaques formed by beta-amyloid protein (Aβ) deposition, neurofibrillary tangles formed by Tau protein hyperphosphorylation, and neuronal loss accompanied by glial cell proliferation. There were many theories about the pathogenesis of AD, including abnormal deposition of Aβ, Tau protein hyperphosphorylation, and so on [76]. At present, there was no specific cure drug, and only symptomatic treatment and therapeutics to delay the progression of AD were given to AD patients.
Aβ abnormal deposition was a crucial factor that drives AD pathogenesis, which was associated with neuroinflammation, oxidative stress, cell apoptosis and autophagy, etc. GPS could improve the learning and memory ability in dementia mice induced by D-galactose combined with sodium nitrite and aluminum trichloride, and the mechanism underlying might be related to the reduced oxidative damage and Aβ expression in brain tissues. Compared with the model group, the activity of SOD and GSH-Px were significantly increased, and the level of MDA was significantly decreased in brain tissues of the GPS group (250 mg/kg). In addition, the expression of Aβ42 decreased by treatment with GPS [77]. Under normal circumstances, microglial cells were in a static state, which might be activated into a classic activated state (M1 state) or alternative activated state (M2 state) when the body was in a pro-inflammatory environment for a long time. The former was harmful; in contrast, the latter was beneficial. A high level of Aβ in the brain could activate microglial cells to secrete pro-inflammatory factors and eventually develop into chronic inflammation. Treatment with 50 mg/L GPS for 24 h could attenuate Aβ-induced inflammation by reducing microglial activation and shifting microglial M1 to M2 state, and the process might be mediated by suppressors of cytokine signaling 1 (SOCS1) [78]. GPS also could improve cognitive dysfunction and chronic inflammation caused by injecting lipopolysaccharide in the hippocampus. GPS treatment (25, 50, and 100 mg/kg) for 21 days significantly decreased pro-inflammatory mediators such as IL-6, IL-1β, and NF-κB levels in the brain [79]. Pretreatment with GPS could effectively inhibit the Aβ1-40-induced increase in inducible nitric oxide synthase (iNOS) expression and cell apoptosis. Compared with Aβ1-40 group, the growth condition of cells incubated with GPS was improved obviously, the number of neurons was increased, and the synapses between adjacent neurons were more and compact [80]. Jia et al. investigated the neuroprotective effect against Aβ-induced cytotoxicity of a purified polysaccharide GPP1 in PC12 cells. The results showed that GPP1 exerted its protective effect via inhibiting oxidative stress and suppressing the mitochondrial apoptotic pathway [81]. Phytoestrogens had been proved to be protective against Aβ-induced neurotoxicity and regarded as a relatively safe agent for AD treatment. GP-17, as a novel phytoestrogen isolated from GP, had reported its neuroprotective effect [82,83]. GP-17 promoted the autophagy-based elimination of Aβ and prevented the formation of Aβ plaques in the hippocampus and cortex of APP/PS1 mice [82]. GP-17 exerted protective effect against Aβ25-35-induced neurotoxicity in nerve growth factor-induced differentiation of PC12 cells, including oxidative stress, apoptosis, and autophagic cell death, which might be related to estrogen receptor-dependent activation of PI3K/Akt pathways, inactivation of GSK-3β, and activation of nuclear factor-erythroid 2-related factor 2/antioxidant responsive element/heme oxygenase 1 (Nrf2/ARE/HO-1) pathways [83].
Most scholars believed that the damage of the cholinergic system in the brain was the main possible mechanism for the formation of AD, thus putting forward the “cholinergic hypothesis”. Cholinergic neurotransmitters were important for normal learning and memory. A decrease in cholinergic function, particularly within the basal forebrain, could result in a decline in memory and cognitive function with age. Scopolamine, an anticholinergic drug, caused memory impairments. The memory-enhancing effects of Gypenoside TN-2 and Gypenoside LXXIV (G-74) were investigated in scopolamine-treated mice in passive-avoidance and Morris water maze tests. Studies had demonstrated that TN-2 and G-74 potently ameliorated memory and learning impairment caused by scopolamine [84,85].
3.4.2. Effect on Vascular Dementia
Vascular dementia (VD) was a clinical syndrome caused by various cerebrovascular disease and characterized by cognitive, learning, and memory impairments. VD was the second-most-common dementia in the elderly, after Alzheimer’s disease. With the aging of China’s population, the incidence of vascular dementia is increasing year by year. Large amounts of studies had demonstrated that the neurons injury or apoptosis were observed in the hippocampus of VD mice, which might be one of the important mechanisms of dementia. GPS had been reported to significantly improve learning and memory impairment and protect the neurons by inhibiting the expression of apoptosis-related protein P38 and caspase-3 [86,87]. It was found that the level of phosphorylated cAMP response element-binding protein (PCREB) in the hippocampus could affect the plasticity of synapses and the ability of learning and memory. The expression level of PCREB mRNA in the hippocampus of VD model group was significantly lower than that of sham operated group. By intraperitoneal injection of 100, 200, and 400 mg/kg GPS, the expression of PCREB was significantly up-regulated, suggesting that the decrease in hippocampal neuron damage and the improvement of learning and memory ability in VD mice might be related to the up-regulation of PCREB expression in hippocampal neurons [88]. In addition, the effect of GPS on improving cognitive function was closely associated with its antioxidant and anti-inflammatory activities. By GPS treatment (100 and 200 mg/kg for 61 days), the oxidative neuronal damage was ameliorated, and the activation of inflammatory astrocytes was reduced in the cortex and hippocampus after chronic cerebral hypoperfusion [89]. Together, GPS had therapeutic potential for the treatment of VD and might serve as a potential new anti-dementia agent.
3.4.3. Anti-Parkinson’s Disease
Parkinson’s disease (PD) is a chronic neurodegenerative disease, which mainly occurs in middle-aged and elderly patients. The main clinical manifestation included bradykinesia, static tremor, autonomic nerve dysfunction, and myotonia. To date, large amounts of studies had demonstrated the anti-Parkinson’s property of GP, as shown in Table 3.
Table 3.
Anti-Parkinson’s disease effect of GP.
Studies on the pathogenesis of PD had demonstrated that the loss of dopaminergic neurons in the brain of PD patients was closely related to oxidative stress [96]. The 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced decreases of glutathione content and SOD activity in the substantia nigra, which resulted in oxidative stress, loss of nigral dopaminergic neurons, and motor dysfunction, were all attenuated by treatment with GPS [90]. In vitro, GPS protected primary cultured dopaminergic neurons against 1-methyl-4-phenylpyridinium ion (MPP+)-induced oxidative injury as well. It was found that GPS pretreatment had stronger protective effect than GPS co-treatment or post-treatment [91]. L-3, 4-Dihydroxyphenylalanine (L-DOPA) was the natural precursor of dopamine and was the most effective therapy for PD. However, chronic L-DOPA administration resulted in a loss of drug efficacy and irreversible adverse effects including motor and non-motor dysfunctions in both patients with PD and animal models of PD [92,93]. Several studies had reported that GP could ameliorate L-DOPA-induced adverse effects and might be developed as a useful agent for adjuvant therapeutics of PD receiving long-term treatment with L-DOPA. GPS (25 and 50 mg/kg) and GPE (50 mg/kg) could effectively attenuate the development of L-DOPA-induced-dyskinesia without compromising the anti-Parkinson effects of L-DOPA by reducing ΔFosB expression and ERK1/2 phosphorylation in the 6-hydroxydopamine (6-OHDA)-lesioned rat model of PD [92]. Studies showed that MPTP-lesioned mice exhibited deficits associated with habit learning and spatial memory, which were further aggravated by treatment with L-DOPA (25 mg/kg). However, treatment with GPS (50 mg/kg) ameliorated memory deficits via increasing the activation of the dopaminergic neuronal system and modulating the phosphorylation of the N-methyl-D-aspartate (NMDA) receptor-mediated signaling system [93]. In addition, GPS (50 mg/kg) and GPE (50 mg/kg) improved the symptoms of anxiety disorders in the MPTP-lesioned rat model of PD with or without L-DOPA treatment [94].
3.5. Effects on Ischemia–Reperfusion(I/R) Injury
Tissue ischemia–reperfusion (I/R) injury is the damage caused by excessive free radical attacks on cells in tissues after blood flow recovery on the basis of ischemia. Numerous evidence had confirmed that the saponins and flavonoids in GP exhibited the improvement effect on tissue I/R injury, including renal, hepatic, and myocardial I/R injury (as shown in Table 4).
Table 4.
Effects on ischemia–reperfusion(I/R) injury.
It had been demonstrated that the generation of I/R injuries is closely associated with inflammation, oxidative damage, and cell apoptosis. Studies in vitro and in vivo had shown that the effect of GPS on I/R-induced injury was mainly attributed to its anti-inflammation, anti-oxidative, and anti-apoptotic activity. An oral treatment with 50 mg/kg GP 1 h prior to ischemia effectively attenuated the I/R-induced increase in the levels of hepatic lipid peroxidation and serum alanine aminotransferase and suppressed the I/R-induced increase in the pro-apoptotic protein levels of Bax and cytochrome c and the activity of caspase-3/8, as well as the I/R-induced decrease in the levels of anti-apoptotic protein Bcl-2 [102]. Ye et al. [101] evaluated the protective effect against I/R-renal injury and its underlying molecular mechanisms. Pretreatment with 50 mg/kg GP significantly inhibited I/R-induced up-regulation of concentrations of serum creatinine and urea nitrogen. Furthermore, pretreatment with GP inhibited the I/R-induced production of pro-inflammatory cytokines, oxidative damage, and apoptosis. In myocardial I/R injury, GPS was shown to reduce infarct size and alleviate the impairments in cardiac structure and function, which was associated with the preservation of mitochondrial function and reduction of oxidative stress in the myocardial cells [98]. In addition, studies had shown that gypenoside had strong capacity to inhibit ER stress-induced cardiomyocyte apoptosis through the blockade of C/EBP-homologous protein (CHOP) pathway and activation of PI3K/Akt pathway [100]. A study by Yu et al. showed that GP protected cardiomyocytes against I/R injury by inhibiting NF-κB activation via the MAPK signaling pathway [97]. Another study by Chang et al. confirmed that gypenoside A functioned its protective effect on myocardial I/R injuries by suppressing miR-143-3p level via the activation of AMPK/FOXO1 pathway [99]. These studies indicated that the effects of GPS on I/R injuries could be multi-pronged.
3.6. Hepatoprotective Effects
At present, numerous studies had demonstrated that GP had the strong protective property for various kinds of liver injury (as shown in Table 5).
Table 5.
Hepatoprotective effects of GP.
3.6.1. CCl4-Induced Liver Injury
Researchers reported that GPP possessed a protective effect against hepatic injury in rats caused by carbon tetrachloride (CCl4). GPP (40 and 80 g/kg for 30 days, i.g.) could stabilize the liver cell membrane, effectively prevent the leakage of substance in cytoplasm, and reduce the level of serum transaminase. In addition, the up-regulated expression of iNOS mRNA in liver tissues of a rat model, which led to the production of nitric oxide (NO) and further induced liver injury, was attenuated following GPP treatment [103]. CCl4 was metabolized by cytochrome P-450 in the liver endoplasmic reticulum to produce a free radical that could lead to lipid peroxidation, liver damage, and necrosis. GPS exhibited a potent protective effect on CCl4-induced acute and chronic liver damage, which was attributed to its free radical scavenging capacity [104,105]. In addition, GPS was reported to also possess anti-fibrotic potential on CCl4-induced liver fibrosis in vivo and vitro studies, which supported its clinical use in alleviating the progression of hepatic fibrosis. GPS could improve the collagenolytic activity, decrease the deposition of collagen in the liver, promote the proliferation of liver cells, and thus suppress the onset and enhance the recovery of liver fibrosis induced by CCl4 in rats [104]. Chen et al. [106] investigated the effects and mechanisms of GPS on ameliorating liver fibrosis induced by CCl4 and found that the transforming growth factor-β (TGF-β) signaling was inhibited by treatment of GPS, consequently suppressing the proliferation of hepatic progenitor cells and inhibiting their differentiation into myofibroblasts.
3.6.2. Exhaustion Exercise-Induced Liver Injury
Generally, extreme exercise leads to the generation of a large number of free radicals, and when the generated free radicals exceed the scavenging capacity of the body’s anti-oxidation defense system, the body is in the state of oxidative stress and further induces cell apoptosis. It was reported that GPP possessed protective effects on exhaustion exercise-induced hepatocyte apoptosis by its anti-oxidative and anti-apoptotic activities. GPP administration decreased the expression of Bax and increased the expression of Bcl-2. Meanwhile, GPP protected liver tissue against oxidative damage induced by exhaustion exercise as evidenced by significant increase in the SOD, GSH-Px, and CAT activities and decrease in the content of MDA [107].
3.6.3. Alcoholic Liver Injury
Studies on alcoholic liver injury indicated that oxidative stress was one of the most important mechanisms, which further led to oxidative damage and inflammatory response. GPS exerted the protective effect on alcoholic liver injury by activating the Nrf2 signaling pathway to improve the level of antioxidant defense system and inhibiting the nuclear transposition of NF-κB to reduce the inflammatory response [108].
3.6.4. High Choline-Induced Liver Injury
The dietary ingestion of high choline was highly linked to liver oxidative stress injury. GPS was reported to exert the significant protection against liver oxidative stress injury in high choline-treated mice [109].
3.6.5. Fatty Liver Disease
GPS was believed to be a promising agent for the prevention of fatty liver disease. The protective effects of GPS on fatty liver disease were investigated in rats treated with high-fat and cholesterol diet and alcohol. GPS could prevent liver fatty degeneration through the up-regulation of PPAR-α expression in the liver to inhibit lipid peroxidation and hepatocyte apoptosis, thus ameliorating hepatic steatosis and mitochondrial damage and improving hepatic function [110]. GP was reported to have a hepatoprotective effect on nonalcoholic fatty liver disease (NAFLD) as well. Studies had shown that GP could alleviate NAFLD with following mechanisms: (1) reduction of inflammatory response by regulating the balance of liver Treg/Th17 cells, reducing the production of pro-inflammatory factors, and increasing the production of anti-inflammatory factors, so as to reduce liver inflammatory response and protect liver tissue; (2) reduction of oxidative stress as evidenced by increased SIRT6 and phase 2 anti-oxidant enzyme expression in liver tissues; (3) structural alterations of gut microbiota [111,112,113].
3.7. Hypoglycemic Effect
Diabetes mellitus is a chronic metabolic disease caused by genetic and environmental factors for a long time, which has become the third category of diseases seriously endangering human health. At present, the existing oral hypoglycemic drugs in China, such as metformin and sulfonylureas, could cause serious adverse effects [114]. The long-term use of insulin will lead to decreased insulin receptor sensitivity of the body, insulin resistance, and finally aggravate the disease. Therefore, it was urgent to seek new, safe, and effective oral hypoglycemic drugs. In recent years, natural medicine has attracted more and more attention because of its long-lasting effect and fewer side effects. Researchers had made great progress in searching for anti-diabetic components from traditional Chinese medicine resources, including GP. At present, the research on anti-diabetes mainly focused on GPS, GPP, and crude extracts (Table 6) [115].
Table 6.
Hypoglycemic effect of GP.
GPS, as the main active ingredient, had been reported for its hypoglycemic effects. Phanoside, a novel dammarane-type saponin with a molecular mass of 914.5 Da, was isolated from ethanol extract, and four stereoisomers differing in configurations at positions 21 and 23 were identified, each of which was found to stimulate insulin release from isolated rat pancreatic islets [122]. Phanoside-induced insulin release might be mediated via the K-ATP channel and L-type Ca2+ channel [123]. However, it could dose-dependently stimulate insulin release at both low and high glucose levels (3.3 mM and 16.7 mM), suggesting that its insulin secretory effect was not glucose-dependent, which could potentially induce severe hypoglycemia and was similar to that of the well-known antidiabetic drug sulfonylurea. Gylongiposide I, which was identified in screening the active components of GP, was different from Phanoside and displayed unique abilities to stimulate insulin release at high glucose levels (16.7 mM) but limited effects at a low glucose concentration (3.3 mM) [124]. In vitro studies had shown [125] that GP exerted stronger inhibitory α-glucosidase activity with IC50 (42.8 µg/mL) as compared with acarbose (IC50 at 53.9 µg/mL). Based on this, GP could be developed as a novel anti-diabetic drug taken to reduce postprandial blood glucose. Compared with the traditional oral hypoglycemic drugs that had no obvious protective effect on the kidney, GPS displayed a protective effect on the kidney while lowering blood glucose a well as significantly improving renal tubule congestion and renal tubule epithelial edema in diabetic mice [116]. AMPK was a key regulator of glucose and lipid metabolism. Activated AMPK decreases blood glucose levels by increasing the glucose uptake and glycogen synthesis, stimulating the translocation of glucose transporter 4 (GluT4) to the cell membrane as well as up-regulating GluT4 expression. Therefore, AMPK activators were considered promising candidates for type-2 diabetes mellitus. Two novel dammarane-type saponins named damulin A and B isolated from GP could stimulate glucose uptake via AMPK activation in L6 myotube cells [117]. Protein tyrosine phosphatase 1B (PTP1B), a negative regulator of the insulin-signaling pathway, could inhibit or terminate the insulin signaling pathway transduction and further regulate negatively insulin sensitivity. Therefore, inhibition of PTP1B was an effective therapeutic approach to treatment type-2 diabetes mellitus. In a recent study conducted by Wang et al. that aimed to find the hypoglycemic active components in GP, 12 compounds and their hydrolysis products were isolated and identified from GPS, and 11 compounds exerted inhibitory activity against PTP1B [126]; these results were in line with previous studies [127]. It was reported that insulin sensitivity could be affected by inflammation. Inflammatory factors such as TNF-α and IL-6 produced by nonspecific inflammation could interfere with the conduction of insulin signaling pathway and thereby lead to decreased insulin sensitivity. Studies showed that insulin sensitivity in diabetic rats treated with GPS was improved by blocking the NF-κB signaling pathway and reducing the secretion of inflammatory factors. However, GPS had no influence on the streptozotocin-induced rat islet cells damage [128].
A large number of studies proved that GPP had hypoglycemic effect. Du et al. [119] established a model of type 2 diabetic rats by a high-fat and high-sugar diet combined with a small dose of streptozotocin to study the effects of GPP on blood glucose and its possible mechanism. The results showed that the fasting blood glucose and glucose tolerance were reduced by the treatment with GPP. Moreover, the fasting serum insulin level was increased in a dose-dependent manner, suggesting that the hypoglycemic effect of GPP was related to the secretion of insulin. In vitro studies showed that GPP had a strong inhibitory effect on α-amylase in a dose-dependent manner, which was equivalent to about 50% of the inhibitory effect of acarbose at the same concentration [118].
The ethanol extract of GP grown in Hainan could significantly reduce the blood glucose level of alloxan-induced hyperglycemia mice. There was no statistically difference between the medium-dose group (2 g/kg) and the metformin group; the high-dose group (4 g/kg) showed a stronger hypoglycemic effect than the metformin group. More importantly, it could significantly inhibit the increase in blood glucose induced by glucose, but it exhibited no obvious effect on the blood glucose of normal mice [120]. Huang et al. [129] estimated the effects of different components of GP on blood biochemical indexes of hyperglycemic mice and found that both GPE and 90% ethanol elution parts via macroporous resin chromatography could reduce the blood glucose of diabetic mice and alleviate the symptoms of “three more and one less”. Yassin et al. [121] explored the effect of GPE (1600 mg/kg/d) on hepatic glucose output (HGO) in spontaneously type 2 diabetic Goto-Kakizaki rats, and the results showed that the hepatic insulin sensitivity was improved, while HGO and plasma glucose levels were reduced. This was in agreement with results from a clinic trial conducted to evaluate the anti-diabetic effect of GP tea in drug-naïve type 2 diabetic patients [130,131]. Moreover, the biosecurity of tea was proved clinically, as no hepatoxicity and nephrotoxicity or other adverse effects such as hypoglycemia were observed in the trial. In addition, the antidiabetic effect of GP was associated with the stimulation of insulin release from the islets. An oral treatment of GP water extract (0.3 g/kg of body weight daily) for two weeks in GK rats improved glucose tolerance and stimulated insulin release. GP-induced insulin release was partly mediated via K-ATP and L-type Ca2+ channels [132].
3.8. Others
In addition to the above significant biological activities, GP also possessed an additional anti-aging effect, anxiolytic effect, anti-gastric ulcer effect, anti-fatigue effect, anti-platelet aggregation activity, antidepressant effect, sedative and hypnotic effect, immunomodulatory effect, antiviral activity, antimicrobial activity, and effects on osteoporosis and hyperuricemia, as Table 7 shown.
Table 7.
Other activities of GP.
4. Conclusions and Perspectives
G. pentaphyllum (Thunb.) Makino is a highly promising TCM that has attracted great attention with a wide range of potent bioactivities, including hypoglycemic, anti-tumor, hepatoprotective, neuroprotective, and so on. As a typical edible and medicinal plant, GP has been developed in the field of animal feed to improve the quality of meat and has a pretty good market prospect. However, GP was widely distributed and rich in natural resources. The chemical composition of GP has not been studied thoroughly. Most research about its pharmacological properties were only limited to GPS. Thereby, further pharmacological research on single gypenoside and flavonoids are required to extend our understanding of pharmacological properties. Another important issue was that the underlying mechanism of bioactivities was still inadequately researched. In the future, the new technologies should be explored to deepen our understanding of GP, which can provide the basis for clinical drug use and preparation development.
Author Contributions
Conceptualization and Funding Acquisition, X.M.; Project Administration and Supervision, L.Y.; Original Draft Preparation and Methodology, C.S. and N.L.; Data Collection, R.R., Y.W., X.S., F.L. and R.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. U20A20136, 81860753, 81560684); Key Research and Development Program of Ningxia (2018BHF2001); Ningxia key research and invention program of science and technology cooperation of the East and the West (No. 2018ZDKJ0219, 2017BY084, 2017BY079).
Conflicts of Interest
Authors have no conflict of interest.
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| ABCA1 | ATP-binding cassette transporter A1 |
| ABCG1 | ATP-binding cassette transporter G1 |
| AD | Alzheimer’s disease |
| ADP | Adenosine diphosphate |
| AIF | Apoptosis-inducing factor |
| Akt | Protein kinase B |
| ALT | Alanine aminotransferase |
| AMPK | AMP-activated protein kinase |
| ARE | Antioxidant responsive element |
| AS | Atherosclerosis |
| AST | Aspartate aminotransferase |
| ATF-4 | Activating transcription factor 4 |
| Atg | Autophagy-related gene |
| Aβ | Beta-amyloid protein |
| BDNF | Brain-derived neurotrophic factor |
| BID | BH3-interacting-domain |
| CAT | Catalase |
| CCl4 | Carbon tetrachloride |
| CDC25A | Cell division cycle protein 25A |
| CDK | Cyclin-dependent kinases |
| Chk2 | Checkpoint kinase 2 |
| CHOP | C/EBP-homologous protein |
| COX-2 | Cyclooxygenase-2 |
| Cry | Crytochrome |
| CTL | Cytotoxic T lymphocytes |
| DFF45 | DNA degradation factor 45 KD |
| DRP-1 | Dynamin-related protein 1 |
| E2F1 | E2F transcription factor 1 |
| ECs | Endothelial cells |
| ER | Endoplasm reticulum |
| ERCC6L | Excision repair cross-complementation group 6 like |
| ERK | Extracellular signal-regulated kinase |
| ERα | Estrogen receptor alpha |
| FAK | Focal adhesion kinase |
| FLIP | FLICE inhibitory protein |
| FOXO1 | Forkhead Box O1 |
| GADD153 | Growth arrest and DNA damage-inducible 153 |
| GluT4 | Glucose transporter 4 |
| GluT9 | Glucose transporter 9 |
| GP | Gynostemma pentaphyllum |
| GPE | Gynostemma pentaphyllum extract |
| GPP | Gynostemma pentaphyllum polysaccharides |
| GPS | Gypenosides |
| GRP78 | Glucose regulated protein 78 KD |
| GSH-Px | Glutathione peroxidase |
| GTP | Guanosine triphosphate |
| H5N1 | Highly pathogenic avian influenza virus |
| HDL | High-density lipoprotein |
| HDL-C | High-density lipoprotein cholesterol |
| HGO | Hepatic glucose output |
| HIF-1α | Hypoxia-inducible factor-1α |
| HO-1 | Heme oxygenase 1 |
| HUVECs | Human umbilical vein endothelial cells |
| I/R | Ischemia–reperfusion |
| ICAM-1 | Intercellular cell adhesion molecule-1 |
| IFN | Interferon |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| IP3R | Inositol triphosphate receptor |
| IκBα | Inhibitor of NF-κB α |
| JNK | c-Jun N-terminal kinase |
| LC3 | Microtubule-associated protein 1 light chain 3 |
| LDL | Low-density lipoprotein |
| LDL-C | Low-density lipoprotein cholesterol |
| LDLR | Low-density lipoprotein receptor |
| L-DOPA | L-3, 4-Dihydroxyphenylalanine |
| LXR | Liver X receptor |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemotactic protein 1 |
| MDA | Malondialdehyde |
| MDCK | Madin-Darby canine kidney |
| MFC | Mouse forestomach carcinoma |
| Mfn2 | Mitofusin 2 |
| MMP-2 | Matrix metalloproteinase-2 |
| MMP-9 | Matrix metalloproteinase-9 |
| MPP+ | 1-Methyl-4-phenylpyridinium ion |
| MPTP | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| mTOR | Mammalian target of rapamycin |
| MyD88 | Myeloid differentiation factor 88 |
| NAFLD | Nonalcoholic fatty liver disease |
| NFAT | Nuclear factor of activated T cells |
| NF-kB | Nuclear factor-kappa B |
| NK | Natural killer |
| NMDA | N-methyl-D-aspartate |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| NOX4 | Nicotinamide adenine dinucleotide phosphate-oxidase 4 |
| Nrf2 | Nuclear factor-erythroid 2-related factor 2 |
| OAT1 | Organic anion transporter 1 |
| OGD/R | Oxygen-glucose deprivation–reoxygenation |
| OPA1 | Optic atrophy protein 1 |
| Oria1 | Calcium release-activated calcium channel protein 1 |
| Ox-LDL | Oxidized low-density lipoprotein |
| PARP | Poly ADP-ribose polymerase |
| PCNA | Proliferating cell nuclear antigen |
| PCREB | Phosphorylated cAMP response element-binding protein |
| PCSK9 | Proprotein convertase subtilisin/kexin 9 |
| PD | Parkinson’s disease |
| PI3K | Phosphoinositide 3-kinases |
| PPAR | Peroxisome proliferator activated receptor |
| PRRSV | Porcine reproductive and respiratory syndrome virus |
| PTP1B | Protein tyrosine phosphatase 1B |
| RANKL | Receptor Activator of Nuclear Factor-kappa B Ligand |
| RCC | Renal cell carcinoma |
| ROS | Reactive oxygen species |
| SERCA | Sarco/endoplasmic-reticulum-Ca2+-ATPase |
| SIRT1 | Silent information regulator 1 |
| SOC | Store-operated Ca2+ channel |
| SOCS1 | Suppressors of cytokine signaling 1 |
| SOD | Superoxide dismutase |
| SOS | Sevenless homolog |
| SRB1 | Scavenger receptor class B type I |
| STAI | State-Trait Anxiety Inventory |
| STIM1 | Stromal interacting molecule 1 |
| T-AOC | Total antioxidant capacity |
| TC | Total cholesterol |
| TCM | Traditional Chinese medicine |
| TG | Triglycerides |
| TGF-β | Transforming growth factor-β |
| TH | Tyrosine hydrolase |
| TLR4 | Toll-like receptor 4 |
| TNF | Tumor necrosis factor |
| TRAIL | TNF-related apoptosis-inducing ligand |
| T-STAI, | Trait Anxiety Scale of the STAI |
| ULK1 | Unc-51-like kinase 1 |
| uPA | Urokinase-type plasminogen activator |
| URAT1 | Urate transporter 1 |
| VD | Vascular dementia |
| XIAP | X-linked inhibitor of apoptosis protein |
| γH2AX | Phosphorylation of histone H2AX |
References
- Yuan, Z.Y.; Xie, M.Z.; Huang, H.Y. Advances in the chemical constituents and pharmacological studies of gynostemma pentaphyllum. Asia-Pac. Tradit. Med. 2019, 15, 190–197. [Google Scholar]
- Razmovski-Naumovski, V.; Huang, T.H.; Tran, V.H.; Li, G.Q.; Duke, C.C.; Roufogalis, B.D. Chemistry and Pharmacology of Gynostemma pentaphyllum. Phytochem. Rev. 2005, 4, 197–219. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.H.; Farimani, M.M.; Yang, J.L. Dammarane-type saponins from Gynostemma pentaphyllum and their potential anti-AD activity. Phytochem. Lett. 2019, 31, 147–154. [Google Scholar] [CrossRef]
- Chiranthanut, N.; Teekachunhatean, S.; Panthong, A.; Khonsung, P.; Kanjanapothi, D.; Lertprasertsuk, N. Toxicity evaluation of standardized extract of Gynostemma pentaphyllum Makino. J. Ethnopharmacol. 2013, 149, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yuan, D.S. Advances in Studies on Gynostemma Germplasm Resources in China. J. Yunnan Agric. Univ. 2009, 24, 459–461. [Google Scholar]
- Fan, D.D.; Kuang, Y.H.; Xiang, S.X.; Zhang, D.; Zhu, J.J.; Wang, Z.M.; Wang, D.Q.; Li, C.Y. Research progress in chemical constituents and pharmacological activities of gynostemma pentaphyllum. Chin. Pharm. J. 2017, 52, 342–352. [Google Scholar]
- Zhou, W.C. Research on extraction and anti-inflammatory activity of new saponins from Gynostemma pentaphyllum seeds. Yunnan Chem. Technol. 2015, 45, 34–37. [Google Scholar]
- Lou, Y.Y.; Mu, L.; Huang, Y.P.; Zhang, J.; Yin, Z.Q.; Pan, K. A new cucurbitane-type saponin from Gynostemma pentaphyllum. Chin. Tradit. Herb. Drugs 2021, 52, 1872–1876. [Google Scholar]
- Ji, X.L.; Shen, Y.B.; Guo, X.D. Isolation, Structures, and Bioactivities of the Polysaccharides from Gynostemma pentaphyllum (Thunb.) Makino: A Review. Biomed Res. Int. 2018, 2018, 1–14. [Google Scholar]
- Xue, J.J.; Gao, J.D.; Chen, Z.J.; Jing, M.; Qiao, J.; Liu, X. Chemical composition and pharmacological action of Gynostemma pentaphyllum. J. Gansu Univ. Chin. Med. 2018, 35, 86–89. [Google Scholar]
- Ma, L.P.; Zhao, P.R.; Zhang, H.F.; Lou, Z.L. Determination of polysaccharide content in different parts of Gynostemma pentaphyllum. J. Henan Med Univ. 2000, 35, 445–446. [Google Scholar]
- Ling, Y.; Lei, Z.N.; Li, X.Y.; Yan, X.Y.; Wang, S.Y.; Yang, W.L.; Nie, Q.H.; Zhang, Q.; Bao, H.O.; Yu, J.M.; et al. Characterization and identification of the chemical constituents of gynostemma pentaphyllum using high performance liquid Chromatography-Electrospray Ionization-Quadrupole Time-of-Flight tandem mass spectrometry (HPLC-ESI-QTOF-MS/MS). Anal. Lett. 2020, 53, 760–773. [Google Scholar] [CrossRef]
- Shen, Z.L.; Wang, Z.B.; Hou, H.F.; Zhang, L.; Huang, J.J. New progress in research on chemical constituents, pharmacological effects and application of Gynostemma pentaphyllum. Ginseng Res. 2020, 32, 59–64. [Google Scholar]
- Zu, M.L.; Piao, X.L.; Gao, J.M.; Xing, S.F.; Liu, L.H. Monomer gypenoside LI from Gynostemma pentaphyllum inhibits cell proliferation and upregulates expression of miR-128-3p in melanoma cells. J. Biochem. Mol. Toxic. 2020, 34, 1–8. [Google Scholar] [CrossRef]
- Zu, M.L.; Duan, Y.; Xie, J.B.; Qi, Y.S.; Xie, P.; Borjigidai, A.; Piao, X.L. Gypenoside LI arrests the cell cycle of breast cancer in G0/G1 phase by down-regulating E2F1. J. Ethnopharmacol. 2021, 273, 114017. [Google Scholar] [CrossRef]
- Ma, J.X.; Hu, X.P.; Liao, C.H.; Xiao, H.T.; Zhu, Q.C.; Li, Y.; Liu, Z.G.; Tao, A.J.; He, Z.D.; Xu, C.S.; et al. Gypenoside l inhibits proliferation of liver and esophageal cancer cells by inducing senescence. Molecules 2019, 24, 1054. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.H.; Zheng, K.; Li, Y.; Xu, H.; Kang, Q.R.; Fan, L.; Hu, X.P.; Jin, Z.; Zeng, Y.; Kong, X.L.; et al. Gypenoside L inhibits autophagic flux and induces cell death in human esophageal cancer cells through endoplasm reticulum stress-mediated Ca2+ release. Oncotarget 2016, 7, 47387–47402. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; Liao, C.H.; Li, Y.; Fan, X.M.; Fan, L.; Xu, H.; Kang, Q.R.; Zeng, Y.; Wu, X.L.; Wu, H.Q.; et al. Gypenoside L, Isolated from Gynostemma pentaphyllum, Induces Cytoplasmic Vacuolation Death in Hepatocellular Carcinoma Cells through Reactive-Oxygen-Species-Mediated Unfolded Protein Response. J. Agric. Food Chem. 2016, 64, 1702–1711. [Google Scholar] [CrossRef]
- Xing, S.F.; Liu, L.H.; Zu, M.L.; Ding, X.F.; Cui, W.Y.; Chang, T.; Piao, X.L. The inhibitory effect of gypenoside stereoisomers, gypenoside L and gypenoside LI, isolated from Gynostemma pentaphyllum on the growth of human lung cancer A549 cells. J. Ethnopharmacol. 2018, 219, 161–172. [Google Scholar] [CrossRef]
- Lu, H.F.; Chen, Y.S.; Yang, J.S.; Chen, J.C.; Lu, K.W.; Chiu, T.H.; Liu, K.C.; Yeh, C.C.; Chen, G.W.; Lin, H.J.; et al. Gypenosides induced G0/G1 arrest via inhibition of cyclin E and induction of apoptosis via activation of caspases-3 and -9 in human lung cancer A-549 cells. In Vivo 2008, 22, 215–221. [Google Scholar]
- Chen, J.C.; Lu, K.W.; Lee, J.H.; Yeh, C.C.; Chung, J.G. Gypenosides induced apoptosis in human colon cancer cells through the mitochondria-dependent pathways and activation of caspase-3. Anticancer Res. 2006, 26, 4313–4326. [Google Scholar]
- Chen, J.C.; Lu, K.W.; Tsai, M.L.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Hsia, T.C.; Yu, C.S.; Chou, S.T.; Kao, M.C.; et al. Gypenosides induced G0/G1 arrest via CHk2 and apoptosis through endoplasmic reticulum stress and mitochondria-dependent pathways in human tongue cancer SCC-4 cells. Oral Oncol. 2009, 45, 273–283. [Google Scholar] [CrossRef]
- Liu, J.S.; Chiang, T.H.; Wang, J.S.; Lin, L.J.; Chao, W.C.; Inbaraj, B.S.; Lu, J.F.; Chen, B.H. Induction of p53-independent growth inhibition in lung carcinoma cell A549 by gypenosides. J. Cell. Mol. Med. 2015, 19, 1697–1709. [Google Scholar] [CrossRef]
- Lu, K.W.; Chen, J.C.; Lai, T.Y.; Yang, J.S.; Weng, S.W.; Ma, Y.S.; Lu, P.J.; Weng, J.R.; Chueh, F.S.; Wood, W.G.; et al. Gypenosides inhibits migration and invasion of human oral cancer SAS cells through the inhibition of matrix metalloproteinase-2 -9 and urokinase-plasminogen by ERK1/2 and NF-kappa B signaling pathways. Hum. Exp. Toxicol. 2011, 30, 406–415. [Google Scholar] [CrossRef]
- Lu, K.W.; Chen, J.C.; Lai, T.Y.; Yang, J.S.; Weng, S.W.; Ma, Y.S.; Lin, H.Y.; Wu, R.S.; Wu, K.C.; Wood, W.G.; et al. Gypenosides suppress growth of human oral cancer SAS cells in vitro and in a murine xenograft model. Integr. Cancer Ther. 2012, 11, 129–140. [Google Scholar] [CrossRef]
- Lu, K.W.; Ma, Y.S.; Yu, F.S.; Huang, Y.P.; Chu, Y.L.; Wu, R.S.; Liao, C.L.; Chueh, F.S.; Chung, J.G. Gypenosides induce cell death and alter gene expression in human oral cancer HSC-3 cells. Exp. Ther. Med. 2017, 14, 2469–2476. [Google Scholar] [CrossRef]
- Shi, L.; Pi, Y.Z.; Luo, C.; Zhang, C.H.; Tan, D.H.; Meng, X.J. In vitro inhibitory activities of six gypenosides on human liver cancer cell line HepG2 and possible role of HIF-1α pathway in them. Chem.-Biol. Interact. 2015, 238, 48–54. [Google Scholar] [CrossRef]
- Sun, D.P.; Li, X.X.; Liu, X.L.; Zhao, D.; Qiu, F.Q.; Li, Y.; Ma, P. Gypenosides induce apoptosis by ca2+ overload mediated by Endoplasmic-Reticulum and Store-Operated ca2+ channels in human hepatoma cells. Cancer Biother. Radio. 2013, 28, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Wang, X.; Wang, Y.; Wang, P.; Xiao, Y. Antiproliferation and anti-migration induced by gypenosides in human colon cancer SW620 and esophageal cancer Eca-109 cells. Hum. Exp. Toxicol. 2014, 33, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Wang, X.B.; Niu, J.F.; Wang, Y.Q.; Wang, P.; Liu, Q.H. Anti-cancer effect and the underlying mechanisms of gypenosides on human colorectal cancer SW-480 cells. PLoS ONE 2014, 9, e95609. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C.; Lu, J.F.; Wang, J.S.; Lin, L.J.; Kuo, H.I.; Chen, B.H. Antiproliferation Effect and Apoptosis Mechanism of Prostate Cancer Cell PC-3 by Flavonoids and Saponins Prepared from Gynostemma pentaphyllum. J. Agric. Food Chem. 2011, 59, 11319–11329. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, L.H.; Ren, Y.G.; Gao, Y.L.; Kang, L.; Qiao, Q. Anticancer and immunoregulatory activity of Gynostemma pentaphyllum polysaccharides in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2014, 69, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Wang, Z.H.; Zhao, Y.X.; Luo, S.J.; Zhang, D.W.; Xiao, S.X.; Peng, Z.H. Isolation and antitumor activities of acidic polysaccharide from Gynostemma pentaphyllum Makino. Carbohyd. Polym. 2012, 89, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Huang, J.J.; Lin, W.J.; Yuan, Z.W.; Feng, S.L.; Xie, Y.; Ma, W.Z. In Vitro Anticancer Activity of a Nonpolar Fraction from Gynostemma pentaphyllum (Thunb.) Makino. Evid.-Based Compl. Alt. 2016, 2016, 1–11. [Google Scholar]
- Xing, S.F.; Liu, L.H.; Zu, M.L.; Lin, M.; Zhai, X.F.; Piao, X.L. Inhibitory Effect of Damulin B from Gynostemma pentaphyllum on Human Lung Cancer Cells. Planta Med. 2019, 85, 394–405. [Google Scholar] [CrossRef]
- Zhang, X.S.; Shi, G.H.; Wu, X.J.; Zhao, Y. Gypensapogenin H from hydrolyzate of total Gynostemma pentaphyllum saponins induces apoptosis in human breast carcinoma cells. Nat. Prod. Res. 2020, 34, 1642–1646. [Google Scholar] [CrossRef]
- Zhang, X.S.; Zhao, C.; Tang, W.Z.; Wu, X.J.; Zhao, Y.Q. Gypensapogenin H, a novel dammarane-type triterpene induces cell cycle arrest and apoptosis on prostate cancer cells. Steroids 2015, 104, 276–283. [Google Scholar] [CrossRef]
- Liu, Y.J.; Liu, J.C.; Wang, Y.F. Effects of Gynostemma pentaphyllum polysaccharides on tumor growth inhibition and immune regulation in MFC gastric cancer-bearing mice. Chin. Tradit. Pat. Med. 2019, 41, 2876–2881. [Google Scholar]
- Liu, X.; Wang, P.J.; Xu, F.X. Study on Gypenosides inhibiting neoplasm growth and elevating immunological function in Lewis lung cancer of mice. J. Anhui Tradit. Chin. Med Coll. 2001, 20, 43–44. [Google Scholar]
- Tan, X.S.; Li, K.N.; Miao, Y.J. Gypenosides inhibits immune escape factors of non-small lung cancer cells by facilitating sTim-3/Tim-3 ratio. Immunol. J. 2021, 37, 520–527. [Google Scholar]
- Liu, H.; Li, X.M.; Duan, Y.; Xie, J.B.; Piao, X.L. Mechanism of gypenosides of Gynostemma pentaphyllum inducing apoptosis of renal cell carcinoma by PI3K/AKT/mTOR pathway. J. Ethnopharmacol. 2021, 271, 113907. [Google Scholar] [CrossRef]
- Khan, I.; Huang, G.X.; Li, X.A.; Liao, W.L.; Leong, W.K.; Xia, W.R.; Bian, X.Q.; Wu, J.L.; Hsiao, W.L.W. Mushroom polysaccharides and jiaogulan saponins exert cancer preventive effects by shaping the gut microbiota and microenvironment in Apc mice. Pharmacol. Res. 2019, 148, 104448. [Google Scholar] [CrossRef]
- Tsui, K.C.; Chiang, T.H.; Wang, J.S.; Lin, L.J.; Chao, W.C.; Chen, B.H.; Lu, J.F. Flavonoids from Gynostemma pentaphyllum Exhibit Differential Induction of Cell Cycle Arrest in H460 and A549 Cancer Cells. Molecules 2014, 19, 17663–17681. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Tan, D.H.; Yan, T.C.; Jiang, D.H.; Hou, M.X. Cytotoxic triterpenes from the acid hydrolyzate of Gynostemma pentaphyllum saponins. J. Asian Nat. Prod. Res. 2018, 20, 182–187. [Google Scholar] [CrossRef]
- Cui, W.Y.; Jin, Y.L.; Liu, H.; Zu, M.L.; Zhai, X.F.; Yang, C.; Gu, Y.L.; Cheng, Y.; Piao, X.L. Dammarane-type saponins from Gynostemma pentaphyllum and their cytotoxicities. Nat. Prod. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Shi, L.; Meng, X.J.; Cao, J.Q.; Zhao, Y.Q. A new triterpenoid saponin from Gynostemma pentaphyllum. Nat. Prod. Res. Former. Nat. Prod. Lett. 2012, 26, 1419–1422. [Google Scholar] [CrossRef]
- Yin, F.; Zhang, Y.N.; Yang, Z.Y.; Hu, L.H. Nine new dammarane saponins from Gynostemma pentaphyllum. Chem. Biodivers. 2006, 3, 771–782. [Google Scholar] [CrossRef]
- Shi, L.; Cao, J.Q.; Shi, S.M.; Zhao, Y.Q. Triterpenoid saponins fromGynostemma pentaphyllum. J. Asian Nat. Prod. Res. 2011, 13, 168–177. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, H.; Luo, Y.; Zhang, J.; Wang, M.; Liao, P.; Cao, L.; Guo, P.; Sun, G.; Sun, X. Gypenoside XVII prevents atherosclerosis by attenuating endothelial apoptosis and oxidative stress: Insight into the ERα-Mediated PI3K/Akt pathway. Int. J. Mol. Sci. 2017, 18, 77. [Google Scholar] [CrossRef] [Green Version]
- Shou, D.F.; Lu, D.Z.; Wang, P. The mechanism of gypenosides regulate cholesterol homeostasis in foam cells. J. Zhejiang Chin. Med. Univ. 2011, 35, 297–300. [Google Scholar]
- Ge, Q.Q.; Wang, X.F.; Zong, L.; Bu, J.S.; Lu, D.Z. Effects of gypenosides on PPAR-γ/LXR-α/ABCA1 signaling pathway in vascular of ApoE-/- as mice. Zhejiang Med. Educ. 2019, 18, 47–50. [Google Scholar]
- Shen, C.Y.; Jiang, J.G.; Huang, C.L.; Zhu, W. Gypenoside LVI attenuates foam cell formation by promoting cholesterol export and inhibiting inflammation response. J. Funct. Foods 2018, 50, 71–77. [Google Scholar] [CrossRef]
- Song, N.; Yang, F.; Cao, H.M.; Zhang, N.; Jia, L.Q.; Chen, W.N.; Yang, G.L. Gypenoside influences the progression of atherosclerosis in the ApoE-/- mouse through mTOR/ULK1 pathway. Chin J Arter. 2018, 26, 127–132. [Google Scholar]
- Quan, Y.; Qian, M.Z. Effect and Mechanism of Gypenosides on the Inflammatory Molecular Expression in High fat Induced Atherosclerosis Rat. Chin. J. Integr. Tradit. West. 2010, 30, 403–406. [Google Scholar]
- Song, N.; Jia, L.Q.; Cao, H.M.; Ma, Y.X.; Chen, N.; Chen, S.; Lv, X.M.; Yang, G.L. Gypenoside inhibits endothelial cell apoptosis in atherosclerosis by modulating mitochondria through PI3K/Akt/Bad pathway. Biomed Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Quan, Y.; Yang, Y.; Wang, H.; Shu, B.; Gong, Q.H.; Qian, M. Gypenosides attenuate cholesterol-induced DNA damage by inhibiting the production of reactive oxygen species in human umbilical vein endothelial cells. Mol. Med. Rep. 2015, 11, 2845–2851. [Google Scholar] [CrossRef]
- Bo, H.; Hou, X.W.; Liu, R.H.; Liu, X.H.; Hu, Z.H. Gypenoside inhibits ox-LDL uptake and foam cell formation through enhancing Sirt1-FOXO1 mediated autophagy flux restoration. Life Sci. 2018, 264, 118721. [Google Scholar]
- Song, N.; Chen, N.; Cao, H.M.; Chen, S.; Wang, Y.; Qu, N.N.; Jia, L.Q.; Wang, L. Effects of gynostemma pentaphyllum saponins on atherosclerosis in ApoE-/- mice by regulating biological rhythm related proteins. Chin. Arch. Tradit. Chin. Med. 2021, 39, 248–251. [Google Scholar]
- Sun, X.N.; Song, N.; Yang, X.; Chen, Y.R.; Chen, N.; Cao, H.M. Mechanism of gynostemma saponin a improving ox-LDL-induced EA.hy926 cell injury by regulating mitochondrial function. Chin. Arch. Tradit. Chin. Med. 2020, 38, 77–80. [Google Scholar]
- Zhang, N.; Song, N.; Cao, H.M.; Jia, L.Q.; Chen, W.N.; Leng, X. Effect of gypenosides on the prevention and treatment of atherosclerosis by autophagosome. Nat. Prod. Res. Dev. 2017, 29, 2112–2116. [Google Scholar]
- Yang, J.X.; Ma, F.H.; Zhang, J.T.; Shou, Z.X.; Tang, N.; Tang, C.; Xie, Z.X.; Sun, J.F.; Wu, N. Gypenosides suppress the inflammatory response to atherosclerosis in rats and interfere the expression of TLR4/MyD88/NF-κB signaling pathway. Genom. Appl. Biol. 2021. [Google Scholar]
- Wu, D.; Jiao, X.; Yuan, Z.L.; He, Y.Q. Gypenosides attenuated LPS-induced endothelial cell inflammatory injury. Chin. J. New Drugs 2020, 29, 323–328. [Google Scholar]
- Huang, T.H.; Razmovski-Naumovski, V.; Salam, N.K.; Duke, R.K.; Tran, V.H.; Duke, C.C.; Roufogalis, B.D. A novel LXR-α activator identified from the natural product Gynostemma pentaphyllum. Biochem. Pharmacol. 2005, 70, 1298–1308. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Gypenosides Regulates Lipid and Carbohydrate Metabolism via Bile Acids Related Pathway. Master’s Thesis, Zunyi Medical University, Zunyi, China, 2017. [Google Scholar]
- Mao, Y. The Effect of Gypenosides Relieving Fat Accumulation in HepG2 Cells. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2017. [Google Scholar]
- Zhang, Y.D.; Jiang, X.Y.; Cao, L.J.; Wang, J.; Jiang, C.H.; Zhao, M.G.; Zhang, J.; Yin, Z.Q. Gypenoside granules improved lipid metabolism in C57BL/6J mice with hyperlipidemia. J. China Pharm. Univ. 2019, 50, 713–720. [Google Scholar]
- Wu, L.S.; Qian, M.Z. Effects of gypenosides on PCSK9 gene expression and blood lipids lowered by simvastatin. Chin. J. Pathophysiol. 2017, 33, 79–85. [Google Scholar]
- Wang, Y.S.; Wang, J.; Wang, X.N.; Jiang, C.H.; Zheng, X.; Zhang, J.; Yin, Z.Q. Therapeutic effects of gypenosides on hypercholesterolemia and it protective effect on liver injury. J. China Pharm. Univ. 2021, 52, 84–91. [Google Scholar]
- Yin, M.J.; Zhang, J.J.; Wang, L.Z.; Li, F.Y.; Li, Z.F.; Xiang, W.; Bie, S.T.; Wang, C.H.; Li, Z. Ten New Dammarane-Type Saponins with Hypolipidemia Activity from a Functional Herbal Tea-Gynostemma pentaphyllum. Molecules 2020, 25, 3737. [Google Scholar] [CrossRef]
- Lee, H.S.; Lim, S.; Jung, J.I.; Kim, S.M.; Lee, J.K.; Kim, Y.H.; Cha, K.M.; Oh, T.K.; Moon, J.M.; Kim, T.Y.; et al. Gynostemma pentaphyllum extract ameliorates High-Fat Diet-Induced obesity in C57BL/6N mice by upregulating SIRT1. Nutrients 2019, 11, 2475. [Google Scholar] [CrossRef] [Green Version]
- Gauhar, R.; Hwang, S.; Jeong, S.; Kim, J.; Song, H.; Park, D.C.; Song, K.; Kim, T.Y.; Oh, W.K.; Huh, T. Heat-processed Gynostemma pentaphyllum extract improves obesity in ob/ob mice by activating AMP-activated protein kinase. Biotechnol. Lett. 2012, 34, 1607–1616. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.F.; Yang, P.Y.; Wan, J.C.; Chang, Q.M.; Wang, T.T.Y.; Lu, W.Y.; Zhang, Y.Q.; Wang, Q.; Yu, L.L. Gypenosides reduced the risk of overweight and insulin resistance in C57BL/6J mice through modulating adipose thermogenesis and gut microbiota. J. Agric. Food Chem. 2017, 65, 9237–9246. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, P.Y.; Shi, H.M.; Sun, X.J.; Lee, S.; Yu, L.L. A novel Gynostemma pentaphyllum saponin and its adipogenesis inhibitory effect through modulating Wnt/β-catenin pathway and cell cycle in mitotic clonal expansion. J. Funct. Foods 2015, 17, 552–562. [Google Scholar] [CrossRef]
- Rao, A.; Clayton, P.; Briskey, D. The effect of an orally-dosed Gynostemma pentaphyllum extract (ActivAMP®) on body composition in overweight, adult men and women: A double-blind, randomised, placebo-controlled study. J. Hum. Nutr. Diet. 2021. [Google Scholar] [CrossRef]
- Wang, J.Z.; Hu, G.; Zai, W.H. Degrading blood-fat effect of saponin from gynostemma pentaphyllum in mice with hyperlipidemia. Chin. J. Ethnomedicine Ethnopharmacy 2007, 41–43. [Google Scholar]
- Song, B.Y. Research progress in the pathogenesis of alzheimer’s disease. J. Xi’an Univ. 2020, 23, 77–79. [Google Scholar]
- Yao, B.C.; Ao, Y.; Zhang, G.; Liu, M.X.; Yao, S.Y.; Li, D.M. Effect and mechanism of Gynostemma pentaphyllum Makino on learning and memory ability in dementia mice. Chin J Public Health 2017, 33, 1708–1711. [Google Scholar]
- Cai, H.; Liang, Q.L.; Ge, G.Q. Gypenoside Attenuates β Amyloid-Induced Inflammation in N9 Microglial Cells via SOCS1 Signaling. Neural Plast. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Shim, I.; Lee, H. Gypenosides attenuate Lipopolysaccharide-Induced neuroinflammation and memory impairment in rats. Evid.-Based Compl. Alt. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.L.; Sun, T.M.; Wang, M.; Li, R.H.; Yao, B.C.; Tan, H.B. The influence of steven leafs on the cell’s model of AD inducted by aβ1-40. Chin. J. Integr. Med. Cardio-Cerebrovasc. Dis. 2012, 10, 201–204. [Google Scholar]
- Jia, D.; Rao, C.G.; Xue, S.X.; Lei, J.L. Purification, characterization and neuroprotective effects of a polysaccharide from Gynostemma pentaphyllum. Carbohyd. Polym. 2015, 122, 93–100. [Google Scholar] [CrossRef]
- Meng, X.B.; Luo, Y.; Liang, T.; Wang, M.X.; Zhao, J.Y.; Sun, G.B.; Sua, X.B. Gypenoside XVII enhances lysosome biogenesis and autophagy flux and accelerates autophagic. J. Alzheimer’s Dis. 2016, 1135–1150. [Google Scholar] [CrossRef]
- Meng, X.B.; Wang, M.; Sun, G.B.; Ye, J.X.; Zhou, Y.H.; Dong, X.; Wang, T.T.; Lu, S.; Sun, X.B. Attenuation of Aβ25–35-induced parallel autophagic and apoptotic cell death by gypenoside XVII through the estrogen receptor-dependent activation of Nrf2/ARE pathways. Toxicol. Appl. Pharm. 2014, 279, 63–75. [Google Scholar] [CrossRef]
- Hong, S.; Yang, J.; Joh, E.; Kim, H.J.; Kim, D. Gypenoside TN-2 ameliorates scopolamine-induced learning deficit in mice. J. Ethnopharmacol. 2011, 134, 1010–1013. [Google Scholar] [CrossRef]
- Joh, E.; Yang, J.; Kim, D. Gypenoside LXXIV ameliorates Scopolamine-Induced learning deficit in mice. Planta Med. 2010, 76, 793–795. [Google Scholar] [CrossRef]
- Yang, X.; Tang, X.; Su, Y.; Cai, Z.; Zeng, D.; Xu, L. Protective effect and mechanism of gypenosides on hippocampus neurons in rats with chronic cerebral ischemia. Chin J. Mod. Appl. Pharm. 2019, 36, 1487–1491. [Google Scholar]
- Qi, M.F.; Luo, C.L.; Qin, W.; Tian, H.; Chen, Y.S. Gynostemma improves vascular dementia-induced cognitive dysfunction in mice. Acta Acad. Med. Zunyi 2012, 35, 371–374. [Google Scholar]
- Luo, C.L.; Tian, H.; Qin, W.; Chen, Y.S. Effect of Gypenosides on the neurons injury and expression of phosphorylated CREB in the hippocampus tissue of mouse with vascular dementia. J. Zunyi Med. Univ. 2014, 37, 320–323. [Google Scholar]
- Zhang, G.L.; Deng, J.P.; Wang, B.H.; Zhao, Z.W.; Li, J.; Gao, L.; Liu, B.L.; Xong, J.R.; Guo, X.D.; Yan, Z.Q.; et al. Gypenosides improve cognitive impairment induced by chronic cerebral hypoperfusion in rats by suppressing oxidative stress and astrocytic activation. Behav. Pharmacol. 2011, 22, 633–644. [Google Scholar] [CrossRef]
- Wang, P.; Niu, L.; Gao, L.; Li, W.X.; Jia, D.; Wang, X.L.; Gao, G.D. Neuroprotective effect of gypenosides against oxidative injury in the substantia nigra of a mouse model of parkinson’s disease. J. Int. Med. Res. 2010, 38, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Niu, L.; Guo, X.D.; Gao, L.; Li, W.X.; Jia, D.; Wang, X.L.; Ma, L.T.; Gao, G.D. Gypenosides protects dopaminergic neurons in primary culture against MPP+-induced oxidative injury. Brain Res. Bull. 2010, 83, 266–271. [Google Scholar] [CrossRef]
- Shin, K.S.; Zhao, T.T.; Park, K.H.; Park, H.J.; Hwang, B.Y.; Lee, C.K.; Lee, M.K. Gypenosides attenuate the development of L-DOPA-induced dyskinesia in 6-hydroxydopamine-lesioned rat model of Parkinson’s disease. BMC Neurosci. 2015, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.T.; Kim, K.S.; Shin, K.S.; Park, H.J.; Kim, H.J.; Lee, K.E.; Lee, M.K. Gypenosides ameliorate memory deficits in MPTP-lesioned mouse model of Parkinson’s disease treated with L-DOPA. BMC Complem. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.S.; Zhao, T.T.; Choi, H.S.; Hwang, B.Y.; Lee, C.K.; Lee, M.K. Effects of gypenosides on anxiety disorders in MPTP-lesioned mouse model of Parkinson’s disease. Brain Res. 2014, 1567, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Park, M.S.; Kim, S.H.; Hwang, B.Y.; Lee, C.K.; Lee, M.K. Neuroprotective Effects of Herbal Ethanol Extracts from Gynostemma pentaphyllum in the 6-Hydroxydopamine-Lesioned Rat Model of Parkinson’s Disease. Molecules 2010, 15, 2814–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, F.Y.; Chen, Z.F.; Wu, H.M.; Guo, P.X. Research Progress on the Pathogenesis of Parkinson’s Disease. Chin. J. Ethnomed. Ethnopharmacy 2020, 29, 60–64. [Google Scholar]
- Yu, H.J.; Shi, L.Y.; Qi, G.X.; Zhao, S.J.; Gao, Y.; Li, Y.Z. Gypenoside protects cardiomyocytes against Ischemia-Reperfusion injury via the inhibition of Mitogen-Activated protein kinase mediated nuclear factor kappa b pathway in vitro and in vivo. Front. Pharmacol. 2016, 7, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.J.; Guan, Q.G.; Guo, L.; Zhang, H.S.; Pang, X.F.; Cheng, Y.; Zhang, X.G.; Sun, Y.X. Gypenosides alleviate myocardial ischemia-reperfusion injury via attenuation of oxidative stress and preservation of mitochondrial function in rat heart. Cell Stress Chaperones 2016, 21, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.P.; Shi, R.; Wang, X.J.; Bao, Y. Gypenoside a protects ischemia/reperfusion injuries by suppressing miR-143-3p level via the activation of AMPK/Foxo1 pathway. Biofactors 2020, 46, 432–440. [Google Scholar] [CrossRef]
- Yu, H.J.; Zhang, H.S.; Zhao, W.H.; Guo, L.; Li, X.Y.; Li, Y.; Zhang, X.G.; Sun, Y.X. Gypenoside protects against myocardial Ischemia-Reperfusion injury by inhibiting cardiomyocytes apoptosis via inhibition of CHOP pathway and activation of PI3K/Akt pathway in vivo and in vitro. Cell. Physiol. Biochem. 2016, 39, 123–136. [Google Scholar] [CrossRef]
- Ye, Q.F.; Zhu, Y.; Ye, S.J.; Liu, H.; She, X.G.; Niu, Y.; Ming, Y.Z. Gypenoside attenuates renal ischemia/reperfusion injury in mice by inhibition of ERK signaling. Exp. Ther. Med. 2016, 11, 1499–1505. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ming, Y.Z.; Wan, Q.Q.; Ye, S.J.; Xie, S.; Zhu, Y.; Wang, Y.F.; Zhong, Z.B.; Li, L.; Ye, Q.F. Gypenoside attenuates hepatic ischemia/reperfusion injury in mice via anti-oxidative and anti-apoptotic bioactivities. Exp. Ther. Med. 2014, 7, 1388–1392. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.M. Protective Effect of Gynostemma pentaphyllum Polysaccharide on Liver Injure Induced by Carbon Tetrachloride in Rats. Chin. J. Exp. Tradit. Med. Formulae 2013, 19, 244–247. [Google Scholar]
- Chen, J.; Tsai, C.; Chen, L.; Chen, H.; Wang, W. Therapeutic effect of gypenoside on chronic liver injury and fibrosis induced by CCl4 in rats. Am. J. Chin. Med. 2000, 28, 175–185. [Google Scholar] [CrossRef]
- Chen, J.X.; Zhang, J.G.; Zhang, L.; Liu, J.B. Study on Liver-protective Effect of Total Saponins of Gynostemma Pentaphyllum. China Pharm. 2007, 16, 7–8. [Google Scholar]
- Chen, J.M.; Li, X.W.; Hu, Y.H.; Liu, W.; Zhou, Q.; Zhang, H.; Mu, Y.P.; Liu, P. Gypenosides ameliorate carbon Tetrachloride-Induced liver fibrosis by inhibiting the differentiation of hepatic progenitor cells into myofibroblasts. Am. J. Chin. Med. 2017, 45, 1061–1074. [Google Scholar] [CrossRef]
- Xu, Y.F.; Qi, B. Protective Effects of Polysaccharides from Gynostemma pentaphyllum Makino on the Exhaustive Exercise-induced Hepatocyte Apoptosis. Food Res. Dev. 2016, 37, 158–162. [Google Scholar]
- Nan, Y.; Zhang, W.; Chang, J.R.; Cao, J.; Zhu, Y.X. Gypenosides protect acute alcoholic liver injury through Nrf2 /NF-κB signaling pathway in mice. Chin. Pharmacol. Bull. 2019, 35, 40–45. [Google Scholar]
- Yang, C.C.; Zhao, Y.; Ren, D.Y.; Yang, X.B. Protective Effect of Saponins-Enriched Fraction of Gynostemma pentaphyllum against High Choline-Induced Vascular Endothelial Dysfunction and Hepatic Damage in Mice. Biol. Pharm. Bull. 2020, 43, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Qin, R.N.; Zhang, J.Y.; Li, C.Y.; Zhang, X.Q.; Xiong, A.H.; Huang, F.; Yin, Z.; Li, K.Y.; Qin, W.Y.; Chen, M.Z.; et al. Protective effects of gypenosides against fatty liver disease induced by high fat and cholesterol diet and alcohol in rats. Arch. Pharm. Res. 2012, 35, 1241–1250. [Google Scholar] [CrossRef]
- Bae, U.J.; Park, E.O.; Park, J.; Jung, S.J.; Ham, H.; Yu, K.W.; Park, Y.J.; Chae, S.W.; Park, B.H. Gypenoside UL4-Rich Gynostemma pentaphyllum Extract Exerts a Hepatoprotective Effect on Diet-Induced Nonalcoholic Fatty Liver Disease. Am. J. Chin. Med. 2018, 46, 1315–1332. [Google Scholar] [CrossRef]
- Shen, S.H.; Zhong, T.Y.; Peng, C.; Fang, J.; Lv, B. Structural modulation of gut microbiota during alleviation of non-alcoholic fatty liver disease with Gynostemma pentaphyllum in rats. BMC Complement. Med. Ther. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Ruan, D.; Lei, J. Beneficial effect of stevenleaf on the imbalance between th17 and treg in nonalcoholic fatty liver disease model rats. Chin J. Mod. Appl. Pharm. 2017, 34, 1683–1688. [Google Scholar]
- Stein, S.A.; Lamos, E.M.; Davis, S.N. A review of the efficacy and safety of oral antidiabetic drugs. Expert Opin. Drug Saf. 2013, 12, 153–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.N.; Shou, Q.Y.; Tian, Y.S.; Gu, G.Z.; Hu, Q.J.; Yang, H.X.; Chen, X.P.; Wang, X.C. Research progress on bioactive component groups and mechanisms of Gynostemma pentaphyllum for treatment of diabetes. Chin. Tradit. Herb. Drugs 2020, 51, 2248–2257. [Google Scholar]
- Lin, Z.Z.; Chen, T. Study on hypoglycemic effect of gypenosides on experimental diabetic mice. J. Longyan Univ. 2011, 29, 51–53. [Google Scholar]
- Nguyen, P.H.; Gauhar, R.; Hwang, S.L.; Dao, T.T.; Park, D.C.; Kim, J.E.; Song, H.; Huh, T.L.; Oh, W.K. New dammarane-type glucosides as potential activators of AMP-activated protein kinase (AMPK) from Gynostemma pentaphyllum. Bioorgan. Med. Chem. 2011, 19, 6254–6260. [Google Scholar] [CrossRef]
- Wei, S.R.; Xue, C.K.; He, X.B.; Shen, K.; Yuan, B.; Jiang, P.; Zhu, J.; Li, Y.; Zeng, L. Experimental study on hypoglycemic effect of Gynostemma pentaphyllum polysaccharide. Chin. J. Gerontol. 2005, 25, 418–420. [Google Scholar]
- Du, X.Y.; Hou, Y.; Tan, H.; Han, Y.; Zhang, Y. Hypoglycemic Activity of Polysaccharide from Gynostemma pentaphyllum on Type 2 Diabetic Rats and its Mechanism. Sci. Technol. Eng. 2011, 11, 5754–5758. [Google Scholar]
- Ou, S.Z.; Mo, Y.N.; Chen, S.M.; Gao, L.F.; Ji, L.M.; Wang, D.M.; He, T.; Yu, D.R. Study on the hypoglycemic effect of the ethanol extract of Hainan gynostemma. China Trop. Med. 2014, 14, 38–40. [Google Scholar]
- Yassin, K.; Huyen, V.T.; Hoa, K.N.; Ostenson, C.G. Herbal extract of gynostemma pentaphyllum decreases hepatic glucose output in type 2 diabetic goto-kakizaki rats. Int. J.Biomed. Sci. 2011, 7, 131–136. [Google Scholar]
- Norberg, Å.; Hoa, N.K.; Liepinsh, E.; Van Phan, D.; Thuan, N.D.; Jörnvall, H.; Sillard, R.; östenson, C. A Novel Insulin-releasing Substance, Phanoside, from the Plant Gynostemma pentaphyllum. J. Biol. Chem. 2004, 279, 41361–41367. [Google Scholar] [CrossRef] [Green Version]
- Hoa, N.K.; Norberg, A.; Sillard, R.; Van Phan, D.; Thuan, N.D.; Dzung, D.T.N.; Jörnvall, H.; Östenson, C. The possible mechanisms by which phanoside stimulates insulin secretion from rat islets. J. Endocrinol. 2007, 192, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, L.C.E.; Rattigan, D.; Ehtesham, E.; Demmou, C.; östenson, C.; Sandström, C. Profiling and activity screening of Dammarane-type triterpen saponins from Gynostemma pentaphyllum with glucose-dependent insulin secretory activity. Sci. Rep.-UK 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Megalli, S.; Davies, N.M.; Roufogalis, B.D. Anti-hyperlipidemic and hypoglycemic effects of Gynostemma pentaphyllum in the Zucker fatty rat. J. Pharm. Pharm. Sci. 2006, 9, 281–291. [Google Scholar]
- Wang, Z.; Yin, Y.; Bian, Y.Y.; Tian, J.L.; Shi, L. Triterpenoids from Gynostemma pentaphyllum and their inhibition activity to α-glycosidase and protein tyrosine phosphatase 1B. Chin. Tradit. Herb. Drugs 2020, 51, 6142–6150. [Google Scholar]
- Zhang, X.S.; Bi, X.L.; Xiao, W.; Cao, J.Q.; Xia, X.C.; Diao, Y.P.; Zhao, Y.Q. Protein tyrosine phosphatase 1B inhibitory effect by dammarane-type triterpenes from hydrolyzate of total Gynostemma pentaphyllum saponins. Bioorg. Med. Chem. Lett. 2013, 23, 297–300. [Google Scholar] [CrossRef]
- Zhu, K.N.; Tian, S.S.; Wang, H.; Tian, Y.S.; Gu, G.Z.; Qiu, Y.Y.; Zhang, L.; Yang, H.X. Study on effect of gypenosides on insulin sensitivity in rats with diabetes mellitus via regulating NF-κB signaling pathway. China J. Chin. Mater. Med. 2021, 46, 4488–4496. [Google Scholar]
- Huang, X.F.; Song, Y.; Song, C.W.; Wang, M.Y.; Liu, H.X.; Yu, S.G.; Fang, N.B. Study on hypoglycemic activity of different components of Gynostemma pentaphyllum. Hubei J. TCM 2013, 35, 67–69. [Google Scholar]
- Huyen, V.T.T.; Phan, D.V.; Thang, P.; Hoa, N.K.; östenson, C.G. Gynostemma pentaphyllum Tea Improves Insulin Sensitivity in Type 2 Diabetic Patients. J. Nutr. Metab. 2013, 2013, 765383. [Google Scholar] [CrossRef] [Green Version]
- Huyen, V.T.; Phan, D.V.; Thang, P.; Hoa, N.K.; Ostenson, C.G. Antidiabetic effect of Gynostemma pentaphyllum tea in randomly assigned type 2 diabetic patients. Horm. Metab. Res. 2010, 42, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Lokman, E.F.; Gu, H.F.; Wan Mohamud, W.N.; östenson, C. Evaluation of antidiabetic effects of the traditional medicinal plant gynostemma pentaphyllum and the possible mechanisms of insulin release. Evid.-Based Compl. Alt. 2015, 2015, 120572. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.X.; Cong, J.; Gong, Q. Study on the effect of gypenosides on antioxidative injury in aged mice skin. Chin. J. Aesthetic Med. 2015, 24, 34–36. [Google Scholar]
- Yao, D.D.; Huang, S.H.; Liu, G.H. Effects of gynostemma pentaphyllum makino on hypothalamus in aging rats and its mechanisms. J. Med. Pr. 2007, 20, 1119–1121. [Google Scholar]
- Zhang, R.X. Study about the Effect of Gynostemma Pentaphyllum Polysaccharide on Anti-Oxidation Capability of Aging Mice Induced by D-Galactose. Master’s Thesis, Guangxi University, Guangxi, China, 2020. [Google Scholar]
- Zhao, T.T.; Shin, K.S.; Choi, H.S.; Lee, M.K. Ameliorating effects of gypenosides on chronic stress-induced anxiety disorders in mice. BMC Complementary Altern. Med. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.; Won, Y.H.; Kim, S.; Noh, S.; Park, S.; Jung, S.; Lee, C.K.; Hwang, B.Y.; Lee, M.K.; Ha, K.; et al. Supplementation with extract of Gynostemma pentaphyllum leaves reduces anxiety in healthy subjects with chronic psychological stress: A randomized, double-blind, placebo-controlled clinical trial. Phytomedicine 2018, 52, 198–205. [Google Scholar] [CrossRef]
- Mu, R.H.; Fang, X.Y.; Wang, S.S.; Li, C.F.; Chen, S.M.; Chen, X.M.; Liu, Q.; Li, Y.C.; Yi, L.T. Antidepressant-like effects of standardized gypenosides: Involvement of brain-derived neurotrophic factor signaling in hippocampus. Psychopharmacology 2016, 233, 3211–3221. [Google Scholar] [CrossRef]
- Shang, X.Y.; Chao, Y.; Zhang, Y.; Lu, C.Y.; Xu, C.L.; Niu, W.N. Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice. Molecules 2016, 21, 1085. [Google Scholar] [CrossRef] [Green Version]
- Rujjanawate, C.; Kanjanapothi, D.; Amornlerdpison, D. The anti-gastric ulcer effect of Gynostemma pentaphyllum Makino. Phytomedicine 2004, 11, 431–435. [Google Scholar] [CrossRef]
- Shan, L.N.; Shi, Y.X. Effects of polysaccharides from Gynostemma pentaphyllum (Thunb.) Makino on physical fatigue. Afr. J. Tradit. Complement Altern. Med. 2014, 11, 112–117. [Google Scholar]
- Kim, Y.H.; Jung, J.I.; Jeon, Y.E.; Kim, S.M.; Oh, T.K.; Lee, J.; Moon, J.M.; Kim, T.Y.; Kim, E.J. Gynostemma pentaphyllum extract and Gypenoside L enhance skeletal muscle differentiation and mitochondrial metabolism by activating the PGC-1α pathway in C2C12 myotubes. Nutr. Res. Pract. 2021, 15, e45. [Google Scholar]
- Yin, X.M. Effects of combination of aspirin and gypenosides on platelet aggregation. Chin. J. Hosp. Pharm. 2001, 21, 468–470. [Google Scholar]
- Jiang, Y.; Chen, X.L. Effects of Gypenosides on sedative hypnosis and neurotransmitters in rats. Zhejiang J. Tradit. Chin. Med. 2016, 51, 459–460. [Google Scholar]
- Srichana, D.; Taengtip, R.; Kondo, S. Antimicrobial activity of Gynostemma pentaphyllum extracts against fungi producing aflatoxin and fumonisin and bacteria causing diarrheal disease. Southeast Asian J. Trop. Med. Public Health 2011, 42, 704–710. [Google Scholar] [PubMed]
- Wang, T.; Guo, Y.S.; Jiang, Y.L.; Li, M.D.; Wu, L.; Liu, H.Q.; Su, P. Study of the immune enhancement effect of Epimedium polysaccharide and Gynostemma polysaccharide on PRRSV attenuated vaccine. Heilongjiang Anim. Sci. Vet. Med. 2021, 122–126. [Google Scholar]
- Sornpet, B.; Potha, T.; Tragoolpua, Y.; Pringproa, K. Antiviral activity of five Asian medicinal pant crude extracts against highly pathogenic H5N1 avian influenza virus. Asian Pac. J. Trop. Med. 2017, 10, 871–876. [Google Scholar] [CrossRef]
- Pang, M.X.; Fang, Y.Y.; Chen, S.H.; Zhu, X.X.; Shan, C.W.; Su, J.; Yu, J.J.; Li, B.; Yang, Y.; Chen, B.; et al. Gypenosides inhibits xanthine oxidoreductase and ameliorates urate excretion in hyperuricemic rats induced by high cholesterol and high fat food (Lipid emulsion). Med. Sci. Monit. 2017, 23, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Han, J.K.; Gao, W.; Su, D.Y.; Liu, Y. Gypenoside inhibits RANKL-induced osteoclastogenesis by regulating NF-κB, AKT, and MAPK signaling pathways. J. Cell. Biochem. 2018, 119, 7310–7318. [Google Scholar] [CrossRef]
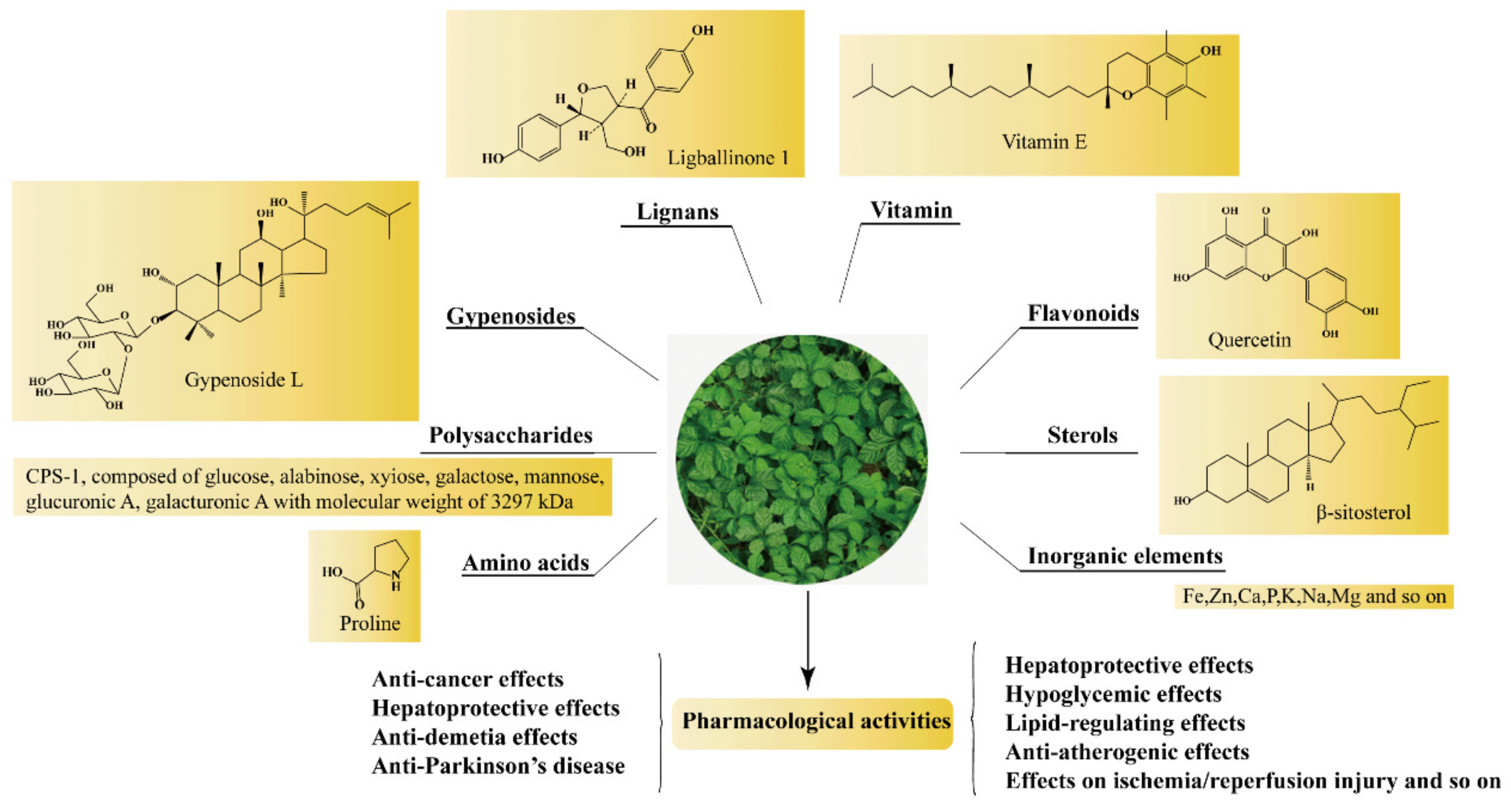
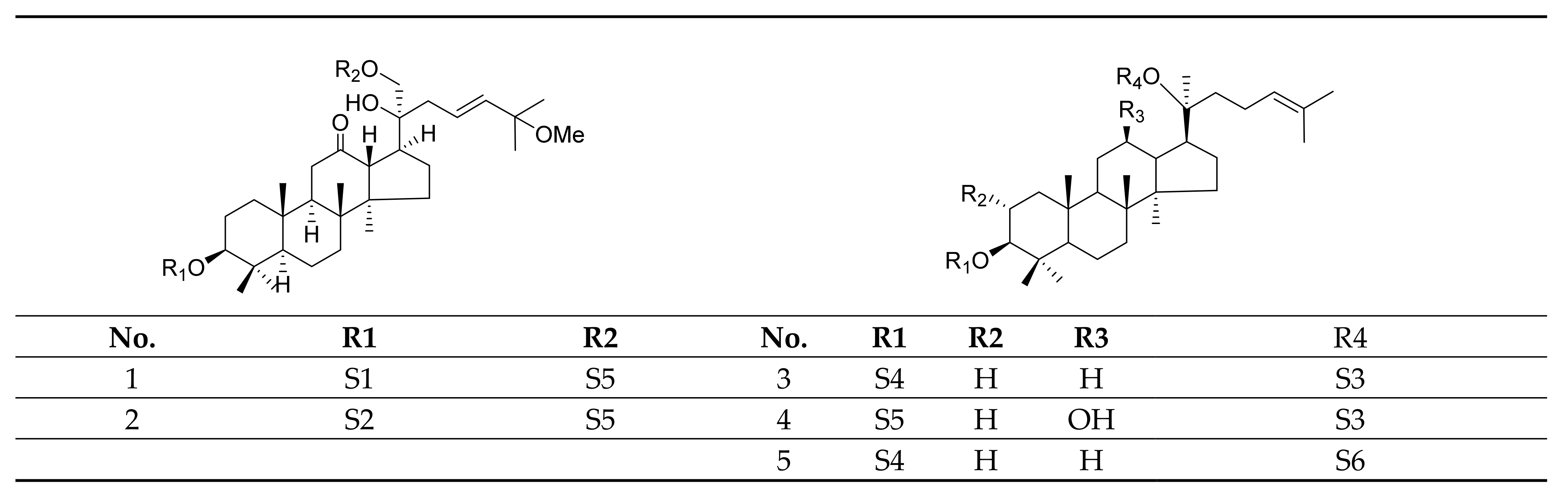
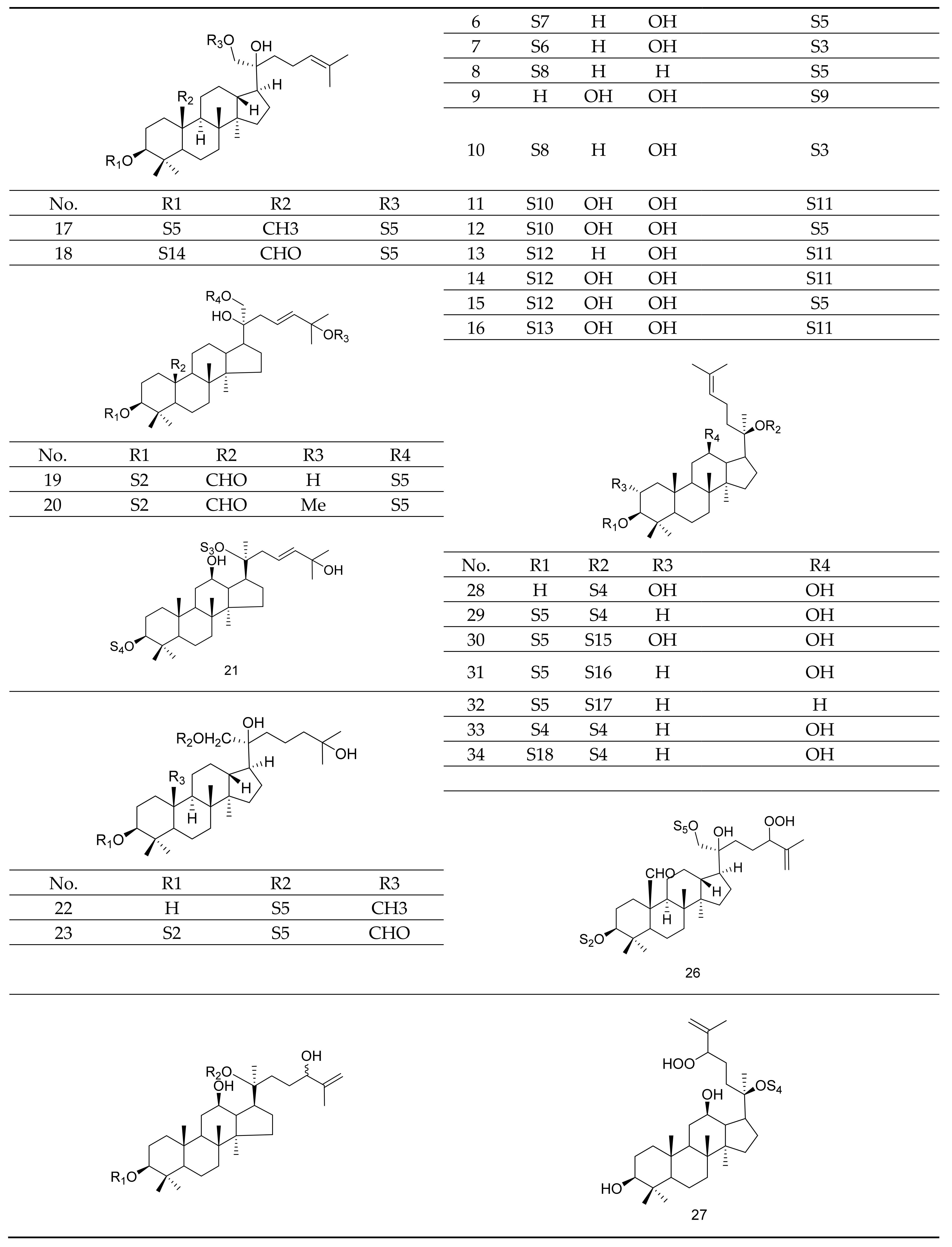
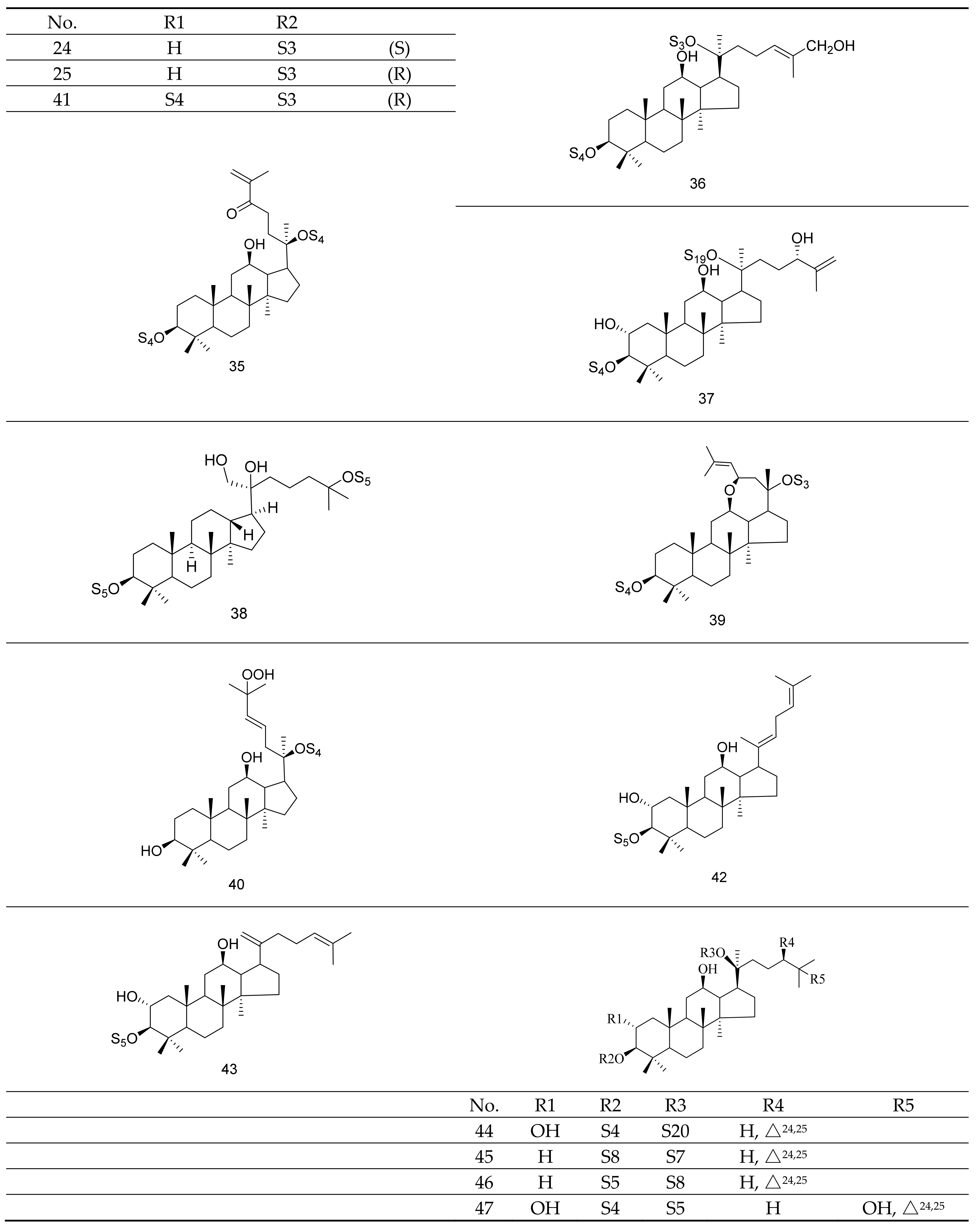
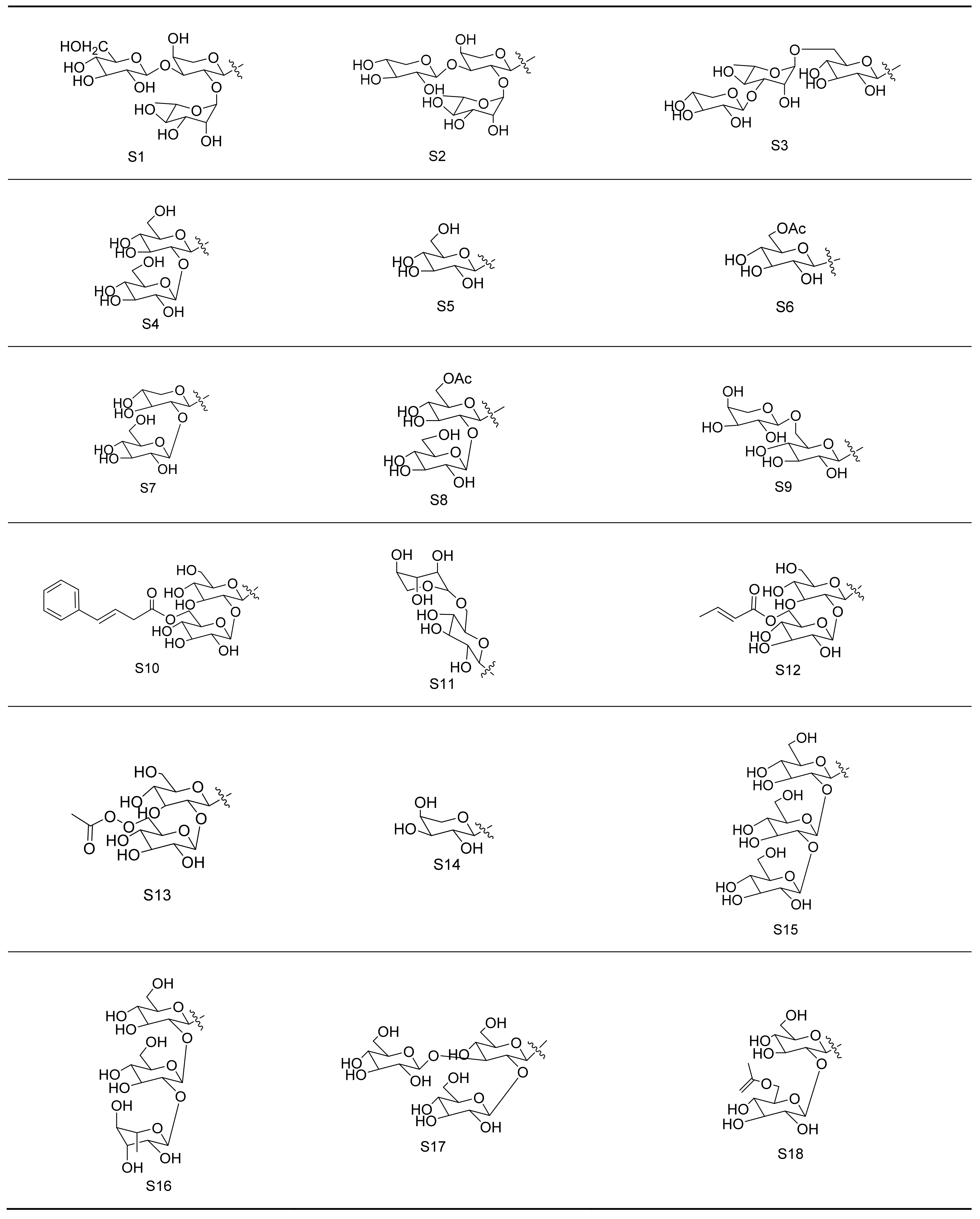
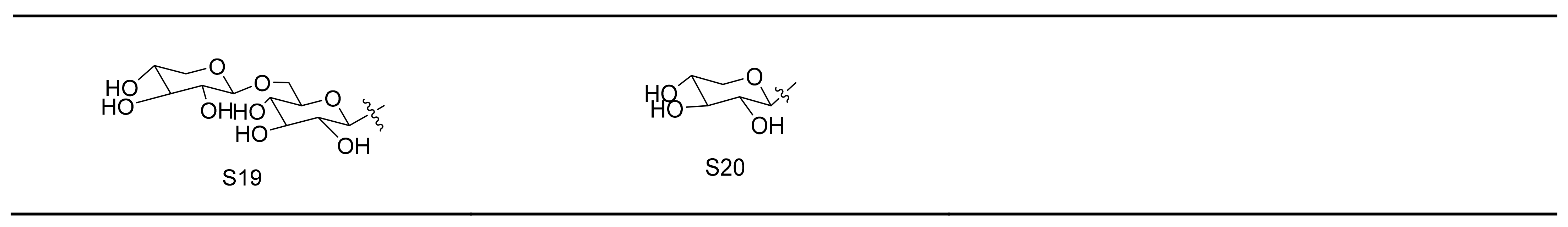
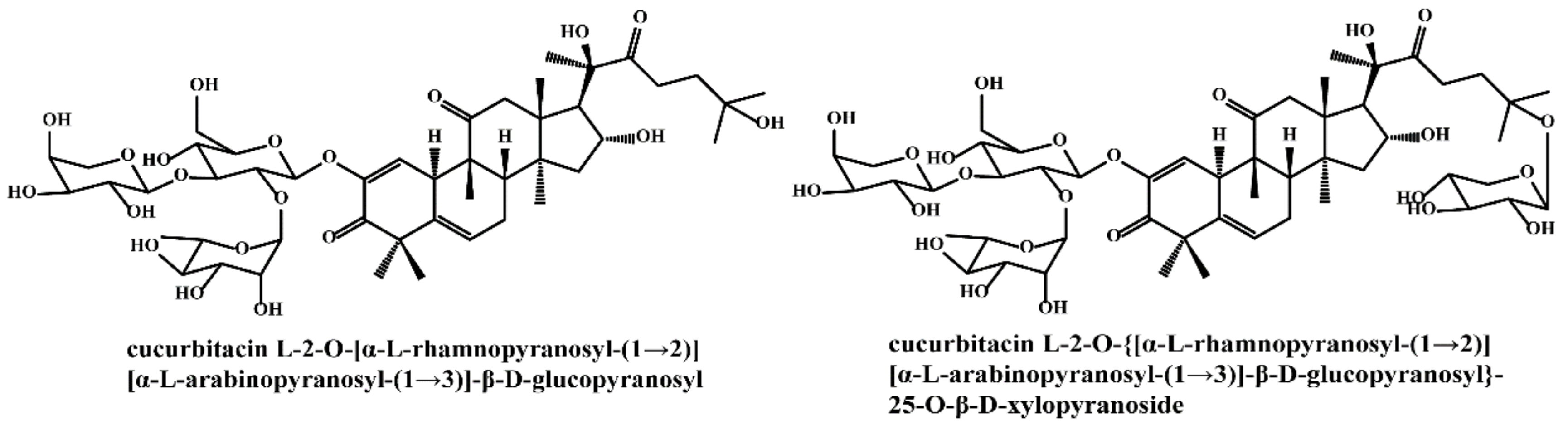
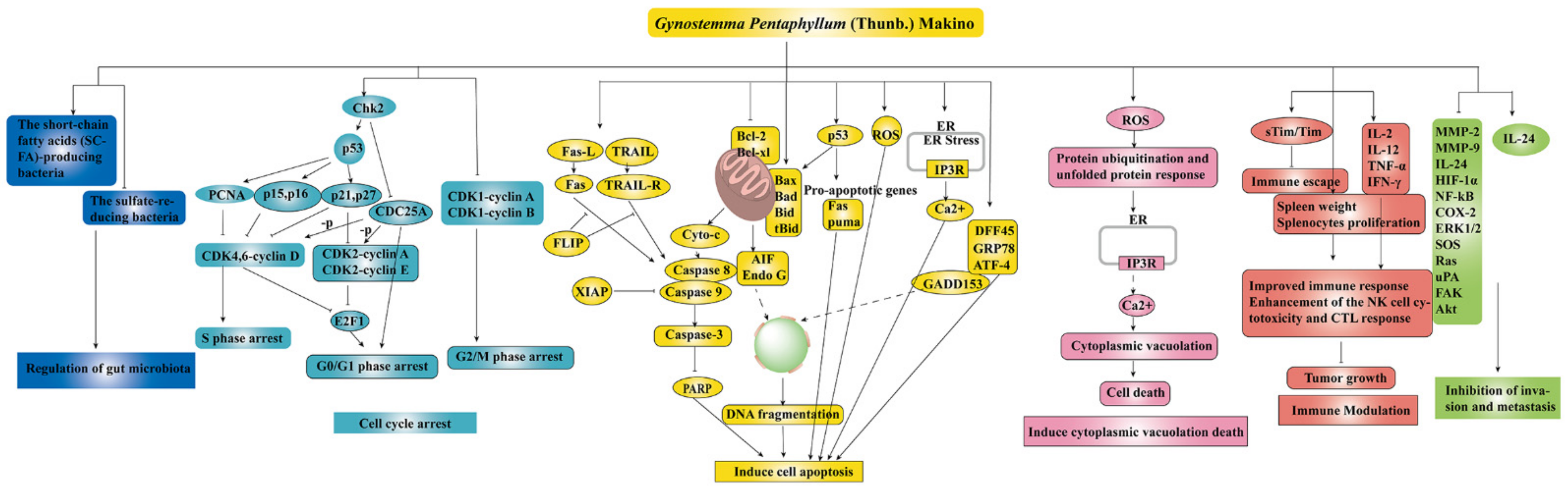
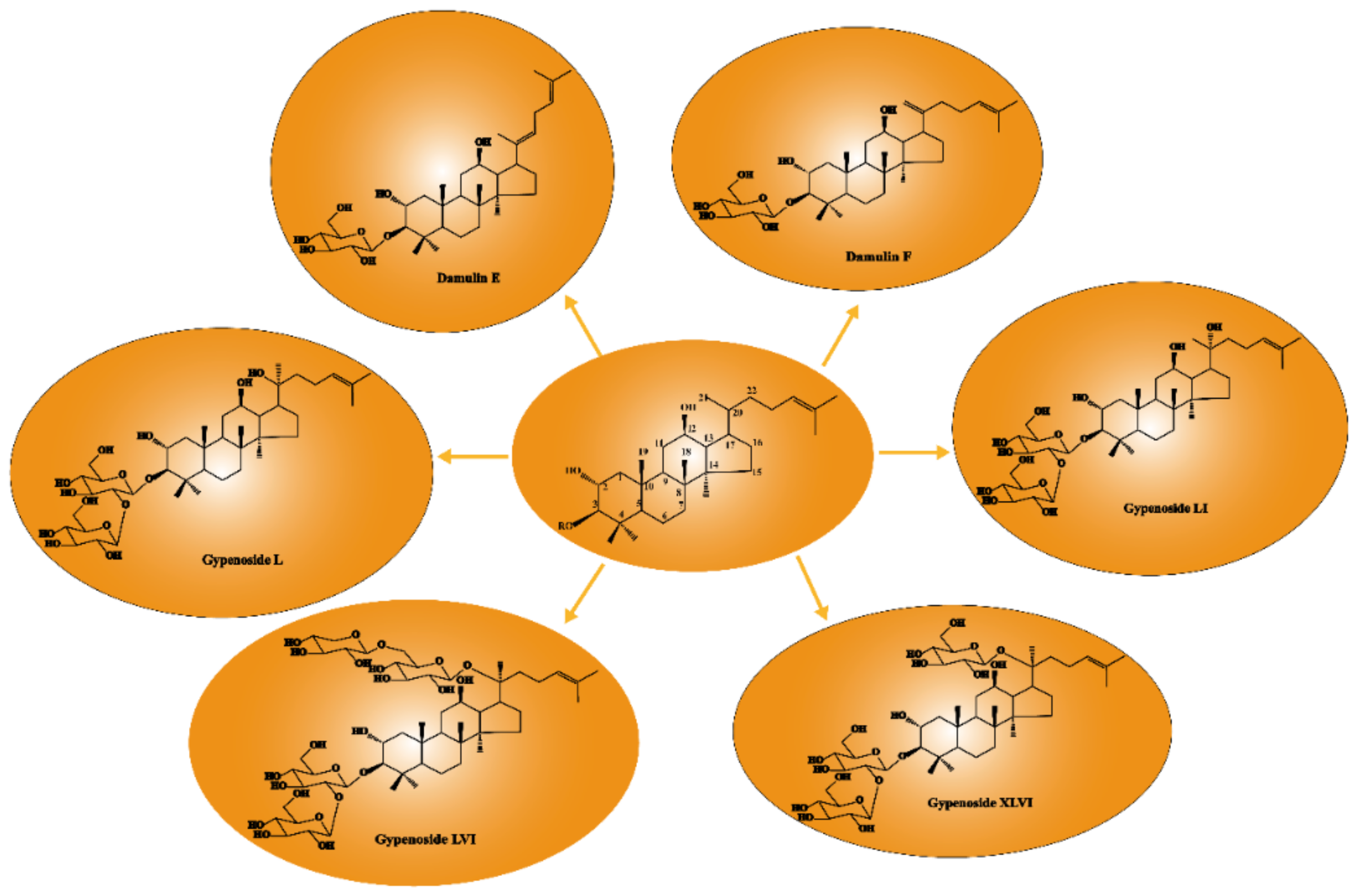
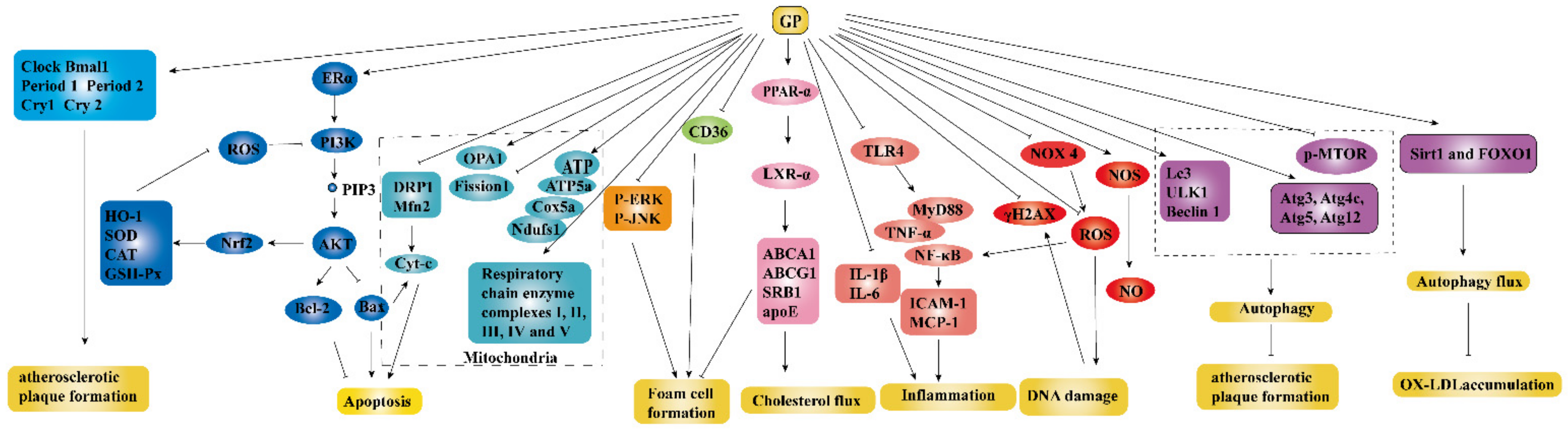
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).