Abstract
The trace element selenium (Se) is a crucial element for many living organisms, including soil microorganisms, plants and animals, including humans. Generally, in Nature Se is taken up in the living cells of microorganisms, plants, animals and humans in several inorganic forms such as selenate, selenite, elemental Se and selenide. These forms are converted to organic forms by biological process, mostly as the two selenoamino acids selenocysteine (SeCys) and selenomethionine (SeMet). The biological systems of plants, animals and humans can fix these amino acids into Se-containing proteins by a modest replacement of methionine with SeMet. While the form SeCys is usually present in the active site of enzymes, which is essential for catalytic activity. Within human cells, organic forms of Se are significant for the accurate functioning of the immune and reproductive systems, the thyroid and the brain, and to enzyme activity within cells. Humans ingest Se through plant and animal foods rich in the element. The concentration of Se in foodstuffs depends on the presence of available forms of Se in soils and its uptake and accumulation by plants and herbivorous animals. Therefore, improving the availability of Se to plants is, therefore, a potential pathway to overcoming human Se deficiencies. Among these prospective pathways, the Se-biofortification of plants has already been established as a pioneering approach for producing Se-enriched agricultural products. To achieve this desirable aim of Se-biofortification, molecular breeding and genetic engineering in combination with novel agronomic and edaphic management approaches should be combined. This current review summarizes the roles, responses, prospects and mechanisms of Se in human nutrition. It also elaborates how biofortification is a plausible approach to resolving Se-deficiency in humans and other animals.
1. Introduction
Selenium (Se) is a micronutrient essential for the proper functioning of plants and animals [1,2]. It was first described by the Swedish chemist Jacob Berzelius in 1817, however, its biological role was not determined until the 1950s when overdoses were linked to cardiac muscle dystrophy and acute hepatic necrosis [3]. In 1973, the beneficial biological role of Se as a key constituent of glutathione peroxidase (GPx) was discovered; thus it contributes to protecting the body from stress-induced oxidative damage to living cells. Subsequently, it was found that Se is not only associated with GPx, it is also connected with numerous other enzymatic activities within organisms’ cells; for example, Se biological forms like SeCys and SeMet are key constituents of iodothyronine deiodinase, which is crucial in the healthy functioning of the endocrine system [1,4].
It is estimated that globally one billion people are facing Se insufficiency [5] since Se is essential for the good functioning of the human immune, endocrine and reproductive systems, and also links with the function of the human brain [6]. An earlier study revealed that prolonged Se deficiency in the human body negatively affects the cardiovascular system and can lead to myocardial infarction, i.e., heart attacks [7]. Besides these examples, Se deficiency is also associated with Keshan and Kashin-Beck diseases, which predominantly occur in children and women of child-bearing age [8,9].
Generally, Se is uptaken by humans through consumption of Se-enriched plant and animal products, in particular from plant sources. Plants take up Se from the soil as selenates [10], which are then converted into the organic forms SeCys and SeMet [11]. Increasing the concentration of Se in edible plants’ parts/products is a viable pathway to overcome human Se deficiency. Recently, Se-biofortification combined with improved agronomic, breeding, molecular, biotechnology, and genetic engineering approaches have been recognized as a leading tool to enrich plant-food products with Se. While there is a large, disperse body of literature on the roles and mechanism of Se in human health and its enrichment in plant foods, a comprehensive review covering all these aspects is lacking. This review summarizes the roles, responses, and mechanisms of Se in humans and the potential to enrich plant foods with this micronutrient through biofortification. The review also explains how biofortification is a plausible approach to resolving Se-deficiency in humans and other animals.
2. The Natural Form of Selenium and Its Deficiency and Toxicity Symptoms
2.1. Natural Form
Selenium (Se) is widespread across the Earth and is present in the atmosphere, lithosphere, hydrosphere and biosphere [12]. The weathering of rocks and the eruption of volcanic gases are key sources of Se into the environment. Additionally, the decomposition of Se-enriched organic matter, through biomethylation by microorganisms, maintains a positive flow of this element into the atmosphere [1]. These mechanisms contribute to the presence of volatile Se compounds, viz., hydrogen selenide (H2Se), dimethyl selenide (DMSe), and selenium oxide (SeO2). Globally, the Se content in arable soils ranges between 0.33 and 2 mg/kg [13]. Se-rich areas are known as seleniferous areas [14]. The Se concentration in soils varies depending on the management of the local environment and the presence of both Se-rich parent materials and microorganisms necessary for its release into the atmosphere [15].
The soils which have originated from igneous rock, granite, sandstone, and limestone are all rich in Se [16]. Conversely, soils in temperate and humid climates are generally low in Se. Irrespective of soil depth, the Se content in mineral-enriched soils fluctuates ~14 mg/kg [17]. Se-rich materials include berzelianite (Cu2Se), klaustalite (PbSe), and naumanite (Ag2Se) [18]. Anthropogenic activities, such as the combustion of fossil fuels, metal smelting, international shipping and the over-use of inorganic fertilizers are primarily responsible for additional contributions of Se into the atmosphere (and thus into agricultural soils), from the before mentioned minerals [17].
Se-rich soils have been observed in the United States, Russia, parts of China, Australia, Canada, and Ireland [4]. In contrast, New Zealand and a wide portion of Europe have soils which are largely Se deficient [1]. Tomza-Marciniak [19] calculated that Se deficiencies exist in more than 70% of countries. However, the total Se content in soil is not a reliable estimate of plant available Se or the amount of Se available to humans and animals through plants [20]. The Se availability of plants is determined by a large number of soil chemical and biochemical characteristics, including sorption, disruption, soil pH, presence of other nutrients and methylation [21]. For example, Se uptake is higher for plants high in sulfur [22]. Se-rich foods include seafood, eggs, chicken, nuts, mushrooms, and green vegetables including spinach, cauliflower, and cabbage. Se concentrations in humans vary with agroclimatic region and daily diet.
In water, Se is present in minute quantities as selenates or selenites. Groundwater contains higher concentrations of Se than seawater [16,18]. This is largely the result of runoff of Se-rich fertilizer from intensively managed agricultural soils, as well as Se secretion from parent rock material [23]. In potable water, 10 µg Se per litre of water is acceptable according to the World Health Organization [24].
2.2. Selenium Deficiency Symptoms
2.2.1. Symptoms in Human
Prolonged Se deficiency adversely affects the cardiovascular system, which may be a cause of myocardial infarction [16]. Se deficiency also causes Keshan and Kashin-Beck diseases, which primarily occur in childbearing women and in children who live in areas deficient in Se [25]. A moderate deficiency of Se in daily food habit reduces immunity, can impair the nervous system and may cause congenital hypothyroidism in fetuses [26]. Additionally, Alzheimer’s disease, depression and anxiety have been associated with prolonged Se deficiencies [27]. Se may contribute to the suppression of HIV and slow the progression to AIDS [28]. Se is also necessary for the developing fetus in women and animals [16,29]. An insufficiency of Se in the human diet affects the thylakoid gland and may lead to moodiness and the impairment of behaviors and cognitive functions [30,31,32]. Se deficiency reduces the activity of 5′-thyronine deiodinase enzymes, leading to low triiodothyronine concentration in blood. Moreover, a deficiency of Se accelerates human ageing [31].
2.2.2. Symptoms in Animals
The majority of Se-deficiency symptoms have been observed in animals when less than 0.1 mg Se is present per kg of animal diet. Most commonly, Se deficiency causes a myo-degenerative syndrome [32] Table 1, also known as white muscle disease (WMD), whereby animal muscles are pale in appearance [33]. WMD may occur in all livestock including birds, and most seriously damages the skeletal (fibres/highly elongated cells) and cardiac muscles [34]. The deficiency of Se also impairs an animal’s immunity [35]. The clinical signs of Se deficiency in animals include reductions in appetite, fertility and growth, as well as muscle weakness [36]. Specific Se deficiency symptoms include heart disease in pigs, placenta retention in cows, and birds both a higher embryonic mortality rate and muscular dystrophy [33,37].

Table 1.
Se responsive diseases in animals.
2.3. Selenium Toxicity
2.3.1. Toxicity in Humans and Animals
Both excessive and insufficiency of Se are detrimental to human health. The consumption of higher doses of Se can be toxic. There is a narrow limit between safe-and-adequate Se uptake and overconsumption leading to toxicity. There are only limited reports on human Se toxicity; this may be because many commonly consumed foods are Se non-accumulators. The symptoms of Se toxicity are hair loss and skin and nail lesions [38], hypotension, tachycardia, muscle contractions dizziness, nausea, vomiting, facial flushing, tremors and muscle soreness. In extreme cases, acute Se toxicity can cause serious intestinal and neurological problems, heart attack, kidney failure and death [39]. Excessive Se uptake can also damage the mucus membranes of the digestive tract, and lead to ongoing nausea, diarrhoea and also increase risk of type 2 diabetes [38]. In the case of animals, the death of poultry due to Se toxicity in wheat grains used as chicken feed was reported in South Dakota, USA [40].
The limit between safe and toxic amounts of Se is small and has yet to be standardized in many geographical locations. The recommended dietary allowance of Se varies in humans with age, gender, pregnancy and lactation Figure 1. Pregnant or lactating women require higher 9% and 27% daily amounts of Se than other women [41]. Human beings must consume around 55 micrograms of Se per day and not exceed the maximum limit of 400 micrograms per day [42]. The World Health Organization recommends a daily average intake of 55 µg of Se, while the recommendation is varied with age, gender, diet and geographic location [43]. The International Food and Nutrition Board has recommended an average daily intake of 40–70 µg Se and 45–55 µg Se for men and women, respectively, and 25 µg Se for children [2,44]. A daily dose of 55–200 µg of Se is recommended for healthy adult humans [45,46]. Table 2 illustrates the maximum recommended daily Se intake limits, above which symptoms of Se-toxicity such as significant hair loss and abnormal nail growth appear [41].
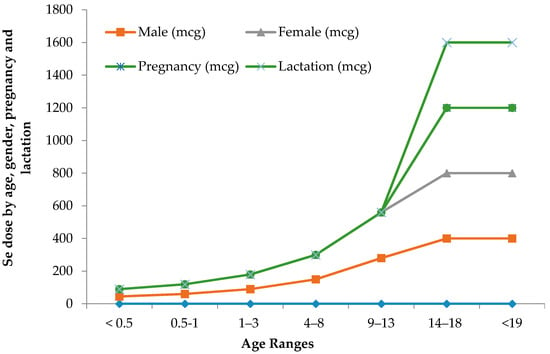
Figure 1.
Recommended maximum daily dietary allowances for Se.
Table 2.
Recommended maximum daily Se intake levels.
In case of the available forms Se, Ríos et al. [47] regarded that >40 μmol Se/L of selenate as toxic, while Hawrylak-Nowak [48] estimates that safe concentrations of Se are between 20 and 15 μmol/L for selenate and selenite, respectively. However, Ríos et al. [49] reconsidered and set an upper limit of 80 μmol Se/L.
2.3.2. Selenium Phytotoxicity
Toxic levels of Se within plant tissues are greater than 5 mg/kg [50]. Se toxicity does not occur when selenate is applied at rates between 10–200 g Se per ha, which are commonly recommended for wheat biofortification. Se is usually not considered essential for taller plants, while soils low in Se are considered to neither inhibit plant growth nor reduce crop yield [50,51]. However, some research has indicated the beneficial effects of low doses of applied Se on crop performance. For example, increased growth of ryegrass (Lolium perenne) and lettuce (Latuca sativa) was recorded when crops were fertilized with Se and exposed to UVB radiation. Low-dose applications of Se are useful for crop plants only when they are under oxidative stress, otherwise, this microelement is not essential.
3. Importance of Selenium for Global Human Nutritional Security in the 21st Century
Se is an essential mineral element, obtained by humans and animals through their diets. Biofortification is a plausible strategy to enrich Se in foods. Options to enhance Se content in human foods include: increasing the number of foods naturally high in Se (e.g., Brazil nuts); agronomic biofortification (i.e., incorporation of Se fertilizers); genetic biofortification; plant breeding strategies; and supplementing organic Se directly to humans or livestock [52,53]. Those living in areas with low endemic levels of Se generally do not consume adequate amounts of Se, as food crops and animal feeds do not take sufficient Se from the soils in which they grow. In the case of plants, the amount of Se in plants depends on the Se concentration in the soils in which the plants are grown. A study of Se concentration in Brazil nuts demonstrated high spatial variability: while a single Brazil nut from one area contained 288% of the recommended daily Se intake, other nuts are grown elsewhere contained only 11% of the recommended daily Se intake [42].
3.1. Health Benefits of Selenium for Humans
Among the total amount of Se in an average human (~3–20 mg), around 47% is found in skeletal muscles/fibre cells and about 4% in the kidneys [54]. Generally, the presence of Se is determined by its concentration in blood serum [55]. Highest Se concentrations are observed in adults around 60 years of age after this Se concentration gradually decreases [56]. A Se deficiency occurs when the blood contains less than 85 µg Se per litre of blood serum. Low Se levels increase the risk of some cancers [57] while over-sufficiency of Se may result in anaemia, hair loss, bone stiffness or blindness [54]. Where Se is present in the air in concentrations above 0.2 mg/m3 it may be inhaled by humans and other animals [58]. Both under- and over-supply of Se are harmful to human health [59].
Daily doses of 100–200 µg Se inhabits of genetic damage and the development of some cancer cells [60]. A daily intake of over 200 µg Se per day may accelerate the presence of cytotoxic T cells within the body. A daily intake of 100 µg Se has been observed to reduce clinical depression and anxiety [61]. Exceeding an intake of 1500 µg day−1 has been observed to be toxic to humans [62]. As the Se concentration in plant and animal foods reflects the endemic Se concentration of the soil in which they are grown, Se concentration in foods varies globally. For example, Se intake is more than 90 µg/day in the United States [59], in Venezuela, it is 326 µg/day [63], while in some European countries the concentration is lower than the recommended value at around 30 µg/day [64]. The Health benefits of Se in the physiological processes of the human body are illustrated in Figure 2.
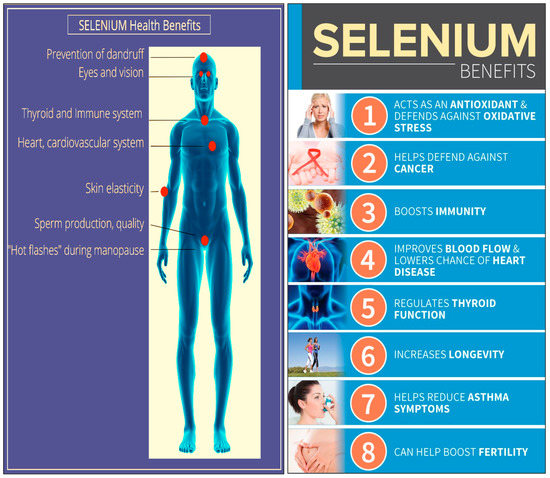
Figure 2.
Health benefits of Se in the physiological processes of the human body.
The production of Se-enriched foods, such as eggs, meat, and milk, is attractive worldwide as these products improve the nutritional status and health of those who may otherwise be Se deficient. However, while this food biofortification would benefit those with Se deficiencies, people with high inherent Se intake through their diet may be adversely affected by the biofortification process. Therefore, it is recommended that people living in areas with high Se concentrations in the soil do not consume excessive amounts of Se-fortified foods [65]. Se is a powerful antioxidant which may contribute to fighting cancers, viral infections, and aging. It is important for the normal functioning of the thyroid, brain, heart, and reproductive system. Se is the only micronutrient (as SeCys protein) mentioned in the human genome [66]. The effect of Se in mitigating some cancers has been reported by Combs and Lu [67]. The numerous health benefits of Se are discussed in the following sub-sections.
3.1.1. Selenium is a Strong Antioxidant
Se acts as a strong antioxidant, which reduces cellular damage caused by free radicals and protects the body from heart disease and some cancers [68] Figure 2. Free radicals are naturally produced as byproducts of living cells and may also be a result of smoking, drinking or mental stress [69]. Furthermore, the production of free radicals has been linked to many human diseases including cardiovascular disease, Alzheimer’s, some cancers, and premature ageing [70].
3.1.2. Selenium Reduces the Risk of Some Cancers
Se reduces the risk of some cancers [71,72]. Se reduces DNA damage and the effects of stress while boosting the body’s immune system. The WHO recommended daily dose of Se improves the quality of life in patients undergoing chemotherapy [73]. These beneficial effects of Se have been observed in Se-enriched plant foods but not in supplements. As well, Se may reduce the side effects of chemotherapy [71,72,73].
3.1.3. Selenium Protects Against Cardiovascular Problems
A Se-sufficient human diet contributes to a healthy heart and cardiovascular system [65,74]. Se reduces the risk of heart attack by mitigating inflammation and oxidative stress in the body which otherwise may contribute to plaque build-up within the lining of arterial walls, leading to atherosclerosis [75]. Arterial thickening contributes to heart attacks and heart disease.
3.1.4. Selenium May Improve Some Mental Illnesses
Insomnia may contribute to anxiety, depression and reduced quality of life [76]. Often patients with poor mental health have been found to have low blood Se levels [77].
3.1.5. Selenium Is Beneficial for Thyroid Health
Selenium helps protect the thyroid against oxidative cell damage and aids the release of thyroid hormones which regulate growth and development with the human body [78,79]. Se is thus beneficial for the normal functioning of the thyroid gland [80,81].
3.1.6. Se Strengthens Immunity and May Reduce Breathing Difficulties
Elevated Se levels in blood serum have been associated with an enhanced human immune system [82,83,84]. Additionally, Se may benefit some asthmatics by reducing swelling in airways [85,86].
3.1.7. Finland Case Study: Selenium Biofortification of Human and Livestock Feed Crops
Heart disease was a leading cause of death in Finland in the 1960s and 1970s. To counter this, the Finnish government required the introduction of Se (as selenate) fertilizer in all multiple-nutrient fertilizer mix used for agriculture from 1984. Table 3 illustrates the success of this Se-biofortification program in Finland. Initially, Se fertilizer was applied at 16 mg Se/kg of fertilizer in grain and horticultural production and 6 mg Se/kg for pasture and hay production. The program was so successful in increasing plant Se concentrations, and thus the level of Se in humans in Finland, that in 1990 a rate of 6 mg Se/kg fertilizer was adopted universally [87,88]. In 1998, Se supplementation was increased to 10 mg Se/kg fertilizer for all fertilizers applied to crops [87]. As a result of the agronomic Se biofortification program, human health in Finland increased markedly: dietary Se intakes trebled and blood plasma Se concentrations approximately doubled within 3 years of the introduction of the biofortification [88]. The prevalence of heart disease and some cancers in Finland have significantly decreased since 1985.
Table 3.
Results of a Se-biofortification program in Finland.
3.2. Importance of Selenium for Both Plants and Animals
3.2.1. For Plants
In plants, Se is most commonly found as SeMet, methyl-SeCys or ɣ-glutamyl-Se-SeCys (ɣ-Glu-MeSeCys) [93]. Supplementing wheat and maize with Se at rates of up to 100 gm Se/ha did not affect crop productivity in terms of grain or stover yield [88]. Additionally, applications of Se have been reported to increase plants’ resistance to oxidative stresses [94,95]. Further, the presence of the antioxidants glutathione peroxidase (GSH-Px) and superoxide dismutase (SOD) is enhanced with Se supplementation and, in some cases, the concentration of lipids are reduced. Hence, Se may enhance the natural defence mechanisms of plant against pests, particularly insects [96]. D’Amato et al. [97] reported that the oil yields from olive trees were enhanced in terms of intensity and stability of color following application of Se. Se biofortification in peach and pear trees increased the Se concentration in fruit and reduced fruit-softening rates, thus increasing the shelf-life of fruit products [98]. However, biofortification with high concentrations of Se negatively affected plant growth in lettuce [48] and retarded germination in mustard [98]. Some crops, such as brassicas (e.g., broccoli, B. oleracea L.) were unaffected by applications of Se [99].
Se fertilizers can be effectively utilized to alleviate poisoning from some toxic metal and metalloids stress in field crops. Dipping the roots of rice seedlings in a Se solution before transplanting reduces the effects of arsenic, which inhibits seedling growth [93,100]. Additionally, rice plant height, number of tillers, chlorophyll content, panicle length and kernel weight were also all significantly enhanced when rice seeds were coated in Se before sowing [101,102]. Se-coating rice seeds also reduce arsenic phytotoxicity seedlings and enhance crop productivity [100,101,102,103].
3.2.2. For Animals
In ruminant animals, Se is more readily absorbed in organic forms than in inorganic [104]. SeMet, a commonly occurring form of Se, can be taken up in animal cells instead of methionine, as these cells do not distinguish between methionine and SeMet during protein synthesis [105]. Organic SeMet is a valuable source of Se to facilitate the rapid synthesis of seleno-proteins [106].
Se-fortified alfalfa, fed daily, is one of the most effective tools to improve Se concentration in animals [107]. This significantly increases the Se concentration in the edible meat and provides adequate Se to humans who consume the meat. The concentration of Se in an animal’s meat is directly proportional to the Se concentration in its feed or forage [108]. Marine animals take up Se from seawater, phytoplankton, and other marine feeds, such as krill [109]. Water-soluble Se is also absorbed through the gills, epidermis and gut of marine animals. There is a strong correlation between low Se levels and mercury absorption in fish [110]. Mercury toxicity reduces with higher Se concentration in fish tissues, this relationship may be important for research into aquatic food farmed under suboptimal water quality conditions.
4. Biofortification—A Sustainable Agricultural Strategy for Reducing Micronutrient Malnutrition
The concentration of Se in food ultimately depends on the concentration of plant-available Se in cultivated soils. The availability of Se in soils is mediated by soil pH, redox potential, cation exchange capacity (CEC) and the soil concentrations of S, Fe, Al and C [111,112]. This microelement is present everywhere but not uniformly distributed. Lyons et al. [113] illustrate the variability in soil Se concentration as shown in Table 4.
Biofortification of crops and animal feed improves the nutritional quality of food products and may help to reduce global malnutrition [88]. Plants are the most abundant sources of Se in many countries, followed by meat and seafood sources [114]. Thus, it is critical to enhancing Se uptake by plants, and ultimately the Se concentration in the human diet, to alleviate Se deficiency-induced human disorders. Se biofortification is the most effective approach to increase Se concentrations from agriculture [115].
Table 4.
Variability in Se concentrations in soils from different locations across the globe.
Table 4.
Variability in Se concentrations in soils from different locations across the globe.
| Locations | Soil Types | pH (H2O) | Total Soil Se µg/kg | Se in Cereal Grain µg/kg | References |
|---|---|---|---|---|---|
| Yangshuo, China | Ishumiso | 8.3 | 700 | 20 | Lyons et al. [113] Zhu et al. [116] |
| Minnipa, South Australia | Calcareous Xerochrepts | 8.6 | 80 | 720 | Lyons et al. [113] Williams et al. [117] |
| Charlick, South Australia | Typic Natrixeralf | 6.6 | 85 | 70 | Lyons et al. [113] Thavarajah et al. [118] |
| East Zimbabwe | Typic Kandiustalf (ex-granitic parent material) | 5.0 | 30.000 | 7 | Lyons et al. [113] Winkel et al. [119] Fordyce et al. [120] |
4.1. Selenium Biofortification through Agronomic Management
Se is a vital micronutrient element which also enhances the antioxidants properties of food crops [121,122]. Se is present in animals (including humans) and plants and influences many physiological processes including promoting growth [123]. It is believed that many populations have less than the necessary intake of Se, which increases the risk of many diseases [124]. Se enriches in plants varies depending on geography and Se availability within soils, and thus Se uptake by humans varies but is tied to diet [125]. Worldwide, the distribution of Se within soils varies topographically, with a diversified range (from near zero up to 1200 mg kg−1) [126]. Therefore, in many locations’ agriculture plants require Se inputs from external sources [127].
Different agronomic management practices can be used to biofortify Se in plants: these include soil inorganic fertilization, foliar application, and also as organic fertilizers. Se-biofortification through agronomic management aims to standardize and regulate the concentration of Se within crop products by incorporating the right time, right source, the right method, and the right amount of Se fertilizer. Agronomic Se bio-fortification has many advantages over direct Se supplementation as the inorganic Se absorbed by the plant is transformed into an organic form, which has a higher bioavailability [128]. The application of Se fertilizers to soil commonly results in increased total and bioavailable Se [88], and thus in higher Se concentration in edible crop products [20,129]. Agronomic Se bio-fortification reduces the risk of Se leaching into groundwater or lost through volatilization, as Se binds to soil organic matter and remains within the soil. Field tests using inorganic Se fertilizers in Finland and New Zealand successfully demonstrated its bioavailability to plants [130,131]. Foliar Se fertilization in rice, wheat, and lettuce [132] suppressed nutritional deficiencies in human populations [133].
4.1.1. Selenium Biofortification through Direct Soil Fertilization with Inorganic Fertilizers
Applying selenate-based fertilizers enhance Se uptake by plants and, subsequently, the Se concentration in animals and humans who consume those plants [126]. This management practice is used where Se is in deficit by adding Se to inorganic fertilizers applied to agronomic soils, most commonly as selenate or as selenite of sodium or barium salts, which are applied to the soil either directly or diluted in a water-based fertilizer [134]. However applying Se broadly at high concentrations is generally not economically sustainable, and so site-specific Se applications which account for existing Se available within the soil and crop demand must be considered [135]. For example, Ramkissoon et al. [88] observed that application of 3.33 µg kg−b of Se (equivalent to 10 g ha−o) to wheat can be made more efficient by its co-application with macronutrient carriers, either to the soil or to the leaves during the awn-peep stage and observed that grain Se concentrations varying from 0.13–0.84 mg kg−w. For raising the grain Se concentrations, soil application of selenate was found 2–15 times more effective than application of granular Se-enriched macronutrient fertilizers such as N, P, K or S. While co-application of Se applied as foliar with an N carrier doubled the Se concentration in wheat grains compared to the application of foliar Se.
In agricultural production systems, Se is taken up from the soil by plants, this must be balanced by the application of Se fertilizer. The amount of Se fertilizer applied depends on the Se concentration in both the fertilizer and the soil. Se also accumulates in soils through atmospheric deposition and from irrigation water and some industrial processes [127].
The solubility, mobility, and bioavailability of Se within a soil generally depend on its exact chemical form and how sturdily it is tied to soil particles. For example, in alkaline soils the dominant form of Se is selenate and the absorption of selenate by plants is affected by the presence (or not) of different ions which may be present in the soil (e.g., K+, Ca2+, Mg2+, SO2–, Cl−) [136,137,138]. Different fertilizers differently influence the amount of Se within a soil as they alter the concentration of different ions within the soil, and thus the availability of Se to crops. Absorption of Se within soils may be reduced due to the presence of competitive ions such as K+, Ca2+, Mg2+, SO2− and Cl− [139].
Se absorption is increased in soils by the addition of chelating agents such as compost and other organic matter. However, adding organic matter to soil may reduce the Se uptake capacity of crops [136]. As Se is not used in agriculture to improve soil fertility or crop productivity and thus management practices which facilitate Se bioavailability to plants are often not prioritized by farmers [123]. Therefore, it may be helpful to identify the principal factors that limit Se concentration within soils and address these to enhance the concentration of Se within the animal and human foods.
Se is well suited to agronomic biofortification of food crops. Research has shown that the selenite form (SE) of Se, when used in biofortification, is readily taken up by plants in a wide range of soils. This SE is transported throughout the plant and stored in its numerous edible parts [140]. In cereal crops, this SE is transformed into seleno-methionine and stored in the endosperm. Therefore, milled products like white flour and polished rice are significant bioavailable sources of Se [137].
Selenate (i.e., Se in its highest oxidation form, +6) is a more effective form for use in soil fertilization than selenite (i.e., Se in +4 oxidation form) [141,142]. Selenate is up to 33 times more effective than selenite in plant fertilization [126]. In soils with a high clay, content selenite is absorbed quickly but is of limited availability to plants. Applications of selenite increase grain yield in lentil by 10% with significant antioxidant activity (66% inhibition) compared to the control (59% inhibition) [143]. The form of selenates is highly water-soluble and available to uptake by plants, while easily leached from the soil solution. In aerated soils with neutral to higher pH, this form of Se is dominated. For example, soils with a high content of Ca and Mg CaSeO4 generally creates MgSeO4, this form is easily soluble and represent total Se soluble in soil [144]. In soils rich in organic matter and water and without air entry selenates are transformed and reduced to less mobile forms. With decreasing pH and redox potential in soil SeO32− dominate, being less available for plants than SeO42− [145].
4.1.2. Selenium Biofortification through the Foliar Application with Inorganic Fertilizers
In some circumstances, soil-based applications of Se-enriched fertilizers are ineffective, largely due to the physical, chemical and biological soil properties, including soil texture, pH, redox potential, microbial activities and inherent Se concentration [146]. As an alternate method of fertilization, the foliar application is widely accepted [126]. The effectiveness of this method for Se supplementation has been successfully reported in various crops such as rice [147], wheat [148], potato [149], soybean [150] and carrot [151]. However, when Se concentration exceeds 100 µg Se/mL of water, it causes phytotoxicity in some crops [151]. During foliar application (SeF), the Se solution must be distributed using well-calibrated equipment; spraying should not occur on rainy or windy days, and applications need to be made at the late vegetative stage, where there is an adequate surface area to facilitate maximum absorption of Se. In the soil application (SeS), Se is effective in the period from early growth of seedlings to plant maturity for uptaking Se by roots [152].
Comparison of the long-term environmental impact of Se application directly into soils or as a foliar spray has shown that foliar application reduces the potential accumulation of Se in the environment. Lower concentrations of Se are used in foliar applications than when fertilizer is applied directly to the soil [153].
Se fertilizer is generally applied in soils with a pH 5.5 or greater. Se translocates within the plant through the phloem and is deposited in the edible portions of crop plants as SeMet. This organic form of Se is found across all cereal grains, particularly in white flour and polished rice, and is readily absorbed by the small intestine of both humans and animals [52].
At the same time, Se biofortification of agricultural soils is a relatively wasteful process. Approximately 12% of Se applied directly to soils is recovered in plants; this amount is higher when foliar applications are used [88]. For example, a field study with the purple-grained wheat cultivar (‘202w17′) and common wheat cultivar (‘Shannong 129′) showed that both soil and foliar application of Se boosted the organic Se concentration in roots, shoots, and grains of both cultivars, but the higher concentration of Se in the grain of two cultivars was noted when Se was applied as foliar. Foliar application of Se enhanced approximately 1.5-fold higher concentration of organic Se in grains of cultivar ‘202w17′ than cultivar ‘Shannong 129′ [152].
Foliar application of Se is safer and more effective, easy and cost-effective for Se biofortification than other Se fertilizer application methods [90]. Foliar application of Se at an appropriate crop growth stage accelerates the efficiency of fertilizer uptake and boosts crop production, reducing wastage. The timing of Se application is critical in cereal crops. The most effective time for Se fertilization is between booting and milking stages as this is when there is a maximum number of green leaves within the field [129].
4.1.3. Selenium Biofortification through Organic Fertilizers
The presence of organic matter in drier environments increases the absorption rate of micronutrients, including Se, and may act as a reservoir of these trace elements for crops [154]. Using Se-enriched organic or green fertilizers is another option for soil amendment to produce Se-biofortified crops [155]. Well decomposed green manure facilitates Se uptake by different plant species; concurrently the decomposition of the organic matter helps to mobilize other major soil nutrients [156]. For example, when Se-enriched Stanleya pinnata was applied to carrots under optimum soil moisture, 90% of the organic Se was converted into inorganic selenate and selenite [156]. Furthermore, applications of SeMet and SeCys from organic sources increased Se uptake by plants which received Se containing inorganic fertilizers [157]. Se-enriched peat has been used as a source of organic Se in cucumber, tomato and lettuce crops [158]. The effectiveness of growing mushrooms in Se-enriched agricultural by-products has been demonstrated by Bhatia et al. [159]. However, Se bio-accessibility may be affected by the formation of indigestible Se -containing polysaccharides or Se association with chitin-containing structures in cell walls [160].
Applications of expensive and inefficient inorganic fertilizers are not necessary to fertilize crops Se [161]. Organic matter within the soil performs a significant role in immobilizing Se and making it available to plants [162]. The presence of multiple chelating compounds in organic matter helps enhance the uptake of Se by plants [152]. Adding organic materials like manures and crop residues to agricultural soils additionally increase the bioavailability of Se in the soil and may balance out Se levels [163]. For example, organic matter has been used to reduce Se toxicity in soils over-rich in Se [164,165]. Applying Se-loaded animal manures to the soil can result in higher Se concentrations in crops [166,167]. The organic biofortification of Se has become more widespread globally as Se-enhanced waste products have been recognized as a useful resource to improve the Se concentrations of agricultural soil and, indirectly, improve the nutritional status and health of humans [168]. Losses of Se through volatilization or leaching to groundwater in maize have been reduced by the application of soil organic matter concurrently with Se fertilizer [169].
Among these four valence states of Se, i.e., Se (0), Se (II), Se (IV) and Se (VI). The selenite Se (IV) and selenate Se (VI) forms of Se are primarily used in the biofortification of crops. Both selenite and selenate are water-soluble, with selenate more soluble than selenite in soil solutions. Under acidic conditions in humid areas (such as many tropical soils), selenite is the more commonly prevalent form of Se [170,171]; it forms strong bonds with metal oxides or soil organic matter has low bioavailability to plants [101]. However, Se, when mixed with Organo-Se complexes (for example Seleno-amino acid), exists as the valence stage of Se (II) which is exceptionally bioavailable. Se accumulation in plants is higher when applied by mixing with organic compounds than the inorganic forms of Se [157]. For example, seleno-amino acids are active compounds and can be applied to the soil in Se-amended organic manures in crop fields [172]. Besides, organo-Se compounds can also be released in soils through the decomposition of plant material and soil microbial matter [173].
4.2. Success of Selenium Biofortification in Food Crops Depends on a Better Understanding of the Genetic Variation of Crop Cultivars
Deep knowledge of Se biogeochemistry, its uptake mechanisms and assimilation by plants is necessary for effective Se bio-fortification of food crops. Plant-based bio-fortification is the most commonly used, most effective, natural strategy to improve deficiencies in nutritional elements like Se in the staple crops across the world [128]. Se in plants is metabolized into plant tissues [174] and varies significantly by plant species and variety [175]. Significant genetic variation in grain Se concentration has been reported for several kinds of cereals including bread wheat [176], barley [177], oat [178] and rice [179]. Similarly, significant genetic variation in seed Se concentration has been observed across legumes including chickpea, lentil [180], mung bean [181] and soybean [182]. Significant genetic variation has also been reported for Se concentrations in various leafy vegetables including onions [183], brassicas [184], and mustard [185].
4.3. Crop Breeding Assisted by Selenium Biofortification
A successful breeding methodology to produce Se biofortified food crops depends on the plant genotype, growing environment, soil physical and chemical properties and the soil microbial population. Breeding-assisted Se biofortification is a time-consuming long-term research endeavor which requires high inputs and resources. Successful implementation of a program to develop Se biofortified food crops requires identification of the most promising parent lines with broad genetic variations; long term crossing and back-crossing; and traits which are stable within a wide range of climate and soil environments [186].
4.4. Molecular and Genetic Engineering for Selenium Biofortification
Modern biotechnology-based Se biofortification facilitates Se transport, accumulation, volatilization and tolerance if toxicity arises [187]. The acceleration of Se transport in transgenic crops results in crop quality enhancement and seleno-amino acid accumulation in the edible parts of food crop which may be beneficial for human and animal health. Overexpression of genes involved in Se transportation in cell plasma membrane enhances the potentiality of Se uptake and subsequent transport within the plant [188]. For example, the overexpression of adenosine triphosphate sulfurylase (ATPS1) transgene in Arabidopsis thaliana results in higher organic Se accumulation in foliage but a decline in total Se accumulation [189]. However, the overexpression of ATPS1 accelerates the concentration of both organic and total Se in the leaves of mustard plants [190].
4.4.1. Biofortification of Selenium through Molecular Approaches
While it is well documented that inter- and intraspecific variation in Se accumulation in plants exists, the genetics of Se accumulation have received little attention. Using recombinant inbred lines, the genetic basis of selenite tolerance of A. thaliana has recently been investigated. Three quantitative trait loci (QTLs) on chromosomes 1, 3 and 5 explained 24% of the variation in Se tolerance as defined by root-length inhibition and 32% of the phenotypic variation in terms of root length [191]. Several QTLs are associated with high Se accumulation in the grains and leaves of crops have been identified [192,193]. Also, QTLs influencing grain Se concentration have been identified using populations derived from crosses between bread wheat genotypes [176], a cross between an indica and a japonica rice variety [194], and an association mapping panel of rice accessions [195]. Yang et al. [196] identified four QTLs affecting grain Se concentration in a genetic mapping population derived from a cross between wild emmer wheat and tetraploid durum wheat. These occurred on chromosomes 5B, 6A and 6B. None of the causal genes underpinning the QTLs affecting Se accumulation in wheat or rice grains is currently known. Two QTLs affecting seed Se concentrations were identified using a population derived from a cross between two soybean cultivars, one of which includes a gene encoding GmSULTR2 which may facilitate Se translocation from the root to the shoot [183]. Plant cultivars with high Se concentrations in edible products can be used in marker-assisted breeding to transfer these high-Se QTLs to high-yielding low Se cultivars [167].
4.4.2. Biofortification of Selenium through Genetic Engineering and Transgenics
Bio-fortification by traditional and molecular breeding has been achieved in crops and specific crop components when genetic diversity is available in the primary, secondary, or tertiary gene pools of the targeted crop. When genetic diversity is unavailable, genetic transformation is the better option. This transgenic approach is advantageous in that once a useful gene has been discovered it can be used to enhance Se accumulation in multiple crops.
Transgenic plants have been engineered with greater Se tolerance, Se accumulation or reduced Se volatilization than their non-transgenic counterparts [197]. An example of this is the SeCys methyltransferase gene of Astragalus bisulcatus (two-grooved poison vetch) which was introduced into Arabidopsis thaliana to overexpress Se-methyl SeCys and g-glutamyl methyl SeCys in plant shoots [198], which resulted in an increased accumulation of Se within the plant. Other genes have been successfully targeted by genetic engineering in the last decade, with positive outcomes for Se bio-fortification. For instance, the overexpression of the Se binding protein gene SBP1 in Arabidopsis thaliana enhanced the resistance of the plant to selenite via a GSH-dependent mechanism [199]. Similarly, the loss-of-function mutations in the gene APX1 coding for a cytosolic ascorbate peroxidase enzyme or the overexpression of the ethylene response factor ERF96 improved Se tolerance and accumulation in A. thaliana [200,201]. In the Se accumulator Brassica juncea, a novel SeCys methyltransferase enzyme has been identified, which is capable of methylating both homocysteine and SeCys substrates [202]. The overexpression of this enzyme in tobacco plants increased the total Se and MeSeCys accumulation in the plants (Chen et al. 2019). Another potential gene target is the NRT1.1B transporter, a member of the rice peptide transporter (PTR) family involved in nitrate transport; its overexpression in rice leads to higher SeMet accumulation in rice grain [203]. Additionally, the overexpression of genes encoding SeCys lyases or the overexpression of the At Se building protein (SBP) 1 increases a plant’s tolerance to selenate or selenite [188]. In addition to the assimilation enzymes, sulphate transporters may be potential targets of genetic engineering. When combined with functional genomics this gene technology could significantly contribute to future Se bio-fortification research [204]. The advancement of modern molecular tools and analytical techniques has progressed research which concentrates on Se bio-fortification to design future and more effective crop breeding strategies. Analytical methods include synchrotron X-ray fluorescence and X-ray absorption near edge structure spectroscopies, while molecular technologies benefit from high-speed and low-cost next-generation sequencing (NGS), and encompass oligo-directed mutagenesis, reverse breeding, RNA-directed gene-methylation, and gene editing [205].
4.5. Selenium Biofortification of Crops by Beneficial Microorganisms
Bio-fortification enhances the nutrient concentration of crops and can be undertaken by fertilizers, conventional plant breeding or by biotechnological approaches or by beneficial microbes. Bio-fortification of food crops increases their nutritional concentrations where the essential elements are in inadequate supply in the soil. Se bio-fortification can be performed successfully in Se-deficient soils by selecting plant species that can assimilate the micronutrient in their edible parts and thus enrich the diet of animals (including humans). Moreover, the excreta of animals fed Se-fortified plant matter again enrich the soil with Se.
Many agricultural soils are deficient in Se and hence biofortification is a well-established technique to improve Se uptake in food crops. Many microbes assist plants to take up nutrients from soils, resist abiotic stress, and improve their growth and yield. These beneficial microbes may also be exploited for bio-fortification. Dark septate fungi, mycorrhizal and root endophytic fungi, plant growth-promoting rhizobacteria (PGPR), etc. are used for Se bio-fortification [206].
Higher concentrations of bacteria colonizing a seleniferous area resulted in better root formation and consequently greater plant-Se uptake in Brassica juncea [207]. The bacteria population enhanced the bioavailability of Se; however, this may be species-dependent. Microbial biomass carbon, microbial biomass nitrogen, adenylate energy charge and metabolic quotient did not influence plant-Se uptake from soil [208].
The symbiotic relationship between food crops and beneficial microorganisms (BMOs) improves crop growth, micronutrient uptake and resistance to different stressors. Endomycorrhizal and ectomycorrhizal fungi, and root endophytic fungi (REF) are well-known BMOs commonly used as biofilm biofertilizers. Arbuscular mycorrhizal fungi (AMF) settle in roots of angiosperms while ectomycorrhizal fungi are found in gymnosperms. REFs generally have a vast range of host plants.
Se bio-fortification by microbes is not new. In soils rich in Se, rhizosphere bacteria helped in the formation of root hairs and Se uptake as selenite in mustard (Brassica juncea) was confirmed by de Souza et al. [208]. In 2015 Yasin et al. [207] reported that various combinations of BMOs increase Se concentration in Brassica juncea growing in a seleniferous area. Different bacteria may differently influence plant growth by mediating different rates of Se uptake from soil. Considering all the microflora which surround the roots of food crops grown in a Se-rich soil, important information may be obtained regarding the suitable bio-fortification approach to be taken. In some Se-enriched soils (e.g., 20 μg Se/g soil) the presence of soil microbes, such as adenosine triphosphate (ATP), adenylate energy charge (AEC), ATP-to-microbial biomass C and metabolic quotient, had a limited effect on Se uptake [208].
4.5.1. Arbuscular Mycorrhizal Fungi (AMF) and Root Endophytic Fungi
Arbuscular mycorrhizal fungi are important in Se bio-fortification as they enhance the uptake of nutrients in host plants. The genomes of AMF encode high-affinity inorganic phosphate transporters [209]. There is a competition between the uptake of phosphate and Se accumulation that results in a decrease in the accumulation-translocation coefficients of Se. This also decreases the Se content in wheat (especially in leaves, stems, spikes and roots) when phosphate fertilizers are applied to Se-enriched crop fields [210]. Some species of Astragalus, namely, A. racemosus and A. bisulacatus are hyper accumulators of Se, whereas, A. glycyphyllos and A. drummondii are non-accumulators. Sulfur deficiency in these species increased Se accumulation; as well, an increase in Se supply enhanced the accumulation of sulfate in shoot and root tissue [211]. Table 5 and Table 6 illustrate AMFs and REFs that are used in Se bio-fortification in food crops.
Table 5.
Arbuscular mycorrhizal fungi used in Se bio-fortification.
Table 6.
Root endophytic fungi used in Se bio-fortification.
Arbuscular mycorrhizal fungi increase Se uptake in plants. When wheat seedlings were inoculated with AMF, namely Glomus versiform and Funneliformis mosseaein in the hydroponic culture medium, the uptake of selenate and enhanced the accumulation of selenite; however, there was no effect on the uptake of SeMet [217,218]. Higher accumulation of Se was observed by up-regulation of three genes that encode sulfate transporters, i.e., TaSultr1:1, TaSultr1:3, and TaSultr2:1, in the roots of mycorrhiza. In the roots of G. versiform and F. mosseae, TaSultr1:1 gene expression was up-regulated 2.18-fold and 2.12-fold, respectively. Allium sativum L. contains diallyl disulfide, which inhibits metastasis of many cancer cells (colon, lung, and skin cancer cells) and WEHI-3 leukaemia cells [219].
4.5.2. Plant Growth-Promoting Rhizobacteria (PGPR)
The interface between roots and soil is the rhizosphere, where critical interactions between beneficial microorganisms and plant roots occur. In 1980, Kloepper demonstrated the beneficial role of PGPR [220]. PGPR promote (directly or indirectly) plant growth in the rhizosphere. Mechanisms like bio-fertilization and photo-stimulation promote plant growth while reducing the use of inorganic chemical fertilizers, combating abiotic stress and reducing plant disease. The WHO recognizes some micronutrients which are essential for the proper functioning of the human body, including Se, iron (Fe) and zinc (Zn). PGPR have a significant role in the bio-fortification of Se, Fe and Zn [221].
Plant roots absorb Se in the forms of selenate, selenite or SeCys and SeMet, but cannot directly take up metal selenides or elemental Se. PGPR increase the level of Se uptake in plants, improving both animal and human health. Mustard (Brassica juncea) utilizes PGPR to increase Se accumulation and volatilization. Here, dimethyl selenide is the form of most volatilized Se; it is five- to seven-hundred times less toxic than elemental Se. This makes PGPR one of the most desirable tools for Se bio-fortification to improve Se concentrations in the human food chain. Endophytic bacteria identified and isolated from Se-supplemented wheat improve plant growth, bio-fortification and serve as biocontrol agents in wheat cultivation in ash-derived volcanic andisol in Southern Chile [222].
PGPR have well-established roles in enhancing Se uptake, crop productivity and stress tolerance [223]. Some PGPRB solubilizes inorganic phosphate in soils. Phosphate solubilizing bacteria are exploited for Se bio-fortification Table 7. Often stable complexes are formed by Se with clay minerals and/or strongly absorbed with aluminium (Al), iron (Fe), manganese (Mn), and hence have low bioavailability to most plants [224]. However, selenite and selenate are widely available to plants. When selenate and selenite (e.g., as Se fertilizer) are applied to agricultural land, they are reduced very fast and converted into forms which make them unavailable to plants (i.e., Se–metal ion complexes) which leads to low Se bioavailability (<10% only) for food crops. The re-solubility of Se in soils is very important for bio-fortification. PGPR function as Se-solubilizing agents [225]. Trivedi et al. [226] identified several endophytic seleno-bacteria from Ricinus communis; further identification at the molecular level showed that these were Alcaligenes faecalis, Paraburkholderia megapolitana, and Stenotrophomonas maltophilia. P. megapolitana play an important role in facilitating the growth of soybean (Glycine max) under drought conditions while increasing Se bio-fortification. The synergistic action between Se bio-fortification and increased drought tolerance is crucial for the cultivation of crops in arid and semi-arid regions where Se is limited.
Table 7.
Plant growth-promoting rhizobacteria (PGPR) which facilitate the uptake of Se.
5. Mechanisms to Uptake and Accumulate Selenium in Plants
5.1. Uptake Mechanisms
Selenium is present in Nature in both organic and inorganic forms. Organic forms are mainly SeMet and SeCys; while inorganic forms include elemental Se, selenide (Se2−), selenite (SeO2−3SeO32−) and selenate (SeO2−4SeO42−) [229]. The uptake, distribution and translocation of Se within plants are determined by plant translocation, the activity of membrane transporters, the presence of other substances, soil physical conditions (e.g., salinity and soil pH), the form and concentration of Se, and the plant species and phase of its development [230]. In comparison to selenite, selenate is more water-soluble and more common in agricultural soils [231]. In acidic soils, Se mainly exists as selenite, while in alkaline soils selenate is the dominant form of Se. Both these forms are metabolized to seleno-compounds but differ in terms of absorption and mobility within the plant [232]. The rates of plant transpiration and xylem loading determine the translocation of Se to shoot tissue [230]. Selenate is more mobile than SeMet, which is more mobile than either selenite or SeCys in wheat and canola [157]. The transport of selenate in plants is via sulfate channels and transporters [191], while phosphate transport mechanisms are responsible for the transport of selenite [232].
The existence of transporters for sulfate and selenate depends on the soil and plant nutritional status [233]. Constitutive active sulfate transporters showed less selectivity for sulfate over selenate than inducible sulfate transporters. Selectivity for Se decreases when there is a higher concentration of external sulfate [233]. The earlier study observed that selenate (a toxic analogue of sulfate) transport inside the plant of Arabidopsis thaliana via sulfate transporters (SULTR1 and SULTR1;2) [234], while elsewhere it was found that the uptake of selenate, sulfate transporter gene SULTR1;2 plays a predominant role in plant roots [235]. Arabidopsis thaliana was found resistant to selenate with lack of sulfate transporter gene SULTR1;2, but no resistance with another sulfate transporter gene SULTR1 [234,235,236]. There was an increase in Se uptake with the starvation of sulfur in Triticum aestivum [233]. Passive diffusion has been observed as a mechanism for selenite uptake [237], while Li et al. [232] found an active transport mechanism for selenite uptake. Terry et al. [197] observed selenite uptake without the involvement of membrane transporters. Li et al. [232] stated that in phosphorous deficiency the uptake of selenite was enhanced. This finding not only supports earlier studies indicating low uptake of selenite with increasing phosphate concentration but also indicates the role of phosphate transporters for selenite uptake.
5.2. Accumulation Mechanisms
During seedling growth, young leaves contain relatively high concentrations of Se [238]. The accumulation of Se usually occurs in the vacuoles of the plant cells [239,240], and through sulfate transporters, Se may be effluxed from the vacuole [241]. Depending on the concentration of Se inside the cell, plants are classified as non-accumulators, secondary-accumulators or hyper-accumulators [242]. Plants which thrive in regions which are rich in Se (i.e., >1000 mg Se/kg of soil) are thus hyper-accumulators of Se. Examples of hyper-accumulators include Xylorhiza, Neptunia, Conopsis, Astragalus and Stanleya species. Plants that accumulate between 100 and 1000 mg Se/kg soil with no toxicity symptoms are secondary-accumulators; these include Medicago sativa, Camelina, Aster, Helianthus, Broccoli, Brassica napus and Brassica juncea varieties. Plants which accumulate less than 100 mg Se/kg of soil are non-accumulators: in Se-enriched soils, these plants would not survive. Non-accumulator plants volatilize Se as dimethyl selenide (DMSe) and show retarded growth [242]. Non-accumulator plants sequester Se in vacuoles when enriched with Se [240].
6. Prospects of Selenium Biofortification
The importance of Se has long been recognized for human and animal life [4,14]. As a trace element, Se is critical for human health and Se deficiency impedes metabolism [243]. Generally, crops biofortified with Se are enriched with many beneficial phytochemicals, such as minerals and antioxidants [244] Figure 3. The response of plants to Se biofortification differs by crop, for example, different responses have been observed in lettuce [47], tomato [245], broccoli [246], cucumber [247] and carrot crops [248].
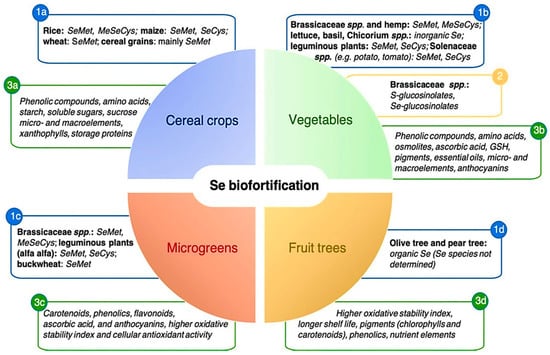
Figure 3.
Se biofortification to improve human plant-foods [244]. Note: (1a), (1b), (1c) and (1d) indicate that SeMet, MeSeCys, SeCys contents are available in cereals, vegetables, fruits, and microgreens; (2) indicate that S-glucosinolate and Se- glucosinolate content are available only in vegetables and (3a), (3b), (3c) and (3d) indicate that phenolic compounds, amino acids, starch, soluble sugars, sucrose, micro-and macro-elements, antioxidants, storage protein etc. are available in cereals, vegetables, fruits, and microgreens.
Selenium is closely bound to clay particles and is found in greater abundance in clay soils than in more sandy soils. Se toxicity is a result of the isomorphic substitution of sulfur for Se, which ultimately adversely affects plant growth. In general, however, food products are low in Se and benefit from Se enhancement, of which biofortification is very important [249]. Generally, vegetables have good prospects for Se biofortification [250] due to their inherent tendency to accumulate Se [251]. Biofortification has also been observed to occur naturally in Brazil, where Amazonian fruits contain sufficient Se to meet daily human requirements (0.03 to 512 μg/g) [252]. There are many costs associated with Se biofortification which impede the adoption of this process; efforts are being made to reduce the costs of biofortification. One application is to mix a mile (1%) Se solution with fertilizer before applying to the soil [253].
Several factors influence the success of Se biofortification: two major considerations are the method of Se fertilization (i.e., in-soil vs. foliar application) and the crop species. Generally, Se has a smaller role in the growth and development of taller plants [254]. Se is assimilated to Se-amino acids through sulfate channels [255]. Malformed proteins which cause toxicity may be produced as a result of excess Se [256], however, lower Se concentrations are beneficial, particularly for hyper-accumulator plant species [6]. The beneficial effects of Se biofortification depend on the rate of Se application, the accumulation rate of the plant, and its capacity for Se tolerance [48]. Malik et al. [257] reported reductions in electrolytic leakage which promoted cell antioxidative processes [200]. In salt-affected areas, a beneficial role of Se in food crops is observed which may be a result of improved plant growth and yield parameters [200], improvement in net photosynthesis and water content [258], and the stimulation of antioxidants. Under heat or drought stress, Se plays a critical role in maintaining cereal grain yields [259,260]. Se also protects plants from insect-pest and fungal attacks [261,262]. Figure 4 illustrates many ways in which Se biofortification improves human and animal health [131].
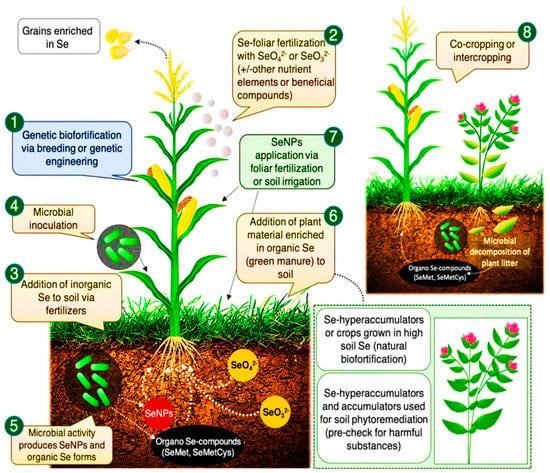
Figure 4.
Se biofortification promotes crop yields and quality parameters. Se-biofortification approaches include (1) genetic tools, (2) through foliar application, (3) soil amendment, (4) agronomic biofortification, (5) broadcasting into soils, (6) green manure with Se, enriched growth and development of plants, (7) nano-sized biofortification to leaves or soil, and (8) intercropping with Se- hyper-accumulator plants. Adapted and modified from Schiavon et al. [132].
7. Conclusions
Se is an essential trace element for all living organisms. Se deficiency causes severe ill health effects in humans and animals. It is estimated that about one billion people globally suffer from Se deficiency. Se is linked in humans to the effective performance of the immune, endocrine and reproductive systems as well as brain function. Excessive Se uptake can contribute to hypotension, tachycardia, tremors, muscle contractions, hair loss, and skin and nail lesions. Therefore, a balanced daily dose of Se should be maintained. The recommended dosage of Se varies with age, gender, pregnancy, lactation and also geographical location and diet. Se is accrued in the human body through the consumption of animal and plant foods. To meet Se deficiency and improve human health where Se is inadequate, Se concentrations in plants should be quantified and balanced. Se-biofortification in combination with agronomic, breeding, molecular, biotechnology, and genetic engineering management approaches are effective strategies for Se enrichment in human plants foods.
Author Contributions
Conceptualization, A.H., S.M. and S.S. (Sukamal Sarkar); writing—original draft preparation, A.H., S.M., S.S. (Sukamal Sarkar), Z.A., H.V., S.G., M.M., R.B., P.K., P.B. and S.S. (Saikat Saha), writing—review and editing, M.S., M.B., T.I. and A.M.L., funding acquisition, M.S., M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This is an international collaborative work. Financial support for APC of the review was funded by an S-grant from the Ministry of Education, Youth and Sports of the Czech Republic.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
The study did not involve humans or animals.
Data Availability Statement
Not applicable.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [PubMed]
- Galan-Chilet, I.; Tellez-Plaza, M.; Guallar, E.; De Marco, G.; Lopez-Izquierdo, R.; Gonzalez-Manzano, I.; Carmen Tormos, M.; Martin-Nuñez, G.M.; Rojo-Martinez, G.; Saez, G.T.; et al. Plasma selenium levels and oxidative stress biomarkers: A gene–environment interaction population-based study. Free Radic. Biol. Med. 2014, 74, 229–236. [Google Scholar] [CrossRef]
- Duntas, L.H.; Benvenga, S. Selenium: An element for life. Endocrine 2015, 48, 756–775. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błazejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef]
- Nothstein, A.K.; Eiche, E.; Riemann, M.; Nick, P.; Winkel, L.H.E.; Göttlicher, J.; Steininger, R.; Brendel, R.; Brasch, M.V.; Konrad, G.; et al. Tracking se assimilation and speciation through the rice plant–nutrient competition, toxicity and distribution. PLoS ONE 2016, 26, e0152081. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Le Duc, D.L. Phytoremediation of selenium using transgenic plants. Curr. Opin. Biotechnol. 2009, 20, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Liu, G.; Yousaf, B.; Ali, M.U.; Abbas, Q.; Munir, M.A.M.; Mian, M.M. Developmental selenium exposure and health risk in daily foodstuffs: A systematic review and meta-analysis. Ecotoxicol. Environ. Saf. 2018, 149, 291–306. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. Adaptive dysfunction of selenoproteins from the perspective of the triage theory: Why modest selenium deficiency may increase risk of diseases of aging. FASEB J. 2011, 25, 1793–1814. [Google Scholar] [CrossRef] [PubMed]
- Shreenath, A.P.; Ameer, M.A.; Dooley, J. Selenium Deficiency. In Treasure Island (FL): StatPearls Publishing; 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482260/ (accessed on 30 January 2021).
- Longchamp, M.; Angeli, N.; Castrec-Rouelle, M. Selenium uptake in Zea mays supplied with selenate or selenite under hydroponic conditions. Plant Soil 2013, 362, 107–117. [Google Scholar] [CrossRef]
- Khanam, A.; Platel, K. Bioaccessibility of selenium, selenomethionine and selenocysteine from foods and influence of heat processing on the same. Food Chem. 2016, 194, 1293–1299. [Google Scholar] [CrossRef]
- Reich, H.J.; Hondal, R.J. Why nature chose selenium? ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P.; Soerensen, A.L.; DiMento, B.P.; Balcom, P.H. The global marine selenium cycle: Insights from measurements and modeling. Glob. Biogeochem. Cycles 2018, 32, 1720–1737. [Google Scholar] [CrossRef]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Selinus, O., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 375–416. [Google Scholar]
- Winkel, L.H.; Trang, P.T.K.; Lan, V.M.; Stengel, C.; Amini, M.; Ha, N.T.; Viet, P.H.; Berg, M. Arsenic pollution of groundwater in Vietnam exacerbated by deep aquifer exploitation for more than a century. Proc. Natl. Acad. Sci. USA 2011, 108, 1246–1251. [Google Scholar] [CrossRef]
- Shahid, M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar]
- Söderlund, M.; Virkanen, J.; Holgersson, S.; Lehto, J. Sorption and speciation of selenium in boreal forest soil. J. Environ. Radioact. 2016, 164, 220–231. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiang, Y.; Zhou, Y.; Yang, Y.; Zhang, J.; Huang, H.; Shang, C.; Luo, L.; Gao, J.; Tang, L. Selenium contamination, consequences and remediation techniques in water and soils: A review. Environ. Res. 2018, 164, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Tomza-Marciniak, A.; Bąkowska, M.; Pilarczyk, B.; Semeniuk, M.; Hendzel, D.; Udała, J.; Balicka-Ramisz, A.; Tylkowska, A. Concentration of selenium in the soil and in selected organs of roe deer (Capreolus capreolus) from the Greater Poland Voivodeship. Acta Sci. Pol. Zootech. 2010, 9, 251–260. [Google Scholar]
- Chilimba, A.D.; Young, S.D.; Black, C.R.; Meacham, M.C.; Lammel, J.; Broadley, M.R. Agronomic biofortification of maize with selenium (Se) in Malawi. Field Crop. Res. 2012, 125, 118–128. [Google Scholar] [CrossRef]
- Lopes, G.; Ávila, F.W.; Guilherme, L.R.G. Selenium behavior in the soil environment and its implication for human health. Ciência Agrotecnologia 2017, 41, 605–615. [Google Scholar] [CrossRef]
- Stroud, J.; Broadley, M.; Foot, I.; Fairweather-Tait, S.; Hart, D.; Hurst, R.; Knott, P.; Mowat, H.; Norman, K.; Scott, P. Soil factors affecting selenium concentration in wheat grain and the fate and speciation of Se fertilisers applied to soil. Plant Soil 2010, 332, 19–30. [Google Scholar] [CrossRef]
- Paikaray, S. Origin, mobilization and distribution of selenium in a soil/water/air system: A global perspective with special reference to the Indian scenario. Clean 2016, 44, 474–487. [Google Scholar] [CrossRef]
- WHO. Selenium in Drinking-Water. In Background Document for Preparation of WHO Guidelines for Drinking-Water Quality; World Health Organization (WHO/SDE/WSH/03.04/13): Geneva, Switzerland, 2003. [Google Scholar]
- Oropeza-Moe, M.; Wisløff, H.; Bernhoft, A. Selenium deficiency associated porcine and human cardiomyopathies. J. Trace Elem. Med. Biol. 2015, 31, 148–156. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Sies, H. Selenium homeostasis and antioxidant selenoproteins in brain: Implications for disorders in the central nervous system. Arch. Biochem. Biophys. 2013, 536, 152–157. [Google Scholar] [CrossRef]
- Pillai, R.; Uyehara-Lock, J.H.; Bellinger, F.P. Selenium and selenoprotein function in brain disorders. IUBMB Life 2014, 66, 229–239. [Google Scholar] [CrossRef]
- Lipinski, B. Rationale for the treatment of cancer with sodium selenite. Med. Hypotheses 2005, 64, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Okunade, K.S.; Olowoselu, O.F.; Osanyin, G.E.; John-Olabode, S.; Akanmu, S.A.; Anorlu, R.I. Selenium deficiency and pregnancy outcome in pregnant women with HIV in Lagos, Nigeria. Int. J. Gynecol. Obstet. 2018, 142, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and thyroid disease: From pathophysiology to treatment. Int. J. Endocrinol. 2017, 1297658. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.H.; Clish, C.B.; Gladyshev, V.N. Selenium deficiency is associated with pro-longevity mechanisms. Cell Rep. 2019, 27, 2785–2797. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef]
- Underwood, E.J.; Suttle, N.F. The mineral nutrition of livestock. In Selenium; Underwood, E.J., Suttle, N.F., Eds.; CABI Publishing: New York, NY, USA, 1999; pp. 421–475. [Google Scholar]
- Whanger, P.D.; Weswig, P.H.; Oldfield, J.E.; Cheeke, P.R.; Muth, O.H. Factors influencing selenium and white muscle disease: Forage types, salts, amino acids and dimethyl sulfoxide. Nutr. Rep. Int. 1972, 6, 21–37. [Google Scholar]
- Turner, R.J.; Finch, J.M. Selenium and the immune response. Proc. Nutr. Soc. 1991, 50, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr.; Combs, S.B. The Role of Selenium in Nutrition; Academic Press: Cambridge, MA, USA, 1986; p. 532. [Google Scholar]
- Andrews, E.D.; Hartley, W.J.; Grant, A.B. Selenium- responsive diseases of animals in New Zealand. New Zealand Vet. J. 1968, 16, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total. Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef]
- Vinceti, M.; Mandrioli, J.; Borella, P.; Michalke, B.; Tsatsakis, A.; Finkelstein, Y. Selenium neurotoxicity in humans: Bridging laboratory and epidemiologic studies. Toxicol. Lett. 2014, 230, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Franke, K.W. A new toxicant occurring naturally in certain samples of plant foodstuffs. 1. Results obtained in preliminary feeding trials. J. Nutr. 1934, 8, 597–609. [Google Scholar] [CrossRef]
- IMFNB. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academy Press: Cambridge, MA, USA, 2000; pp. 1–20. [Google Scholar]
- Silva Junior, E.C.; Wadt, L.H.O.; Silva, K.E.; Lima, R.M.B.; Batista, K.D.; Guedes, M.C.; Carvalho, G.S.; Carvalho, T.S.; Reis, A.R.; Lopes, G.; et al. Natural variation of selenium in Brazil nuts and soils from the Amazon region. Chemosphere 2017, 188, 650–658. [Google Scholar] [CrossRef]
- Post, M.; Lubiński, W.; Lubiński, J.; Krzystolik, K.; Baszuk, P.; Muszyńska, M.; Marciniak, W. Serum selenium levels are associated with age-related cataract. Ann. Agric. Environ. Med. 2018, 25, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Pophaly, S.D.; Singh, P.; Kumar, H.; Tomar, S.K.; Singh, R. Selenium enrichment of lactic acid bacteria and bifidobacteria: A functional food perspective. Trends Food Sci. Tech. 2014, 39, 135–145. [Google Scholar] [CrossRef]
- USDA-ARS. USDA National Nutrient Database for Standard Reference; Release 25; USDA-ARS: Washington, DC, USA, 2002. Available online: http://www.nal.usda.gov/fnic/selenium (accessed on 30 January 2021).
- WHO. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks. 2009. Available online: http://www.who.int/healthinfo/global_burden_disease/GlobalHealthRisks_reportannex.pdf (accessed on 28 January 2021).
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant. Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef]
- Ríos, J.J.; Blasco, B.; Leyva, R.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Nutritional balance changes in lettuce plant grown under different doses and forms of selenium. J. Plant. Nutr. 2013, 36, 1344–1354. [Google Scholar] [CrossRef]
- Reilly, C. Selenium in Food and Health; Blackie: London, UK, 1996. [Google Scholar]
- Shrift, A. Aspects of selenium metabolism in higher plants. Ann. Rev. Plant Physiol. 1969, 20, 475–494. [Google Scholar] [CrossRef]
- Lyons, G.H.; Stangoulis, J.C.R.; Graham, R.D. High-selenium wheat: Biofortification for better health. Nutr. Res. Rev. 2003, 16, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wang, P.; Yamaji, N.; Ma, J.F. Plant nutrition for human nutrition: Hints from rice research and future perspectives. Mol. Plant 2020, 13, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Lyons, M.P.; Papazyan, T.T.; Surai, P.F. Selenium in food chain and animal nutrition: Lessons from nature. Asian-Australas. J. Anim. Sci. 2007, 20, 1135–1155. [Google Scholar] [CrossRef]
- Kipp, A.P.; Strohm, D.; Brigelius-Flohé, R.; Schomburg, L.; Bechthold, A.; Leschik-Bonnet, E.; Heseker, H.; DGE, G.N.S. Revised reference values for selenium intake. J. Trace Elem. Med. Biol. 2015, 32, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Tamari, Y.; Kim, E.S. Longitudinal study of the dietary selenium intake of exclusively breast-fed infants during early lactation in Korea and Japan. J. Trace Elem. Med. Biol. 1999, 13, 129–133. [Google Scholar] [CrossRef]
- Hsueh, Y.M.; Su, C.T.; Shiue, H.S.; Chen, W.J.; Pu, Y.S.; Lin, Y.C.; Tsai, C.S.; Huang, C.Y. Levels of plasma selenium and urinary total arsenic interact to affect the risk for prostate cancer. Food Chem. Toxicol. 2017, 107, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Bampidis, V.; Azimonti, G.; Bastos, M.D.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Durjava, M.K.; López-Alonso, M.; Puente, S.L.; Marcon, F.; et al. Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae CNCM I-3060 (selenised yeast inactivated) for all animal species. EFSA J. 2018, 16, e05386. [Google Scholar]
- Fraczek, A.; Pasternak, K. Selenium in medicine and treatment. J. Elem. 2013, 18, 145–163. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Kurek, E. Binding and conversion of selenium in Candida utilis ATCC 9950 yeasts in bioreactor culture. Molecules 2017, 22, 352. [Google Scholar] [CrossRef] [PubMed]
- Zwolak, I.; Zaporowska, H. Selenium interactions and toxicity: A review. Selenium interactions and toxicity. Cell Biol. Toxicol. 2012, 28, 31–46. [Google Scholar] [CrossRef]
- Stoffaneller, R.; Morse, N.L. A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Selenium: Significance, and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The argument for increasing Se intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef]
- Combs, G.F.; Lü, J. Selenium as a cancer preventive agent. In Selenium; Hatfield, D.L., Berry, M.J., Gladyshev, V.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar] [CrossRef]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-dependent antioxidant enzymes: Actions and properties of selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhang, B.; Xu, R.; Wang, Y.; Ding, X.; Li, P. Antioxidant activity in vitro of the selenium-contained protein from the Se-enriched Bifidobacterium animalis 01. Anaerobe 2010, 16, 380–386. [Google Scholar] [CrossRef]
- Elahi, M.M.; Kong, Y.X.; Matata, B.M. Oxidative stress as a mediator of cardiovascular disease. Oxidative Med. Cell. Longev. 2009, 5, 259–269. [Google Scholar] [CrossRef]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). Jama 2009, 301, 39–51. [Google Scholar] [CrossRef]
- Klein, E.A.; Thompson, I.M.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the risk of prostate cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). Jama 2011, 306, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Dennert, G.; Horneber, M. Selenium for alleviating the side effects of chemotherapy, radiotherapy and surgery in cancer patients. Cochrane Database Syst. Rev. 2006, 3, CD005037. [Google Scholar] [CrossRef] [PubMed]
- Rees, K.; Hartley, L.; Day, C.; Flowers, N.; Clarke, A.; Stranges, S. Selenium supplementation for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Farrokhian, A.; Bahmani, F.; Taghizadeh, M.; Mirhashemi, S.M.; Aarabi, M.H.; Raygan, F.; Aghadavod, E.; Asemi, Z. Selenium supplementation affects insulin resistance and serum hs-CRP in patients with type 2 diabetes and coronary heart disease. Horm. Metab. Res. 2016, 48, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, R.G.; Tufik, S.; Andersen, M.L. Benefits of selenium in the treatment of depression and sleep disorders. Sleep Breath. 2019, 23, 933–934. [Google Scholar] [CrossRef]
- Triggiani, V.; Tafaro, E.; Giagulli, V.A.; Sabbà, C.; Resta, F.; Licchelli, B.; Guastamacchia, E. Role of iodine, selenium and other micronutrients in thyroid function and disorders. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 277–294. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium, selenoproteins and the thyroid gland: Interactions in health and disease. Nat. Rev. Endocrinol. 2012, 8, 160–171. [Google Scholar] [CrossRef]
- Köhrle, J. Selenium and the thyroid. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 392–401. [Google Scholar] [CrossRef]
- Drutel, A.; Archambeaud, F.; Caron, P. Selenium and the thyroid gland: More good news for clinicians. Clin. Endocrinol. 2013, 78, 155–164. [Google Scholar] [CrossRef]
- Stuss, M.; Michalska-Kasiczak, M.; Sewerynek, E. The role of selenium in thyroid gland pathophysiology. Endokrynol. Pol. 2017, 68, 440–465. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Riaz, M.; Mehmood, K.T. Selenium in human health and disease: A review. J. Postgrad. Med Inst. 2012, 26, 120–133. [Google Scholar]
- Brigelius-Flohé, R. Selenium in human health and disease: An overview. In Selenium; Springer: Berlin/Heidelberg, Germany, 2018; pp. 3–26. [Google Scholar]
- Shaheen, S.O.; Newson, R.B.; Rayman, M.P.; Wong, A.P.; Tumilty, M.K.; Phillips, J.M.; Potts, J.F.; Kelly, F.J.; White, P.T.; Burney, P.G. Randomised, double blind, placebo-controlled trial of selenium supplementation in adult asthma. Thorax 2007, 62, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Norton, G.; Duan, G.-L.; Lei, M.; Zhu, Y.G.; Meharg, A.; Price, A. Identification of quantitative trait loci for rice grain element composition on an arsenic impacted soil: Influence of flowering time on genetic loci. Ann. Appl. Biol. 2012, 161, 46–56. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.J.; Breward, N.; et al. Biofortification of UK food crops with selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef]
- Broadley, M.R.; Alcock, J.; Alford, J.; Cartwright, P.; Foot, I.; Fairweather-Tait, S.J.; Hart, D.J.; Hurst, R.; Knott, P.; McGrath, S.P.; et al. Selenium biofortification of high-yielding winter wheat (Triticum aestivum L.) by liquid or granular Se fertilisation. Plant Soil 2010, 332, 5–18. [Google Scholar] [CrossRef]
- Aro, A.; Alfthan, G.; Varo, P. Effects of supplementation of fertilizers on human selenium status in Finland. Analyst 1995, 120, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Eurola, M.; Ekholm, P.; Ylinen, M.; Koivistoinen, P.; Varo, P. Effects of Selenium Fertilization on the Selenium Content of Selected Finnish Fruits and Vegetables. Acta Agric. Scand. 1989, 39, 345–350. [Google Scholar] [CrossRef]
- Mäkelä, A.L.; Wang, W.C.; Hämäläinen, M.; Näntö, V.; Laihonen, P.; Kotilainen, H.; Meng, L.X.; Mäkelä, P. Environmental effects of nationwide selenium fertilization in Finland. Biol. Trace Elem. Res. 1995, 47, 289–298. [Google Scholar] [CrossRef]
- Varo, P.; Alfthan, G.; Huttunen, J.K.; Aro, A. Nationwide selenium supplementation in Finland: Effects on diet, blood and tissue levels, and health. In Selenium in Biology and Human Health; Burk, R.F., Ed.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 197–218. [Google Scholar]
- Hart, D.; Fairweather-Tait, S.; Broadley, M.; Dickinson, S.; Foot, I.; Knott, P.; McGrath, S.; Mowat, H.; Norman, K.; Scott, P. Selenium concentration and speciation in biofortified flour and bread: Retention of selenium during grain biofortification, processing and production of Se-enriched food. Food Chem. 2011, 126, 1771–1778. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium in higher plants: Physiological role, antioxidant metabolism and abiotic stress tolerance. J. Plant. Sci. 2010, 5, 354–375. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Hladun, K.R.; Parker, D.R.; Tran, K.D.; Trumble, J.T. Effects of selenium accumulation on phytotoxicity, herbivory, and pollination ecology in radish (Raphanus sativus L.). Environ. Pollut. 2013, 172, 70–75. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; Proietti, P.; Nasini, L.; Del Buono, D.; Tedeschini, E.; Businelli, D. Increase in the selenium content of extra virgin olive oil: Quantitative and qualitative implications. Grasas Aceites 2014, 65, e025. [Google Scholar] [CrossRef]
- Pezzarossa, B.; Remorini, D.; Gentile, M.L.; Massai, R. Effects of foliar and fruit addition of sodium selenate on selenium accumulation and fruit quality. J. Sci. Food Agric. 2012, 92, 781–786. [Google Scholar] [CrossRef]
- Hsu, F.C.; Wirtz, M.; Heppel, S.C.; Bogs, J.; Krämer, U.; Khan, M.S.; Bub, A.; Hell, R.; Rausch, T. Generation of Se-fortified broccoli as functional food: Impact of Se fertilization on S metabolism. Plant Cell Environ. 2011, 34, 192–207. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Ghosh, D.; Chandra Santra, S. Evaluation of effectiveness of seed priming with selenium in rice during germination under arsenic stress. Plant. Physiol. Biochem. 2016, 109, 571–578. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Effect of selenium induced seed priming on arsenic accumulation in rice plant and subsequent transmission in human food chain. Ecotoxicol. Environ. Saf. 2018, 152, 67–77. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se mitigates As-induced phytotoxicity in rice seedlings by enhancing essential micronutrient uptake and translocation and reducing As translocation. Environ. Sci. Pollut. Res. 2018, 25, 26978. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se alleviate As induced phytotoxicity during germination and seedling growth by restricting As translocation in rice (Oryza sativa L c.v. IET-4094). Ecotoxicol. Environ. Saf. 2017, 145, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Gao, J.; Huang, K. Effects of different selenium sources on tissue selenium concentrations, blood GSH-Px activities and plasma interleukin levels in finishing lambs. Biol. Trace Elem. Res. 2007, 116, 91–102. [Google Scholar] [CrossRef]
- Shiobara, Y.; Yoshida, T.; Suzuki, K.T. Effects of dietary selenium species on Se concentrations in hair, blood, and urine. Toxicol. Appl. Pharmacol. 1998, 152, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Swanson, C.; Patterson, B.; Levander, O.; Veillon, C.; Taylor, P.; Helzlsouer, K.; McAdam, P.; Zech, L. Human [74Se] selenomethionine metabolism: A kinetic model. Am. J. Clin. Nutr. 1991, 54, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.A.; Bobe, G.; Hunter, J.K.; Vorachek, W.R.; Stewart, W.C.; Vanegas, J.A.; Estill, C.T.; Mosher, W.D.; Pirelli, G.J. Effect of feeding selenium-fertilized alfalfa hay on performance of weaned beef calves. PLoS ONE 2013, 8, e58188. [Google Scholar] [CrossRef]
- Benes, S.E.; Robinson, P.H.; Cun, G.S. Depletion of selenium in blood, liver and muscle from beef heifers previously fed forages containing high levels of selenium. Sci. Total Environ. 2015, 536, 603–608. [Google Scholar] [CrossRef]
- Quijano, M.; Moreno, P.; Gutiérrez, A.M.; Pérez-Conde, M.C.; Camara, C. Selenium speciation in animal tissues after enzymatic digestion by high-performance liquid chromatography coupled to inductively coupled plasma mass spectrometry. J. Mass Spectrom. 2000, 35, 878–884. [Google Scholar] [CrossRef]
- Ralston, N.V.; Ralston, C.R.; Blackwell, J.L.; Raymond, L.J. Dietary and tissue selenium in relation to methylmercury toxicity. Neurotoxicology 2008, 29, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr. Selenium in global food systems. Brit. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef]
- Lin, Z.Q. Uptake and accumulation of selenium in plants in relation to chemical soeciationa nd biotransformation. In Development and Uses of Biofortified Agricultural Products; Banuelos, G., Lin, Z.-Q., Eds.; CRC: Boca Raton, FL, USA, 2008; pp. 45–56. [Google Scholar]
- Lyons, G.H.; Genc, Y.; Graham, R. Biofortication in the food chain, and use of selenium and phyto-compounds in risk reduction and control of prostate cancer. In Development and Uses of Biofortified Agricultural Products; Banuelos, G., Lin, Z.-Q., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 17–44. [Google Scholar]
- ODS. Selenium: Dietary Supplement Fact Sheet. Health Information. US Department of Health and Human Services, National Institutes of Health; Office of Dietary Supplements: Washington, DC, USA, 2016.
- Saltzman, A.; Birol, E.; Bouis, H.E.; Boy, E.; De Moura, F.F.; Islam, Y.; Pfeiffer, W.H. Biofortification: Progress toward a more nourishing future. Glob. Food Secur. 2013, 2, 9–17. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, N.; Li, S.; Li, L.; Su, H.; Liu, C. Distribution and transport of selenium in Yutangba, China: Impact of human activities. Sci. Total Environ. 2008, 392, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.N.; Lombi, E.; Sun, G.X.; Scheckel, K.; Zhu, Y.G.; Feng, X.; Zhu, J.; Carey, A.M.; Adomako, E.; Lawgali, Y.; et al. Selenium characterization in the global rice supply chain. Environ. Sci. Technol. 2009, 43, 6024–6030. [Google Scholar] [CrossRef] [PubMed]
- Thavarajah, P.; Sarker, A.; Materne, M.; Vandemark, G.; Shrestha, R.; Idrissi, O.; Hacikamiloglu, O.; Bucak, B.; Vandenberg, A. A global survey of effects of genotype and environment on selenium concentration in lentils (Lens culinaris L.): Implications for nutritional fortification strategies. Food Chem. 2011, 125, 72–76. [Google Scholar] [CrossRef]
- Winkel, L.H.; Johnson, C.A.; Lenz, M.; Grundl, T.; Leupin, O.X.; Amini, M.; Charlet, L. Environmental selenium research: From microscopic processes to global understanding. Environ. Sci. Technol. 2012, 46, 571–579. [Google Scholar] [CrossRef]
- Fordyce, F.M.; Brereton, N.; Hughes, J.; Luo, W.; Lewis, J. An initial study to assess the use of geological parent materials to predict the Se concentration in overlying soils and in five staple foodstuffs produced on them in Scotland. Sci. Total Environ. 2010, 408, 5295–5305. [Google Scholar] [CrossRef]
- Goicoechea, N.; Garmendia, I.; Fabbrin, E.G.; Bettoni, M.M.; Palop, J.A.; Sanmartín, C. Selenium fertilization and mycorrhizal technology may interfere in enhancing bioactive compounds in edible tissues of lettuces. Sci. Hortic. 2015, 195, 163–172. [Google Scholar] [CrossRef]
- Mahn, A. Modelling of the effect of selenium fertilization on the content of bioactive compounds in broccoli heads. Food Chem. 2017, 233, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Germ, M.; Stibilj, V. Selenium and plants. Acta Agric. Slov. 2008, 89. [Google Scholar] [CrossRef]
- Haug, A.; Graham, R.; Christophersen, O.; Lyons, G. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228. [Google Scholar]
- Sillanpaeae, M.; Jansson, H. Status of cadmium, lead, cobalt and selenium in soils and plants of thirty countries. FAO Soils Bull. 1992, 195. [Google Scholar]
- Ros, G.H.; van Rotterdam, A.M.D.; Bussink, D.W.; Bindraban, P.S. Selenium fertilization strategies for bio-fortification of food: An agro-ecosystem approach. Plant Soil 2016, 404, 99–112. [Google Scholar] [CrossRef]
- Wen, H.; Carignan, J. Reviews on atmospheric selenium: Emissions, speciation and fate. Atmos. Environ. 2007, 41, 7151–7165. [Google Scholar] [CrossRef]
- Chasteen, T.G.; Bentley, R. Biomethylation of selenium and tellurium: Microorganisms and plants. Chem. Rev. 2003, 103, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Ajwa, H.A.; Wu, L.; Guo, X.; Akohoue, S.; Zambrzuski, S. Selenium-induced growth reduction in Brassica land races considered for phytoremediation. Ecotoxicol. Environ. Saf. 1997, 36, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Poblaciones, M.J.; Rodrigo, S.; Santamaria, O.; Chen, Y.; McGrath, S.P. Selenium accumulation and speciation in biofortified chickpea (Cicer arietinum L.) under Mediterranean Plant Soil conditions. J. Sci. Food Agric. 2014, 94, 1101–1106. [Google Scholar] [CrossRef]
- Premarathna, L.; McLaughlin, M.J.; Kirby, J.K.; Hettiarachchi, G.M.; Stacey, S.; Chittleborough, D.J. Selenate-enriched urea granules are a highly Effective fertilizer for selenium biofortification of paddy rice grain. J. Agric. Food Chem. 2012, 60, 6037–6044. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.H.; Sambo, P.; Masi, A.; Malagoli, M. Impact of selenium fertilization on chemical composition and Antioxidant constituents of tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef]
- Ramos, S.J.; Faquin, V.; Guilherme, L.R.G.; Castro, E.M.; Avila, F.W.; Carvalho, G.S.; Bastos, C.E.A.; Oliveira, C. Selenium biofortification and antioxidant activity in lettuce plants fed with selenate and selenite. Plant Soil Environ. 2010, 56, 584–588. [Google Scholar] [CrossRef]
- Alfthan, G.; Aspila, P.; Ekholm, P.; Eurola, M.; Hartikainen, H.; Hero, H.; Hietaniemi, V.; Root, T.; Salminen, P.; Venäläinen, E.-R.; et al. Nationwide Supplementation of Sodium Selenate to Commercial Fertilizers: History and 25-Year Results from the Finnish Selenium Monitoring Programme; CAB International and Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2011. [Google Scholar]
- Winkler, J. Biofortification: Improving the nutritional quality of staple crops. In Access not Excess; Pasternak, C., Ed.; Smith-Gordon Publishing: London, UK, 2011; pp. 100–112. [Google Scholar]
- Dhillon, S.K.; Hundal, B.K.; Dhillon, K.S. Bioavailability of selenium to forage crops in a sandy loam soil amended with Se-rich plant materials. Chemosphere 2007, 66, 1734–1743. [Google Scholar] [CrossRef]
- Bruulsema, T.W.; Heffer, P.; Welch, R.M.; Cakmak, I.; Moran, K. Fertilizing Crops to Improve Human Health: A Scientific Review Volume 1 Food and Nutrition Security Editorial Committee. Better Crop. Plant Food 2012, 96, 29–31. [Google Scholar]
- Ramkissoon, C.; Degryse, F.; da Silva, R.C.; Baird, R.; Young, S.D.; Bailey, E.H.; McLaughlin, M.J. Improving the efficacy of selenium fertilizers for wheat biofortification. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.L.; Wang, S.C.; Gu, B.Q.; Yang, Y.X.; Song, H.B.; Xue, W.L.; Liang, W.S.; Zhang, P.Y. Further investigation on the role of selenium deficiency in the aetiology and pathogenesis of Keshan disease. Biomed. Environ. Sci. 1997, 10, 316–326. [Google Scholar] [PubMed]
- Lyons, G. Selenium in cereals: Improving the efficiency of agronomic biofortification in the UK. Plant Soil 2010, 332, 1–4. [Google Scholar] [CrossRef]
- Boldrin, P.F.; de Figueiredo, M.A.; Yang, Y.; Luo, H.; Giri, S.; Hart, J.J.; Faquin, V.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Selenium promotes sulfur accumulation and plant growth in wheat ( Triticum aestivum ). Physiol. Plant 2016, 158, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Winkel, L.H.; Lin, Z.Q. Selenium in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects; Springer: Berlin/Heidelberg, Germany, 2017; Volume 11, p. 324. [Google Scholar]
- Thavarajah, D.; Abare, A.; Mapa, I.; Coyne, C.J.; Thavarajah, P.; Kumar, S. Selecting lentil accessions for global selenium biofortification. Plants 2017, 6, 34. [Google Scholar] [CrossRef]
- Ekanayake, L.J.; Thavarajah, D.; Vial, E.; Schatz, B.; McGee, R.; Thavarajah, P. Selenium fertilization on lentil (Lens culinaris Medikus) grain yield, seed selenium concentration, and antioxidant activity. Field Crop. Res. 2015, 177, 9–14. [Google Scholar] [CrossRef]
- Eich-Greatorex, A.; Sogn, T.A.; Øgaard, A.F.; Aasen, I. Plant availability of inorganic and organic selenium fertilizer as influenced by soil organic matter content and pH. Nutr. Cycl. Agroecosystems 2007, 79, 221–231. [Google Scholar] [CrossRef]
- Christophersen, O.A.; Lyons, G.; Haug, A.; Steinnes, E. Selenium; Springer: Berlin/Heidelberg, Germany, 2013; pp. 429–463. [Google Scholar]
- Hartfiel, W.; Bahners, N. Selenium deficiency in the Federal Republic of Germany. Biol. Trace Elem. 1988, 15, 1–12. [Google Scholar] [CrossRef]
- Hu, Q.; Chen, L.; Xu, J.; Zhang, Y.; Pan, G. Determination of selenium concentration in rice and the effect of foliar application of Se-enriched fertiliser or sodium selenite on the selenium content of rice. J. Sci. Food Agric. 2002, 82, 869–872. [Google Scholar] [CrossRef]
- Smrkolj, P.; Stibilj, V.; Kreft, I.; Germ, M. Selenium species in buckwheat cultivated with foliar addition of Se (VI) and various levels of UV-B radiation. Food Chem. 2006, 96, 675–681. [Google Scholar] [CrossRef]
- Poggi, V.; Arcioni, A.; Filippini, P.; Pifferi, P.G. Foliar application of selenite and selenate to potato (Solanum tuberosum): Effect of a ligand agent on selenium content of tubers. J. Agric. Food Chem. 2000, 48, 4749–4751. [Google Scholar] [CrossRef]
- Yang, F.; Chen, L.; Hu, Q.; Pan, G. Effect of the application of selenium on selenium content of soybean and its products. Biol. Trace Elem. Res. 2003, 93, 249–256. [Google Scholar] [CrossRef]
- Xia, Q.; Yang, Z.; Shui, Y.; Liu, X.; Chen, J.; Khan, S.; Wang, J.; Gao, Z. Methods of Selenium Application Differentially Modulate Plant Growth, Selenium Accumulation and Speciation, Protein, Anthocyanins and Concentrations of Mineral Elements in Purple-Grained Wheat. Front. Plant. Sci. 2020, 11, 1114. [Google Scholar] [CrossRef]
- Mao, H.; Wang, J.; Wang, Z.; Zan, Y.; Lyons, G.; Zou, C. Using agronomic biofortification to boost zinc, selenium, and iodine concentrations of food crops grown on the loess plateau in China. J. Soil Sci. Plant. Nutr. 2014, 14, 459–470. [Google Scholar] [CrossRef]
- Tveitnes, S.; Singh, B.R.; Ruud, L. Selenium concentration in spring wheat as influenced by basal application and top dressing of selenium-enriched fertilizers. Fertil. Res. 1995, 45, 163–167. [Google Scholar] [CrossRef]
- Fernández-Martínez, A.; Charlet, L. Selenium environmental cycling and bioavailability: A structural chemist point of view. Rev. Environ. Sci. Biotechnol. 2009, 8, 81–110. [Google Scholar] [CrossRef]
- Freeman, J.L.; Bañuelos, G.S. Selection of salt and boron tolerant selenium hyperaccumulator Stanleya pinnata genotypes and characterization of Se phytoremediation from agricultural drainage sediments. Environ. Sci. Technol. 2011, 45, 9703–9710. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.; Pickering, I.J.; Yang, S.I.; Freeman, J.L. Selenium biofortification of broccoli and carrots grown in soil amended with Se-enriched hyperaccumulator Stanleya pinnata. Food Chem. 2015, 166, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Kikkert, J.; Berkelaar, E. Plant uptake and translocation of inorganic and organic forms of selenium. Arch. Environ. Contam. Toxicol. 2013, 65, 458–465. [Google Scholar] [CrossRef]
- Bhatia, P.; Aureli, F.; D’Amato, M.; Prakash, R.; Cameotra, S.S.; Nagaraja, T.P.; Cubadda, F. Selenium bioaccessibility and speciation in biofortified Pleurotus mushrooms grown on selenium-rich agricultural residues. Food Chem. 2013, 140, 225–230. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.Q.; Broadley, M. Selenium biofortification. In Selenium in Plants; Springer: Berlin/Heidelberg, Germany, 2017; pp. 231–255. [Google Scholar] [CrossRef]
- Li, Z.; Liang, D.; Peng, Q.; Cui, Z.; Huang, J.; Lin, Z. Interaction between selenium and soil organic matter and its impact on soil selenium bioavailability: A review. Geoderma 2017, 295, 69–79. [Google Scholar] [CrossRef]
- Serafin Muñoz, A.H.; Kubachka, K.; Wrobel, K.; Gutierrez Corona, J.F.; Yathavakilla, S.K.; Caruso, J.A.; Wrobel, K. Se-enriched mycelia of Pleurotus ostreatus: Distribution of selenium in cell walls and cell membranes/cytosol. J. Agric. Food Chem. 2006, 54, 3440–3444. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Yamada, H.; Kyuma, K.; Hattori, T.; Kigasawa, S. Selenium in soil humic acid. Soil Sci. Plant. Nutr. 1991, 37, 241–248. [Google Scholar] [CrossRef]
- Park, J.H.; Lamb, D.; Paneerselvam, P.; Choppala, G.; Bolan, N.; Chung, J.W. Role of organic amendments on enhanced bioremediation of heavy metal(loid) contaminated soils. J. Hazard. Mater. 2011, 185, 549–574. [Google Scholar] [CrossRef]
- Dhillon, K.S.; Dhillon, S.K.; Dogra, R. Selenium accumulation by forage and grain crops and volatilization from seleniferous soils amended with different organic materials. Chemosphere 2010, 78, 548–556. [Google Scholar] [CrossRef]
- Sharma, S.; Bansal, A.; Dogra, R.; Dhillon, S.K.; Dhillon, K.S. Effect of organic amendments on uptake of selenium and biochemical grain composition of wheat and rape grown on seleniferous soils in northwestern India. J. Plant. Nutr. Soil Sci. 2011, 174, 269–275. [Google Scholar] [CrossRef]
- Bhatia, P.; Prakash, R.; Prakash, N.T. Enhanced antioxidant properties as a function of selenium uptake by edible mushrooms cultivated on selenium-accumulated waste post-harvest wheat and paddy residues. Int. J. Recycl. Org. Waste Agric. 2014, 3, 127–132. [Google Scholar] [CrossRef]
- Wu, Z.; Bañuelos, G.S.; Lin, Z.Q.; Liu, Y.; Yuan, L.; Yin, X.; Li, M. Biofortification and phytoremediation of selenium in China. Front. Plant. Sci. 2015, 6, 136. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q. Cultivation of the Indian fig Opuntia in selenium-rich drainage sediments under field conditions. Soil Use Manag. 2010, 26, 167–175. [Google Scholar] [CrossRef]
- De Feudis, M.; D’Amato, R.; Businelli, D.; Guiducci, M. Fate of selenium in soil: A case study in a maize (Zea mays L.) field under two irrigation regimes and fertilized with sodium selenite. Sci. Total Environ. 2019, 659, 131–139. [Google Scholar] [CrossRef]
- El-Ramady, H.R.; Domokos-Szabolcsy, É.; Shalaby, T.A.; Prokisch, J.; Fári, M. Selenium in agriculture: Water, air, soil, plants, food, animals and nanoselenium. In CO2 Sequestration, Biofuels and Depollution; Springer: Berlin/Heidelberg, Germany, 2015; pp. 153–232. [Google Scholar]
- Kaur, S.; Kaur, N.; Siddique, K.H.; Nayyar, H. Beneficial elements for agricultural crops and their functional relevance in defence against stresses. Arch. Agron. Soil Sci. 2016, 62, 905–920. [Google Scholar] [CrossRef]
- Sogn, L.; Skorge, P.; Frøslie, A.; Aasen, I.; Stabbetorp, H.; Ruud, L. Effects of selenium enriched complex fertilizers on selenium concentration of small grains. In Human and Animal Health in Relation to Circulation Processes of Selenium and Cadmium; The Norwegian Academy of Science and Letters: Oslo, Norway, 1991; pp. 199–211. [Google Scholar]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral Elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, Se and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; Schneberg, K.A.; Pilon-Smits, E.A. Sulfur–selenium–molybdenum interactions distinguish selenium hyperaccumulator Stanleya pinnata from non-hyperaccumulator Brassica juncea (Brassicaceae). Planta 2014, 239, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M.; Berthomieu, P.; Clairotte, M.; Shibagaki, N.; Davidian, J.C.; Gosti, F. Unequal functional redundancy between the two Arabidopsis thaliana high-affinity sulphate transporters SULTR1;1 and SULTR1;2. New Phytol. 2008, 180, 608–619. [Google Scholar] [CrossRef]
- Pu, Z.E.; Yu, M.; He, Q.Y.; Chen, G.Y.; Wang, J.R.; Liu, Y.X.; Jiang, Q.; Li, W.; Dai, S.; Wei, Y.; et al. Quantitative trait loci associated with micronutrient concentrations in two recombinant inbred wheat lines. J. Integr. Agric. 2014, 13, 2322–2329. [Google Scholar] [CrossRef]
- BuN, M.; Hui, L.; Lashari, M.S.; Shah, A.N.; Licao, C.; Weining, S. Nutritional characteristics and starch properties of Tibetan barley. Int. J. Agric. Policy Res. 2015, 3, 293–299. [Google Scholar]
- Eurola, M.; Hietaniemi, V.; Kontturi, M. Selenium content of Finnish oats in 1997–1999, effect of cultivars and cultivation techniques. Agric. Food Sci. 2004, 13, 46–53. [Google Scholar] [CrossRef]
- Jha, A.B.; Warkentin, T.D. Biofortification of pulse crops: Status and future perspectives. Plants 2020, 9, 73. [Google Scholar] [CrossRef]
- Ray, H.; Bett, K.; Tar’an, B.; Vandenberg, A.; Thavarajah, D.; Warkentin, T. Mineral micronutrient content of cultivars of field pea, chickpea, common bean, and lentil grown in Saskatchewan, Canada. Crop. Sci. 2014, 54, 1698–1708. [Google Scholar] [CrossRef]
- Nair, R.M.; Thavarajah, P.; Giri, R.R.; Ledesma, D.; Yang, R.Y.; Hanson, P.; Easdown, W.; Hughes, J.A. Mineral and phenolic concentrations of mungbean [Vigna radiata (L.) R. Wilczek var. radiata] grown in semi-arid tropical India. J. Food Compos. Anal. 2015, 39, 23–32. [Google Scholar] [CrossRef]
- Ramamurthy, R.K.; Jedlicka, J.; Graef, G.L.; Waters, B.M. Identification of new QTLs for seed mineral, cysteine, and methionine concentrations in soybean [Glycine max (L.) Merr.]. Mol. Breed. 2014, 34, 431–445. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Randle, W.M. Short-day onion cultivars differ in bulb selenium and sulphur accumulation which can affect bulb pungency. Euphytica 1997, 96, 385–390. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Randle, W.M. Genetic variances and selection potential for selenium accumulation in a rapid-cycling Brassica oleracea population. J. Am. Soc. Hortic. Sci. 2001, 126, 329–335. [Google Scholar] [CrossRef]
- Meenakshi, J.V.; Johnson, N.L.; Manyong, V.M.; DeGroote, H.; Javelosa, J. How cost effective is biofortification in combating micronutrient malnutrition? World Dev. 2010, 38, 64–75. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H. Selenium in Plants. In Progress in Botany. Progress in Botany (Genetics—Physiology—Systematics—Ecology); Lüttge, U., Beyschlag, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 76. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2015, 117, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Sors, T.G.; Ellis, D.R.; Na, G.N.; Lahner, B.; Lee, S.; Leustek, T.; Pickering, I.J.; Salt, D.E. Analysis of sulfur and selenium assimilation in Astragalus plants with varying capacities to accumulate selenium. Plant J. 2005, 42, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Terry, N.; Leduc, D.L.; Pilon-Smits, E.A.H.; Mackey, B. Field trial of transgenic Indian mustard plants shows enhanced phytoremediation of selenium-contaminated sediment. Environ. Sci. Technol. 2005, 39, 1771–1777. [Google Scholar] [CrossRef]
- Zhang, L.; Byrne, P.F.; Pilon-Smits, E.A. Mapping quantitative trait loci associated with selenate tolerance in Arabidopsis thaliana. New Phytol. 2006, 170, 33–42. [Google Scholar] [CrossRef]
- Ates, D.; Sever, T.; Aldemir, S.; Yagmur, B.; Temel, H.Y.; Kaya, H.B.; Alsaleh, A.; Kahraman, A.; Ozkan, H.; Vandenberg, A.; et al. Identification QTLs Controlling Genes for Se Uptake in Lentil Seeds. PLoS ONE 2016, 11, e0149210. [Google Scholar]
- Wang, J.; Zhou, C.; Xiao, X.; Xie, Y.; Zhu, L.; Ma, Z. Enhanced Iron and Selenium Uptake in Plants by Volatile Emissions of Bacillus amyloliquefaciens (BF06). Appl. Sci. 2017, 7, 85. [Google Scholar] [CrossRef]
- Huang, Y.; Sun, C.; Min, J.; Chen, Y.; Tong, C.; Bao, J. Association mapping of quantitative trait loci for mineral element contents in whole grain rice (Oryza sativa L.). J. Agric. Food Chem. 2015, 63, 10885–10892. [Google Scholar] [CrossRef]
- Yang, R.; Wang, R.; Xue, W.; Yan, J.; Zhao, G.; Fahima, T.; Cheng, J. QTL location and analysis of selenium content in tetraploid wheat grain. Guizhou Agric. Sci. 2013, 10, 1–4. [Google Scholar]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in greater plants. Annu. Rev. Plant. Physiol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Raina, M.; Sharma, A.; Nazir, M.; Kumari, P.; Rustagi, A.; Hami, A.; Bhau, B.S.; Zargar, S.M.; Kumar, D. Exploring the new dimensions of selenium research to understand the underlying mechanism of its uptake, translocation, and accumulation. Physiol. Plant. 2020. [Google Scholar] [CrossRef] [PubMed]
- Agalou, A.; Roussis, A.; Spaink, H.P. The Arabidopsis selenium-binding protein confers tolerance to toxic levels of selenium. Funct Plant Biol. 2005, 32, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Yang, J.; Liu, C.; Chen, Z.; Yao, Z.; Cao, S. Overexpression of ethylene response factor ERF96 gene enhances selenium tolerance in Arabidopsis. Plant Physiol Plant Soil with accelerated HIV disease progression among HIV-1-infected pregnant women in Tanzania. J. Nutr. 2020, 134, 2556–2560. [Google Scholar]
- Chen, M.; Zeng, L.; Luo, X.; Mehboob, M.Z.; Ao, T.; Lang, M. Identification and functional characterization of a novel selenocysteine methyltransferase from Brassica juncea L. J. Exp. Bot. 2019, 70, 6401–6416. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, B.; Deng, K.; Gao, X.; Sun, G.; Zhang, Z.; Li, P.; Wang, W.; Li, H.; Zhang, Z.; et al. NRT1.1B improves selenium concentrations in rice grains by facilitating selenomethinone translocation. Plant Biotechnol. J. 2019, 17, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Poletti, S.; Sautter, C. Biofortification of the crops with micronutrients using plant breeding and/or transgenic strategies. Minerva Biotecnol. 2005, 17, 1–11. [Google Scholar]
- Carvalho, S.M.P.; Vasconcelos, M.W. Producing more with less: Strategies and novel technologies for plant-based food biofortification. Food Res. Int. 2013, 54, 961–971. [Google Scholar] [CrossRef]
- Ye, Y.; Qu, J.; Pu, Y.; Rao, S.; Xu, F.; Wu, C. Selenium Biofortification of Crop Food by Beneficial Microorganisms. J. Fungi 2020, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Yasin, M.; El-Mehdawi, A.F.; Anwar, A.; Pilon Smits, E.A.H.; Faisal, M. Microbial enhanced selenium and iron biofortification of wheat (Triticum aestivum L.) Applications in Phytoremediation and Biofortification. Int. J. Phytoremed. 2015, 17, 341–347. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.P.; Chu, D.; Zhao, M.; Zayed, A.M.; Ruzin, S.E.; Schichnes, D.; Terry, N. Rhizosphere bacteria enhance selenium accumulation and volatilization by indian mustard. Plant. Physiol. 1999, 119, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Dong, T.; Ye, J.; Hou, Z. Selenium accumulation in wheat (Triticum aestivum L) as affected by coapplication of either selenite or selenate with phosphorus. Soil Sci. Plant Nutr. 2017, 63, 37–44. [Google Scholar] [CrossRef]
- Cabannes, E.; Buchner, P.; Broadley, M.R.; Hawkesford, M.J. A comparison of sulfate and selenium accumulation in relation to the expression of sulfate transporter genes in Astragalus species. Plant Physiol. 2011, 157, 2227–2239. [Google Scholar] [CrossRef]
- Lindblom, S.D.; Valdez-Barillas, J.R.; Fakra, S.C.; Marcus, M.A.; Wangeline, A.L.; Pilon-Smits, E.A.H. Influence of microbial associations on selenium localization and speciation in roots of Astragalus and Stanleya hyperaccumulators. Environ. Exp. Bot. 2013, 88, 33–42. [Google Scholar] [CrossRef]
- Patharajan, S.; Raaman, N. Influence of arbuscular mycorrhizal fungi on growth and selenium uptake by garlic plants. Arch. Phytopathol. Plant. Prot. 2012, 45, 138–151. [Google Scholar] [CrossRef]
- Larsen, E.H.; Lobinski, R.; Burger-Meÿer, K.; Hansen, M.; Ruzik, R.; Mazurowska, L.; Kik, C. Uptake and speciation of selenium in garlic cultivated in soil amended with symbiotic fungi (mycorrhiza) and selenate. Anal. Bioanal. Chem. 2006, 385, 1098–1108. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, S.; Wen, B.; Huang, H.; Luo, L. Accumulation and Speciation of Selenium in Plants as Affected by Arbuscular Mycorrhizal Fungus Glomus mosseae. Biol. Trace Elem. Res. 2011, 143, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Li, J.; Ma, X.; Niu, H.; Hou, S.; Wu, F. Effect of arbuscular mycorrhizal fungi on uptake of selenate, selenite, and selenomethionine by roots of winter wheat. Plant Soil 2019, 438, 71–83. [Google Scholar] [CrossRef]
- Sanmartín, P.; DeAraujo, A.; Vasanthakumar, A. Melding the old with the new: Trends in methods used to identify, monitor, and control microorganisms on cultural heritage materials. Microb. Ecol. 2018, 76, 64–80. [Google Scholar] [CrossRef]
- Fordyce, F.; Masara, D.; Appleton, J. Final Report on Stream Sediment, Soil and Forage Chemistry as Indicators of Cattle Mineral Status in North-East Zimbabwe; British Geological Survey: Nottingham, UK, 1994; 70p, (WC/94/003); Available online: http://nora.nerc.ac.uk/id/eprint/8350/1/WC94003.pdf (accessed on 24 January 2021).
- Patel, P.J.; Trivedi, G.R.; Shah, R.K.; Saraf, M. Selenorhizobacteria: As biofortification tool in sustainable agriculture. Biocatal. Agric. Biotechnol. 2018, 14, 198–203. [Google Scholar] [CrossRef]
- Kloepper, J.W. Effects of Rhizosphere Colonization by Plant Growth-Promoting Rhizobacteria on Potato Plant Development and Yield. Phytopathology 1980, 70, 1078. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Islam, M.T. Co-inoculation with Enterobacter and Rhizobacteria on Yield and Nutrient Uptake by Wheat (Triticum aestivum L.) in the Alluvial Soil under Indo-Gangetic Plain of India. J. Plant Growth Regul. 2017, 36, 608–617. [Google Scholar] [CrossRef]
- Raghavendra, M.P.; Nayaka, S.C.; Nuthan, B.R. Role of Rhizosphere Microflora in Potassium Solubilization. Potassium Solubilizing Microorg. Sustain. Agric. 2016, 43–59. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.J.; Jorquera, M.A.; Azcón, R.; Paredes, C.; Rengel, Z.; de la Luz Mora, M. Endophytic bacteria from selenium-supplemented wheat plants could be useful for plant-growth promotion, biofortification and Gaeumannomyces graminis biocontrol in wheat production. Biol. Fertil. Soils 2014, 50, 983–990. [Google Scholar] [CrossRef]
- Abera Tuffa, Y. Phenotypic, Symbiotic and Plant Growth Promoting Properties of Soybean Nodulating Rhizobia under Greenhouse and Field Conditions in Ethiopia. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2019. [Google Scholar]
- Nakamaru, Y.M.; Altansuvd, J. Speciation and bioavailability of selenium and antimony in non-flooded and wetland soils: A review. Chemosphere 2014, 111, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Abadin, Z.U.; Yasin, M.; Faisal, M. Bacterial-Mediated Selenium Biofortification of Triticum aestivum: Strategy for Improvement in Selenium Phytoremediation and Biofortification. Agric. Important Microbes Sustain. Agric. 2017, 299–315. [Google Scholar] [CrossRef]
- Trivedi, G.; Patel, P.; Saraf, M. Synergistic effect of endophytic selenobacteria on biofortification and growth of Glycine max under drought stress. South. Afr. J. Bot. 2019. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.; Jorquera, M.; Azcón, R.; Borie, F.; Cornejo, P.; Mora, M. Enhanced selenium content in wheat grain by co-inoculation of selenobacteria and arbuscular mycorrhizal fungi: A preliminary study as a potential Se biofortification strategy. J. Cereal Sci. 2013, 57, 275–280. [Google Scholar] [CrossRef]
- Wang, P.; Wang, H.; Liu, Q.; Tian, X.; Shi, Y.; Zhang, X. QTL mapping of selenium content using a RIL population in wheat. PLoS ONE 2017, 12, e0184351. [Google Scholar] [CrossRef]
- Bodnar, M.; Konieczka, P.; Namiesnik, J. The properties, functions, and use of selenium compounds in living organisms. J. Environ. Sci. Health Part C 2012, 30, 225–252. [Google Scholar] [CrossRef]
- Renkema, H.; Koopmans, A.; Kersbergen, L.; Kikkert, J.; Hale, B.; Berkelaar, E. The effect of transpiration on selenium uptake and mobility in durum wheat and spring canola. Plant Soil 2012, 354, 239–250. [Google Scholar] [CrossRef]
- Missana, T.; Alonso, U.; García-Gutiérrez, M. Experimental study and modeling of selenite sorption onto illite and smectite clays. J. Colloid Interface Sci. 2009, 334, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef]
- El Kassis, E.; Cathala, E.; Rouached, H.; Fourcroy, P.; Berthomieu, P.; Terry, N.; Davidian, J.C. Characterization of a selenate-resistant Arabidopsis mutant. Root growth as a potential target for selenate toxicity. Plant Physiol. 2007, 143, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Saito, K. Molecular biology and functional genomics for identification of regulatory networks of plant sulfate uptake and assimilatory metabolism. In Sulfur Metabolism in Phototrophic Organisms; Springer: Berlin/Heidelberg, Germany, 2008; pp. 149–159. [Google Scholar]
- Shibagaki, N.; Rose, A.; McDermott, J.P.; Fujiwara, T.; Hayashi, H.; Yoneyama, T.; Davies, J.P. Selenate-resistant mutants of Arabidopsis thaliana identify Sultr1;2, a sulfate transporter required for efficient transport of sulfate into roots. Plant J. 2002, 29, 475–486. [Google Scholar] [CrossRef]
- Ellis, D.R.; Salt, D.E. Plants, selenium, and human health. Curr. Opin. Plant Biol. 2002, 6, 273–279. [Google Scholar] [CrossRef]
- Cappa, J.J.; Cappa, P.J.; El Mehdawi, A.F.; McAleer, J.M.; Simmons, M.P.; Pilon-Smits, E.A. Characterization of selenium and sulfur accumulation across the genus Stanleya (Brassicaceae): A field survey and common-garden experiment. Am. J. Bot. 2014, 101, 830–839. [Google Scholar] [CrossRef]
- Ximénez-Embún, P.; Alonso, I.; Madrid-Albarrán, Y.; Cámara, C. Establishment of selenium uptake and species distribution in lupine, Indian mustard, and sunflower plants. J. Agric. Food Chem. 2004, 52, 832–838. [Google Scholar] [CrossRef]
- Mazej, D.; Osvald, J.; Stibilj, V. Selenium species in leaves of chicory, dandelion, lamb’s lettuce and parsley. Food Chem. 2008, 107, 75–83. [Google Scholar] [CrossRef]
- Gigolashvili, T.; Kopriva, S. Transporters in plant sulphur metabolism. Front. Plant Sci. 2014, 5, 422. [Google Scholar] [CrossRef] [PubMed]
- Galeas, M.L.; Zhang, L.H.; Freeman, J.L.; Wegner, M.; Pilon-Smits, E.A.H. Seasonal fluctuations of selenium and sulfur accumulation in selenium-hyperaccumulators and related non-accumulators. New Phytol. 2007, 173, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Jarzyńska, G.; Falandysz, J. Selenium and 17 other largely essential and toxic metals in muscle and organ meats of Red Deer (Cervus elaphus)—consequences to human health. Environ. Int. 2011, 37, 882–888. [Google Scholar] [CrossRef]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current knowledge on selenium biofortification to improve the nutraceutical profile of food: A comprehensive review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef] [PubMed]
- Pezzarossa, B.; Rosellini, I.; Borghesi, E.; Tonutti, P.; Malorgio, F. Effects of Se enrich- ment on yield, fruit composition and ripening of tomato (Solanum lycopersicum) plants grown in hydroponics. Sci. Hortic. 2014, 165, 106–110. [Google Scholar] [CrossRef]
- Bachiega, P.; Salgado, J.M.; de Carvalho, J.E.; Ruiz, A.L.T.G.; Schwarz, K.; Tezotto, T.; Morzelle, M.C. Antioxidant and antiproliferative activities in different maturation stages of broccoli (Brassica oleracea Italica) biofortified with selenium. Food Chem. 2016, 190, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Businelli, D.; D’Amato, R.; Onofri, A.; Tedeschin, E.; Tei, F. Se-enrichment of cucumber (Cucumis sativus L.), lettuce (Lactuca sativa L.) and tomato (Solanum lycopersicum L.) through fortification in pre-transplanting. Sci. Hortic. 2015, 197, 697–704. [Google Scholar] [CrossRef]
- Smoleń, S.; Skoczylas, L.; Ledwozyw-Smolen, I.; Rakoczy, R.; Kopec, A.; Ewa Piatkowska, E.; Biezanowska-Kopec, R.; Koronowicz, A.; Kapusta-Duch, J. Biofortification of carrot (Daucus carota L.) with iodine and selenium in a field experiment. Front. Plant. Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Joy, E.J.M.; Kalimbira, A.A.; Gashu, D.; Ferguson, E.L.; Sturgess, J.; Dangour, A.D.; Banda, L.; Chiutsi-Phiri, G.; Bailey, E.H. Can selenium deficiency in Malawi be alleviated through consumption of agro-biofortified maize flour? Study protocol for a randomised, double-blind, controlled trial. Trials 2019, 20, 795. [Google Scholar] [CrossRef]
- Davis, C.D. Nutritional interactions: Credentialing of molecular targets for cancer prevention. Exp. Biol. Med. 2007, 232, 176–183. [Google Scholar]
- Malagoli, M.; Schiavon, M.; Dall’acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. Plant. Sci. 2015, 6, 1–5. [Google Scholar] [CrossRef]
- Chang, J.C.; Gutenmann, W.H.; Reid, C.M.; Lisk, D.J. Selenium content of Brazil nuts from two geographic locations in Brazil. Chemosphere 1995, 30, 801–802. [Google Scholar] [CrossRef]
- Whelan, R.; Barrow, N.J. Slow-release selenium fertilizers to correct selenium sheep in Western Australia. Fertil. Res. 1994, 38, 183–188. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilon-Smits, E.A. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef]
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta Gen. Subj. 2018. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D. A tale of two toxicities: Malformed selenoproteins and oxidative stress both contribute to seleni- um stress in plants. Ann. Bot. 2013, 112, 965–972. [Google Scholar] [CrossRef]
- Malik, J.A.; Goel, S.; Kaur, N.; Sharma, S.; Singh, I.; Nayyar, H. Selenium antagonizes the toxic effects of arsenic on mungbean (Phaseolus aureus Roxb.) plants by restricting its uptake and enhancing the antioxidative and detoxification mechanisms. Environ. Exp. Bot. 2012, 77, 242–248. [Google Scholar] [CrossRef]
- Habibi, G. Physiological, photochemical and ionic responses of sunflower seedlings to exogenous selenium sup- ply under salt stress. Acta Physiol. Plant 2017, 39, 213. [Google Scholar] [CrossRef]
- Manojlović, M.S.; Lončarić, Z.; Cabilovski, R.R.; Popović, B.; Karalić, K.; Ivezić, V.; Ademi, A.; Singh, B.R. Biofortification of wheat cultivars with selenium. Soil Plant Sci. 2019, 69, 715–724. [Google Scholar] [CrossRef]
- Sarwar, N.; Akhtar, M.; Kamran, M.A.; Imran, M.; Riaz, M.A.; Kamran, K.; Hussain, S. Selenium biofortification in food crops: Key mechanisms and future perspectives. J. Food Compos. Anal. 2020, 103615. [Google Scholar] [CrossRef]
- Mechora, Š. Selenium as a protective agent against pests: A review. Plants 2019, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jia, W.; Hu, C.; Nie, M.; Ming, J.; Cheng, Q.; Cai, M.; Sun, X.; Li, X.; Zheng, X.; et al. Selenium as a potential fungicide could protect oilseed rape leaves from Sclerotinia sclerotiorum infection. Environ. Pollut. 2020, 257, 113495. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).