
Phenolic Constituents, Antioxidant and Cytoprotective Activities, Enzyme Inhibition Abilities of Five Fractions from Vaccinium dunalianum Wight


 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Total Phenolic Content and Total Flavonoid Content
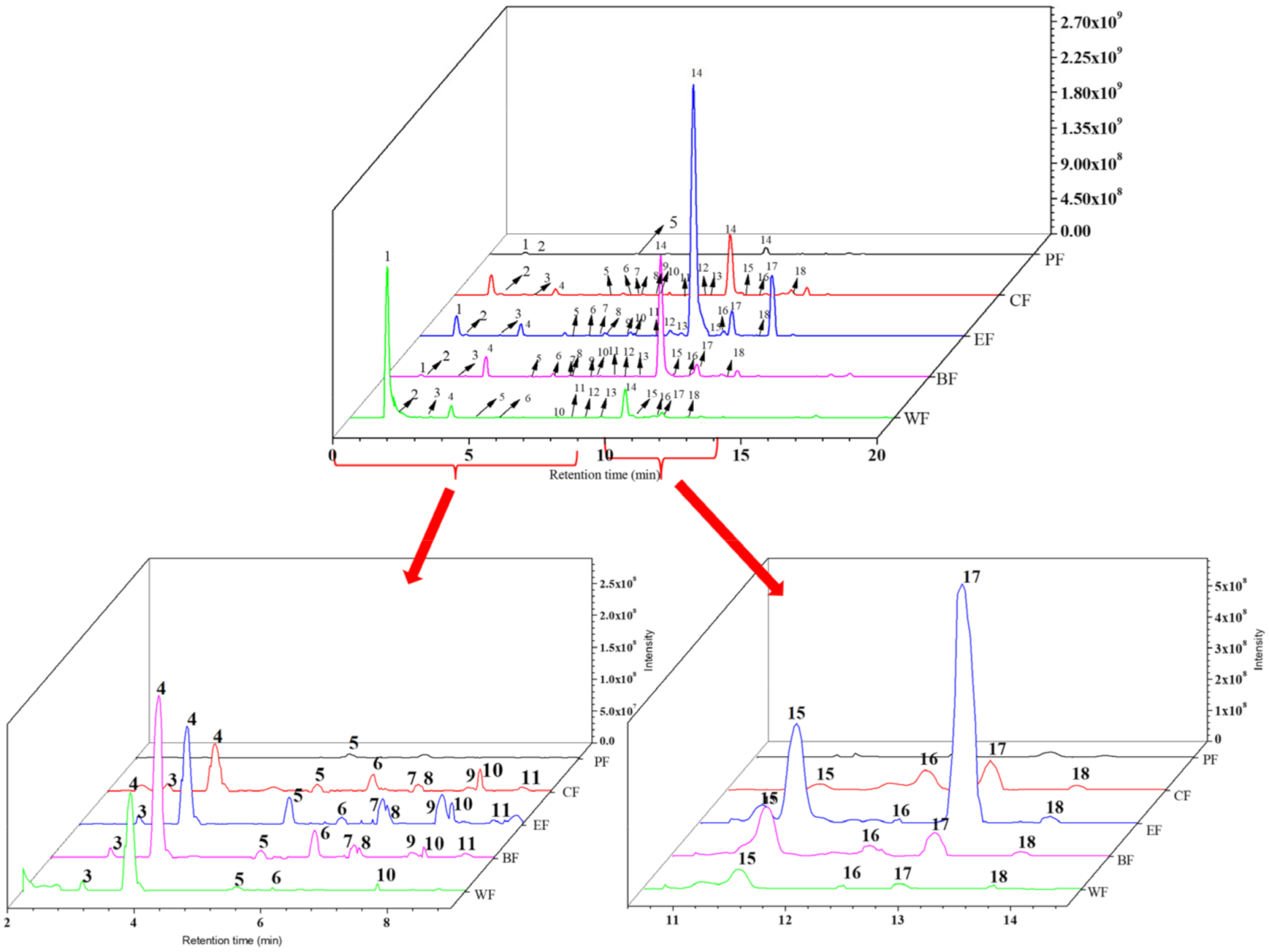
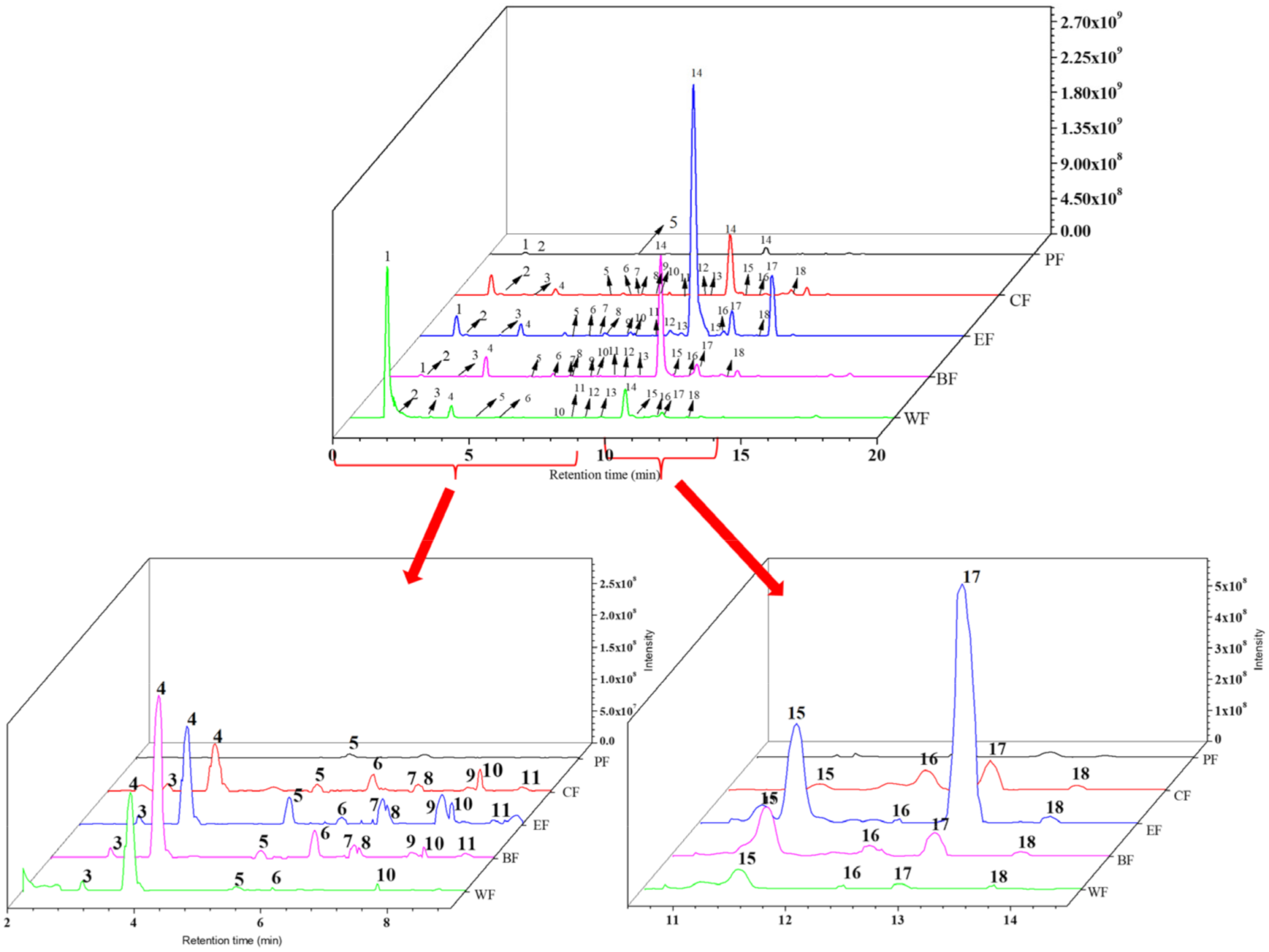
2.2. Chemical Composition Analysis of Five Fractions
2.3. Enzyme Inhibition Activity of Five Fractions
2.3.1. α-Glucosidase Inhibitory Activity
2.3.2. Inhibitory Activity of Pancreatic Lipase
2.3.3. Antioxidant Effects of Five Fractions
2.4. Cytoprotective Effect on Oxidative Damage in H2O2-Induced HepG2 Cells
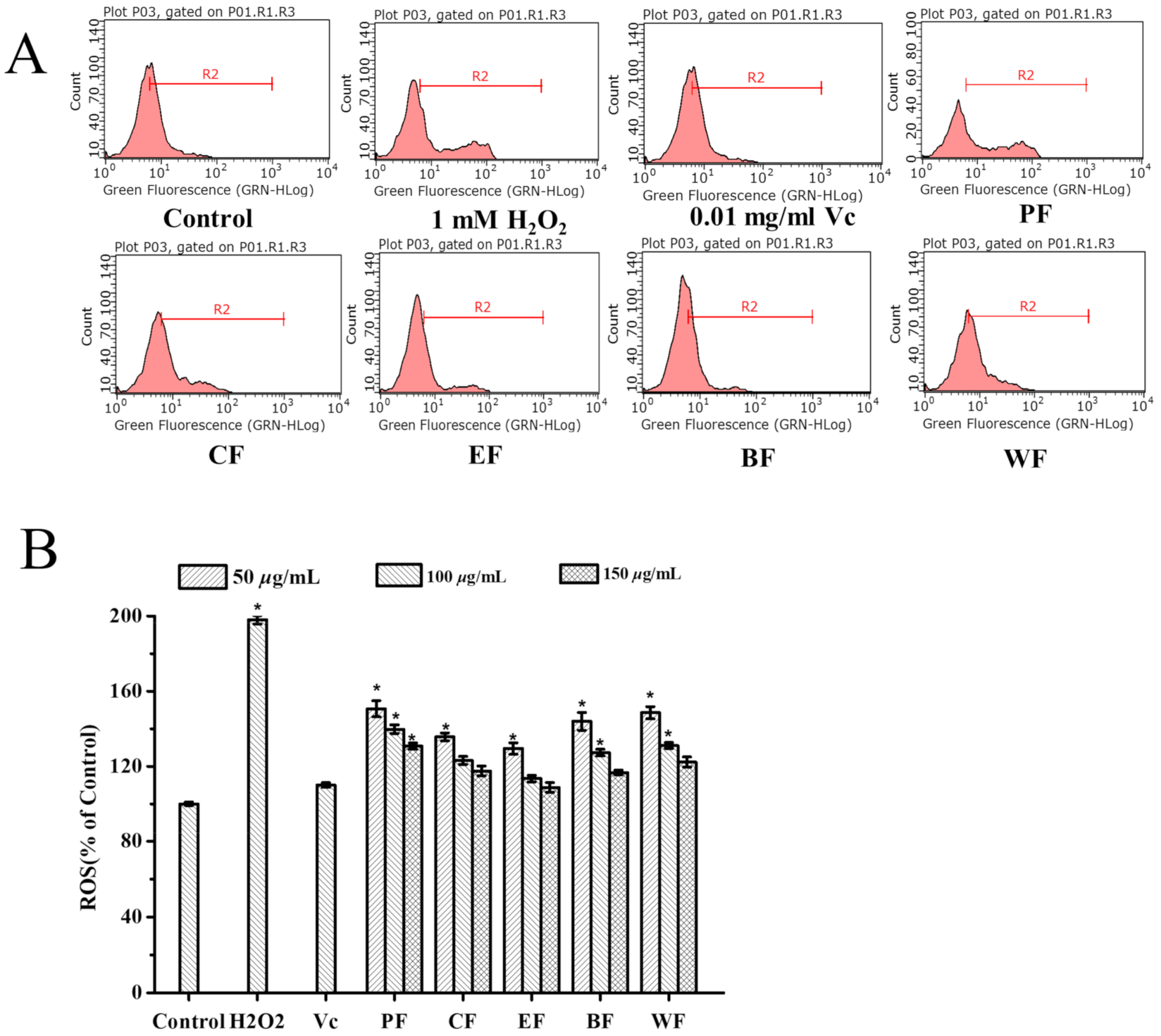
2.4.1. Inhibitory Effect on Intracellular Reactive Oxygen (ROS) Generation
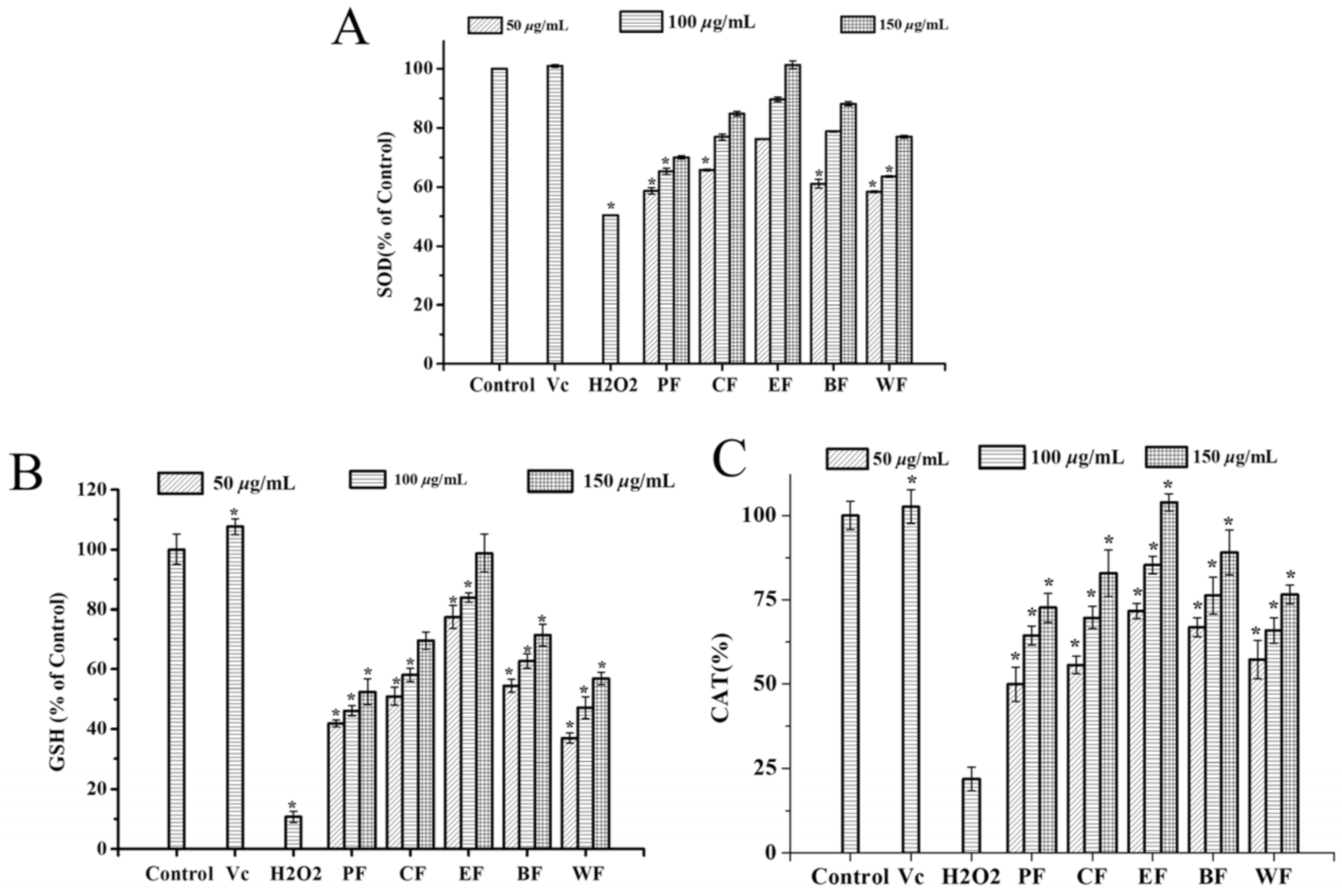
2.4.2. Effect on Intracellular Antioxidant Enzyme Activities
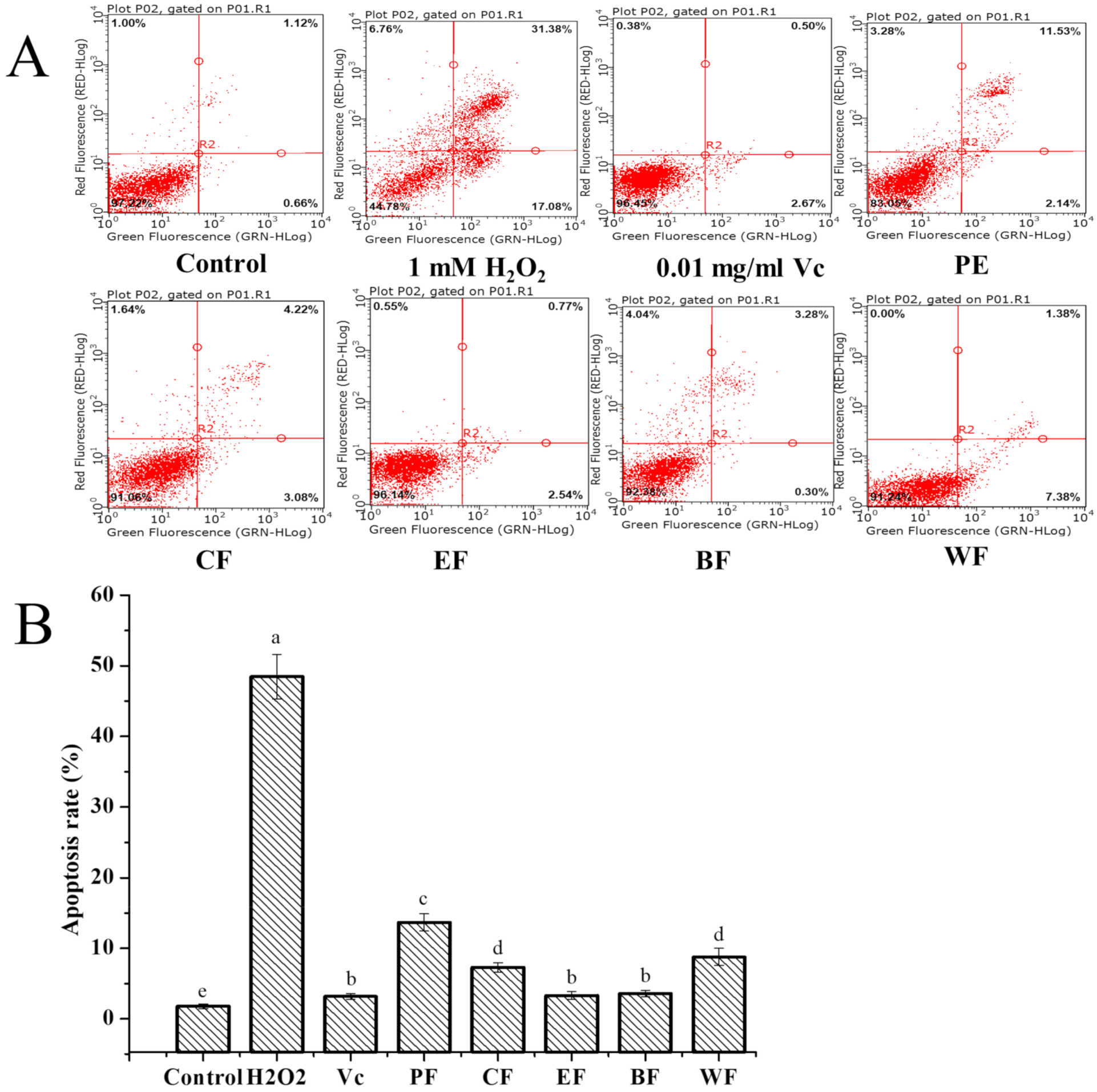
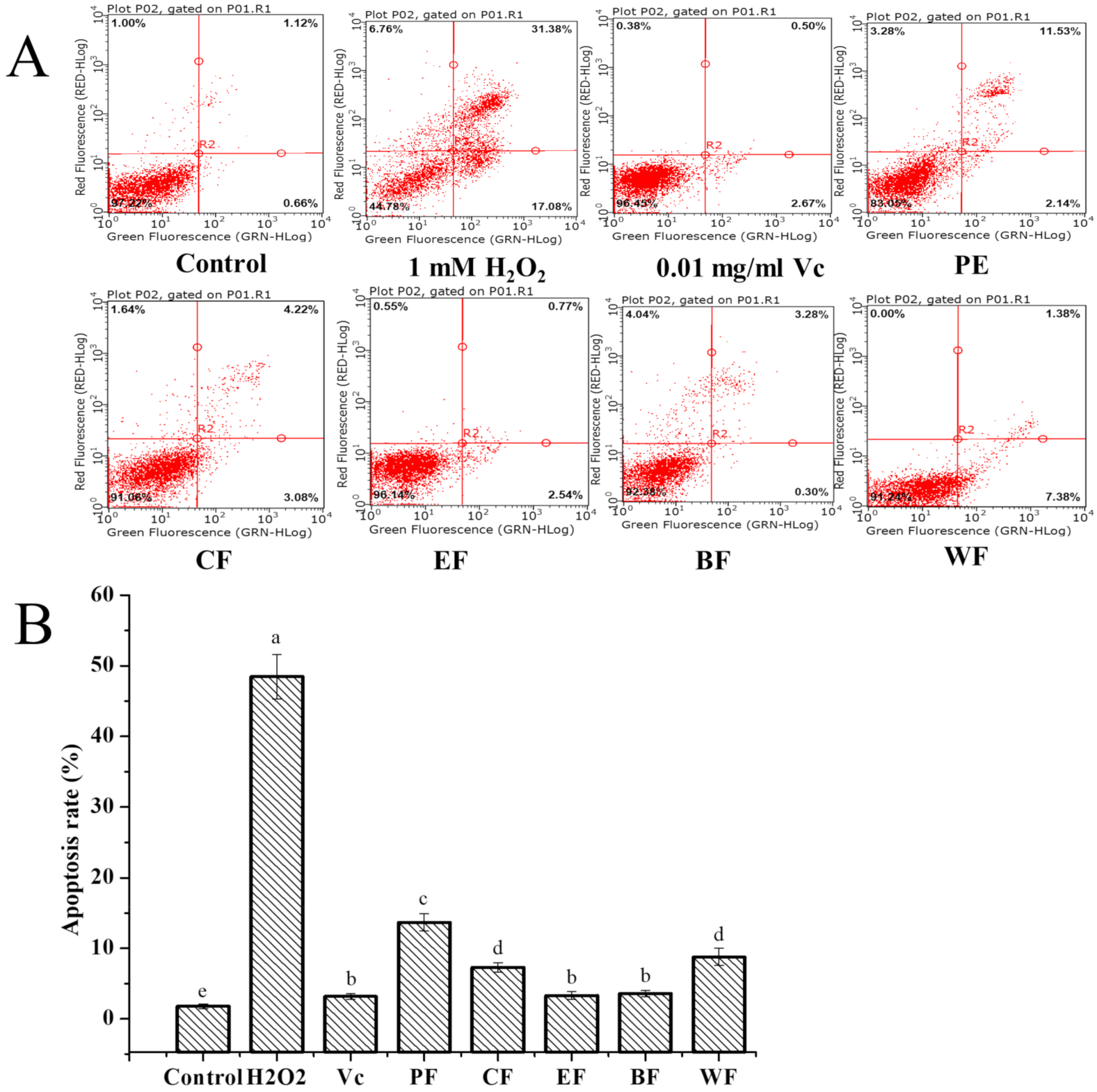
2.4.3. Cytoprotective Activity against H2O2-Induced Cell Apoptosis
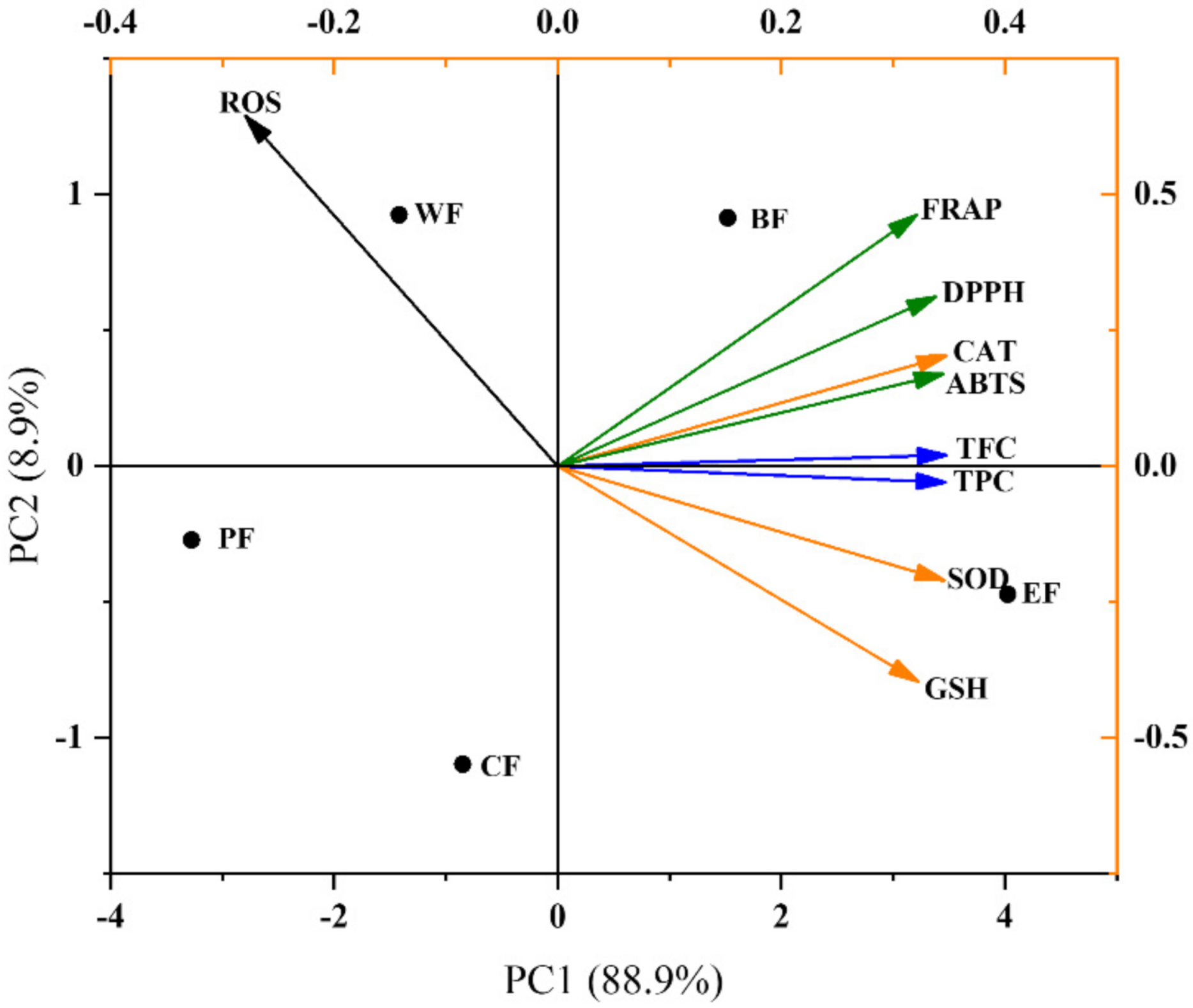
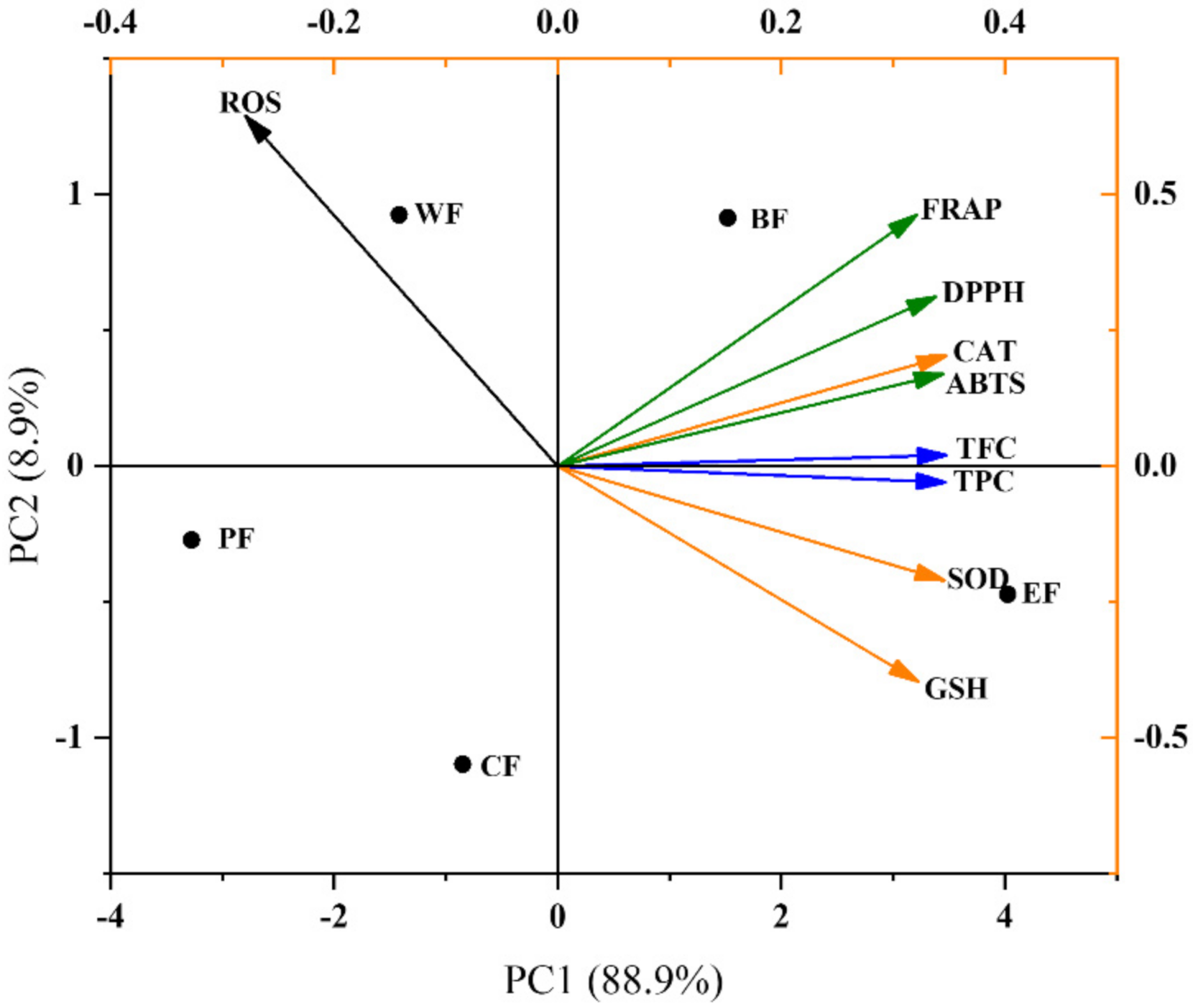
2.5. Multivariate Analysis
3. Materials and Methods
3.1. Materials and Reagents
3.2. Plant Material
3.3. Extraction and Preparation of Five Fractions of Vaccinium Dunalianum
3.4. Determination of Total Phenolic Content and Total Flavonoid Content
3.4.1. Determination of Total Phenolic Content
3.4.2. Determination of Total Flavonoid Content
3.4.3. Identification and Quantification of Phenolics by UHPLC-MS/MS
3.5. Enzyme Inhibition Ability Assay
3.5.1. Inhibitory Activity of α-Glucosidase
3.5.2. Inhibitory Activity of Pancreatic Lipase
3.6. Antioxidant Activity Assay
3.6.1. DPPH-Radical Scavenging Capacity Assay
3.6.2. Determination of FRAP Reducing Ability
3.6.3. Determination of ABTS+ Free Radical Scavenging Capacity
3.7. Cytoprotective Effect on Oxidative Damage in H2O2-Induced HepG2 Cells
3.7.1. Cell Culture and Cell Viability
3.7.2. Measurement of Intracellular Reactive Oxygen Species (ROS)
3.7.3. Determination of Cell Apoptosis
3.7.4. Determination of Intracellular Antioxidant Enzyme Activity
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.R.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Zhao, T.; Sun, M.; Kong, L.; Xue, Q.; Wang, Y.; Wang, Y.; Khan, A.; Cao, J.; Cheng, G. Bioactivity-Guided Isolation of Phytochemicals from Vaccinium dunalianum Wight and Their Antioxidant and Enzyme Inhibitory Activities. Molecules 2021, 26, 2075. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of citrus flavonoids, naringin and naringenin, on metabolic syndrome and their mechanisms of action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. 2005, 579, 200–213. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar] [CrossRef]
- Xu, J.; Cao, J.; Yue, J.; Zhang, X.; Zhao, Y. New triterpenoids from acorns of Quercus liaotungensis and their inhibitory activity against α-glucosidase, α-amylase and protein-tyrosine phosphatase 1B. J. Funct. Foods 2018, 41, 232–239. [Google Scholar] [CrossRef]
- van de Laar, F.A.; Lucassen, P.L.; Akkermans, R.P.; van de Lisdonk, E.H.; Rutten, G.E.; van Weel, C. Alpha-glucosidase inhibitors for patients with type 2 diabetes: Results from a Cochrane systematic review and meta-analysis. Diabetes Care 2005, 28, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Rasouli, H.; Hosseini-Ghazvini, S.M.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 8, 1942–1954. [Google Scholar] [CrossRef]
- Butsat, S.; Siriamornpun, S. Effect of solvent types and extraction times on phenolic and flavonoid contents and antioxidant activity in leaf extracts of Amomum chinense C. Int. Food Res. J. 2016, 23, 180–187. [Google Scholar]
- Jiang, Y.; Fang, Z.; Leonard, W.; Zhang, P. Phenolic compounds in Lycium berry: Composition, health benefits and industrial applications. J. Funct. Foods 2021, 77, 104340. [Google Scholar] [CrossRef]
- Umeno, A.; Horie, M.; Murotomi, K.; Nakajima, Y.; Yoshida, Y. Antioxidative and Antidiabetic Effects of Natural Polyphenols and Isoflavones. Molecules 2016, 21, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-K.; Zhou, X.-L.; Wang, X.-Q.; Zhang, J.-X.; Yang, M.-L.; Liu, Y.-P.; Cao, J.-X.; Cheng, G.-G. Que Zui tea ameliorates hepatic lipid accumulation and oxidative stress in high fat diet induced nonalcoholic fatty liver disease. Food Res. Int. 2022, 156, 111196. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Raffa, D.; Maggio, B.; Raimondi, M.V.; Plescia, F.; Daidone, G. Recent discoveries of anticancer flavonoids. Eur. J. Med. Chem. 2017, 142, 213–228. [Google Scholar] [CrossRef]
- Gao, S.-H.; Zhao, T.-R.; Liu, Y.-P.; Wang, Y.-F.; Cheng, G.-G.; Cao, J.-X. Phenolic constituents, antioxidant activity and neuroprotective effects of ethanol extracts of fruits, leaves and flower buds from Vaccinium dunalianum Wight. Food Chem. 2022, 374, 131752. [Google Scholar] [CrossRef]
- Russell, W.; Duthie, G. Plant secondary metabolites and gut health: The case for phenolic acids. Proc. Nutr. Soc. 2011, 70, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Mendes, V.; Costa, V.; Mateus, N. Involvement of the modulation of cancer cell redox status in the anti-tumoral effect of phenolic compounds. RSC Adv. 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, M.; Zhang, H.-Q.; Sun, S.; Xia, B.; Wu, F.-H. In vivo hypoglycemic effects of phenolics from the root bark of Morus alba. Fitoterapia 2009, 80, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.-S.; Cho, C.-W.; Hong, H.-D.; Lee, Y.-C.; Choi, U.-K.; Kim, Y.-C. Hypolipidemic and antioxidant properties of phenolic compound-rich extracts from white ginseng (Panax ginseng) in cholesterol-fed rabbits. Molecules 2013, 18, 12548–12560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.-J.; Park, J.-H.; Choi, H.-N.; Kim, J.-I. α-glucosidase inhibitory activities of myricetin in animal models of diabetes mellitus. Food Sci. Biotechnol. 2015, 24, 1897–1900. [Google Scholar] [CrossRef]
- Berton, A.; Sebban-Kreuzer, C.; Rouvellac, S.; Lopez, C.; Crenon, I. Individual and combined action of pancreatic lipase and pancreatic lipase-related proteins 1 and 2 on native versus homogenized milk fat globules. Mol. Nutr. Food Res. 2009, 53, 1592–1602. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Born, M.; Carrupt, P.-A.; Zini, R.; Brée, F.; Tillement, J.-P.; Hostettmann, K.; Testa, B. Electrochemical Behaviour and Antioxidant Activity of Some Natural Polyphenols. Helv. Chim. Acta 1996, 79, 1147–1158. [Google Scholar] [CrossRef]
- Das, N.P.; Ramanathan, L. Studies on Flavonoids and Related Compounds as Antioxidants in Food. In Lipid-Soluble Antioxidants: Biochemistry and Clinical Applications; Birkhäuser: Basel, Switzerland, 1992; pp. 295–306. [Google Scholar]
- Wang, L.L.; Yu, Q.L.; Han, L.; Ma, X.L.; Song, R.D.; Zhao, S.N.; Zhang, W.H. Study on the effect of reactive oxygen species-mediated oxidative stress on the activation of mitochondrial apoptosis and the tenderness of yak meat. Food Chem. 2018, 244, 394–402. [Google Scholar] [CrossRef]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Wu, H.; Jiao, C.; Wu, B.; Yu, H.; Yan, B.; Liang, Y.J.E.P. Cytotoxicity induced by iodinated haloacetamides via ROS accumulation and apoptosis in HepG-2 cells. Environ. Pollut. 2018, 242, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, Q.; He, L.; Shu, Y.; Zhang, S.; Tan, S.; Lin, T.J.O.M.; Cellular Longevity, Active Fragment of Veronica ciliata Fisch. Attenuates t-BHP-Induced Oxidative Stress Injury in HepG2 Cells through Antioxidant and Antiapoptosis Activities. Oxidative Med. Cell. Longev. 2017, 2017, 4727151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L.J.F.; Toxicology, C. Protective effects of rambutan (Nephelium lappaceum) peel phenolics on H2O2-induced oxidative damages in HepG2 cells and d-galactose-induced aging mice. Food Chem. Toxicol. 2017, 108, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Biology, M.J.N.R.M.C. Opinion: Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nature reviews. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar]
- Gamez-Meza, N.; Noriega-Rodriguez, J.A.; Medina-Juarez, L.A.; Ortega-Garcia, J.; Cazarez-Casanova, R.; Angulo-Guerrero, O. Antioxidant activity in soybean oil of extracts from Thompson grape bagasse. J. Am. Oil Chem. Soc. 1999, 76, 1445–1447. [Google Scholar] [CrossRef]
- Zengin, G. Products, A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crops Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Cai, S.; Wang, O.; Wu, W.; Zhu, S.; Zhou, F.; Ji, B.; Gao, F.; Zhang, D.; Liu, J.; Cheng, Q. Comparative Study of the Effects of Solid-State Fermentation with Three Filamentous Fungi on the Total Phenolics Content (TPC), Flavonoids, and Antioxidant Activities of Subfractions from Oats (Avena sativa L.). J. Agric. Food Chem. 2012, 60, 507–513. [Google Scholar] [CrossRef]
- Abate, G.; Zhang, L.; Pucci, M.; Morbini, G.; Mac Sweeney, E.; Maccarinelli, G.; Ribaudo, G.; Gianoncelli, A.; Uberti, D.; Memo, M.; et al. Phytochemical Analysis and Anti-Inflammatory Activity of Different Ethanolic Phyto-Extracts of Artemisia annua L. Biomolecules 2021, 11, 975. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Y.; Gao, F.; Zhao, Y.; Cai, S.; Pang, M. The free, esterified, and insoluble-bound phenolic profiles of Rhus chinensis Mill. fruits and their pancreatic lipase inhibitory activities with molecular docking analysis. J. Funct. Foods 2018, 40, 729–735. [Google Scholar] [CrossRef]
- Yang, M.; Ma, Y.; Wang, Z.; Khan, A.; Zhou, W.; Zhao, T.; Cao, J.; Cheng, G.; Cai, S. Phenolic constituents, antioxidant and cytoprotective activities of crude extract and fractions from cultivated artichoke inflorescence. Ind. Crops Prod. 2020, 143, 111433. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Technology, Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amamcharla, J.K.; Metzger, L.E.J.I.D.J. Modification of the ferric reducing antioxidant power (FRAP) assay to determine the susceptibility of raw milk to oxidation. Int. Dairy J. 2014, 34, 177–179. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, Y.; Yang, M.; Cao, J.; Cheng, G.J.F.C. UHPLC-ESI-HRMS/MS analysis on phenolic compositions of different E Se tea extracts and their antioxidant and cytoprotective activities. Food Chem. 2020, 318, 126512. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ma, Y.; Jia, Y.; Pang, M.; Cheng, G.; Cai, S.J.F.C. Phenolic profiles, antioxidant activities and cytoprotective effects of different phenolic fractions from oil palm (Elaeis guineensis Jacq.) fruits treated by ultra-high pressure. Food Chem. 2019, 288, 68–77. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Y.; Patel, G.; Xue, Q.; Njateng, G.S.; Cai, S.; Cheng, G.; Kai, G. In vitro and in vivo anti-inflammatory effects of different extracts from Epigynum auritum through down-regulation of NF-κB and MAPK signaling pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | TFC 1 | TPC 2 | Enzyme Inhibition Activity (IC50 μg/mL) | Antioxidant Activity | |||
|---|---|---|---|---|---|---|---|
| α-Glucosidase | Pancreatic Lipase | DPPH (nmol TE/mg Extract) 3 | FRAP (nmol TE/mg Extract) 4 | ABTS (nmol TE/mg Extract) 5 | |||
| PF | 2.71 ± 0.09 a | 12.68 ± 0.37 a | 498.56 ± 12.34 e | 814.32 ± 6.53 e | - | - | 1.35 ± 0.14 a |
| CF | 9.07 ± 0.45 c | 21.34 ± 0.56 c | 201.76 ± 6.51 c | 253.52 ± 11.86 c | 0.12 ± 0.01 a | 0.21 ± 0.01 a | 5.25 ± 0.12 b |
| EF | 23.42 ± 1.10 e | 59.65 ± 0.93 e | 84.21 ± 2.21 a | 122.53 ± 5.45 a | 0.44 ± 0.03 d | 0.91 ± 0.04 d | 12.85 ± 0.23 e |
| BF | 18.79 ± 0.18 d | 42.09 ± 0.86 d | 125.55 ± 4.75 b | 178.76 ± 4.85 b | 0.35 ± 0.01 c | 0.84 ± 0.02 c | 9.26 ± 0.12 d |
| WF | 5.54 ± 0.15 b | 15.77 ± 0.21 b | 266.36 ± 1.98 d | 435.21 ± 8.86 d | 0.21 ± 0.01 b | 0.51 ± 0.02 b | 6.55 ± 0.28 c |
| Peak | Compounds | tR (min) | [M − H]-(m/z) | Error (ppm) | Molecular Formula | MS/MS Fragment Ions | Extraction Site |
|---|---|---|---|---|---|---|---|
| 1 | quinic acid A | 1.17 | 191.0553 | 1.389 | C7H12O6 | 85.03, 93.04, 191.06 | PF, CF, EF, BF, WF |
| 2 | arbutin B | 1.51 | 271.0823 | 7.044 | C19H12O2 | 86.06, 100.08 | PF, CF, EF, BF, WF |
| 3 | 6-O-caffeoyl-D- Glucopyranose B | 3.00 | 341.0877 | 3.024 | C15H18O9 | 59.01, 135.04, 161.02, 179.03, 221.04 | CF, EF, BF, WF |
| 4 | chlorogenic acid B | 3.84 | 353.0878 | 3.091 | C16H18O9 | 85.03, 127.04, 191.06 | CF, EF, BF, WF |
| 5 | caffeic acid A | 5.35 | 179.0341 | 1.144 | C9H8O4 | 89.04, 134.04, 135.04 | PF, CF, EF, BF, WF |
| 6 | p-hydroxybenzaldehyde A | 6.21 | 121.0282 | 1.371 | C7H6O2 | 85.03, 191.06 | CF, EF, BF, WF |
| 7 | methyl caffeate B | 6.83 | 193.0134 | 1.452 | C9H5O5 | 91.02, 109.03, 137.02 | CF, EF, BF |
| 8 | Ampeloptin C | 6.86 | 319.0459 | 3.311 | C15H12O8 | 57.03, 83.01, 125.02, 137.02, 193.01 | CF, EF, BF |
| 9 | quercetin-3-O-arabinoside B | 7.77 | 435.0906 | 2.868 | C20H19O11 | 435.09 | CF, EF, BF |
| 10 | 5-O-(E)-Feruloylquinic acid C | 7.84 | 367.1034 | 2.81 | C17H20O9 | 93.03, 134.03, 191.06 | CF, EF, BF, WF |
| 11 | Neoeriocitrin C | 8.59 | 595.1671 | 2.291 | C27H32O15 | 161.02, 323.08, 433.11 | CF, EF, BF, WF |
| 12 | eriodictyol-7-glucoside C | 9.01 | 449.1088 | 2.209 | C21H22O11 | 125.02, 161.02, 179.03 | CF, EF, BF, WF |
| 13 | methyl chlorogenate C | 9.42 | 367.1067 | 2.564 | C17H20O9 | 93.03, 134.04, 191.06 | CF, EF, BF, WF |
| 14 | 6’-O-caffeoylarbutin B | 10.49 | 433.1137 | 1.886 | C21H22O10 | 161.02, 179.03 | PF, CF, EF, BF, WF |
| 15 | paeoniflorin-3-O-glucoside chloride C | 11.17 | 463.1247 | 2.531 | C22H23ClO11 | 139.04, 161.02 | CF, EF, BF |
| 16 | p-hydroxybenzoic acid A | 12.37 | 137.0233 | 0.296 | C7H5O3 | 52.03, 65.04, 93.03, 94.04 | CF, EF, BF, WF |
| 17 | robustaside A B | 12.96 | 417.1224 | 2.712 | C21H22O9 | 145.03, 163.04 | CF, EF, BF, WF |
| 18 | kaempferol-3-O-β-D-glucoside B | 13.92 | 447.1396 | 2.453 | C21H20O11 | 145.03, 160.02, 175.04, 193.05 | CF, EF, BF, WF |
| Peak | Compound | PF | CF | EF | BF | WF |
|---|---|---|---|---|---|---|
| 1 | quinic acid A | 4815.52 ± 9.85 b | 22,049.23 ± 49.02 d | 1253.7 ± 3.05 a | 19,221.03 ± 21.05 c | 239,190.25 ± 39.42 e |
| 2 | Arbutin B | 62.83 ± 2.05 a | 1542.88 ± 3.06 c | 701.59 ± 7.09 b | 2817.86 ± 16.23 d | 7109.58 ± 7.65 e |
| 3 | 6-O-caffeoyl-D- Glucopyranose B | - | 670.24 ± 14.02 a | 801.55 ± 25.03 b | 621.75 ± 15.17 a | 1013.09 ± 22.06 c |
| 4 | chlorogenic acid B | - | 25,730.28 ± 32.05 a | 42,713.84 ± 12.23 b | 129,305.52 ± 102.23 c | 47,967.83 ± 67.51 d |
| 5 | caffeic acid A | 141.64 ± 2.46 a | 2066.85 ± 11.03 c | 3534.38 ± 26.85 d | 960.22 ± 10.41 b | 151.57 ± 9.51 a |
| 6 | p-hydroxybenzaldehyde A | - | 4015.03 ± 22.92 c | 897.75 ± 12.67 b | 8736.15 ± 72.76 d | 88.08 ± 2.52 a |
| 7 | methyl caffeate B | - | 622.65 ± 25.06 a | 7723.50 ± 25.03 c | 4014.19 ± 15.01 b | - |
| 8 | Ampeloptin C | - | 1275.64 ± 26.01 a | 15,092.11 ± 75.63 c | 1711.32 ± 12.53 b | - |
| 9 | quercetin-3-O-arabinoside B | - | 82.23 ± 2.51 a | 9451.56 ± 51.02 c | 899.04 ± 18.02 b | - |
| 10 | 5-O-(E)-Feruloylquinic acid C | - | 2207.23 ± 19.04 c | 2109.58 ± 10.32 c | 1233.83 ± 5.13 b | 753.55 ± 10.15 a |
| 11 | Neoeriocitrin C | - | 82.23 ± 2.54 b | 63.96 ± 2.31 a | 1179.58 ± 17.64 d | 133.28 ± 11.06 c |
| 12 | eriodictyol-7-glucoside C | - | 168.36 ± 12.74 b | 1470.51 ± 24.11 d | 554.48 ± 10.07 c | 54.22 ± 2.32 a |
| 13 | methyl chlorogenate C | - | 361.98 ± 10.02 b | 6077.21 ± 17.03 d | 1902.92 ± 29.51 c | 52.01 ± 4.15 a |
| 14 | 6’-O-caffeoylarbutin B | 28.27 ± 2.53 a | 156,036.82 ± 40.07 c | 679,611.55 ± 17.56 e | 387,775.65 ± 61.99 d | 10,425.32 ± 54.32 b |
| 15 | paeoniflorin-3-O-glucoside chloride C | - | 2379.87 ± 27.22 a | 20,963.06 ± 45.06 c | 4376.52 ± 53.03 b | - |
| 16 | p-hydroxybenzoic acid A | - | 4608.03 ± 48.53 c | 86.55 ± 2.16 a | 1767.16 ± 29.42 b | 52.56 ± 12.95 a |
| 17 | robustaside A B | - | 9905.46 ± 50.08 c | 72,950.26 ± 150.02 d | 7932.82 ± 62.33 b | 180.75 ± 23.25 a |
| 18 | kaempferol-3-O-β-D-glucoside B | - | 504.39 ± 4.51 c | 981.48 ± 20.59 d | 156.46 ± 13.32 b | 75.15 ± 2.61 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.-S.; Gu, Q.-H.; Zhang, J.-K.; Tao, J.-H.; Zhao, T.-R.; Cao, J.-X.; Cheng, G.-G.; Lai, G.-F.; Liu, Y.-P. Phenolic Constituents, Antioxidant and Cytoprotective Activities, Enzyme Inhibition Abilities of Five Fractions from Vaccinium dunalianum Wight. Molecules 2022, 27, 3432. https://doi.org/10.3390/molecules27113432
Cheng C-S, Gu Q-H, Zhang J-K, Tao J-H, Zhao T-R, Cao J-X, Cheng G-G, Lai G-F, Liu Y-P. Phenolic Constituents, Antioxidant and Cytoprotective Activities, Enzyme Inhibition Abilities of Five Fractions from Vaccinium dunalianum Wight. Molecules. 2022; 27(11):3432. https://doi.org/10.3390/molecules27113432
Chicago/Turabian StyleCheng, Chang-Shu, Qing-Hui Gu, Jin-Ke Zhang, Jun-Hong Tao, Tian-Rui Zhao, Jian-Xin Cao, Gui-Guang Cheng, Guo-Fang Lai, and Ya-Ping Liu. 2022. "Phenolic Constituents, Antioxidant and Cytoprotective Activities, Enzyme Inhibition Abilities of Five Fractions from Vaccinium dunalianum Wight" Molecules 27, no. 11: 3432. https://doi.org/10.3390/molecules27113432
APA StyleCheng, C. -S., Gu, Q. -H., Zhang, J. -K., Tao, J. -H., Zhao, T. -R., Cao, J. -X., Cheng, G. -G., Lai, G. -F., & Liu, Y. -P. (2022). Phenolic Constituents, Antioxidant and Cytoprotective Activities, Enzyme Inhibition Abilities of Five Fractions from Vaccinium dunalianum Wight. Molecules, 27(11), 3432. https://doi.org/10.3390/molecules27113432