

Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
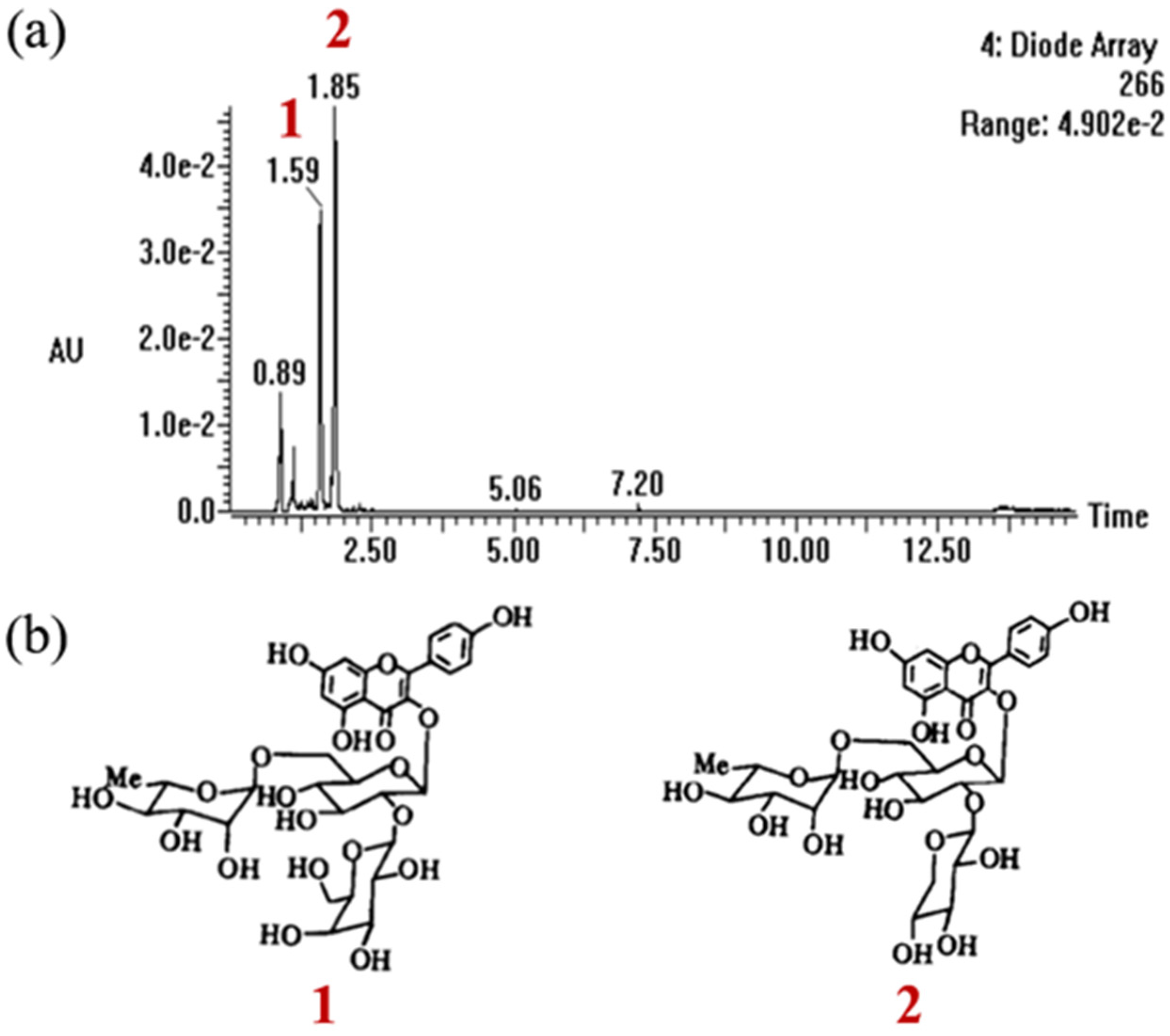
2.1. Characterization Analysis of CSCE
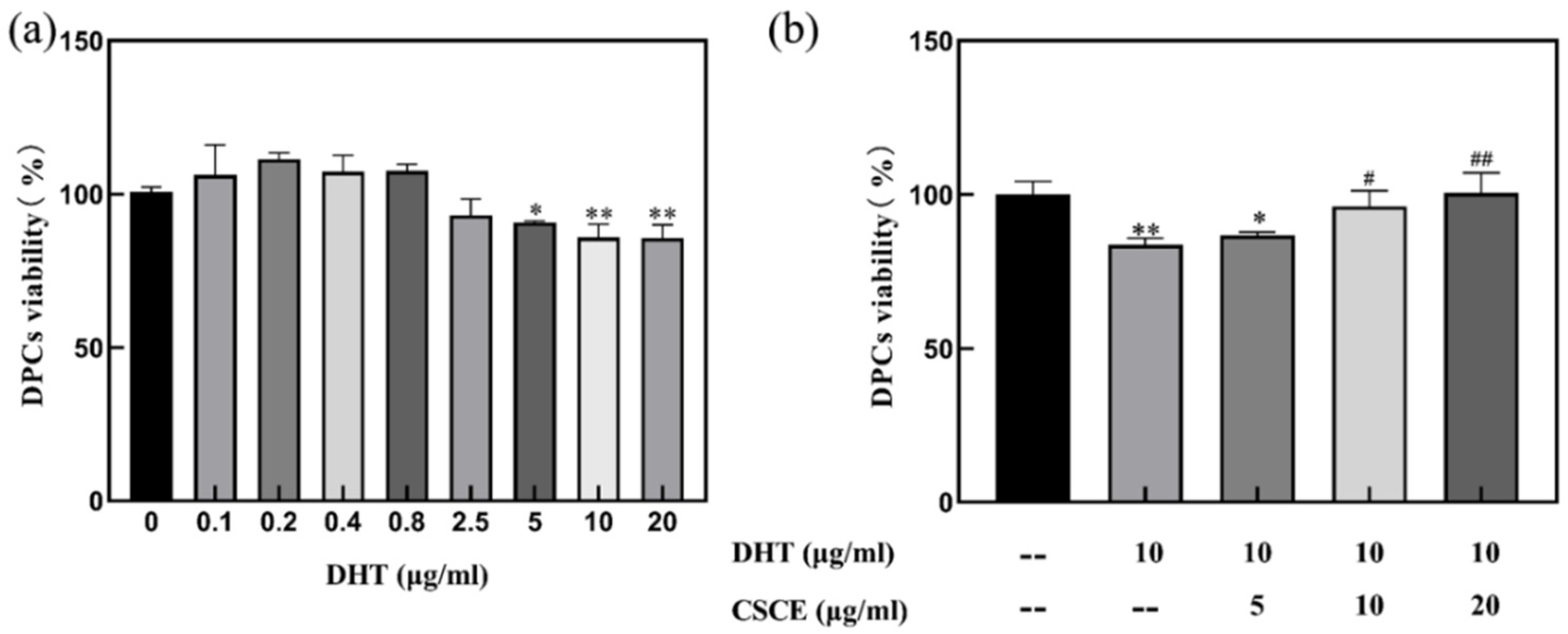
2.2. CSCE Alleviated DHT-Induced Cell Damage
2.3. CSCE Alleviated DHT-Induced Cell Inflammation
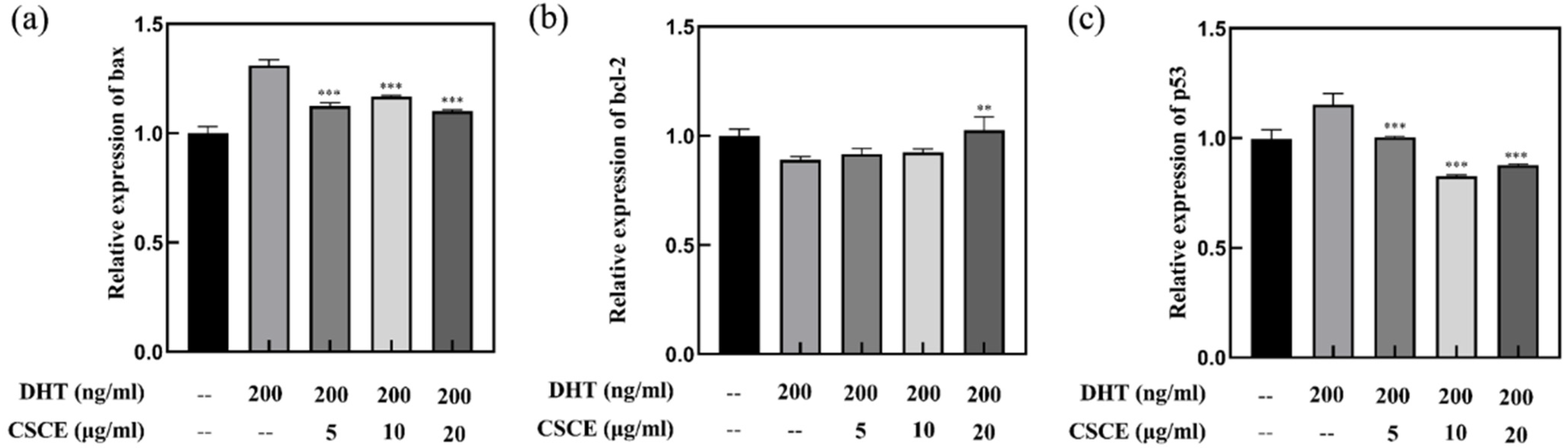
2.4. CSCE Downregulated Bax and p53 mRNA While Upregulated bcl-2 mRNA
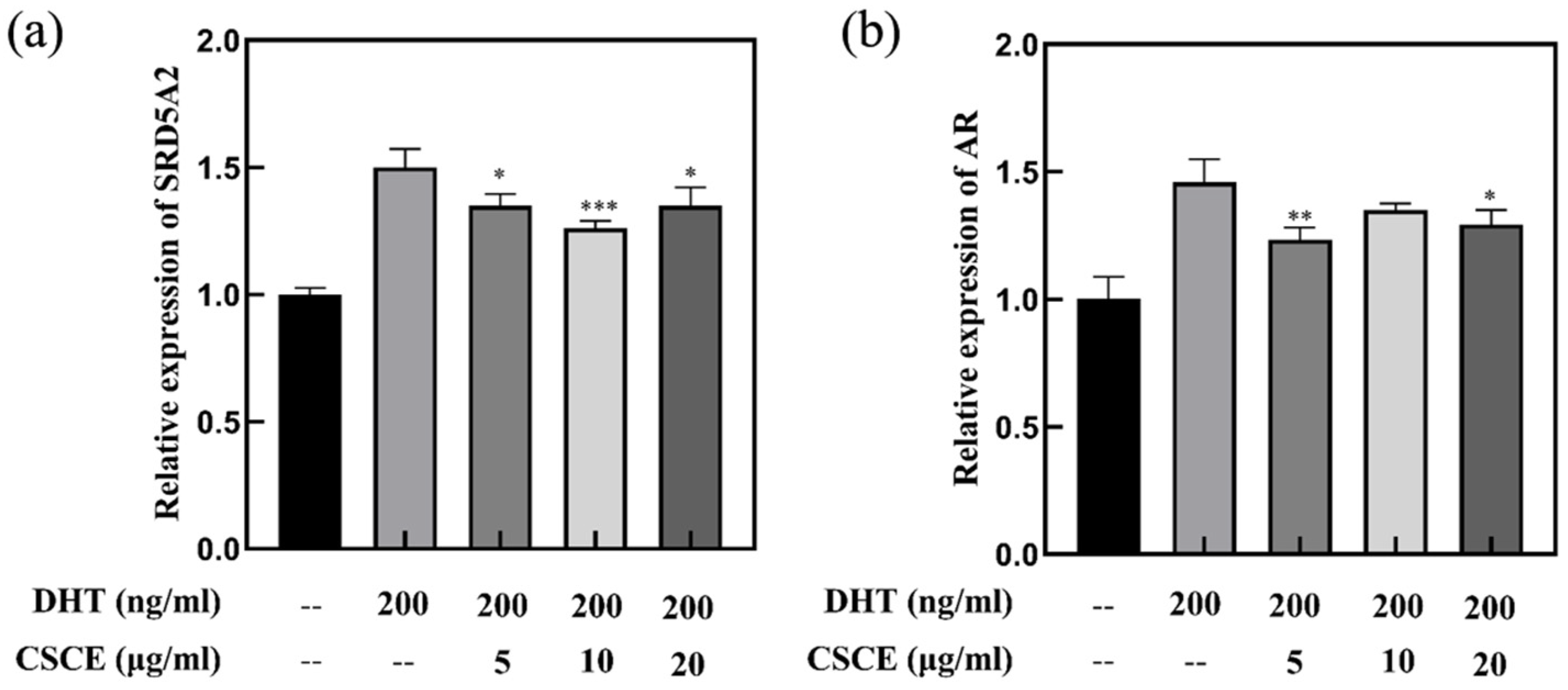
2.5. Anti-Androgenic Effect of CSCE
2.6. CSCE Delayed DHT-Induced Cell Senescence
3. Discussion
4. Materials and Methods
4.1. Preparation of Camellia Seed Cake Extract
4.2. Phytochemical Analysis by UPLC-MS
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. IL-6 and IL-1α ELISA Detection
4.6. Senescence-Associated β-Galactosidase (SA-β-Gal) Assay
4.7. Quantitative Measurement of mRNA Expression of Cytokines
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Severi, G.; Sinclair, R.; Hopper, J.; English, D.; McCredie, M.; Boyle, P.; Giles, G. Androgenetic alopecia in men aged 40–69 years: Prevalence and risk factors. Br. J. Dermatol. 2015, 149, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Sawant, N.; Chikhalkar, S.; Mehta, V.; Ravi, M.; Madke, B.; Khopkar, U. Androgenetic Alopecia: Quality-of-life and Associated Lifestyle Patterns. Int. J. Trichol. 2010, 2, 81–85. [Google Scholar] [CrossRef]
- Kaliyadan, F.; Nambiar, A.; Vijayaraghavan, S. Androgenetic alopecia: An update. Indian J. Dermatol. Venereol. Leprol. 2013, 79, 613. [Google Scholar] [CrossRef] [PubMed]
- English, R.S. A hypothetical pathogenesis model for androgenic alopecia: Clarifying the dihydrotestosterone paradox and rate-limiting recovery factors. Med. Hypotheses 2018, 111, 73–81. [Google Scholar] [CrossRef]
- Inui, S.; Itami, S. Molecular basis of androgenetic alopecia: From androgen to paracrine mediators through dermal papilla. J. Dermatol. Sci. 2011, 61, 1–6. [Google Scholar] [CrossRef]
- Lolli, F.; Pallotti, F.; Rossi, A.; Fortuna, M.C.; Caro, G.; Lenzi, A.; Sansone, A.; Lombardo, F. Androgenetic alopecia: A review. Endocrine 2017, 57, 9–17. [Google Scholar] [CrossRef]
- Deslypere, J.-P.; Young, M.; Wilson, J.; McPhaul, M. Testosterone and 5 alpha-dihydrotestosterone interact differently with the androgen receptor to enhance transcription of the MMTV-CAT reporter gene. Mol. Cell. Endocrinol. 1992, 88, 15–22. [Google Scholar] [CrossRef]
- Kwack, M.H.; Ahn, J.S.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dihydrotestosterone-inducible IL-6 inhibits elongation of human hair shafts by suppressing matrix cell proliferation and promotes regression of hair follicles in mice. J. Investig. Dermatol. 2012, 132, 43–49. [Google Scholar] [CrossRef]
- Randall, V.A.; Hibberts, N.A.; Hamada, K. A comparison of the culture and growth of dermal papilla cells from hair follicles from non-balding and balding (androgenetic alopecia) scalp. Br. J. Dermatol. 1996, 134, 437–444. [Google Scholar] [CrossRef]
- Bahta, A.W.; Farjo, N.; Farjo, B.; Philpott, M.P. Premature senescence of balding dermal papilla cells in vitro is associated with p16(INK4a) expression. J. Investig. Dermatol. 2008, 128, 1088–1094. [Google Scholar] [CrossRef] [Green Version]
- Upton, J.H.; Hannen, R.F.; Bahta, A.W.; Farjo, N.; Farjo, B.; Philpott, M.P. Oxidative Stress–Associated Senescence in Dermal Papilla Cells of Men with Androgenetic Alopecia. J. Investig. Dermatol. 2015, 135, 1244–1252. [Google Scholar] [CrossRef]
- Drake, L.; Hordonsky, M. The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. J. Am. Acad. Dermatol. 1999, 41, 550–554. [Google Scholar] [PubMed]
- Kuttenn, F.; Mowszowicz, I.; Wright, F.; Baudot, N.; Jaffiol, C.; Robin, M.; Mauvais-Jarvis, P. Male Pseudohermaphroditism: A Comparative Study of One Patient with 5α-Reductase Deficiency and Three Patients with the Complete Form of Testicular Feminization. J. Clin. Endocrinol. Metab. 1979, 6, 861. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Miki, K.; Yamaguchi, Y.; Takauji, Y.; Yamakami, Y.; Hossain, M.N.; Ayusawa, D.; Fujii, M. Extract of Plumbago zeylanica enhances the growth of hair follicle dermal papilla cells with down-regulation of 5α-reductase type II. J. Cosmet. Dermatol. 2020, 19, 3083–3090. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Choi, Y.-H.; Kim, J.; Park, S.Y.; Nam, Y.J.; Lee, S.Y.; Jeon, J.H.; Jin, M.H.; Lee, S. Polygonum multiflorum extract support hair growth by elongating anagen phase and abrogating the effect of androgen in cultured human dermal papilla cells. BMC Complementary Med. Ther. 2020, 20, 144. [Google Scholar] [CrossRef]
- Wang, J.; Shen, H.; Chen, T.; Ma, L. Hair growth-promoting effects of Camellia seed cake extract in human dermal papilla cells and C57BL/6 mice. J. Cosmet. Dermatol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Arita, J.; Yamaguchi, A.; Saito, K.; Okonogi, S.; Morisaki, N.; Iwasaki, S.; Murakoshi, I. Two flavonol glycosides from seeds of Camellia sinensis. Phytochemistry 1991, 30, 991–995. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Ahluwalia, G.; Shander, D. Apoptosis in the hair follicle. J. Investig. Dermatol. 2006, 126, 258–264. [Google Scholar] [CrossRef]
- Kwon, O.S.; Pyo, H.K.; Oh, Y.J.; Han, J.H.; Lee, S.R.; Chung, J.H.; Eun, H.C.; Kim, K.H. Promotive effect of minoxidil combined with all-trans retinoic acid (tretinoin) on human hair growth in vitro. J. Korean Med. Sci. 2007, 22, 283–289. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, H.; Wang, J. Alantolactone induces apoptosis of human cervical cancer cells via reactive oxygen species generation, glutathione depletion and inhibition of the Bcl-2/Bax signaling pathway. Oncol. Lett. 2016, 11, 4203–4207. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, F.; Jahanpour, S.; Hazem, K.; Aghbali, A.; Baradran, B.; Pakdel, S.M.V. Garlic (Allium sativum) fresh juice induces apoptosis in human oral squamous cell carcinoma: The involvement of caspase-3, Bax and Bcl-2. J. Dent. Res. Dent. Clin. Dent. Prospect. 2015, 9, 267. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Bai, M.; Suo, Y.; Zhang, G.; Cao, X. Matrine inhibited proliferation and increased apoptosis in human breast cancer MCF-7 cells via upregulation of Bax and downregulation of Bcl-2. Int. J. Clin. Exp. Pathol. 2015, 8, 14793. [Google Scholar] [PubMed]
- Zhao, X.F.; Yang, M.H.; Zheng, G.E. Effects of ultraviolet radiation on p53, Bax and Bcl-2 expression in rat lens epithelial cells. Recent Adv. Ophthalmol. 2017, 16, 2309–2312. [Google Scholar]
- Mousa, A.M.; Al-Fadhli, A.S.; Rao, M.S.; Kilarkaje, N. Gestational lead exposure induces developmental abnormalities and up-regulates apoptosis of fetal cerebellar cells in rats. Drug Chem. Toxicol. 2015, 38, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Komarova, E.A.; Siebenhaar, F.; Botchkareva, N.V.; Sharov, A.A.; Komarov, P.G.; Maurer, M.; Gudkov, A.V.; Gilchrest, B.A. p53 Involvement in the Control of Murine Hair Follicle Regression. Am. J. Pathol. 2001, 158, 1913–1919. [Google Scholar] [CrossRef]
- MagalhEs, J.D.; Passos, J.F. Stress, cell senescence and organismal ageing. Mech. Ageing Dev. 2017, 170, 2–9. [Google Scholar] [CrossRef]
- Rambwawasvika, H.; Dzomba, P.; Gwatidzo, L. Hair growth promoting effect of dicerocaryum senecioides phytochemicals. Int. J. Med. Chem. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Oh, J.W.; Kloepper, J.; Langan, E.A.; Kim, Y.; Yeo, J.; Kim, M.J.; Hsi, T.-C.; Rose, C.; Yoon, G.S.; Lee, S.-J.; et al. A guide to studying human hair follicle cycling in vivo. J. Investig. Dermatol. 2016, 136, 34–44. [Google Scholar] [CrossRef]
- Hiipakka, R.A.; Zhang, H.-Z.; Dai, W.; Dai, Q.; Liao, S. Structure-activity relationships for inhibition of human 5alpha-reductases by polyphenols. Biochem. Pharmacol. 2002, 63, 1165–1176. [Google Scholar] [CrossRef]
- Randall, V.A. Androgens and hair growth. Blackwell Publ. Inc. 2008, 21, 314–328. [Google Scholar] [CrossRef]
- Sinclair, R. Male pattern androgenetic alopecia. Bmj 1998, 317, 865–869. [Google Scholar] [CrossRef]
- Inui, S. Androgen-inducible TGF-beta1 from balding dermal papilla cells inhibits epithelial cell growth: A clue to understand paradoxical effects of androgen on human hair growth. Faseb J. 2002, 16, 1967–1969. [Google Scholar] [CrossRef] [PubMed]
- Hibino, T.; Nishiyama, T. Role of TGF-β2 in the human hair cycle. J. Dermatol. Sci. 2004, 35, 9–18. [Google Scholar] [CrossRef]
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Investig. Dermatol. 2008, 128, 262–269. [Google Scholar] [CrossRef]
- Hamada, K.; Randall, V. Inhibitory autocrine factors produced by the mesenchyme-derived hair follicle dermal papilla may be a key to male pattern baldness. Br. J. Derm. 2010, 154, 609–618. [Google Scholar] [CrossRef]
- Yang, Y.C.; Fu, H.C.; Wu, C.Y.; Wei, K.T.; Huang, K.E.; Kang, H.Y. Androgen receptor accelerates premature senescence of human dermal papilla cells in association with DNA Damage. PLoS ONE 2013, 8, e79434. [Google Scholar] [CrossRef]
- Prins, G.S. Molecular biology of the androgen receptor. Mayo Clin. Proc. 2000, 75, S32–S35. [Google Scholar] [CrossRef]
- Hibberts, N. Balding hair follicle dermal papilla cells contain higher levels of androgen receptors than those from non-balding scalp. J. Endocrinol. 1998, 156, 59–65. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Huang, Y.-C.; Huang, K.-S.; Chan, C.-C.; Chiu, H.-Y.; Tsai, R.-Y.; Chan, J.-Y.; Lin, S.-J. Stress-induced premature senescence of dermal papilla cells compromises hair follicle epithelial-mesenchymal interaction. J. Dermatol. Sci. 2017, 86, 114–122. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Jee, H.J.; Um, J.H.; Kim, Y.M.; Bae, S.S.; Yun, J. Cooperation between p21 and Akt is required for p53-dependent cellular senescence. Aging Cell 2017, 16, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Shi, J.; Zhang, Y.; Lian, A.; Han, X.; Zuo, K.; Liu, M.; Zheng, T.; Zou, F.; Liu, X.; et al. NANOG Attenuates hair follicle-derived mesenchymal stem cell senescence by upregulating PBX1 and activating AKT signaling. Oxidative Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Shen, H.; Fang, C.; Chen, T.; Wang, J. Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells. Molecules 2022, 27, 6443. https://doi.org/10.3390/molecules27196443
Ma L, Shen H, Fang C, Chen T, Wang J. Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells. Molecules. 2022; 27(19):6443. https://doi.org/10.3390/molecules27196443
Chicago/Turabian StyleMa, Ling, Huchi Shen, Chengge Fang, Timson Chen, and Jing Wang. 2022. "Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells" Molecules 27, no. 19: 6443. https://doi.org/10.3390/molecules27196443
APA StyleMa, L., Shen, H., Fang, C., Chen, T., & Wang, J. (2022). Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells. Molecules, 27(19), 6443. https://doi.org/10.3390/molecules27196443