

Talaromyces sp. Ethyl Acetate Crude Extract as Potential Mosquitocide to Control Culex pipiens quinquefasciatus
 and
and
Abstract
:1. Introduction
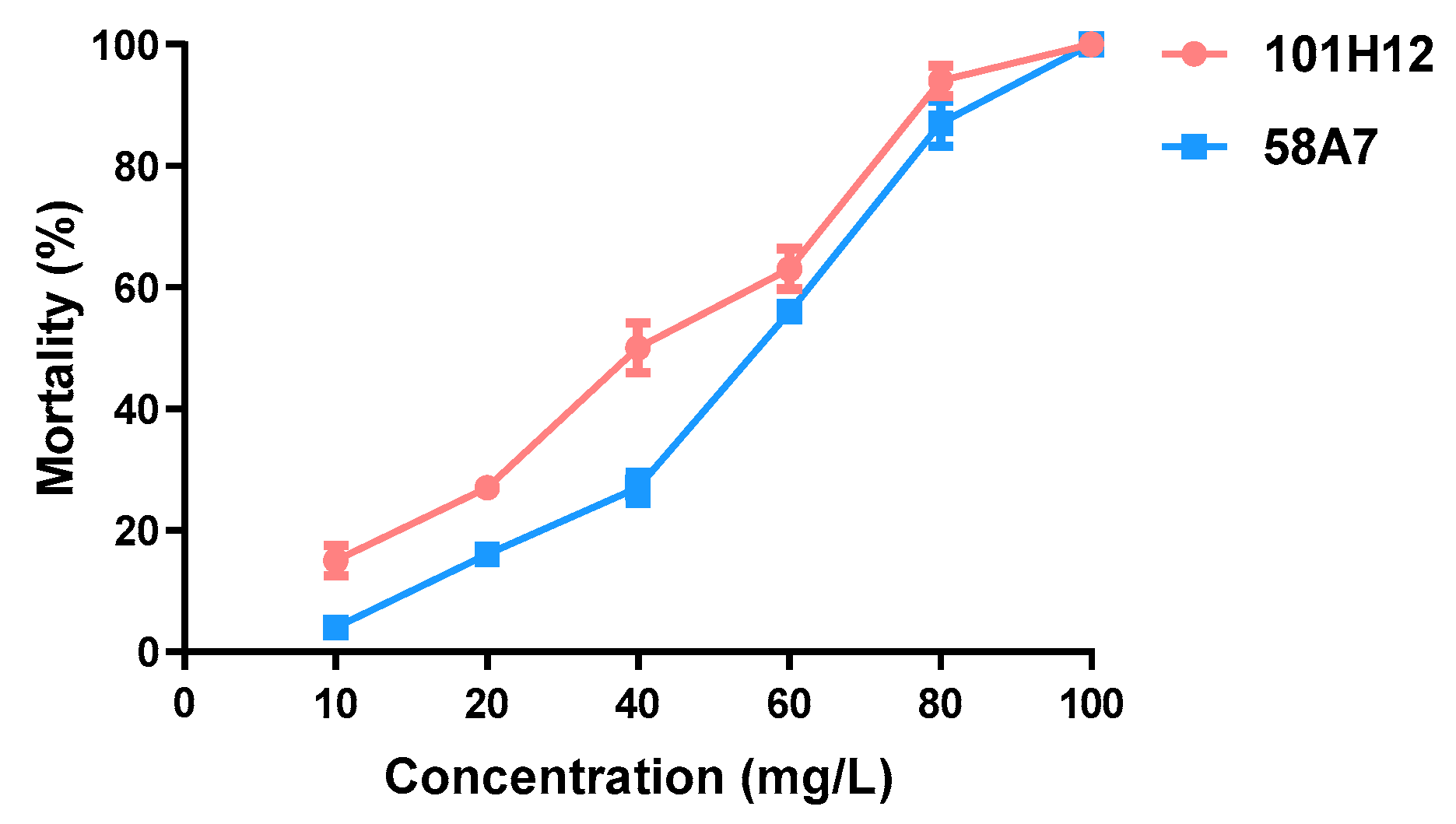
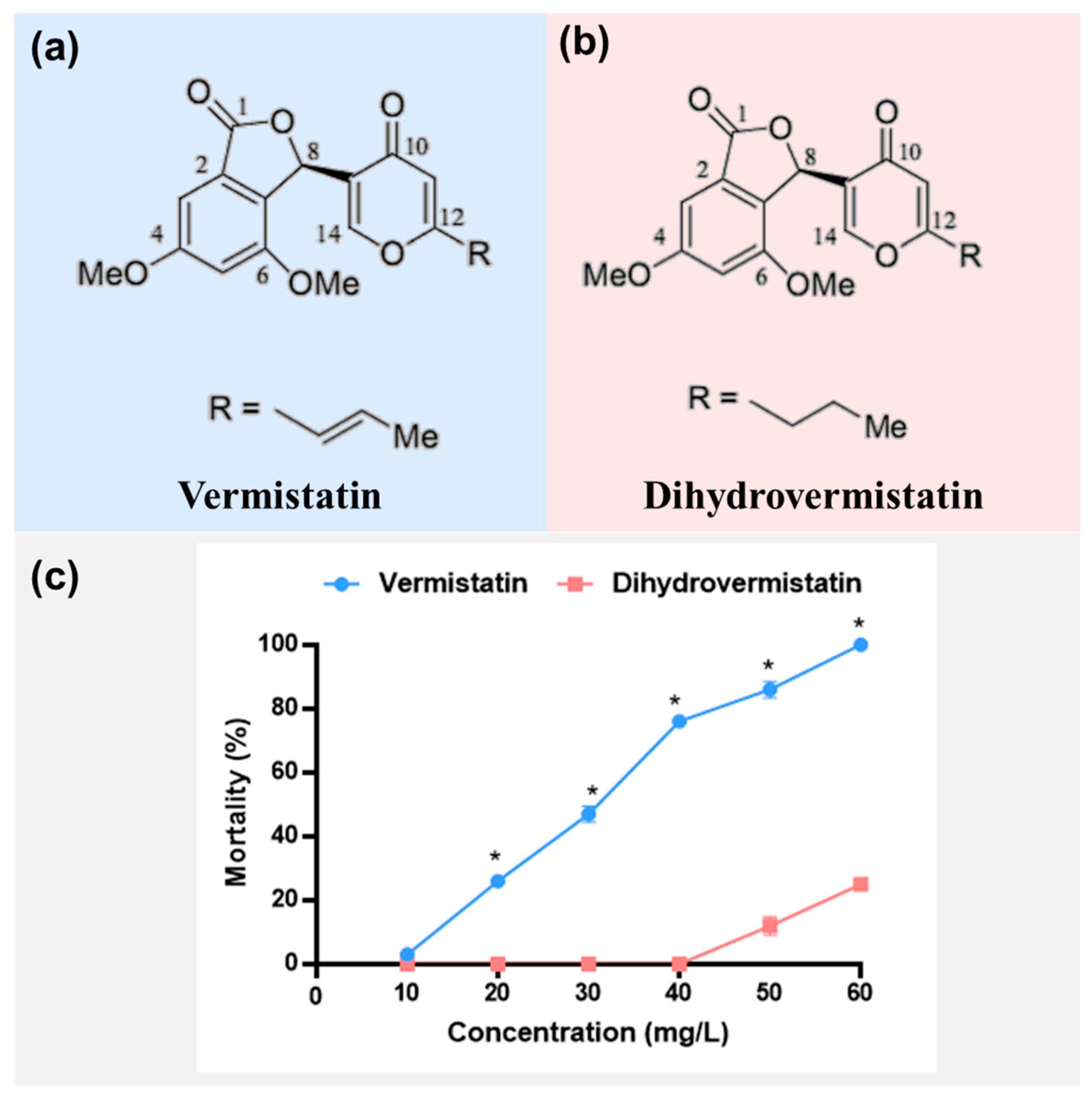
2. Results
3. Discussion
4. Materials and Methods
4.1. Rearing Mosquitos
4.2. Fungal Culture and Metabolite Extraction
4.3. Isolation and Identification of Compounds
- Vermistatin: white crystal; 1H NMR (400 MHz, CDCl3) δH 7.36 (1H, s, H-14), 7.19 (1H, s, H-3), 6.91 (1H, s, H-5), 6.61 (1H, s, H-8), 6.54 (1H, m, H-16), 6.10 (1H, s, H-11), 6.00 (1H, d, J = 15.6 Hz, H-15), 3.81 (3H, s, 4-OCH3), 3.72 (3H, s, 6-OCH3) 1.86 (3H, s, H-17); 13C-NMR (100 MHz, CDCl3) δC 177.4 (C-10), 170.2 (C-12), 163.2 (C-14), 162.3 (C-1), 155.0 (C-4), 154.0 (C-6), 136.1 (C-16), 129.5 (C-2), 127.9 (C-9), 123.6 (C-15), 123.3 (C-7), 113.0 (C-3), 105.3 (C-5), 99.2 (C-11), 73.7 (C-8), 56.1 (4-OCH3), 55.9 (6-OCH3), 18.7 (C-17).
- Dihyrovermistatin: white solid; 1H NMR (400 MHz, CDCl3) δH 7.36 (1H, s, H-14), 6.91 (1H, s, H-3), 6.61 (1H, s, H-5), 6.40 (1H, s, H-8), 6.13 (1H, s, H-11), 3.81 (3H, s, 4-OCH3), 3.72 (3H, s, 6-OCH3), 2.41 (2H, m, H-15), 1.60 (2H, m, H-16), 0.91 (3H, t, J = 7.4 Hz, H-17); 13C-NMR (100 MHz, CDCl3) δC 177.1 (C-12), 170.2 (C-10), 169.5 (C-14), 163.2 (C-1), 155.0 (C-4), 154.5 (C-6), 129.5 (C-2), 127.9 (C-9), 123.6 (C-7), 114.5 (C-12), 105.3 (C-3), 99.2 (C-5), 73.7 (C-8), 56.1 (4-OCH3), 55.9 (6-OCH3), 35.4 (C-15), 20.1 (C-16), 13.5 (C-17).
4.4. Larvicidal Bioassay
4.5. Identification of Fungal Species
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- World Health Organization. World Malaria Report: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020; ISBN 978-92-4-001579-1.
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile Virus Dissemination and Tissue Tropisms in Orally Infected Culex Pipiens Quinquefasciatus. Vector-Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Van Den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and Geographical Expansion of Japanese Encephalitis Virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef]
- Guo, X.X.; Li, C.X.; Deng, Y.Q.; Xing, D.; Liu, Q.M.; Wu, Q.; Sun, A.J.; Dong, Y.D.; Cao, W.C.; Qin, C.F.; et al. Culex Pipiens Quinquefasciatus: A Potential Vector to Transmit Zika Virus. Emerg. Microbes Infect. 2016, 5, e102-5. [Google Scholar] [CrossRef]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The Importance of Vector Control for the Control and Elimination of Vector-Borne Diseases. PLoS Neglected Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The Effect of Malaria Control on Plasmodium Falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Tantely, M.L.; Tortosa, P.; Alout, H.; Berticat, C.; Berthomieu, A.; Rutee, A.; Dehecq, J.S.; Makoundou, P.; Labbé, P.; Pasteur, N.; et al. Insecticide Resistance in Culex Pipiens Quinquefasciatus and Aedes Albopictus Mosquitoes from La Réunion Island. Insect Biochem. Mol. Biol. 2010, 40, 317–324. [Google Scholar] [CrossRef]
- Blackwell, M. The Fungi: 1, 2, 3… 5.1 Million Species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Fungal Kingd. 2017, 5, 79–95. [Google Scholar] [CrossRef]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A Review of the Microbial Production of Bioactive Natural Products and Biologics. Front. Microbiol. 2019, 10, 1404. [Google Scholar] [CrossRef]
- Nielsen, J.C.; Nielsen, J. Development of Fungal Cell Factories for the Production of Secondary Metabolites: Linking Genomics and Metabolism. Synth. Syst. Biotechnol. 2017, 2, 5–12. [Google Scholar] [CrossRef]
- Scholte, E.J.; Knols, B.G.J.; Samson, R.A.; Takken, W. Entomopathogenic Fungi for Mosquito Control: A Review. J. Insect Sci. 2004, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Strasser, H.; Vey, A.; Butt, T.M. Are There Any Risks in Using Entomopathogenic Fungi for Pest Control, with Particular Reference to the Bioactive Metabolites of Metarhizium, Tolypocladium and Beauveria Species? Biocontrol Sci. Technol. 2000, 10, 717–735. [Google Scholar] [CrossRef]
- Niu, G.; Annamalai, T.; Wang, X.; Li, S.; Munga, S.; Niu, G.; Tse-Dinh, Y.C.; Li, J. A Diverse Global Fungal Library for Drug Discovery. PeerJ 2020, 8, e10392. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Hao, Y.; Wang, X.; Gao, J.M.; Li, J. Fungal Metabolite Asperaculane B Inhibits Malaria Infection and Transmission. Molecules 2020, 25, 3018. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Niu, G.; Guan, L.; Ramelow, J.; Zhan, Z.; Zhou, X.; Li, J. Discovery of Mosquitocides from Fungal Extracts through a High-Throughput Cytotoxicity-Screening Approach. Parasites Vectors 2021, 14, 595. [Google Scholar] [CrossRef]
- Zhai, M.M.; Li, J.; Jiang, C.X.; Shi, Y.P.; Di, D.L.; Crews, P.; Wu, Q.X. The Bioactive Secondary Metabolites from Talaromyces Species. Nat. Prod. Bioprospect. 2016, 6, 1–24. [Google Scholar] [CrossRef]
- Lei, L.R.; Gong, L.Q.; Jin, M.Y.; Wang, R.; Liu, R.; Gao, J.; Liu, M.D.; Huang, L.; Wang, G.Z.; Wang, D.; et al. Research Advances in the Structures and Biological Activities of Secondary Metabolites from Talaromyces. Front. Microbiol. 2022, 13, 984801. [Google Scholar] [CrossRef]
- Wu, B.; Ohlendorf, B.; Oesker, V.; Wiese, J.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Acetylcholinesterase Inhibitors from a Marine Fungus Talaromyces Sp. Strain LF458. Mar. Biotechnol. 2015, 17, 110–119. [Google Scholar] [CrossRef]
- Fuska, J.; Uhrín, D.; Proksa, B.; Votický, Z.; Ruppeldt, J. The Structure of Vermistatin, a New Metabolite from Penicillium Vermicula Tum. J. Antibiot. 1986, 39, 1605–1608. [Google Scholar] [CrossRef]
- Fuska, J.; Fuskova, A.P.N. Vermistatin, an Antibiotic with Cytotoxic Effects, Produced from Penicillium-Vermiculatum. Biologia 1979, 34, 735–739. [Google Scholar]
- Ningsih, B.N.S.; Rukachaisirikul, V.; Phongpaichit, S.; Muanprasat, C.; Preedanon, S.; Sakayaroj, J.; Intayot, R.; Jungsuttiwong, S. Talarostatin, a Vermistatin Derivative from the Soil-Derived Fungus Talaromyces Thailandensis PSU-SPSF059. Nat. Prod. Res. 2023, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.K.; Huang, H.R.; She, Z.G.; Cai, J.W.; Lan, L.; Zhang, J.Y.; Fu, L.W.; Vrijmoed, L.L.P.; Lin, Y.C. Structural and Biological Properties of Vermistatin and Two New Vermistatin Derivatives Isolated from the Marine-Mangrove Endophytic Fungus Guignardia Sp. No. 4382. Helv. Chim. Acta 2007, 90, 1925–1931. [Google Scholar] [CrossRef]
- Liu, Z.; Xia, G.; Chen, S.; Liu, Y.; Li, H.; She, Z. Eurothiocin a and B, Sulfur-Containingbenzofurans from a Soft Coral-Derived Fungus Eurotium Rubrum SH-823. Mar. Drugs 2014, 12, 3669–3680. [Google Scholar] [CrossRef]
- He, X.P.; Su, D.Q.; Ding, L.J.; He, S. Secondary Metabolites of Co-Culture of Alternaria Alternate YX-25 and Streptomyces Exfoliatus YX-32. Chinese Tradit. Herb. Drugs 2018, 49, 5772–5779. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhuang, Y.; Kong, F.; Zhang, C.; Zhu, W. Phenolic Polyketides from the Co-Cultivation of Marine-Derived Penicillium Sp. WC-29-5 and Streptomyces Fradiae 007. Mar. Drugs 2014, 12, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Tomoda, H.; Okuda, T.; Wang, H.; Tabata, N.; Masuma, R.; Yamaguchi, Y.; Omura, S. Funicone-Related Compounds, Potentiators of Antifungal Miconazole Activity, Produced by Talaromyces Flavus FKI-0076. J. Antibiot. 2002, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.A.; Stierle, D.B.; Girtsman, T. Caspase-1 Inhibitors from an Extremophilic Fungus That Target Specific Leukemia Cell Lines. J. Nat. Prod. 2012, 75, 344–350. [Google Scholar] [CrossRef]
- Gubiani, J.R.; Wijeratne, E.M.K.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A.L. An Epigenetic Modifier Induces Production of (10′S)-Verruculide B, an Inhibitor of Protein Tyrosine Phosphatases by Phoma Sp. Nov. LG0217, a Fungal Endophyte of Parkinsonia Microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef]
- Xia, X.; Liu, F.; Li, M.; Company, G.M.; Vrijmoed, L.L.P. 1H and 13 C NMR Assignments for 6-Demethylvermistatin and Two Penicillide Derivatives from the Mangrove Fungus Guignardia Sp. (No. 4382) from The. Magn. Reson. Chem. 2018, 46, 693–696. [Google Scholar] [CrossRef]
- Rusman, Y. Isolation of New Secondary Metabolites from Sponge-Associated and Plant-Derived Endophytic Fungi; Cuvillier Verlag: Göttingen, Germany, 2006. [Google Scholar]
- Xu, Q.; Liu, H.; Zhang, L.; Liu, N. Resistance in the Mosquito, Culex Quinquefasciatus, and Possible Mechanisms for Resistance. Pest Manag. Sci. 2005, 61, 1096–1102. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Ma, G.; Yang, F.; Zhan, Z.; Guan, L.; Kuang, W.; Wang, J.; Li, J.; Han, F.; et al. Piperonyl Butoxide Synergizes the Larvicidal Activity of Origanum Vulgare Essential Oil and Its Major Constituents against the Larvae of Aedes Albopictus and Culex Pipiens Quinquefasciatus. J. Asia Pac. Entomol. 2023, 26, 102025. [Google Scholar] [CrossRef]
- Sanei-Dehkordi, A.; Heiran, R.; Moemenbellah-Fard, M.D.; Sayah, S.; Osanloo, M. Nanoliposomes Containing Carvacrol and Carvacrol-Rich Essential Oils as Effective Mosquitoes Larvicides. Bionanoscience 2022, 12, 359–369. [Google Scholar] [CrossRef]
- Komai, S.; Hosoe, T.; Itabashi, T.; Nozawa, K.; Yaguchi, T.; Fukushima, K.; Kawai, K. New Penicillide Derivatives Isolated from Penicillium Simplicissimum. J. Nat. Med. 2006, 60, 185–190. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: Geneva, Switzerland, 2005.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | LC50 | 95% CL | LC90 | 95% CL | Regression Equation | x2 Value | p |
|---|---|---|---|---|---|---|---|
| 101H2 | 31.898 | 26.957~37.137 | 93.917 | 76.049~126.177 | y = −4.109 + 2.733x | 45.713 | 0.002 |
| 58A7 | 44.269 | 38.391~50.672 | 102.008 | 84.596~133.503 | y = −5.819 + 3.535x | 53.634 | 0.000 |
| Fraction 14 | 12.128 | 10.894~13.395 | 22.160 | 19.432~26.644 | y = −5.306 + 4.896x | 34.659 | 0.010 |
| Vermistatin | 28.132 | 26.325~29.912 | 52.001 | 47.870~57.588 | y = −6.961 + 4.803x | 21.772 | 0.474 |
| Dihydrovermistatin | 83.869 | 71.655~118.768 | 145.325 | 106.911~295.192 | y = −10.326 + 5.368x | 8.960 | 0.994 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Xu, Z.; Liu, Y.; Yang, F.; Guan, L.; Yang, J.; Li, J.; Niu, G.; Li, J.; Jin, L. Talaromyces sp. Ethyl Acetate Crude Extract as Potential Mosquitocide to Control Culex pipiens quinquefasciatus. Molecules 2023, 28, 6642. https://doi.org/10.3390/molecules28186642
Chen J, Xu Z, Liu Y, Yang F, Guan L, Yang J, Li J, Niu G, Li J, Jin L. Talaromyces sp. Ethyl Acetate Crude Extract as Potential Mosquitocide to Control Culex pipiens quinquefasciatus. Molecules. 2023; 28(18):6642. https://doi.org/10.3390/molecules28186642
Chicago/Turabian StyleChen, Junhui, Zhiyong Xu, Yangqing Liu, Feiying Yang, Limei Guan, Jian Yang, Jianghuai Li, Guodong Niu, Jun Li, and Liang Jin. 2023. "Talaromyces sp. Ethyl Acetate Crude Extract as Potential Mosquitocide to Control Culex pipiens quinquefasciatus" Molecules 28, no. 18: 6642. https://doi.org/10.3390/molecules28186642