
Assessing the Effects of Thiazole-Carboxamide Derivatives on the Biophysical Properties of AMPA Receptor Complexes as a Potential Neuroprotective Agent
 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. TC Compounds Potently Inhibit AMPA Receptor Subunits in Transfected HEK293T Cells
2.2. Modulation of AMPA Receptor Kinetics by TC Compounds
3. Discussion
4. Materials and Methods
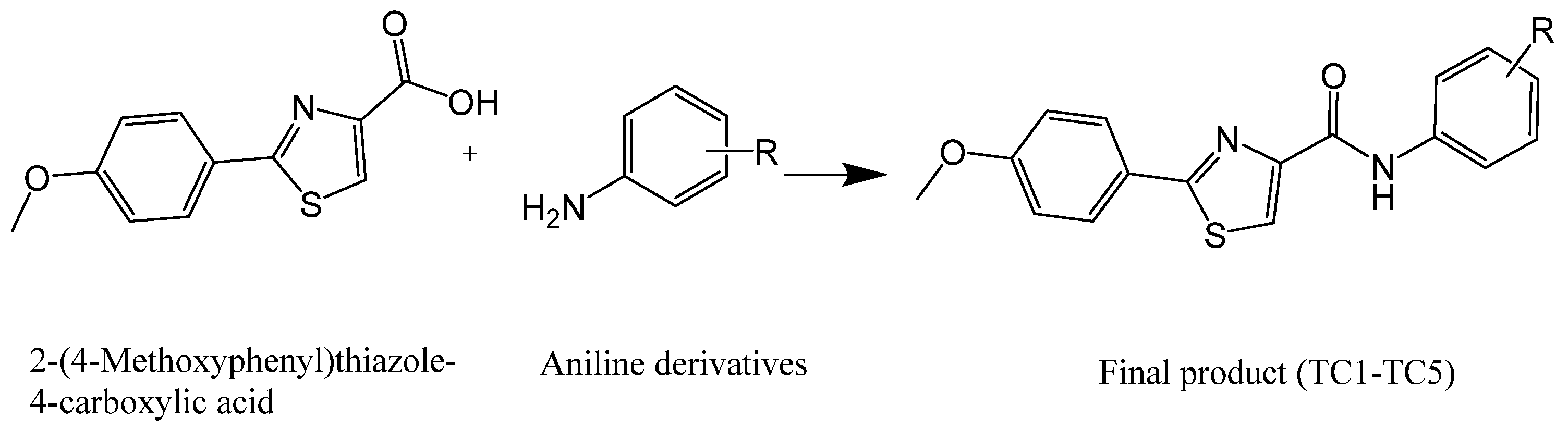
4.1. Chemistry
4.2. Electrophysiological Insights
4.2.1. Preparation of Plasmid DNA
4.2.2. Expression and Recording in HEK293T Cells
4.2.3. Inhibition Assessment and Compound Administration
4.2.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sohal, V.S.; Rubenstein, J.L.R. Excitation-inhibition balance as a framework for investigating mechanisms in neuropsychiatric disorders. Mol. Psychiatry 2019, 24, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Uzunova, G.; Pallanti, S.; Hollander, E. Excitatory/inhibitory imbalance in autism spectrum disorders: Implications for interventions and therapeutics. World J. Biol. Psychiatry 2016, 17, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Van den Bos, M.A.J.; Higashihara, M.; Geevasinga, N.; Menon, P.; Kiernan, M.C.; Vucic, S. Imbalance of cortical facilitatory and inhibitory circuits underlies hyperexcitability in ALS. Neurology 2018, 91, e1669–e1676. [Google Scholar] [CrossRef] [PubMed]
- Selten, M.; Van Bokhoven, H.; Kasri, N.N. Inhibitory control of the excitatory/inhibitory balance in psychiatric disorders. F1000Research 2018, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Yung, D.; Li, J.-Y. Organization of ipsilateral excitatory and inhibitory pathways in the human motor cortex. J. Neurophysiol. 2003, 89, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, D.J.; Good, M.A.; Seeburg, P.H.; Sprengel, R.; Rawlins, J.N.P.; Bannerman, D.M. The role of the GluR-A (GluR1) AMPA receptor subunit in learning and memory. Prog. Brain Res. 2008, 169, 159–178. [Google Scholar] [PubMed]
- Shimshek, D.R.; Bus, T.; Schupp, B.; Jensen, V.; Marx, V.; Layer, L.E.; Köhr, G.; Sprengel, R. Different forms of AMPA receptor mediated LTP and their correlation to the spatial working memory formation. Front. Mol. Neurosci. 2017, 10, 214. [Google Scholar] [CrossRef]
- Mitsushima, D.; Ishihara, K.; Sano, A.; Kessels, H.W.; Takahashi, T. Contextual learning requires synaptic AMPA receptor delivery in the hippocampus. Proc. Natl. Acad. Sci. USA 2011, 108, 12503–12508. [Google Scholar] [CrossRef]
- Qneibi, M.; Bdir, S.; Bdair, M.; Aldwaik, S.A.; Sandouka, D.; Heeh, M.; Idais, T.I. AMPA receptor neurotransmission and therapeutic applications: A comprehensive review of their multifaceted modulation. Eur. J. Med. Chem. 2024, 266, 116151. [Google Scholar] [CrossRef]
- Niescier, R.F.; Lin, Y.-C. The potential role of AMPA receptor trafficking in Autism and other neurodevelopmental conditions. Neuroscience 2021, 479, 180–191. [Google Scholar] [CrossRef]
- Babaei, P. NMDA and AMPA receptors dysregulation in Alzheimer’s disease. Eur. J. Pharmacol. 2021, 908, 174310. [Google Scholar] [CrossRef]
- Egbenya, D.L.; Hussain, S.; Lai, Y.-C.; Xia, J.; Anderson, A.E.; Davanger, S. Changes in synaptic AMPA receptor concentration and composition in chronic temporal lobe epilepsy. Mol. Cell. Neurosci. 2018, 92, 93–103. [Google Scholar] [CrossRef]
- Kerr, M.P. The impact of epilepsy on patients’ lives. Acta Neurol. Scand. 2012, 126, 1–9. [Google Scholar] [CrossRef]
- Ostendorf, A.P.; Gedela, S. (Eds.) Effect of Epilepsy on Families, Communities, and Society; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Vazquez, B.; Devinsky, O. Epilepsy and anxiety. Epilepsy Behav. 2003, 4, 20–25. [Google Scholar] [CrossRef]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef]
- Mayer, M.L.; Armstrong, N. Structure and function of glutamate receptor ion channels. Annu. Rev. Physiol. 2004, 66, 161–181. [Google Scholar] [CrossRef]
- Niciu, M.J.; Kelmendi, B.; Sanacora, G. Overview of glutamatergic neurotransmission in the nervous system. Pharmacol. Biochem. Behav. 2012, 100, 656–664. [Google Scholar] [CrossRef]
- Choi, D.W. Excitotoxic cell death. J. Neurobiol. 1992, 23, 1261–1276. [Google Scholar] [CrossRef]
- Zappala, M.; Grasso, S.; Micale, N.; Polimeni, S.; De Micheli, C. Synthesis and structure-activity relationships of 2,3-benzodiazepines as AMPA receptor antagonists. Mini Rev. Med. Chem. 2001, 1, 243–253. [Google Scholar] [CrossRef]
- Qneibi, M.; Jumaa, H.; Bdir, S.; Al-Maharik, N. Electrophysiological Assessment of newly synthesized 2,3-Benzodiazepine derivatives for inhibiting the AMPA receptor Channel. Molecules 2023, 28, 6067. [Google Scholar] [CrossRef]
- Qneibi, M.S.; Micale, N.; Grasso, S.; Niu, L. Mechanism of inhibition of GluA2 AMPA receptor channel opening by 2,3-benzodiazepine derivatives: Functional consequences of replacing a 7,8-methylenedioxy with a 7,8-ethylenedioxy moiety. Biochemistry 2012, 51, 1787–1795. [Google Scholar] [CrossRef]
- Patel, N.C. Perampanel (Fycompa): AMPA receptor antagonist for the treatment of seizure. Innov. Drug Synth. 2015, 14, 271–282. [Google Scholar]
- Steinhoff, B.J. The AMPA receptor antagonist perampanel in the adjunctive treatment of partial-onset seizures: Clinical trial evidence and experience. Ther. Adv. Neurol. Disord. 2015, 8, 137–147. [Google Scholar] [CrossRef]
- Guseynov, A.D.; Pisarev, S.A.; Shulga, D.A.; Palyulin, V.A.; Fedorov, M.V.; Karlov, D.S. Computational characterization of the glutamate receptor antagonist perampanel and its close analogs: Density functional exploration of conformational space and molecular docking study. J. Mol. Model. 2019, 25, 312. [Google Scholar] [CrossRef]
- Hawash, M.; Jaradat, N.; Abualhasan, M.; Şüküroğlu, M.K.; Qaoud, M.T.; Kahraman, D.C.; Daraghmeh, H.; Maslamani, L.; Sawafta, M.; Ratrout, A.; et al. Design, synthesis, molecular docking studies and biological evaluation of thiazole carboxamide derivatives as COX inhibitors. BMC Chem. 2023, 17, 11. [Google Scholar] [CrossRef]
- Bryson, H.M.; Fulton, B.; Benfield, P. Riluzole: A review of its pharmacodynamic and pharmacokinetic properties and therapeutic potential in amyotrophic lateral sclerosis. Drugs 1996, 52, 549–563. [Google Scholar] [CrossRef]
- Hawash, M. Thiazole Derivatives as Modulators of GluA2 AMPA Receptors: Potent Allosteric Effects and Neuroprotective Potential. Biomolecules 2023, 13, 1694. [Google Scholar] [CrossRef]
- Rameau, G.A.; Tukey, D.S.; Garcin-Hosfield, E.D.; Titcombe, R.F.; Misra, C.; Khatri, L.; Getzoff, E.D.; Ziff, E.B. Biphasic coupling of neuronal nitric oxide synthase phosphorylation to the NMDA receptor regulates AMPA receptor trafficking and neuronal cell death. J. Neurosci. 2007, 27, 3445–3455. [Google Scholar] [CrossRef]
- Singh, S.; Loke, Y.K.; Furberg, C.D. Thiazolidinediones and heart failure: A teleo-analysis. Diabetes Care 2007, 30, 2148–2153. [Google Scholar] [CrossRef]
- Loke, Y.K.; Singh, S.; Furberg, C.D. Long-term use of thiazolidinediones and fractures in type 2 diabetes: A meta-analysis. CMAJ 2009, 180, 32–39. [Google Scholar] [CrossRef]
- Wallach, J.D.; Wang, K.; Zhang, A.D.; Cheng, D.; Nardini, H.K.G.; Lin, H.; Bracken, M.B.; Desai, M.; Krumholz, H.M.; Ross, J.S. Updating insights into Rosiglitazone and cardiovascular risk through shared data: Individual patient and summary level meta-analyses. BMJ 2020, 368, l7078. [Google Scholar] [CrossRef]
- Nissen, S.E. The Rise and Fall of Rosiglitazone; Oxford University Press: Oxford, UK, 2010; pp. 773–776. [Google Scholar]
- Zarate, C.A.; Manji, H.K. The Role of AMPA receptor modulation in the treatment of neuropsychiatric diseases. Exp. Neurol. 2008, 211, 7–10. [Google Scholar] [CrossRef]
- Fontana, A.C.; Poli, A.N.; Gour, J.; Srikanth, Y.V.; Anastasi, N.; Ashok, D.; Khatiwada, A.; Reeb, K.L.; Cheng, M.H.; Bahar, I.; et al. Synthesis and Structure—Activity Relationships for Glutamate Transporter Allosteric Modulators. J. Med. Chem. 2024, 67, 6119–6143. [Google Scholar] [CrossRef]
- Qneibi, M.; Hawash, M.; Gümüş, M.; Çapan, İ.; Sert, Y.; Bdir, S.; Koca, İ.; Bdair, M. Deciphering the Biophysical Properties of Ion Channel Gating Pores by Coumarin–Benzodiazepine Hybrid Derivatives: Selective AMPA Receptor Antagonists. Mol. Neurobiol. 2023, 1–12. [Google Scholar] [CrossRef]
- Charsouei, S.; Jabalameli, M.R.; Karimi-Moghadam, A. Molecular insights into the role of AMPA receptors in the synaptic plasticity, pathogenesis and treatment of epilepsy: Therapeutic potentials of perampanel and antisense oligonucleotide (ASO) technology. Acta Neurol. Belg. 2020, 120, 531–544. [Google Scholar] [CrossRef]
- Qneibi, M.; Jaradat, N.; Hawash, M.; Olgac, A.; Emwas, N. Ortho versus Meta Chlorophenyl-2,3-Benzodiazepine Analogues: Synthesis, Molecular Modeling, and Biological Activity as AMPAR Antagonists. ACS Omega 2020, 5, 3588–3595. [Google Scholar] [CrossRef]
- Deng, X.-Q.; Song, M.-X.; Gong, G.-H.; Wang, S.-B.; Quan, Z.-S. Synthesis and anticonvulsant evaluation of some new 6-(substituted-phenyl) thiazolo [3,2-b][1,2,4] triazole derivatives in mice. Iran. J. Pharm. Res. IJPR 2014, 13, 459. [Google Scholar]
- Xi, N.; Bo, Y.; Doherty, E.M.; Fotsch, C.; Gavva, N.R.; Han, N.; Hungate, R.W.; Klionsky, L.; Liu, Q.; Tamir, R.; et al. Synthesis and evaluation of thiazole carboxamides as vanilloid receptor 1 (TRPV1) antagonists. Bioorg. Med. Chem. Lett. 2005, 15, 5211–5217. [Google Scholar] [CrossRef]
- Malik, S.; Bahare, R.S.; Khan, S.A. Design, synthesis and anticonvulsant evaluation of N-(benzo [d] thiazol-2-ylcarbamoyl)-2-methyl-4-oxoquinazoline-3(4H)-carbothioamide derivatives: A hybrid pharmacophore approach. Eur. J. Med. Chem. 2013, 67, 1–13. [Google Scholar] [CrossRef]
- Qneibi, M.; Hamed, O.; Jaradat, N.; Hawash, M.; Al-Kerm, R.; Al-Kerm, R.; Sobuh, S.; Tarazi, S. The AMPA receptor biophysical gating properties and binding site: Focus on novel curcumin-based diazepines as non-competitive antagonists. Bioorg. Chem. 2021, 116, 105406. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qneibi, M.; Hawash, M.; Bdir, S.; Bdair, M.; Aldwaik, S.A. Assessing the Effects of Thiazole-Carboxamide Derivatives on the Biophysical Properties of AMPA Receptor Complexes as a Potential Neuroprotective Agent. Molecules 2024, 29, 3232. https://doi.org/10.3390/molecules29133232
Qneibi M, Hawash M, Bdir S, Bdair M, Aldwaik SA. Assessing the Effects of Thiazole-Carboxamide Derivatives on the Biophysical Properties of AMPA Receptor Complexes as a Potential Neuroprotective Agent. Molecules. 2024; 29(13):3232. https://doi.org/10.3390/molecules29133232
Chicago/Turabian StyleQneibi, Mohammad, Mohammed Hawash, Sosana Bdir, Mohammad Bdair, and Samia Ammar Aldwaik. 2024. "Assessing the Effects of Thiazole-Carboxamide Derivatives on the Biophysical Properties of AMPA Receptor Complexes as a Potential Neuroprotective Agent" Molecules 29, no. 13: 3232. https://doi.org/10.3390/molecules29133232