
Hyperpolarized 13C NMR Reveals Pathway Regulation in Lactococcus lactis and Metabolic Similarities and Differences Across the Tree of Life
 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
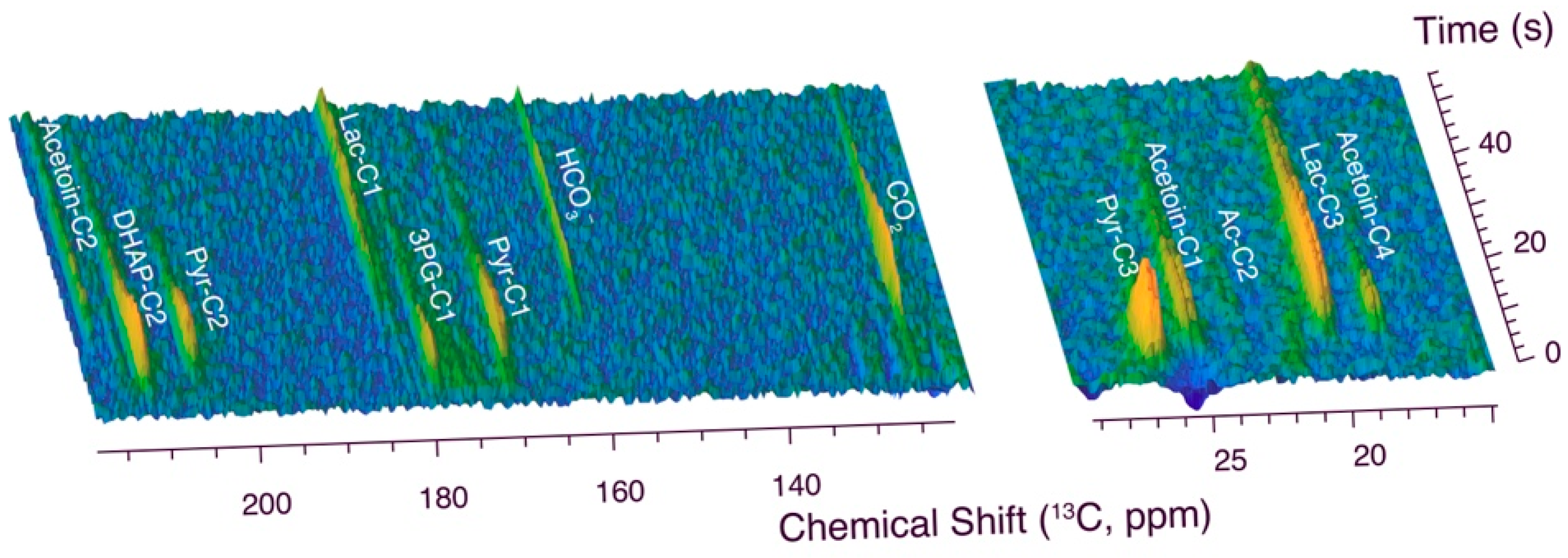
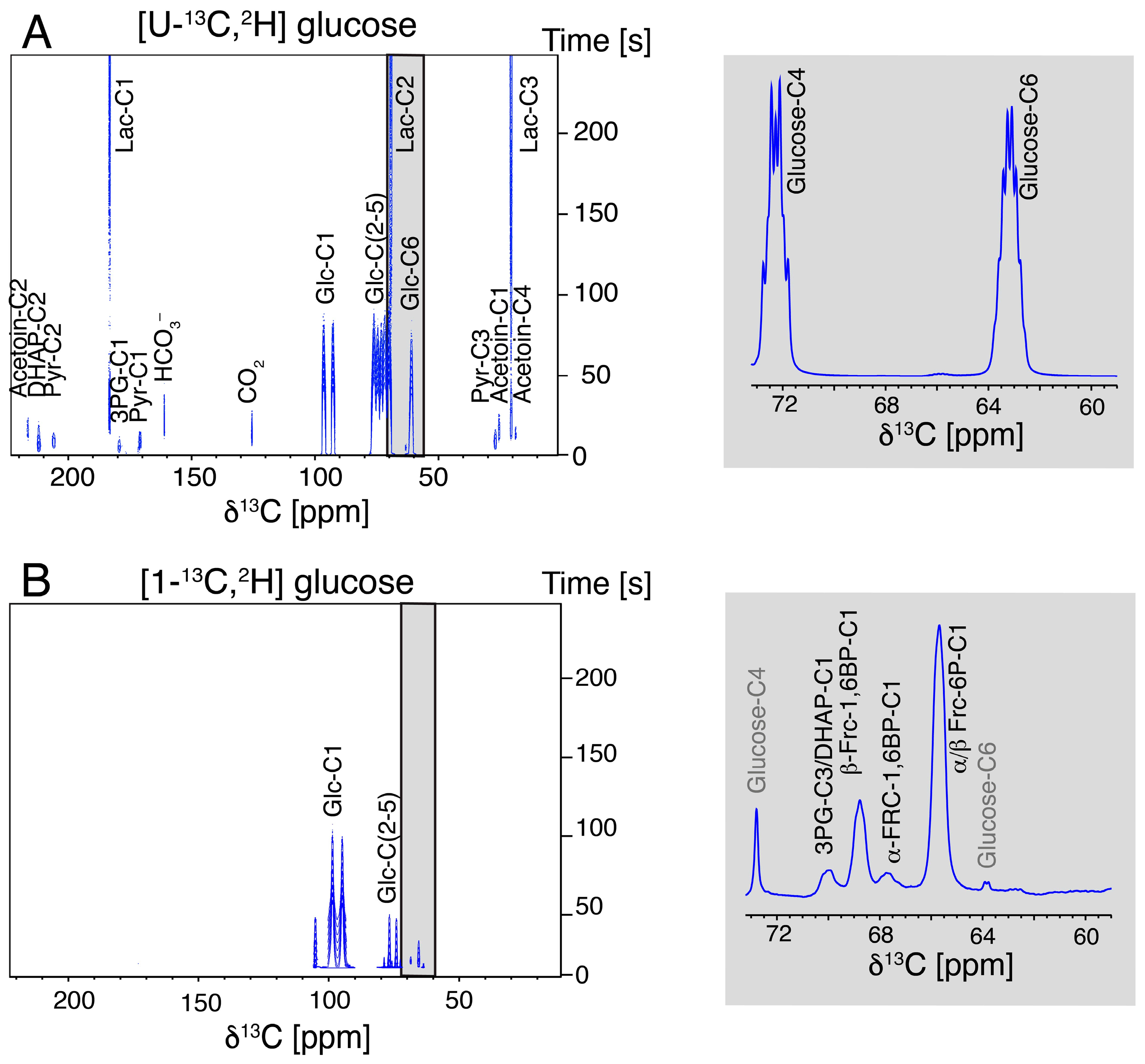
2.1. DNP NMR Using Hyperpolarized [U-13C,2H]glucose in L. lactis
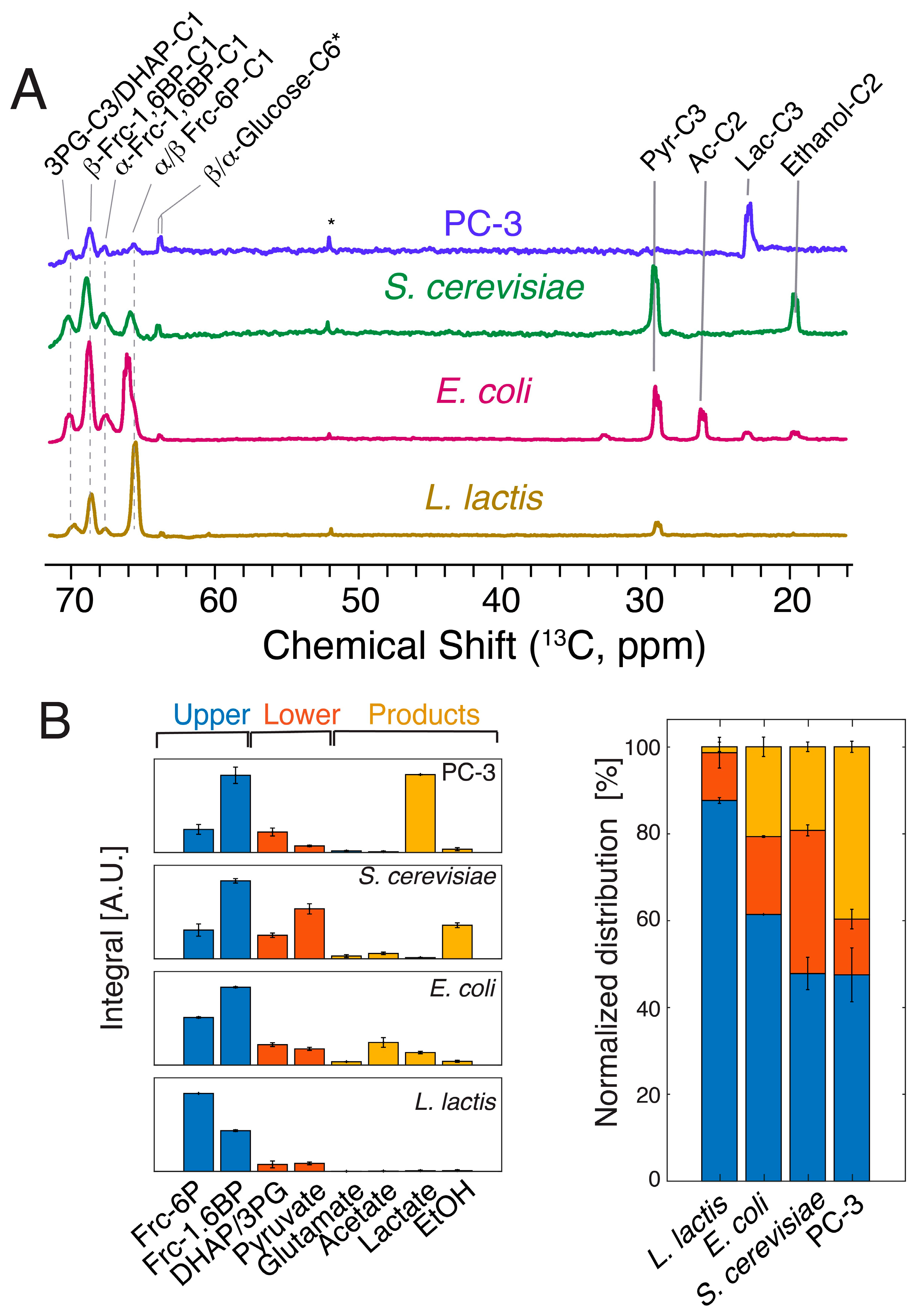
2.2. Hyperpolarized [1-13C,2H]glucose Probes Upper Glycolysis in L. lactis and Other Model Cell Lines
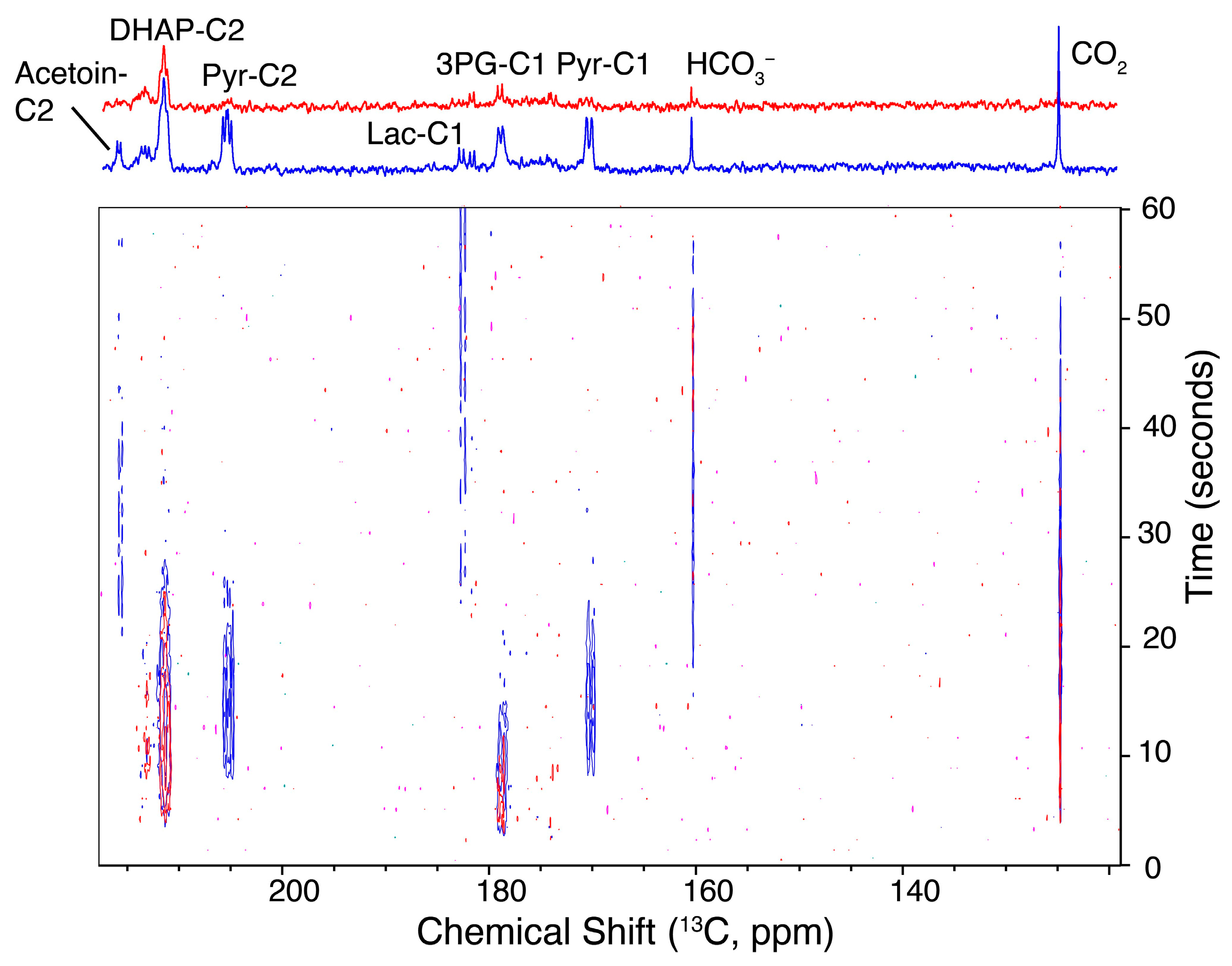
2.3. Influx and Passage through Upper Glycolysis Limits Conversion through the Entire Pathway
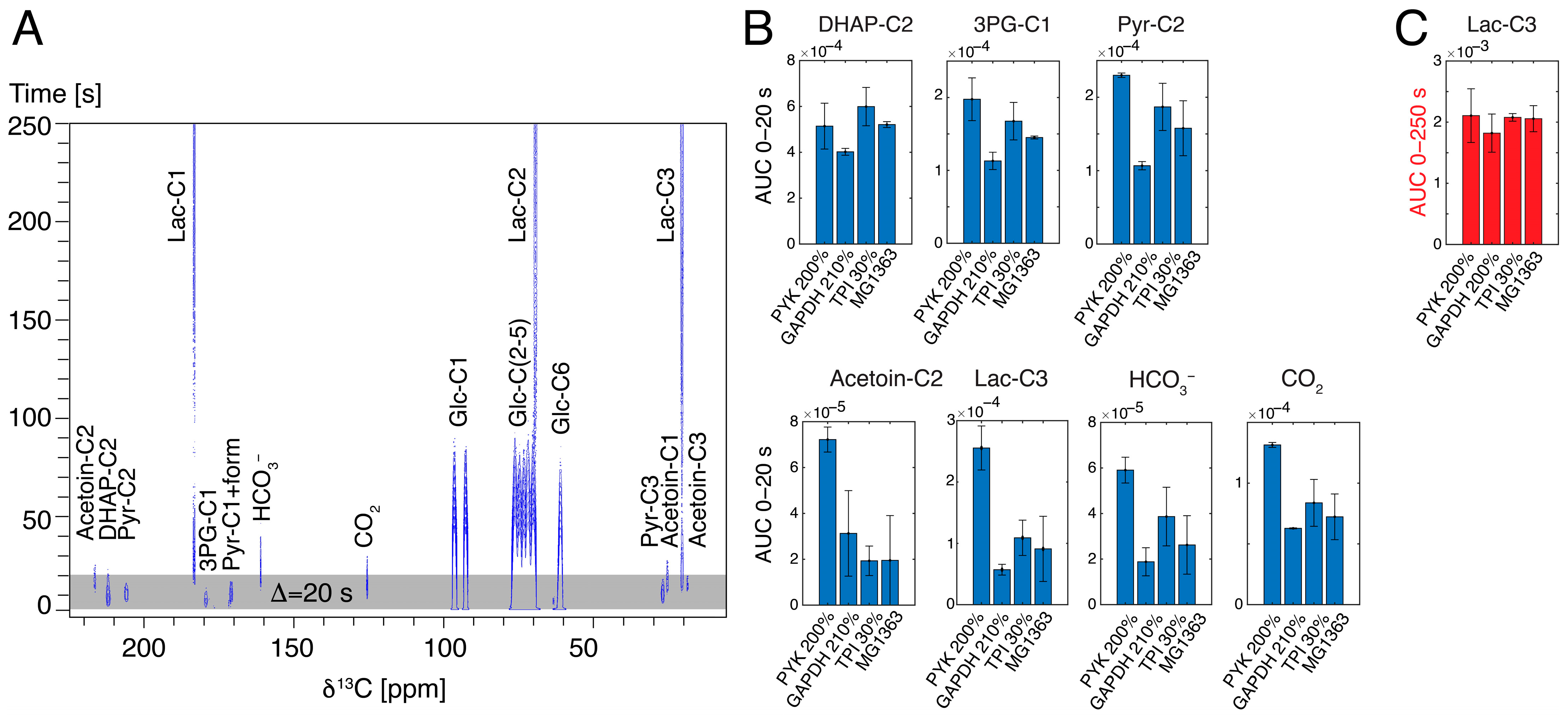
2.4. Hyperpolarized NMR Using Mutant Strains Shows Altered Kinetics on a Timescale of Seconds, Despite Similar Kinetics on a Timescale of Minutes
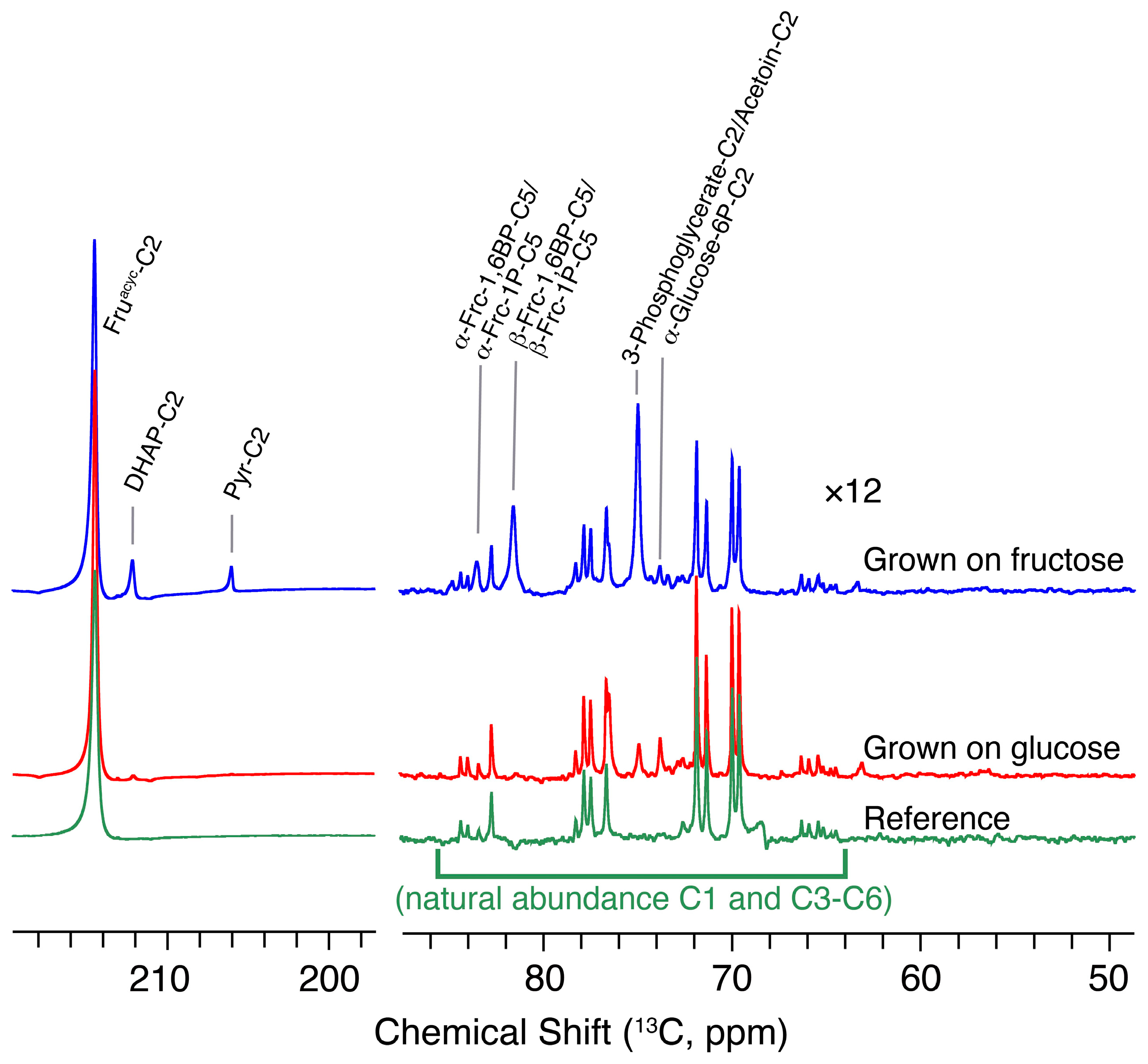
2.5. In-Cell Spectroscopic Visualization of Response to Enzymatic Inhibitor and to Altered Carbon Source
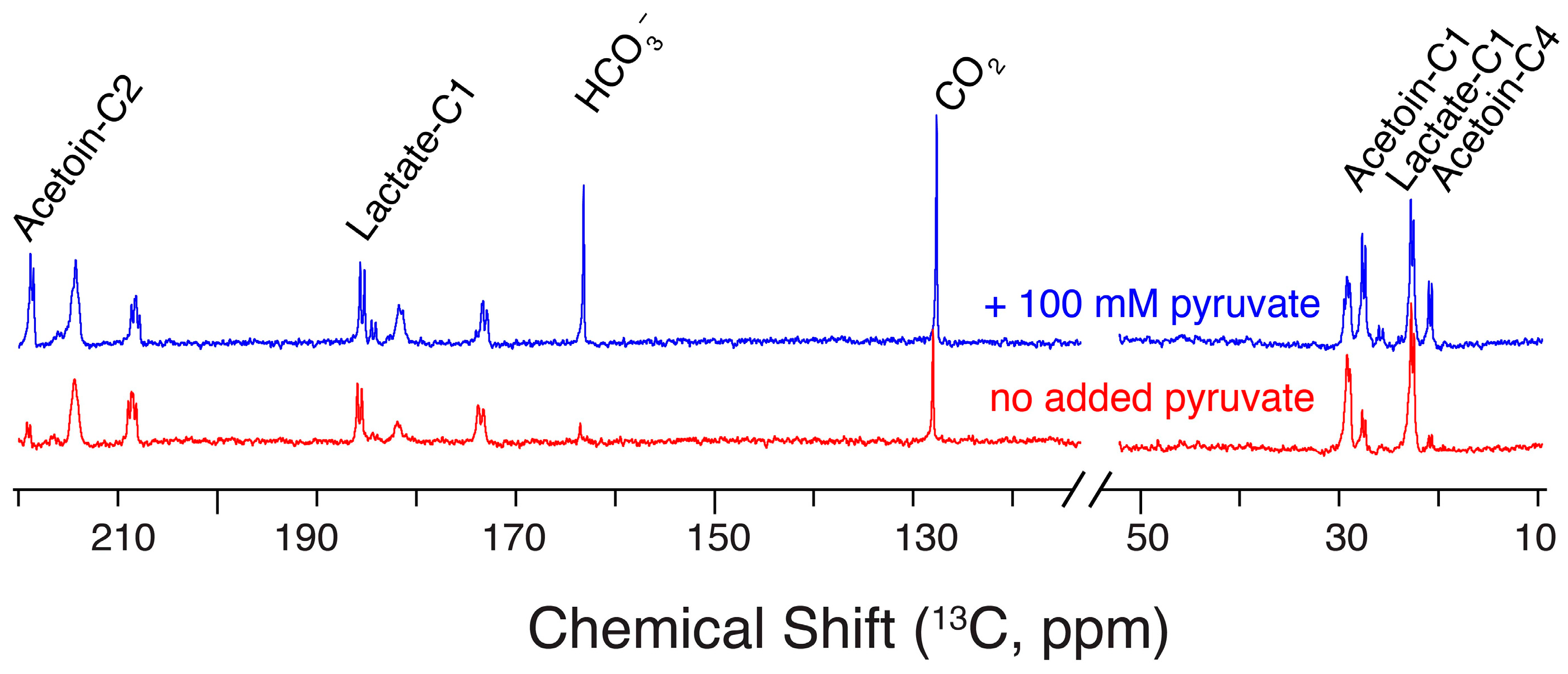
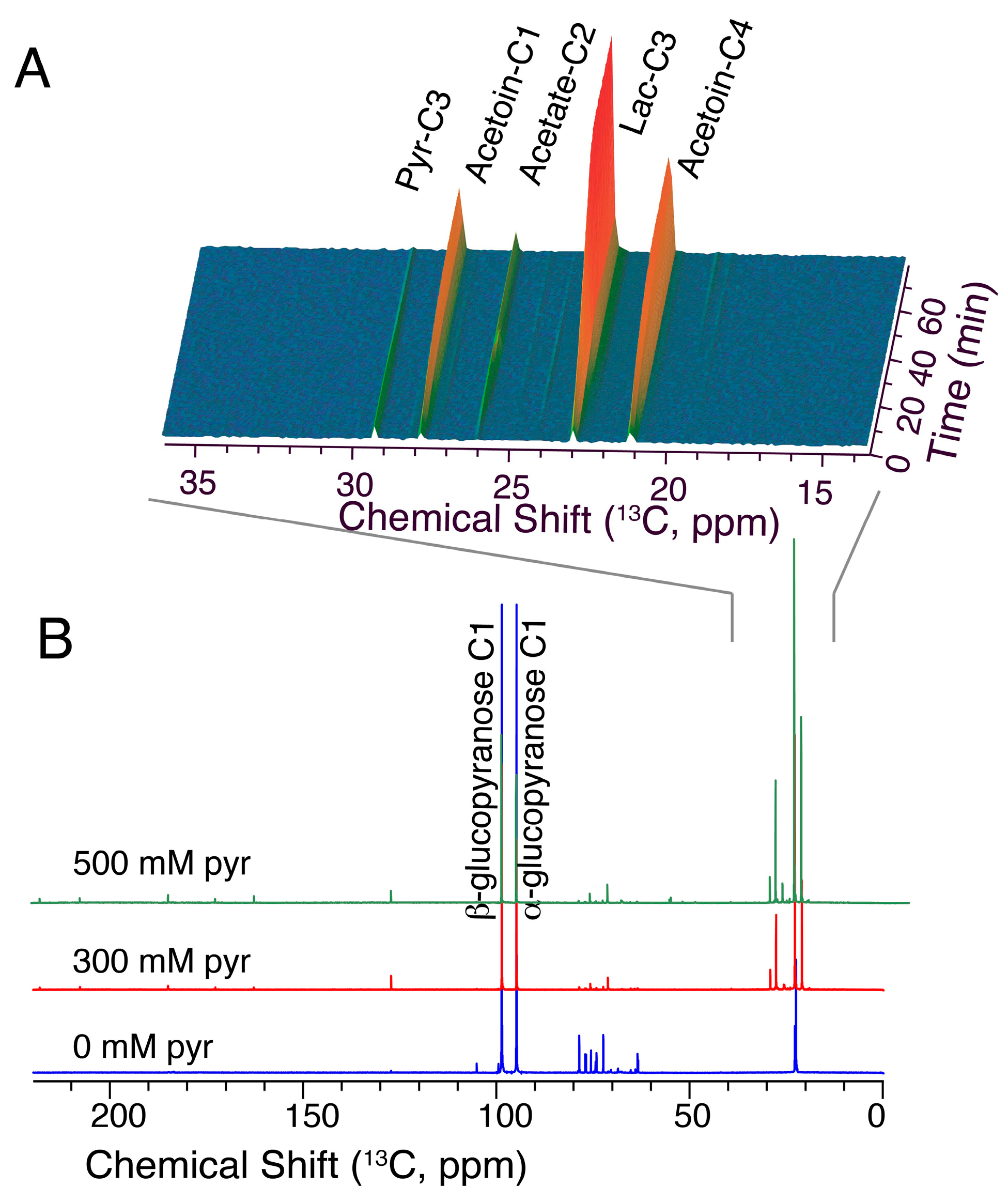
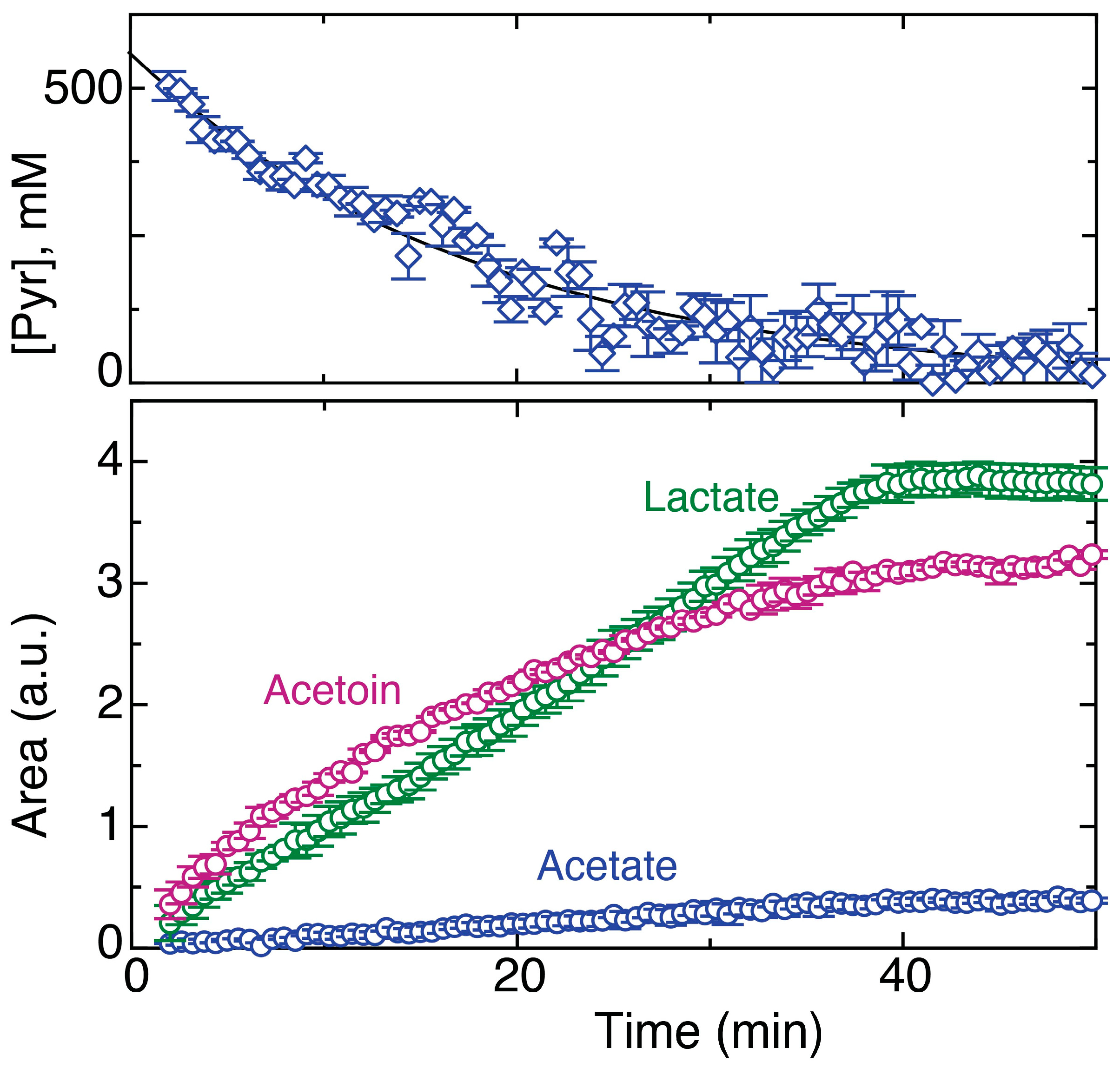
2.6. Glucose/Pyruvate Substrate Mixtures Direct Glucose Carbon towards Acetoin
3. Materials and Methods
3.1. Chemicals and Cells
3.2. Cell Culture
3.3. Inhibition of Glycolysis with Iodoacetate
3.4. Fructose as Carbon Source and dDNP Substrate
3.5. Pyruvate/Glucose Substrate Mixtures
3.6. NMR Experiments
3.7. DNP
3.8. dDNP NMR
3.9. Data Analysis and Fitting
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Park, J.O.; Rubin, S.A.; Xu, Y.-F.; Amador-Noguez, D.; Fan, J.; Shlomi, T.; Rabinowitz, J.D. Metabolite Concentrations, Fluxes and Free Energies Imply Efficient Enzyme Usage. Nat. Chem. Biol. 2016, 12, 482–489. [Google Scholar] [CrossRef]
- Park, J.O.; Tanner, L.B.; Wei, M.H.; Khana, D.B.; Jacobson, T.B.; Zhang, Z.; Rubin, S.A.; Li, S.H.-J.; Higgins, M.B.; Stevenson, D.M.; et al. Near-Equilibrium Glycolysis Supports Metabolic Homeostasis and Energy Yield. Nat. Chem. Biol. 2019, 15, 1001–1008. [Google Scholar] [CrossRef]
- Meier, S.; Jensen, P.R.; Duus, J.Ø. Real-Time Detection of Central Carbon Metabolism in Living Escherichia Coli and Its Response to Perturbations. FEBS Lett. 2011, 585, 3133–3138. [Google Scholar] [CrossRef] [PubMed]
- Sannelli, F.; Jensen, P.R.; Meier, S. In-Cell NMR Approach for Real-Time Exploration of Pathway Versatility: Substrate Mixtures in Nonengineered Yeast. Anal. Chem. 2023, 95, 7262–7270. [Google Scholar] [CrossRef]
- Bar-Even, A.; Flamholz, A.; Noor, E.; Milo, R. Rethinking Glycolysis: On the Biochemical Logic of Metabolic Pathways. Nat. Chem. Biol. 2012, 8, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Locasale, J.W. Thermodynamic Constraints on the Regulation of Metabolic Fluxes. J. Biol. Chem. 2018, 293, 19725–19739. [Google Scholar] [CrossRef]
- Ardenkjær-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in Signal-to-Noise Ratio of >10,000 Times in Liquid-State NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158–10163. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Sannelli, F.; Stauning, L.T.; Meier, S. Enhanced 13C NMR Detects Extended Reaction Networks in Living Cells. Chem. Commun. 2021, 57, 10572–10575. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.; Degani, H.; Frydman, L. Hyperpolarized 13C NMR Studies of Glucose Metabolism in Living Breast Cancer Cell Cultures. NMR Biomed. 2013, 26, 1831–1843. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.B.; Serrao, E.M.; Kennedy, B.W.C.; Hu, D.-E.; Kettunen, M.I.; Brindle, K.M. Magnetic Resonance Imaging of Tumor Glycolysis Using Hyperpolarized 13C-Labeled Glucose. Nat. Med. 2014, 20, 93–97. [Google Scholar] [CrossRef]
- Cavallari, E.; Carrera, C.; Sorge, M.; Bonne, G.; Muchir, A.; Aime, S.; Reineri, F. The 13C Hyperpolarized Pyruvate Generated by ParaHydrogen Detects the Response of the Heart to Altered Metabolism in Real Time. Sci. Rep. 2018, 8, 8366. [Google Scholar] [CrossRef]
- Pravdivtsev, A.N.; Buntkowsky, G.; Duckett, S.B.; Koptyug, I.V.; Hövener, J. Parahydrogen—Induced Polarization of Amino Acids. Angew. Chem. Int. Ed. 2021, 60, 23496–23507. [Google Scholar] [CrossRef]
- Stewart, N.J.; Nakano, H.; Sugai, S.; Tomohiro, M.; Kase, Y.; Uchio, Y.; Yamaguchi, T.; Matsuo, Y.; Naganuma, T.; Takeda, N.; et al. Hyperpolarized 13 C Magnetic Resonance Imaging of Fumarate Metabolism by Parahydrogen-induced Polarization: A Proof-of-Concept in Vivo Study. ChemPhysChem 2021, 22, 915–923. [Google Scholar] [CrossRef]
- Miclet, E.; Abergel, D.; Bornet, A.; Milani, J.; Jannin, S.; Bodenhausen, G. Toward Quantitative Measurements of Enzyme Kinetics by Dissolution Dynamic Nuclear Polarization. J. Phys. Chem. Lett. 2014, 5, 3290–3295. [Google Scholar] [CrossRef]
- Bornet, A.; Ji, X.; Mammoli, D.; Vuichoud, B.; Milani, J.; Bodenhausen, G.; Jannin, S. Long-Lived States of Magnetically Equivalent Spins Populated by Dissolution-DNP and Revealed by Enzymatic Reactions. Chem. A Eur. J 2014, 20, 17113–17118. [Google Scholar] [CrossRef] [PubMed]
- Theillet, F.-X.; Luchinat, E. In-Cell NMR: Why and How? Prog. Nucl. Magn. Reson. Spectrosc. 2022, 132–133, 1–112. [Google Scholar] [CrossRef]
- Keshari, K.R.; Wilson, D.M.; Chen, A.P.; Bok, R.; Larson, P.E.Z.; Hu, S.; Criekinge, M.V.; Macdonald, J.M.; Vigneron, D.B.; Kurhanewicz, J. Hyperpolarized [2-13C]-Fructose: A Hemiketal DNP Substrate for In Vivo Metabolic Imaging. J. Am. Chem. Soc. 2009, 131, 17591–17596. [Google Scholar] [CrossRef]
- Karlsson, M.; Jensen, P.R.; Duus, J.Ø.; Meier, S.; Lerche, M.H. Development of Dissolution DNP-MR Substrates for Metabolic Research. Appl. Magn. Reson. 2012, 43, 223–236. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, Z.; He, L.; Li, C.; Liu, M. NMR Spectroscopy for Metabolomics in the Living System: Recent Progress and Future Challenges. Anal. Bioanal. Chem. 2024, 416, 2319–2334. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Jensen, P.R.; Duus, J.Ø. Direct Observation of Metabolic Differences in Living Escherichia Coli Strains K-12 and BL21. ChemBioChem 2012, 13, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Wang, K.-C.; Sannelli, F.; Hoof, J.B.; Wendland, J.; Jensen, P.R. Visualizing Metabolism in Biotechnologically Important Yeasts with dDNP NMR Reveals Evolutionary Strategies and Glycolytic Logic. Anal. Chem. 2024, 96, 10901–10910. [Google Scholar] [CrossRef] [PubMed]
- Sannelli, F.; Wang, K.-C.; Jensen, P.R.; Meier, S. Rapid Probing of Glucose Influx into Cancer Cell Metabolism: Using Adjuvant and a pH-Dependent Collection of Central Metabolites to Improve in-Cell D-DNP NMR. Anal. Methods 2023, 15, 4870–4882. [Google Scholar] [CrossRef] [PubMed]
- Song, A.A.-L.; In, L.L.A.; Lim, S.H.E.; Rahim, R.A. A Review on Lactococcus lactis: From Food to Factory. Microb. Cell Fact. 2017, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Pool, W.A.; Kok, J.; Kuipers, O.P.; Santos, H. Overview on Sugar Metabolism and Its Control in Lactococcus lactis—The Input from in Vivo NMR. FEMS Microbiol. Rev. 2005, 29, 531–554. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Ramos, A.; Shearman, C.; Gasson, M.J.; Almeida, J.S.; Santos, H. Metabolic Characterization of Lactococcus lactis Deficient in Lactate Dehydrogenase Using in Vivo 13C-NMR. Eur. J. Biochem. 2000, 267, 3859–3868. [Google Scholar] [CrossRef]
- Ebrahimi, P.; Larsen, F.H.; Jensen, H.M.; Vogensen, F.K.; Engelsen, S.B. Real-Time Metabolomic Analysis of Lactic Acid Bacteria as Monitored by in Vitro NMR and Chemometrics. Metabolomics 2016, 12, 77. [Google Scholar] [CrossRef]
- Sriram, R.; Sun, J.; Villanueva-Meyer, J.; Mutch, C.; De Los Santos, J.; Peters, J.; Korenchan, D.E.; Neumann, K.; Van Criekinge, M.; Kurhanewicz, J.; et al. Detection of Bacteria-Specific Metabolism Using Hyperpolarized [2-13C]Pyruvate. ACS Infect. Dis. 2018, 4, 797–805. [Google Scholar] [CrossRef]
- Linares, D.M.; Kok, J.; Poolman, B. Genome Sequences of Lactococcus lactis MG1363 (Revised) and NZ9000 and Comparative Physiological Studies. J. Bacteriol. 2010, 192, 5806–5812. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Bachmann, H.; van Pelt-Klein Jan, E.; Douwenga, S.; Smid, E.J.; Teusink, B.; van Mastrigt, O. Lifestyle, Metabolism and Environmental Adaptation in Lactococcus lactis. FEMS Microbiol. Rev. 2020, 44, 804–820. [Google Scholar] [CrossRef]
- Voit, E.O.; Almeida, J.; Marino, S.; Lall, R.; Goel, G.; Neves, A.R.; Santos, H. Regulation of Glycolysis in Lactococcus lactis: An Unfinished Systems Biological Case Study. IEE Proc. Syst. Biol. 2006, 153, 286. [Google Scholar] [CrossRef]
- Neves, A.R.; Ramos, A.; Nunes, M.C.; Kleerebezem, M.; Hugenholtz, J.; De Vos, W.M.; Almeida, J.; Santos, H. In Vivo Nuclear Magnetic Resonance Studies of Glycolytic Kinetics in Lactococcus lactis. Biotechnol. Bioeng. 1999, 64, 200–212. [Google Scholar] [CrossRef]
- Ramos, A.; Neves, A.R.; Ventura, R.; Maycock, C.; López, P.; Santos, H. Effect of Pyruvate Kinase Overproduction on Glucose Metabolism of Lactococcus lactis. Microbiology 2004, 150, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Nielsen, J.; Förster, J. Modeling Lactococcus lactis Using a Genome-Scale Flux Model. BMC Microbiol. 2005, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Lu, M.; Choi, W.J.; Park, C.; Oh, H.B.; Lee, S.Y.; Lee, J. Dynamic Modeling of Lactic Acid Fermentation Metabolism with Lactococcus lactis. J. Microbiol. Biotechnol. 2011, 21, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Teusink, B.; Walsh, M.C.; Van Dam, K.; Westerhoff, H.V. The Danger of Metabolic Pathways with Turbo Design. Trends Biochem. Sci. 1998, 23, 162–169. [Google Scholar] [CrossRef]
- Jensen, P.R.; Matos, M.R.A.; Sonnenschein, N.; Meier, S. Combined In-Cell NMR and Simulation Approach to Probe Redox-Dependent Pathway Control. Anal. Chem. 2019, 91, 5395–5402. [Google Scholar] [CrossRef]
- Andersen, H.W.; Solem, C.; Hammer, K.; Jensen, P.R. Twofold Reduction of Phosphofructokinase Activity in Lactococcus lactis Results in Strong Decreases in Growth Rate and in Glycolytic Flux. J. Bacteriol. 2001, 183, 3458–3467. [Google Scholar] [CrossRef]
- Jeong, H.; Tombor, B.; Albert, R.; Oltvai, Z.N. The Large-Scale Organization of Metabolic Networks. Nature 2000, 407, 651–654. [Google Scholar] [CrossRef]
- Ma, H.; Zeng, A.-P. Reconstruction of Metabolic Networks from Genome Data and Analysis of Their Global Structure for Various Organisms. Bioinformatics 2003, 19, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Ramos, A.; Costa, H.; Van Swam, I.I.; Hugenholtz, J.; Kleerebezem, M.; De Vos, W.; Santos, H. Effect of Different NADH Oxidase Levels on Glucose Metabolism by Lactococcus lactis: Kinetics of Intracellular Metabolite Pools Determined by In Vivo Nuclear Magnetic Resonance. Appl. Environ. Microbiol. 2002, 68, 6332–6342. [Google Scholar] [CrossRef]
- Papagianni, M.; Avramidis, N.; Filiousis, G. Glycolysis and the Regulation of Glucose Transport in Lactococcus lactis spp. Lactis in Batch and Fed-Batch Culture. Microb. Cell. Fact. 2007, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Orlenko, A.; Hermansen, R.A.; Liberles, D.A. Flux Control in Glycolysis Varies Across the Tree of Life. J. Mol. Evol. 2016, 82, 146–161. [Google Scholar] [CrossRef]
- Liu, J.; Chan, S.H.J.; Chen, J.; Solem, C.; Jensen, P.R. Systems Biology—A Guide for Understanding and Developing Improved Strains of Lactic Acid Bacteria. Front. Microbiol. 2019, 10, 876. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619. [Google Scholar] [CrossRef]
- Liu, N.; Santala, S.; Stephanopoulos, G. Mixed Carbon Substrates: A Necessary Nuisance or a Missed Opportunity? Curr. Opin. Biotechnol. 2020, 62, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.M. Differential Effects of Iodoacetamide and Iodoacetate on Glycolysis and Glutathione Metabolism of Cultured Astrocytes. Front. Neuroenerg. 2009, 1, 463. [Google Scholar] [CrossRef]
- Day, S.E.; Kettunen, M.I.; Gallagher, F.A.; Hu, D.-E.; Lerche, M.; Wolber, J.; Golman, K.; Ardenkjaer-Larsen, J.H.; Brindle, K.M. Detecting Tumor Response to Treatment Using Hyperpolarized 13C Magnetic Resonance Imaging and Spectroscopy. Nat. Med. 2007, 13, 1382–1387. [Google Scholar] [CrossRef]
- Lodi, A.; Woods, S.M.; Ronen, S.M. Treatment with the MEK Inhibitor U0126 Induces Decreased Hyperpolarized Pyruvate to Lactate Conversion in Breast, but Not Prostate, Cancer Cells: U0126-induced Changes in Hyperpolarized Lactate. NMR Biomed. 2013, 26, 299–306. [Google Scholar] [CrossRef]
- Benthin, S.; Nielsen, J.; Villadsen, J. Two Uptake Systems for Fructose in Lactococcus lactis subsp. Cremoris FD1 Produce Glycolytic and Gluconeogenic Fructose Phosphates and Induce Oscillations in Growth and Lactic Acid Formation. Appl. Environ. Microbiol. 1993, 59, 3206–3211. [Google Scholar] [CrossRef]
- Dorau, R.; Chen, L.; Liu, J.; Jensen, P.R.; Solem, C. Efficient Production of α-Acetolactate by Whole Cell Catalytic Transformation of Fermentation-Derived Pyruvate. Microb. Cell Fact. 2019, 18, 217. [Google Scholar] [CrossRef]
- Xu, P.; Qiu, J.; Gao, C.; Ma, C. Biotechnological Routes to Pyruvate Production. J. Biosci. Bioeng. 2008, 105, 169–175. [Google Scholar] [CrossRef]
- Ramos, A.; Jordan, K.N.; Cogan, T.M.; Santos, H. 13C Nuclear Magnetic Resonance Studies of Citrate and Glucose Cometabolism by Lactococcus lactis. Appl Environ. Microbiol. 1994, 60, 1739–1748. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.-J.; Kim, S.; Hahn, J.-S. Efficient Production of Acetoin in Saccharomyces Cerevisiae by Disruption of 2,3-Butanediol Dehydrogenase and Expression of NADH Oxidase. Sci. Rep. 2016, 6, 27667. [Google Scholar] [CrossRef] [PubMed]
- Grütering, C.; Harhues, T.; Speen, F.; Keller, R.; Zimmermann, M.; Jensen, P.R.; Wessling, M.; Blank, L.M. Acetoin Production by Resting Cells of Lactococcus lactis for Direct Electrochemical Synthesis of 2-Butanone. Green Chem. 2023, 25, 9218–9225. [Google Scholar] [CrossRef]
- Kaneko, T.; Takahashi, M.; Suzuki, H. Acetoin Fermentation by Citrate-Positive Lactococcus lactis subsp. Lactis 3022 Grown Aerobically in the Presence of Hemin or Cu2+. Appl. Environ. Microbiol. 1990, 56, 2644–2649. [Google Scholar] [CrossRef] [PubMed]
- Garrigues, C.; Loubiere, P.; Lindley, N.D.; Cocaign-Bousquet, M. Control of the Shift from Homolactic Acid to Mixed-Acid Fermentation in Lactococcus lactis: Predominant Role of the NADH/NAD+ Ratio. J. Bacteriol. 1997, 179, 5282–5287. [Google Scholar] [CrossRef]
- Cocaign-Bousquet, M.; Garrigues, C.; Loubiere, P.; Lindley, N.D. Physiology of Pyruvate Metabolism in Lactococcus lactis. Antonie Van Leeuwenhoek 1996, 70, 253–267. [Google Scholar] [CrossRef]
- Lee, J.H.; Okuno, Y.; Cavagnero, S. Sensitivity Enhancement in Solution NMR: Emerging Ideas and New Frontiers. J. Magn. Reson. 2014, 241, 18–31. [Google Scholar] [CrossRef]
- Elliott, S.J.; Stern, Q.; Ceillier, M.; El Daraï, T.; Cousin, S.F.; Cala, O.; Jannin, S. Practical Dissolution Dynamic Nuclear Polarization. Prog. Nucl. Magn. Reson. Spectrosc. 2021, 126–127, 59–100. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meier, S.; Zahid, A.L.N.; Jørgensen, L.R.; Wang, K.-C.; Jensen, P.R.; Jensen, P.R. Hyperpolarized 13C NMR Reveals Pathway Regulation in Lactococcus lactis and Metabolic Similarities and Differences Across the Tree of Life. Molecules 2024, 29, 4133. https://doi.org/10.3390/molecules29174133
Meier S, Zahid ALN, Jørgensen LR, Wang K-C, Jensen PR, Jensen PR. Hyperpolarized 13C NMR Reveals Pathway Regulation in Lactococcus lactis and Metabolic Similarities and Differences Across the Tree of Life. Molecules. 2024; 29(17):4133. https://doi.org/10.3390/molecules29174133
Chicago/Turabian StyleMeier, Sebastian, Alexandra L. N. Zahid, Lucas Rebien Jørgensen, Ke-Chuan Wang, Peter Ruhdal Jensen, and Pernille Rose Jensen. 2024. "Hyperpolarized 13C NMR Reveals Pathway Regulation in Lactococcus lactis and Metabolic Similarities and Differences Across the Tree of Life" Molecules 29, no. 17: 4133. https://doi.org/10.3390/molecules29174133