


Targeting CBP and p300: Emerging Anticancer Agents



Abstract
1. Introduction
2. Wnt/β-Catenin Signaling Pathway
3. CBP and p300
3.1. CBP/β-Catenin Inhibitors
3.1.1. Non-Specific Antagonists of CBP/β-Catenin Signaling
3.1.2. Specific Antagonists of CBP/β-Catenin Signaling
3.2. p300/β-Catenin Inhibitors
3.2.1. Specific Antagonists of p300/β-Catenin Signaling
3.2.2. Non-Specific Antagonists of p300/β-Catenin Signaling
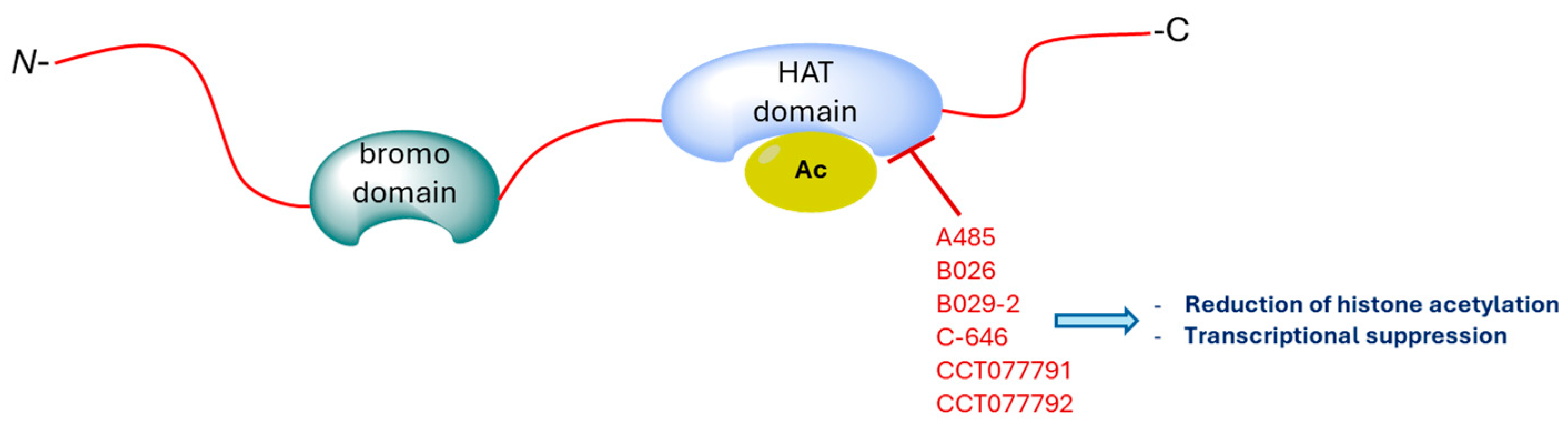
4. CBP/p300 Bromodomain Inhibitors
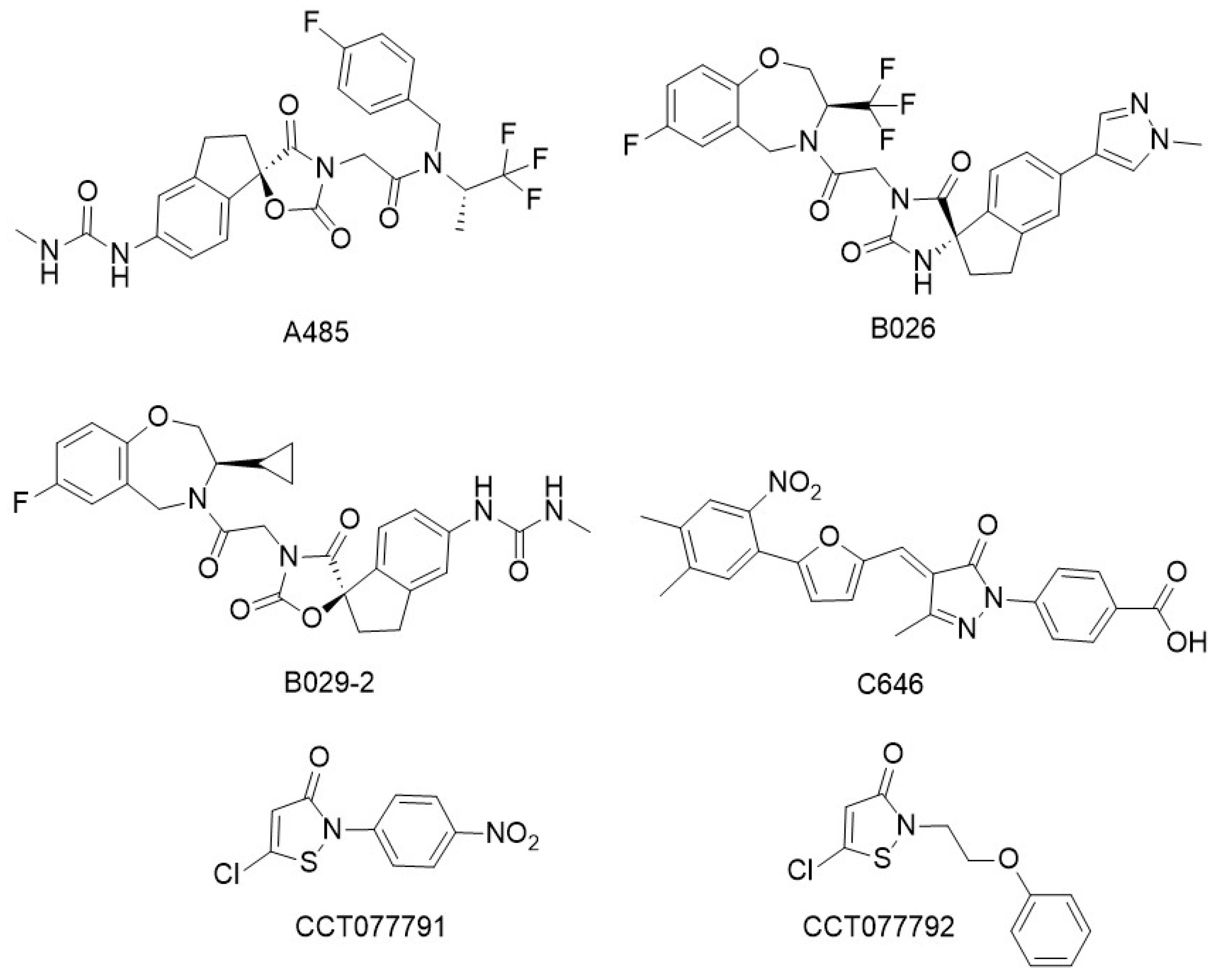
5. CBP/p300 HAT Inhibitors
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| 25(OH)D | 25-hydroxyvitamin D |
| β-TrCP | β-transducing repeats-containing proteins |
| APC | adenomatous polyposis coli |
| APL | acute promyelocytic leukemia |
| ATO | arsenic trioxide |
| ATRA | all-trans retinoic acid |
| BCL9 | B-cell lymphoma 9 |
| BCL9L | B-cell lymphoma 9-like |
| [BRD4(1)] | bromodomain-containing protein 4 |
| BRDT | bromodomain testis-specific protein |
| CBP | cyclic AMP response element-binding binding protein |
| CK1 | casein kinase 1 |
| CML | chronic myelogenous leukemia |
| CRC | colorectal cancer |
| CREB | cyclic AMP response element-binding |
| DLBCL | diffuse large B-cell lymphoma |
| DVL | disheveled |
| DVL-1 | disheveled 1 |
| DYRKs | dual-specificity tyrosine phosphorylation-regulated kinases |
| EBV | Epstein–Barr virus |
| Fzd | frizzled |
| GSK-3β | glycogen synthase kinase-3β |
| H3K27 | histone H3 lysine 27 |
| H3K27Ac | histone H3 lysine 27 acetylation |
| HAT | histone acetyltransferase |
| LEF | lymphoid enhancer-binding factor |
| LRP | low-density lipoprotein receptor-related proteins |
| LRP5 | low-density lipoprotein receptor-related protein 5 |
| LRP6 | low-density lipoprotein receptor-related protein 6 |
| MMPs | matrix metalloproteinases |
| NMC | NUT midline carcinoma |
| NPC | nasopharyngeal carcinoma |
| NUT | nuclear protein of the testis |
| RAREs | retinoic acid response elements |
| RARα | retinoic acid receptor alpha |
| RXR | retinoid X receptors |
| SSC | somatic stem cells |
| Tcf | T-cell factor |
References
- Grossman, S.R. P300/CBP/P53 Interaction and Regulation of the P53 Response. Eur. J. Biochem. 2001, 268, 2773–2778. [Google Scholar] [CrossRef] [PubMed]
- Nagasaka, M.; Miyajima, C.; Aoki, H.; Aoyama, M.; Morishita, D.; Inoue, Y.; Hayashi, H. Insights into Regulators of P53 Acetylation. Cells 2022, 11, 3825. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Bastian, I.N.; Long, M.D.; Dow, M.; Li, W.; Liu, T.; Ngu, R.K.; Antonucci, L.; Huang, J.Y.; Phung, Q.T.; et al. Activation of NF-ΚB and P300/CBP Potentiates Cancer Chemoimmunotherapy through Induction of MHC-I Antigen Presentation. Proc. Natl. Acad. Sci. USA 2021, 118, e2025840118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiong, F.; Wu, G.; Wang, D.; Liu, W.; Chen, J.; Qi, Y.; Wang, B.; Chen, Y. SMAD Proteins in TGF-β Signalling Pathway in Cancer: Regulatory Mechanisms and Clinical Applications. Diagnostics 2023, 13, 2769. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Tiu, B.; Sakamoto, K.M. CBP/P300 Acetyltransferase Activity in Hematologic Malignancies. Mol. Genet. Metab. 2016, 119, 37–43. [Google Scholar] [CrossRef]
- Albadari, N.; Deng, S.; Li, W. The Transcriptional Factors HIF-1 and HIF-2 and Their Novel Inhibitors in Cancer Therapy. Expert. Opin. Drug Discov. 2019, 14, 667–682. [Google Scholar] [CrossRef]
- Sun, Y.; Kolligs, F.T.; Hottiger, M.O.; Mosavin, R.; Fearon, E.R.; Nabel, G.J. Regulation of β-Catenin Transformation by the P300 Transcriptional Coactivator. Proc. Natl. Acad. Sci. USA 2000, 97, 12613–12618. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Choi, B.-R.; Cave, C.; Na, C.H.; Sockanathan, S. GDE2-Dependent Activation of Canonical Wnt Signaling in Neurons Regulates Oligodendrocyte Maturation. Cell Rep. 2020, 31, 107540. [Google Scholar] [CrossRef]
- Salik, B.; Yi, H.; Hassan, N.; Santiappillai, N.; Vick, B.; Connerty, P.; Duly, A.; Trahair, T.; Woo, A.J.; Beck, D.; et al. Targeting RSPO3-LGR4 Signaling for Leukemia Stem Cell Eradication in Acute Myeloid Leukemia. Cancer Cell 2020, 38, 263–278.e6. [Google Scholar] [CrossRef]
- Soleas, J.P.; D’Arcangelo, E.; Huang, L.; Karoubi, G.; Nostro, M.C.; McGuigan, A.P.; Waddell, T.K. Assembly of Lung Progenitors into Developmentally-Inspired Geometry Drives Differentiation via Cellular Tension. Biomaterials 2020, 254, 120128. [Google Scholar] [CrossRef] [PubMed]
- Valenta, T.; Hausmann, G.; Basler, K. The Many Faces and Functions of β-Catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Coluccia, A.; Bufano, M.; La Regina, G.; Puxeddu, M.; Toto, A.; Paone, A.; Bouzidi, A.; Musto, G.; Badolati, N.; Orlando, V.; et al. Anticancer Activity of (S)-5-Chloro-3-((3,5-Dimethylphenyl)Sulfonyl)-N-(1-Oxo-1-((Pyridin-4-Ylmethyl)Amino)Propan-2-Yl)-1H-Indole-2-Carboxamide (RS4690), a New Dishevelled 1 Inhibitor. Cancers 2022, 14, 1358. [Google Scholar] [CrossRef]
- Rubinfeld, B.; Albert, I.; Porfiri, E.; Fiol, C.; Munemitsu, S.; Polakis, P. Binding of GSK3β to the APC-β-Catenin Complex and Regulation of Complex Assembly. Science 1996, 272, 1023–1026. [Google Scholar] [CrossRef]
- Brack, A.S.; Murphy-Seiler, F.; Hanifi, J.; Deka, J.; Eyckerman, S.; Keller, C.; Aguet, M.; Rando, T.A. BCL9 Is an Essential Component of Canonical Wnt Signaling That Mediates the Differentiation of Myogenic Progenitors during Muscle Regeneration. Dev. Biol. 2009, 335, 93–105. [Google Scholar] [CrossRef]
- Cantù, C.; Zimmerli, D.; Hausmann, G.; Valenta, T.; Moor, A.; Aguet, M.; Basler, K. Pax6-Dependent, but β-Catenin-Independent, Function of Bcl9 Proteins in Mouse Lens Development. Genes Dev. 2014, 28, 1879–1884. [Google Scholar] [CrossRef]
- Kotolloshi, R.; Gajda, M.; Grimm, M.-O.; Steinbach, D. Wnt/β-Catenin Signalling and Its Cofactor BCL9L Have an Oncogenic Effect in Bladder Cancer Cells. Int. J. Mol. Sci. 2022, 23, 5319. [Google Scholar] [CrossRef]
- Hecht, A. The P300/CBP Acetyltransferases Function as Transcriptional Coactivators of Beta-Catenin in Vertebrates. EMBO J. 2000, 19, 1839–1850. [Google Scholar] [CrossRef]
- Schwab, K.R.; Patterson, L.T.; Hartman, H.A.; Song, N.; Lang, R.A.; Lin, X.; Potter, S.S. Pygo1 and Pygo2 Roles in Wnt Signaling in Mammalian Kidney Development. BMC Biol. 2007, 5, 15. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt Signaling Pathway in Non-Small Cell Lung Cancer. JNCI J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.H.; Weis, W.I. The Structure of the β-Catenin/E-Cadherin Complex and the Molecular Basis of Diverse Ligand Recognition by β-Catenin. Cell 2001, 105, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.-Q.; You, A.-B.; Zhu, X.-D.; Zhang, W.; Zhang, Y.-Y.; Zhang, S.-Z.; Zhang, K.; Cai, H.; Shi, W.-K.; Li, X.-L.; et al. MiR-182-5p Promotes Hepatocellular Carcinoma Progression by Repressing FOXO3a. J. Hematol. Oncol. 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Weng, W.; Zhang, Q.; Wu, Y.; Ni, S.; Tan, C.; Xu, M.; Sun, H.; Liu, C.; Wei, P.; et al. The LncRNA NEAT1 Activates Wnt/β-Catenin Signaling and Promotes Colorectal Cancer Progression via Interacting with DDX5. J. Hematol. Oncol. 2018, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Spaan, I.; Raymakers, R.A.; van de Stolpe, A.; Peperzak, V. Wnt Signaling in Multiple Myeloma: A Central Player in Disease with Therapeutic Potential. J. Hematol. Oncol. 2018, 11, 67. [Google Scholar] [CrossRef]
- Nalli, M.; Masci, D.; Urbani, A.; La Regina, G.; Silvestri, R. Emerging Direct Targeting β-Catenin Agents. Molecules 2022, 27, 7735. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-Catenin Signaling Pathway in Cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Teo, J.-L.; Kahn, M. The Wnt Signaling Pathway in Cellular Proliferation and Differentiation: A Tale of Two Coactivators. Adv. Drug Deliv. Rev. 2010, 62, 1149–1155. [Google Scholar] [CrossRef]
- Giordano, A.; Avantaggiati, M.L. P300 and CBP: Partners for Life and Death. J. Cell Physiol. 1999, 181, 218–230. [Google Scholar] [CrossRef]
- Goodman, R.H.; Smolik, S. CBP/P300 in Cell Growth, Transformation, and Development. Genes Dev. 2000, 14, 1553–1577. [Google Scholar] [CrossRef] [PubMed]
- Eckner, R.; Ewen, M.E.; Newsome, D.; Gerdes, M.; DeCaprio, J.A.; Lawrence, J.B.; Livingston, D.M. Molecular Cloning and Functional Analysis of the Adenovirus E1A-Associated 300-KD Protein (P300) Reveals a Protein with Properties of a Transcriptional Adaptor. Genes Dev. 1994, 8, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiati, M.L.; Ogryzko, V.; Gardner, K.; Giordano, A.; Levine, A.S.; Kelly, K. Recruitment of P300/CBP in P53-Dependent Signal Pathways. Cell 1997, 89, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Marzio, G.; Wagener, C.; Gutierrez, M.I.; Cartwright, P.; Helin, K.; Giacca, M. E2F Family Members Are Differentially Regulated by Reversible Acetylation. J. Biol. Chem. 2000, 275, 10887–10892. [Google Scholar] [CrossRef]
- Chan, H.M.; La Thangue, N.B. P300/CBP Proteins: HATs for Transcriptional Bridges and Scaffolds. J. Cell Sci. 2001, 114, 2363–2373. [Google Scholar] [CrossRef]
- Iyer, N.G.; Özdag, H.; Caldas, C. P300/CBP and Cancer. Oncogene 2004, 23, 4225–4231. [Google Scholar] [CrossRef]
- Durbin, A.D.; Wang, T.; Wimalasena, V.K.; Zimmerman, M.W.; Li, D.; Dharia, N.V.; Mariani, L.; Shendy, N.A.M.; Nance, S.; Patel, A.G.; et al. EP300 Selectively Controls the Enhancer Landscape of MYCN -Amplified Neuroblastoma. Cancer Discov. 2022, 12, 730–751. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, B.; Liu, X.; Zhang, X.D.; Zhang, L.; Liu, T. Histone Acetyltransferases CBP/P300 in Tumorigenesis and CBP/P300 Inhibitors as Promising Novel Anticancer Agents. Theranostics 2022, 12, 4935–4948. [Google Scholar] [CrossRef]
- Raisner, R.; Kharbanda, S.; Jin, L.; Jeng, E.; Chan, E.; Merchant, M.; Haverty, P.M.; Bainer, R.; Cheung, T.; Arnott, D.; et al. Enhancer Activity Requires CBP/P300 Bromodomain-Dependent Histone H3K27 Acetylation. Cell Rep. 2018, 24, 1722–1729. [Google Scholar] [CrossRef]
- Strachowska, M.; Robaszkiewicz, A. Characteristics of Anticancer Activity of CBP/P300 Inhibitors—Features of Their Classes, Intracellular Targets and Future Perspectives of Their Application in Cancer Treatment. Pharmacol. Ther. 2024, 257, 108636. [Google Scholar] [CrossRef]
- Shi, Y.; Mello, C. A CBP/P300 Homolog Specifies Multiple Differentiation Pathways in Caenorhabditis Elegans. Genes Dev. 1998, 12, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Dyson, H.J.; Wright, P.E. Intrinsically Unstructured Proteins and Their Functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Kim, Y.-M.; Kahn, M. Wnt/Catenin Signaling in Adult Stem Cell Physiology and Disease. Stem Cell Rev. Rep. 2014, 10, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Masiello, D.; McMillian, M.; Nguyen, C.; Wu, Y.; Melendez, E.; Smbatyan, G.; Kida, A.; He, Y.; Teo, J.-L.; et al. CBP/Catenin Antagonist Safely Eliminates Drug-Resistant Leukemia-Initiating Cells. Oncogene 2016, 35, 3705–3717. [Google Scholar] [CrossRef] [PubMed]
- Manegold, P.; Lai, K.; Wu, Y.; Teo, J.-L.; Lenz, H.-J.; Genyk, Y.; Pandol, S.; Wu, K.; Lin, D.; Chen, Y.; et al. Differentiation Therapy Targeting the β-Catenin/CBP Interaction in Pancreatic Cancer. Cancers 2018, 10, 95. [Google Scholar] [CrossRef]
- Gang, E.J.; Hsieh, Y.-T.; Pham, J.; Zhao, Y.; Nguyen, C.; Huantes, S.; Park, E.; Naing, K.; Klemm, L.; Swaminathan, S.; et al. Small-Molecule Inhibition of CBP/Catenin Interactions Eliminates Drug-Resistant Clones in Acute Lymphoblastic Leukemia. Oncogene 2014, 33, 2169–2178. [Google Scholar] [CrossRef]
- Thomas, P.D.; Kahn, M. Kat3 Coactivators in Somatic Stem Cells and Cancer Stem Cells: Biological Roles, Evolution, and Pharmacologic Manipulation. Cell Biol. Toxicol. 2016, 32, 61–81. [Google Scholar] [CrossRef]
- Sasaki, T.; Hwang, H.; Nguyen, C.; Kloner, R.A.; Kahn, M. The Small Molecule Wnt Signaling Modulator ICG-001 Improves Contractile Function in Chronically Infarcted Rat Myocardium. PLoS ONE 2013, 8, e75010. [Google Scholar] [CrossRef]
- Henderson, W.R.; Chi, E.Y.; Ye, X.; Nguyen, C.; Tien, Y.; Zhou, B.; Borok, Z.; Knight, D.A.; Kahn, M. Inhibition of Wnt/β-Catenin/CREB Binding Protein (CBP) Signaling Reverses Pulmonary Fibrosis. Proc. Natl. Acad. Sci. USA 2010, 107, 14309–14314. [Google Scholar] [CrossRef]
- Teo, J.-L.; Ma, H.; Nguyen, C.; Lam, C.; Kahn, M. Specific Inhibition of CBP/β-Catenin Interaction Rescues Defects in Neuronal Differentiation Caused by a Presenilin-1 Mutation. Proc. Natl. Acad. Sci. USA 2005, 102, 12171–12176. [Google Scholar] [CrossRef]
- Kahn, M. Taking the Road Less Traveled—The Therapeutic Potential of CBP/β-Catenin Antagonists. Expert. Opin. Ther. Targets 2021, 25, 701–719. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, J.J.; Chale, R.S.; Abad, A.C.; Schally, A.V. Acute Promyelocytic Leukemia (APL): A Review of the Literature. Oncotarget 2020, 11, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Breitman, T.R.; Selonick, S.E.; Collins, S.J. Induction of Differentiation of the Human Promyelocytic Leukemia Cell Line (HL-60) by Retinoic Acid. Proc. Natl. Acad. Sci. USA 1980, 77, 2936–2940. [Google Scholar] [CrossRef] [PubMed]
- Chambon, P. The Nuclear Receptor Superfamily: A Personal Retrospect on the First Two Decades. Mol. Endocrinol. 2005, 19, 1418–1428. [Google Scholar] [CrossRef]
- Kishimoto, M.; Fujiki, R.; Takezawa, S.; Sasaki, Y.; Nakamura, T.; Yamaoka, K.; Kitagawa, H.; Kato, S. Nuclear Receptor Mediated Gene Regulation through Chromatin Remodeling and Histone Modifications. Endocr. J. 2006, 53, 157–172. [Google Scholar] [CrossRef]
- Dillard, A.C.; Lane, M.A. Retinol Decreases Β-catenin Protein Levels in Retinoic Acid-resistant Colon Cancer Cell Lines. Mol. Carcinog. 2007, 46, 315–329. [Google Scholar] [CrossRef]
- Huynh, T.T.; Sultan, M.; Vidovic, D.; Dean, C.A.; Cruickshank, B.M.; Lee, K.; Loung, C.-Y.; Holloway, R.W.; Hoskin, D.W.; Waisman, D.M.; et al. Retinoic Acid and Arsenic Trioxide Induce Lasting Differentiation and Demethylation of Target Genes in APL Cells. Sci. Rep. 2019, 9, 9414. [Google Scholar] [CrossRef]
- Mondul, A.M.; Weinstein, S.J.; Layne, T.M.; Albanes, D. Vitamin D and Cancer Risk and Mortality: State of the Science, Gaps, and Challenges. Epidemiol. Rev. 2017, 39, 28–48. [Google Scholar] [CrossRef]
- Giovannucci, E. The Epidemiology of Vitamin D and Cancer Incidence and Mortality: A Review (United States). Cancer Causes Control 2005, 16, 83–95. [Google Scholar] [CrossRef]
- Bettoun, D.J.; Burris, T.P.; Houck, K.A.; Buck, D.W.; Stayrook, K.R.; Khalifa, B.; Lu, J.; Chin, W.W.; Nagpal, S. Retinoid X Receptor Is a Nonsilent Major Contributor to Vitamin D Receptor-Mediated Transcriptional Activation. Mol. Endocrinol. 2003, 17, 2320–2328. [Google Scholar] [CrossRef]
- Kahn, M. Can We Safely Target the WNT Pathway? Nat. Rev. Drug Discov. 2014, 13, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Omary, M.B.; Lugea, A.; Lowe, A.W.; Pandol, S.J. The Pancreatic Stellate Cell: A Star on the Rise in Pancreatic Diseases. J. Clin. Investig. 2007, 117, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Che, M.; Kweon, S.-M.; Teo, J.-L.; Yuan, Y.-C.; Melstrom, L.G.; Waldron, R.T.; Lugea, A.; Urrutia, R.A.; Pandol, S.J.; Lai, K.K.Y. Targeting the CBP/β-Catenin Interaction to Suppress Activation of Cancer-Promoting Pancreatic Stellate Cells. Cancers 2020, 12, 1476. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, M.; Kestler, H.A.; Holzmann, K.; Ellenrieder, V.; Schneiderhan, W.; Siech, M.; Adler, G.; Bachem, M.G.; Gress, T.M. Transcriptome Analysis of Human Hepatic and Pancreatic Stellate Cells: Organ-Specific Variations of a Common Transcriptional Phenotype. J. Mol. Med. 2005, 83, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chiang, Y.C.; Chan, L.S.; Chau, W.Y.; Lung, M.L.; Kahn, M.; Lo, K.W.; Mak, N.K.; Lung, H.L. The CBP/β-Catenin Antagonist, ICG-001, Inhibits Tumor Metastasis via Blocking of the MiR-134/ITGB1 Axis-Mediated Cell Adhesion in Nasopharyngeal Carcinoma. Cancers 2022, 14, 3125. [Google Scholar] [CrossRef] [PubMed]
- Danieau, G.; Morice, S.; Renault, S.; Brion, R.; Biteau, K.; Amiaud, J.; Cadé, M.; Heymann, D.; Lézot, F.; Verrecchia, F.; et al. ICG-001, an Inhibitor of the β-Catenin and CAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells. Pharmaceuticals 2021, 14, 421. [Google Scholar] [CrossRef]
- Choi, J.-H.; Jang, T.-Y.; Jeon, S.-E.; Kim, J.-H.; Lee, C.-J.; Yun, H.-J.; Jung, J.-Y.; Park, S.-Y.; Nam, J.-S. The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression. Int. J. Mol. Sci. 2021, 22, 13413. [Google Scholar] [CrossRef]
- Okazaki, H.; Sato, S.; Koyama, K.; Morizumi, S.; Abe, S.; Azuma, M.; Chen, Y.; Goto, H.; Aono, Y.; Ogawa, H.; et al. The Novel Inhibitor PRI-724 for Wnt/β-Catenin/CBP Signaling Ameliorates Bleomycin-Induced Pulmonary Fibrosis in Mice. Exp. Lung Res. 2019, 45, 188–199. [Google Scholar] [CrossRef]
- Ding, H.; Chen, J.; Qin, J.; Chen, R.; Yi, Z. TGF-β-Induced α-SMA Expression Is Mediated by C/EBPβ Acetylation in Human Alveolar Epithelial Cells. Mol. Med. 2021, 27, 22. [Google Scholar] [CrossRef]
- Gabata, R.; Harada, K.; Mizutani, Y.; Ouchi, H.; Yoshimura, K.; Sato, Y.; Kitao, A.; Kimura, K.; Kouji, H.; Miyashita, T.; et al. Anti-Tumor Activity of the Small Molecule Inhibitor PRI-724 Against β-Catenin-Activated Hepatocellular Carcinoma. Anticancer Res. 2020, 40, 5211–5219. [Google Scholar] [CrossRef]
- Yamada, K.; Hori, Y.; Inoue, S.; Yamamoto, Y.; Iso, K.; Kamiyama, H.; Yamaguchi, A.; Kimura, T.; Uesugi, M.; Ito, J.; et al. E7386, a Selective Inhibitor of the Interaction between β-Catenin and CBP, Exerts Antitumor Activity in Tumor Models with Activated Canonical Wnt Signaling. Cancer Res. 2021, 81, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.; Yamada, K.; Kato, Y.; Ozawa, Y.; Odagami, T.; Matsui, J.; Matsushima, T.; Nomoto, K.; Kouji, H.; Owa, T. Abstract 5172: E7386, an Orally Active CBP/Beta-Catenin Modulator, Induces T Cells Infiltration into Tumor and Enhances Antitumor Activity of Anti-PD-1 MAb in Wnt1 Tumor Syngeneic Mice Model. Cancer Res. 2017, 77, 5172. [Google Scholar] [CrossRef]
- Kahn, M. Symmetric Division Versus Asymmetric Division: A Tale of Two Coactivators. Future Med. Chem. 2011, 3, 1745–1763. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Yasuda, S.; Teo, J.-L.; Nguyen, C.; McMillan, M.; Hsieh, C.-L.; Suemori, H.; Nakatsuji, N.; Yamamoto, M.; Miyabayashi, T.; et al. Wnt Signaling Orchestration with a Small Molecule DYRK Inhibitor Provides Long-Term Xeno-Free Human Pluripotent Cell Expansion. Stem Cells Transl. Med. 2012, 1, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Nguyen, C.; Yasuda, S.-Y.; McMillan, M.; Hasegawa, K.; Kahn, M. Specific Direct Small Molecule P300/β-Catenin Antagonists Maintain Stem Cell Potency. Curr. Mol. Pharmacol. 2016, 9, 272–279. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, K.; Nguyen, C.; Smbatyan, G.; Melendez, E.; Higuchi, Y.; Chen, Y.; Kahn, M. Small Molecule P300/Catenin Antagonist Enhances Hematopoietic Recovery after Radiation. PLoS ONE 2017, 12, e0177245. [Google Scholar] [CrossRef]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.-L.; Oh, S.W.; Kim, H.Y.; et al. A Small Molecule Inhibitor of β-Catenin/Cyclic AMP Response Element-Binding Protein Transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef]
- Miyabayashi, T.; Teo, J.-L.; Yamamoto, M.; McMillan, M.; Nguyen, C.; Kahn, M. Wnt/β-Catenin/CBP Signaling Maintains Long-Term Murine Embryonic Stem Cell Pluripotency. Proc. Natl. Acad. Sci. USA 2007, 104, 5668–5673. [Google Scholar] [CrossRef]
- Boni, J.; Rubio-Perez, C.; López-Bigas, N.; Fillat, C.; de la Luna, S. The DYRK Family of Kinases in Cancer: Molecular Functions and Therapeutic Opportunities. Cancers 2020, 12, 2106. [Google Scholar] [CrossRef]
- Xu, J.; Fang, S.; Wang, N.; Li, B.; Huang, Y.; Fan, Q.; Shi, J.; Liu, H.; Shao, Z. Dual-Specificity Tyrosine Phosphorylation-Regulated Kinase Inhibitor ID-8 Promotes Human Somatic Cell Reprogramming by Activating PDK4 Expression. Stem Cell Rev. Rep. 2022, 18, 2074–2087. [Google Scholar] [CrossRef]
- Monteiro, M.B.; Ramm, S.; Chandrasekaran, V.; Boswell, S.A.; Weber, E.J.; Lidberg, K.A.; Kelly, E.J.; Vaidya, V.S. A High-Throughput Screen Identifies DYRK1A Inhibitor ID-8 That Stimulates Human Kidney Tubular Epithelial Cell Proliferation. J. Am. Soc. Nephrol. 2018, 29, 2820–2833. [Google Scholar] [CrossRef]
- Mujtaba, S.; He, Y.; Zeng, L.; Yan, S.; Plotnikova, O.; Sachchidanand; Sanchez, R.; Zeleznik-Le, N.J.; Ronai, Z.; Zhou, M.-M. Structural Mechanism of the Bromodomain of the Coactivator CBP in P53 Transcriptional Activation. Mol. Cell 2004, 13, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Delvecchio, M.; Gaucher, J.; Aguilar-Gurrieri, C.; Ortega, E.; Panne, D. Structure of the P300 Catalytic Core and Implications for Chromatin Targeting and HAT Regulation. Nat. Struct. Mol. Biol. 2013, 20, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Yu, L.-R.; Wang, L.; Zhang, Z.; Kasper, L.H.; Lee, J.-E.; Wang, C.; Brindle, P.K.; Dent, S.Y.R.; Ge, K. Distinct Roles of GCN5/PCAF-Mediated H3K9ac and CBP/P300-Mediated H3K18/27ac in Nuclear Receptor Transactivation. EMBO J. 2011, 30, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Conery, A.R.; Centore, R.C.; Neiss, A.; Keller, P.J.; Joshi, S.; Spillane, K.L.; Sandy, P.; Hatton, C.; Pardo, E.; Zawadzke, L.; et al. Bromodomain Inhibition of the Transcriptional Coactivators CBP/EP300 as a Therapeutic Strategy to Target the IRF4 Network in Multiple Myeloma. Elife 2016, 5, 10483. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.A.; Magnuson, S.; Pastor, R.; Tsui, V.H.-W.; Murray, J.; Crawford; Albrecht, B.K.; Cote, A.; Taylor, A.M.; Lai, K.W.; et al. 4,5,6,7-Tetrahydro-1H-Pyrazolo [4,3-C]Pyridin-3-Amine Compounds as CBP and/or EP300 Inhibitors. WO-2016086200-A9, 23 June 2016. [Google Scholar]
- Jin, L.; Garcia, J.; Chan, E.; de la Cruz, C.; Segal, E.; Merchant, M.; Kharbanda, S.; Raisner, R.; Haverty, P.M.; Modrusan, Z.; et al. Therapeutic Targeting of the CBP/P300 Bromodomain Blocks the Growth of Castration-Resistant Prostate Cancer. Cancer Res. 2017, 77, 5564–5575. [Google Scholar] [CrossRef]
- Romero, F.A.; Murray, J.; Lai, K.W.; Tsui, V.; Albrecht, B.K.; An, L.; Beresini, M.H.; de Leon Boenig, G.; Bronner, S.M.; Chan, E.W.; et al. GNE-781, A Highly Advanced Potent and Selective Bromodomain Inhibitor of Cyclic Adenosine Monophosphate Response Element Binding Protein, Binding Protein (CBP). J. Med. Chem. 2017, 60, 9162–9183. [Google Scholar] [CrossRef]
- Welti, J.; Sharp, A.; Brooks, N.; Yuan, W.; McNair, C.; Chand, S.N.; Pal, A.; Figueiredo, I.; Riisnaes, R.; Gurel, B.; et al. Targeting the P300/CBP Axis in Lethal Prostate Cancer. Cancer Discov. 2021, 11, 1118–1137. [Google Scholar] [CrossRef]
- Penney, K.L.; Schumacher, F.R.; Kraft, P.; Mucci, L.A.; Sesso, H.D.; Ma, J.; Niu, Y.; Cheong, J.K.; Hunter, D.J.; Stampfer, M.J.; et al. Association of KLK3 (PSA) Genetic Variants with Prostate Cancer Risk and PSA Levels. Carcinogenesis 2011, 32, 853–859. [Google Scholar] [CrossRef]
- Diesch, J.; Le Pannérer, M.-M.; Winkler, R.; Casquero, R.; Muhar, M.; van der Garde, M.; Maher, M.; Herráez, C.M.; Bech-Serra, J.J.; Fellner, M.; et al. Inhibition of CBP Synergizes with the RNA-Dependent Mechanisms of Azacitidine by Limiting Protein Synthesis. Nat. Commun. 2021, 12, 6060. [Google Scholar] [CrossRef]
- Hammitzsch, A.; Tallant, C.; Fedorov, O.; O’Mahony, A.; Brennan, P.E.; Hay, D.A.; Martinez, F.O.; Al-Mossawi, M.H.; de Wit, J.; Vecellio, M.; et al. CBP30, a Selective CBP/P300 Bromodomain Inhibitor, Suppresses Human Th17 Responses. Proc. Natl. Acad. Sci. USA 2015, 112, 10768–10773. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.A.; Fedorov, O.; Martin, S.; Singleton, D.C.; Tallant, C.; Wells, C.; Picaud, S.; Philpott, M.; Monteiro, O.P.; Rogers, C.M.; et al. Discovery and Optimization of Small-Molecule Ligands for the CBP/P300 Bromodomains. J. Am. Chem. Soc. 2014, 136, 9308–9319. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carpizo, V.; Ruiz-Llorente, S.; Sarmentero, J.; Graña-Castro, O.; Pisano, D.G.; Barrero, M.J. CREBBP/EP300 Bromodomains Are Critical to Sustain the GATA1/MYC Regulatory Axis in Proliferation. Epigenetics Chromatin 2018, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Spriano, F.; Gaudio, E.; Cascione, L.; Tarantelli, C.; Melle, F.; Motta, G.; Priebe, V.; Rinaldi, A.; Golino, G.; Mensah, A.A.; et al. Antitumor Activity of the Dual BET and CBP/EP300 Inhibitor NEO2734. Blood Adv. 2020, 4, 4124–4135. [Google Scholar] [CrossRef]
- Canales, T.M.; van Gils, N.; Vermue, E.; Rutten, A.; Denkers, F.; van der Deure, T.; Giles, F.; Smit, L. PS967 Preclinical activity of the novel oral dual bet-CBP/P300 inhibitors, NEO1132 and NEO2734, in acute myeloid leukemia. Hemasphere 2019, 3, 435–436. [Google Scholar] [CrossRef]
- van Gils, N.; Martiañez Canales, T.; Vermue, E.; Rutten, A.; Denkers, F.; van der Deure, T.; Ossenkoppele, G.J.; Giles, F.; Smit, L. The Novel Oral BET-CBP/P300 Dual Inhibitor NEO2734 Is Highly Effective in Eradicating Acute Myeloid Leukemia Blasts and Stem/Progenitor Cells. Hemasphere 2021, 5, e610. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Qi, J.; Picaud, S.; Shen, Y.; Smith, W.B.; Fedorov, O.; Morse, E.M.; Keates, T.; Hickman, T.T.; Felletar, I.; et al. Selective Inhibition of BET Bromodomains. Nature 2010, 468, 1067–1073. [Google Scholar] [CrossRef]
- Jiang, G.; Deng, W.; Liu, Y.; Wang, C. General Mechanism of JQ1 in Inhibiting Various Types of Cancer. Mol. Med. Rep. 2020, 21, 1021–1034. [Google Scholar] [CrossRef]
- Picaud, S.; Fedorov, O.; Thanasopoulou, A.; Leonards, K.; Jones, K.; Meier, J.; Olzscha, H.; Monteiro, O.; Martin, S.; Philpott, M.; et al. Generation of a Selective Small Molecule Inhibitor of the CBP/P300 Bromodomain for Leukemia Therapy. Cancer Res. 2015, 75, 5106–5119. [Google Scholar] [CrossRef]
- Strachowska, M.; Gronkowska, K.; Michlewska, S.; Robaszkiewicz, A. CBP/P300 Bromodomain Inhibitor–I–CBP112 Declines Transcription of the Key ABC Transporters and Sensitizes Cancer Cells to Chemotherapy Drugs. Cancers 2021, 13, 4614. [Google Scholar] [CrossRef]
- Ogryzko, V.V.; Schiltz, R.L.; Russanova, V.; Howard, B.H.; Nakatani, Y. The Transcriptional Coactivators P300 and CBP Are Histone Acetyltransferases. Cell 1996, 87, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. The CBP Co-Activator Is a Histone Acetyltransferase. Nature 1996, 384, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Puttagunta, R.; Tedeschi, A.; Sória, M.G.; Hervera, A.; Lindner, R.; Rathore, K.I.; Gaub, P.; Joshi, Y.; Nguyen, T.; Schmandke, A.; et al. PCAF-Dependent Epigenetic Changes Promote Axonal Regeneration in the Central Nervous System. Nat. Commun. 2014, 5, 3527. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, C.; Yamaguchi, K.; Itoh, Y.; Nishimura, T.; Umemura, A.; Minami, M.; Yasui, K.; Mitsuyoshi, H.; Fujii, H.; Tochiki, N.; et al. High Expression of P300 in HCC Predicts Shortened Overall Survival in Association with Enhanced Epithelial Mesenchymal Transition of HCC Cells. Cancer Lett. 2011, 310, 140–147. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Sevinç, K.; Gürhan Sevinç, G.; Cribbs, A.P.; Philpott, M.; Uyulur, F.; Morova, T.; Dunford, J.E.; Göklemez, S.; Arı, Ş.; et al. Bromodomain Inhibition of the Coactivators CBP/EP300 Facilitate Cellular Reprogramming. Nat. Chem. Biol. 2019, 15, 519–528. [Google Scholar] [CrossRef]
- Petrif, F.; Giles, R.H.; Dauwerse, H.G.; Saris, J.J.; Hennekam, R.C.M.; Masuno, M.; Tommerup, N.; van Ommen, G.-J.B.; Goodman, R.H.; Peters, D.J.M.; et al. Rubinstein-Taybi Syndrome Caused by Mutations in the Transcriptional Co-Activator CBP. Nature 1995, 376, 348–351. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, F.; Asai, T.; Lai, F.; Man, N.; Xu, H.; Chen, S.; Greenblatt, S.; Hamard, P.-J.; Ando, K.; et al. Loss of P300 Accelerates MDS-Associated Leukemogenesis. Leukemia 2017, 31, 1382–1390. [Google Scholar] [CrossRef]
- Lasko, L.M.; Jakob, C.G.; Edalji, R.P.; Qiu, W.; Montgomery, D.; Digiammarino, E.L.; Hansen, T.M.; Risi, R.M.; Frey, R.; Manaves, V.; et al. Discovery of a Selective Catalytic P300/CBP Inhibitor That Targets Lineage-Specific Tumours. Nature 2017, 550, 128–132. [Google Scholar] [CrossRef]
- Sayapina, M.S.; Savelov, N.A.; Karseladze, A.I.; Bulanov, A.A.; Tryakin, A.A.; Nosov, D.A.; Garin, A.M.; Tjulandin, S.A. Nuclear Protein of the Testis Midline Carcinoma Masquerading as a Primary Mediastinal Seminoma. Rare Tumors 2016, 8, 85–87. [Google Scholar] [CrossRef]
- Zhang, X.; Zegar, T.; Lucas, A.; Morrison-Smith, C.; Knox, T.; French, C.A.; Knapp, S.; Müller, S.; Siveke, J.T. Therapeutic Targeting of P300/CBP HAT Domain for the Treatment of NUT Midline Carcinoma. Oncogene 2020, 39, 4770–4779. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, R.; Li, Z.; Mei, L.; Wan, S.; Ding, H.; Chen, Z.; Xing, J.; Feng, H.; Han, J.; et al. Discovery of Highly Potent, Selective, and Orally Efficacious P300/CBP Histone Acetyltransferases Inhibitors. J. Med. Chem. 2020, 63, 1337–1360. [Google Scholar] [CrossRef] [PubMed]
- Bowers, E.M.; Yan, G.; Mukherjee, C.; Orry, A.; Wang, L.; Holbert, M.A.; Crump, N.T.; Hazzalin, C.A.; Liszczak, G.; Yuan, H.; et al. Virtual Ligand Screening of the P300/CBP Histone Acetyltransferase: Identification of a Selective Small Molecule Inhibitor. Chem. Biol. 2010, 17, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.-Y.; Chen, S.-J.; Xiao, S.-H.; Sun, Q.-J.; Ding, C.-H.; Zheng, B.-N.; Zhu, X.-Y.; Liu, S.-Q.; Yang, F.; Yang, Y.-X.; et al. Targeting P300/CBP Attenuates Hepatocellular Carcinoma Progression through Epigenetic Regulation of Metabolism. Cancer Res. 2021, 81, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Sasaki, M.; Mitachi, T.; Oike, T.; Higuchi, S.; Tominaga, Y.; Kohno, T. Targeting P300 Addiction in CBP-Deficient Cancers Causes Synthetic Lethality by Apoptotic Cell Death Due to Abrogation of MYC Expression. Cancer Discov. 2016, 6, 430–445. [Google Scholar] [CrossRef]
- Giotopoulos, G.; Chan, W.-I.; Horton, S.J.; Ruau, D.; Gallipoli, P.; Fowler, A.; Crawley, C.; Papaemmanuil, E.; Campbell, P.J.; Göttgens, B.; et al. The Epigenetic Regulators CBP and P300 Facilitate Leukemogenesis and Represent Therapeutic Targets in Acute Myeloid Leukemia. Oncogene 2016, 35, 279–289. [Google Scholar] [CrossRef]
- Huang, X.; Yan, J.; Zhang, M.; Wang, Y.; Chen, Y.; Fu, X.; Wei, R.; Zheng, X.; Liu, Z.; Zhang, X.; et al. Targeting Epigenetic Crosstalk as a Therapeutic Strategy for EZH2-Aberrant Solid Tumors. Cell 2018, 175, 186–199.e19. [Google Scholar] [CrossRef]
- Roth, S.Y.; Denu, J.M.; Allis, C.D. Histone Acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef]
- Stimson, L.; Rowlands, M.G.; Newbatt, Y.M.; Smith, N.F.; Raynaud, F.I.; Rogers, P.; Bavetsias, V.; Gorsuch, S.; Jarman, M.; Bannister, A.; et al. Isothiazolones as Inhibitors of PCAF and P300 Histone Acetyltransferase Activity. Mol. Cancer Ther. 2005, 4, 1521–1532. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New Colorimetric Cytotoxicity Assay for Anticancer-Drug Screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Compounds | Ref. |
|---|---|---|
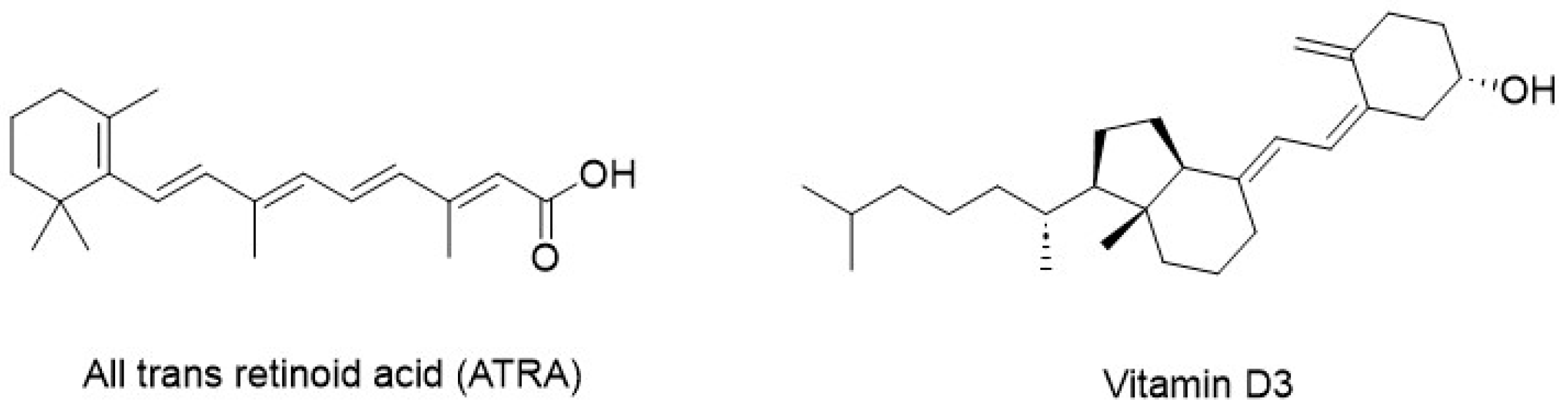
| Non-specific inhibitors of CBP/β-catenin signaling | ATRA, Vitamin D3 | [51,56] |
| Specific inhibitors of CBP/β-catenin signaling | ICG-001, PRI-724, C-82, E-7386 | [61,62,63,64,70,72] |
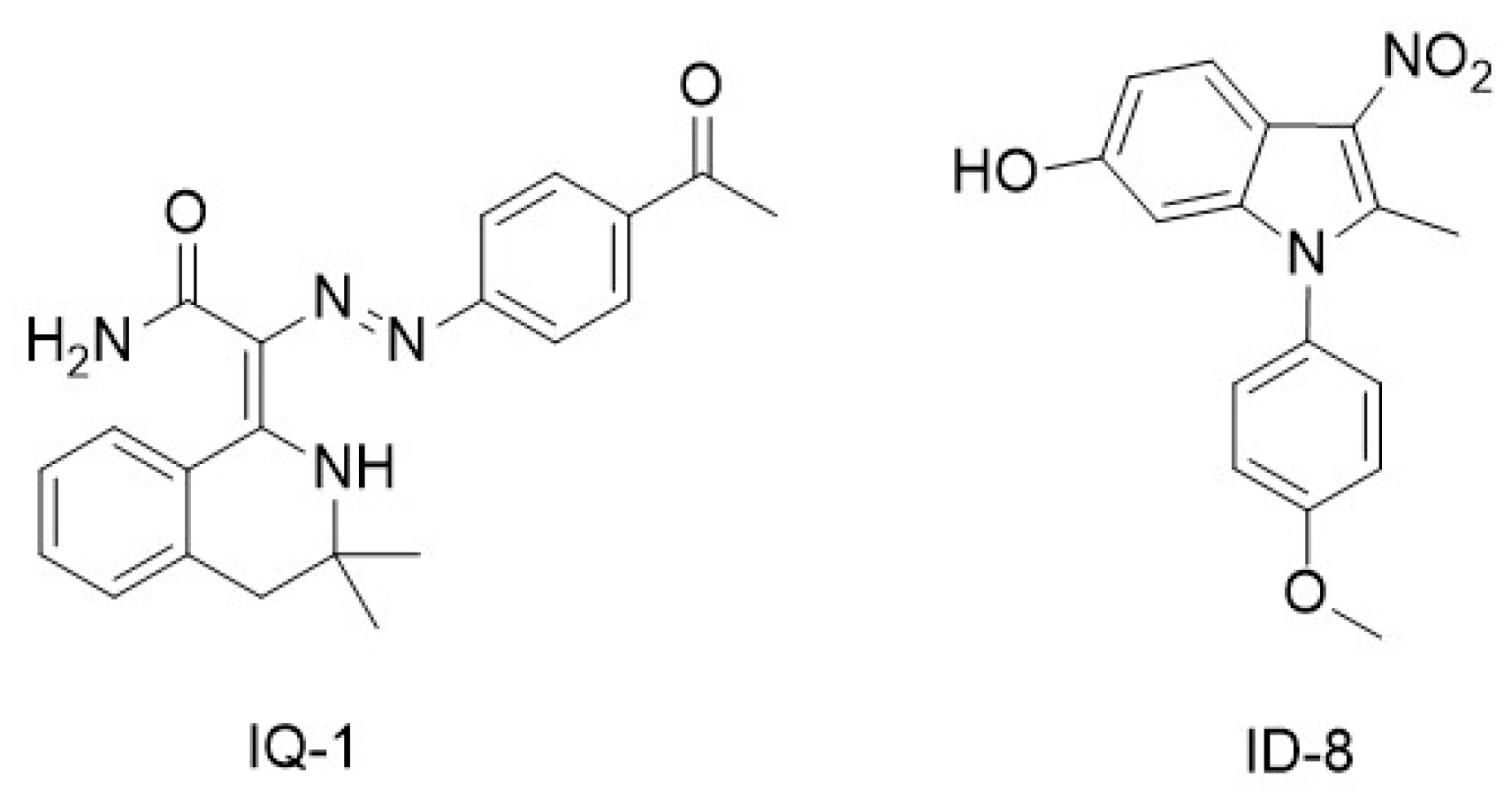
| Non-specific inhibitors of p300/β-catenin signaling | IQ-1, ID-8 | [79,81] |
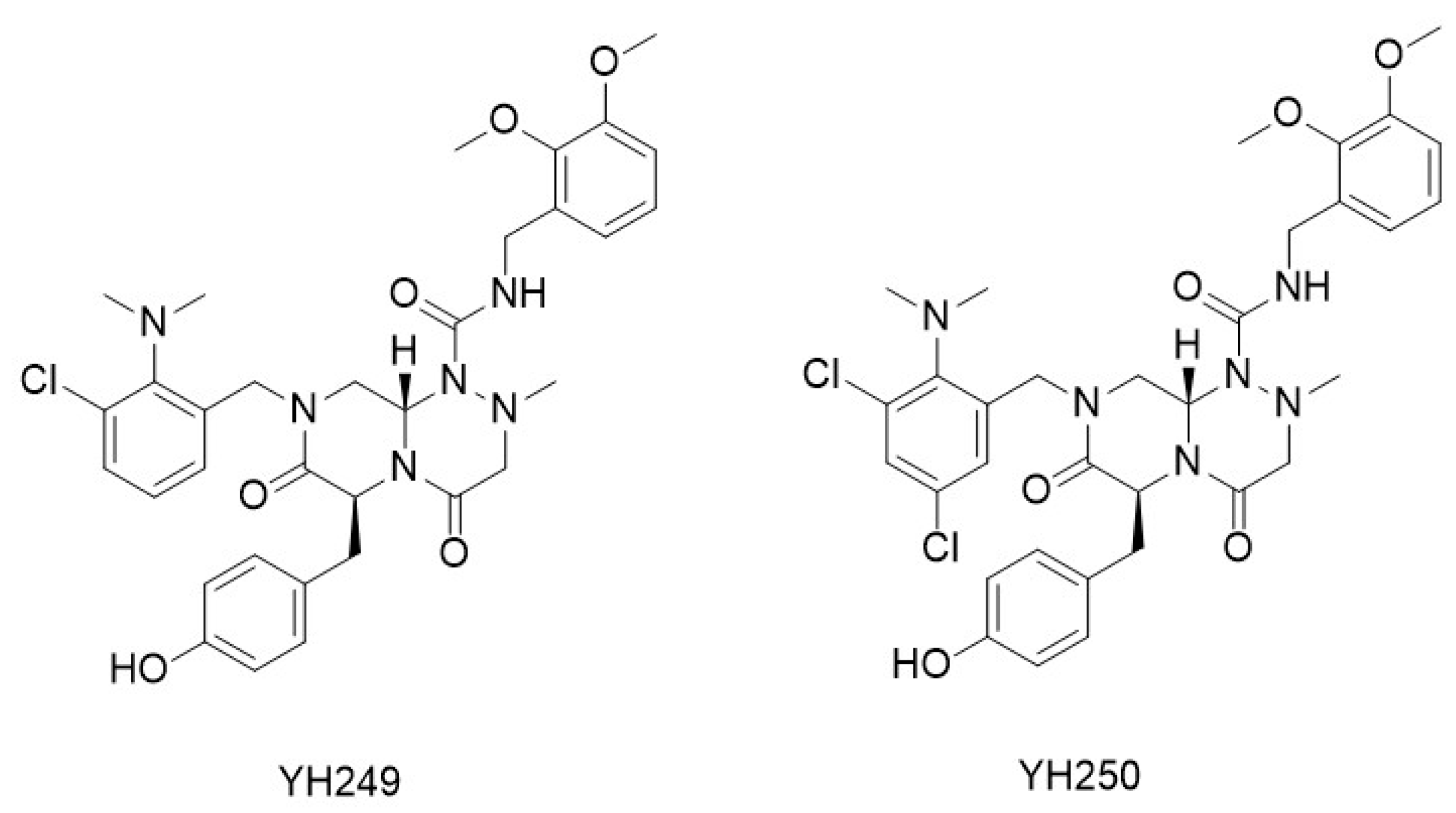
| Specific inhibitors of p300/β-catenin signaling | YH249, YH250 | [76,77,78] |
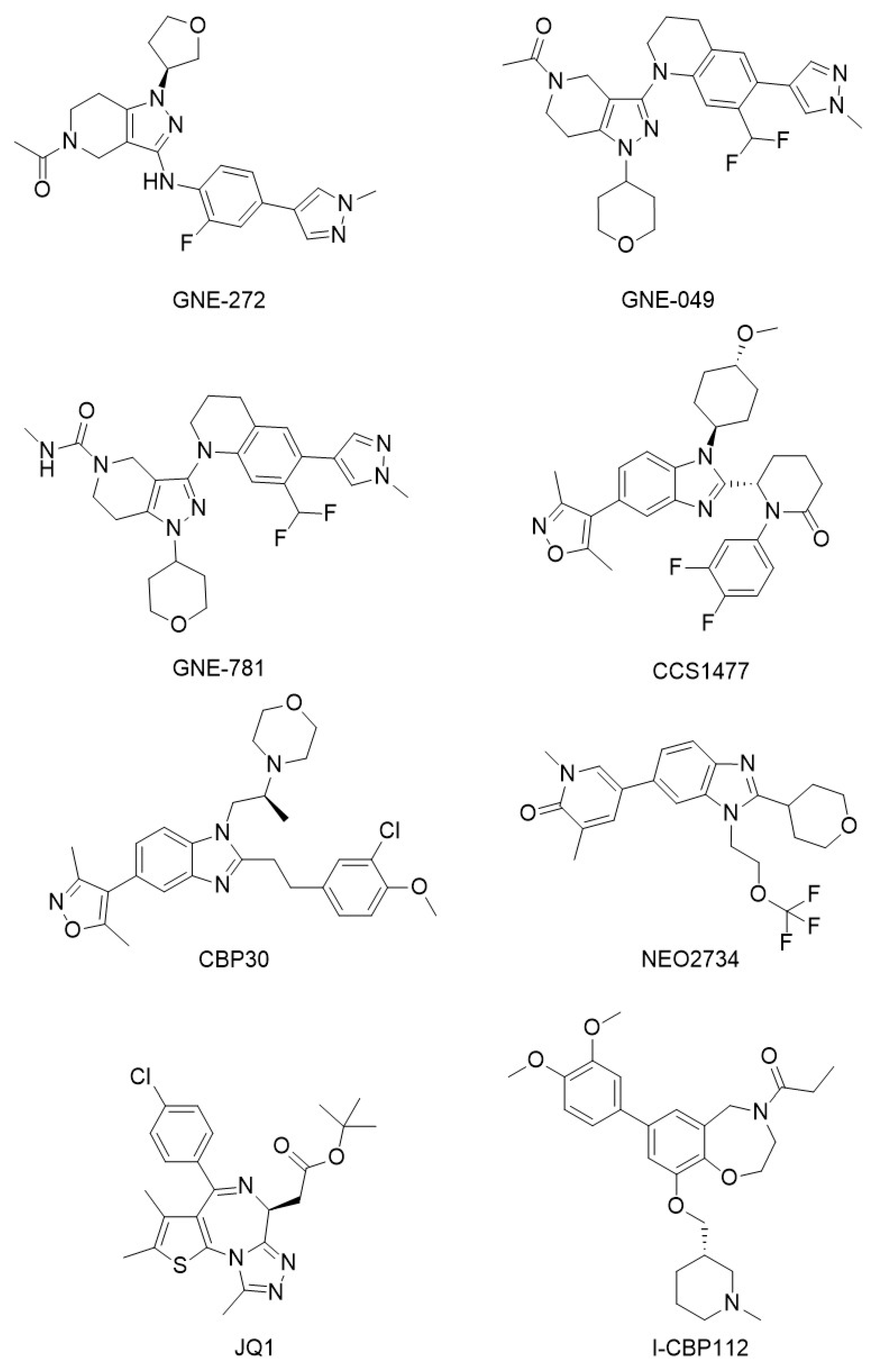
| CBP/p300 bromodomain inhibitors | GNE-272, GNE0-49, GNE-781, CCS1477, CBP30, NEO2734, JQ1, I-CBP112 | [87,88,89,91,94,95,100,102] |
| CBP/p300 HAT inhibitors | A-485, B026, B029-2, C646, CCT077791, CCT077792 | [109,114,115,116,119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masci, D.; Puxeddu, M.; Silvestri, R.; La Regina, G. Targeting CBP and p300: Emerging Anticancer Agents. Molecules 2024, 29, 4524. https://doi.org/10.3390/molecules29194524
Masci D, Puxeddu M, Silvestri R, La Regina G. Targeting CBP and p300: Emerging Anticancer Agents. Molecules. 2024; 29(19):4524. https://doi.org/10.3390/molecules29194524
Chicago/Turabian StyleMasci, Domiziana, Michela Puxeddu, Romano Silvestri, and Giuseppe La Regina. 2024. "Targeting CBP and p300: Emerging Anticancer Agents" Molecules 29, no. 19: 4524. https://doi.org/10.3390/molecules29194524
APA StyleMasci, D., Puxeddu, M., Silvestri, R., & La Regina, G. (2024). Targeting CBP and p300: Emerging Anticancer Agents. Molecules, 29(19), 4524. https://doi.org/10.3390/molecules29194524