
Studies on the Effects of Fermentation on the Phenolic Profile and Biological Activity of Three Cultivars of Kale

 , ,
, ,  ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Results and Discussion
2.1. Phenolic Compounds Profile
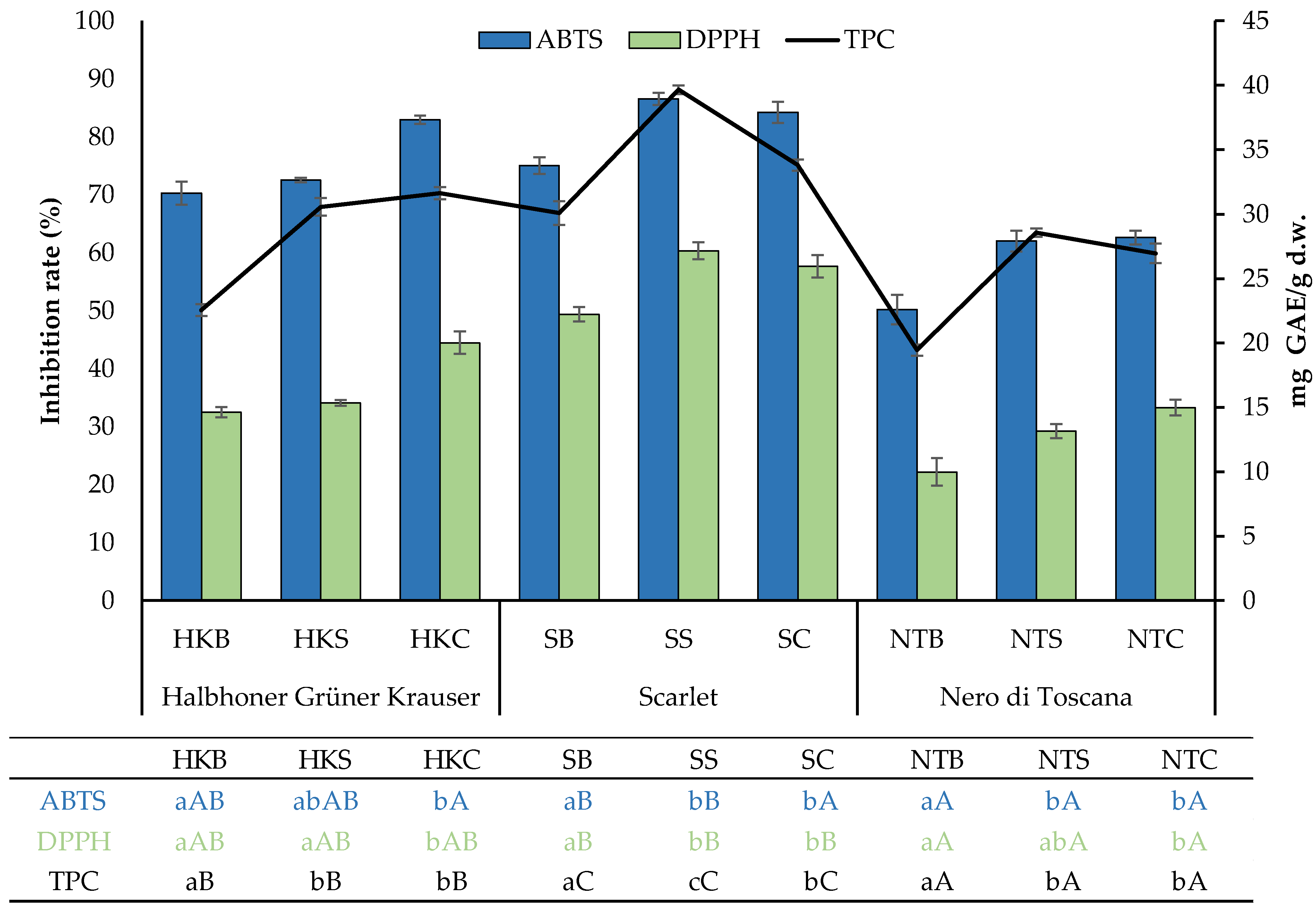
2.2. TPC and Antioxidant Activity
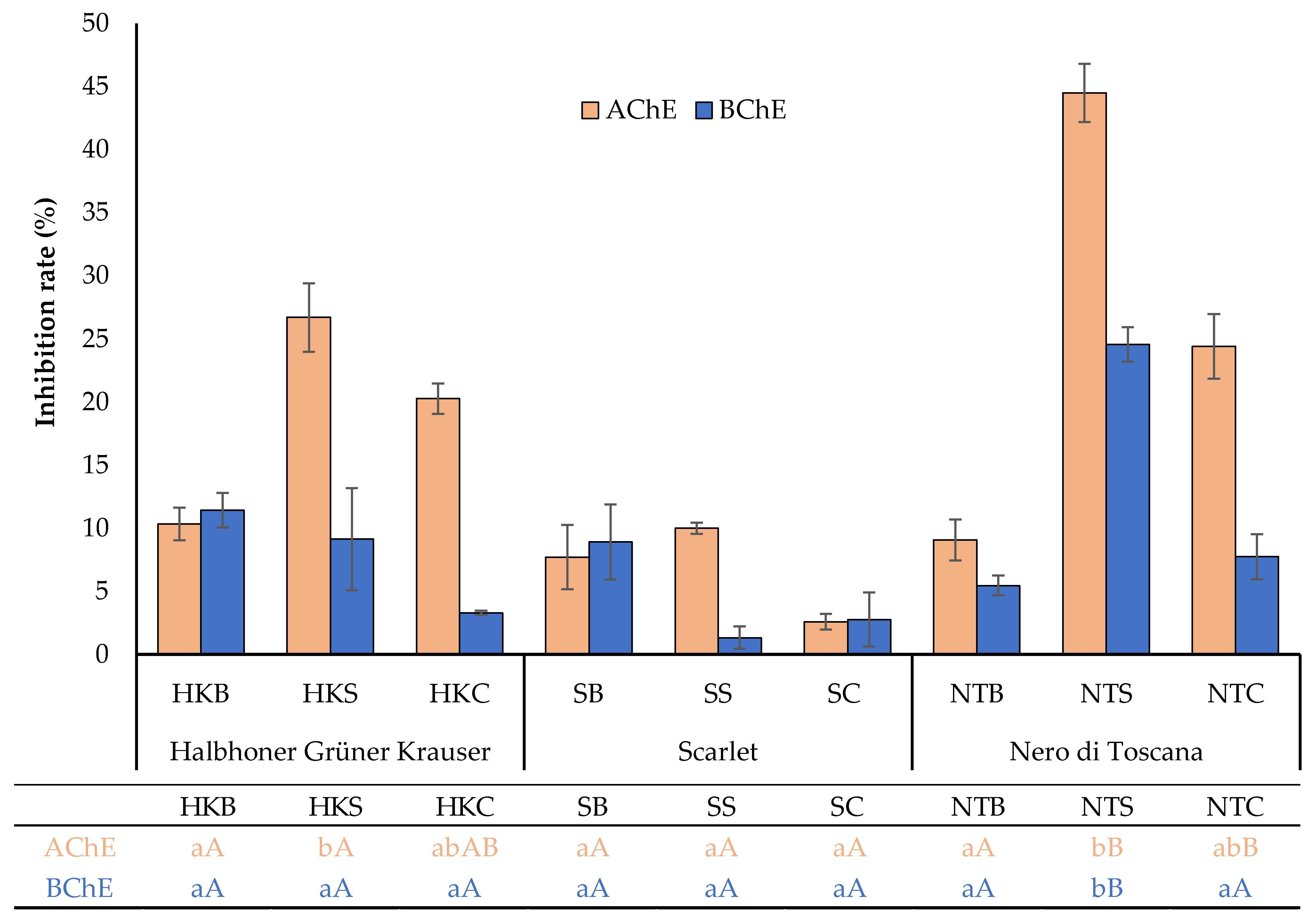
2.3. Cholinesterase Inhibitory Activity
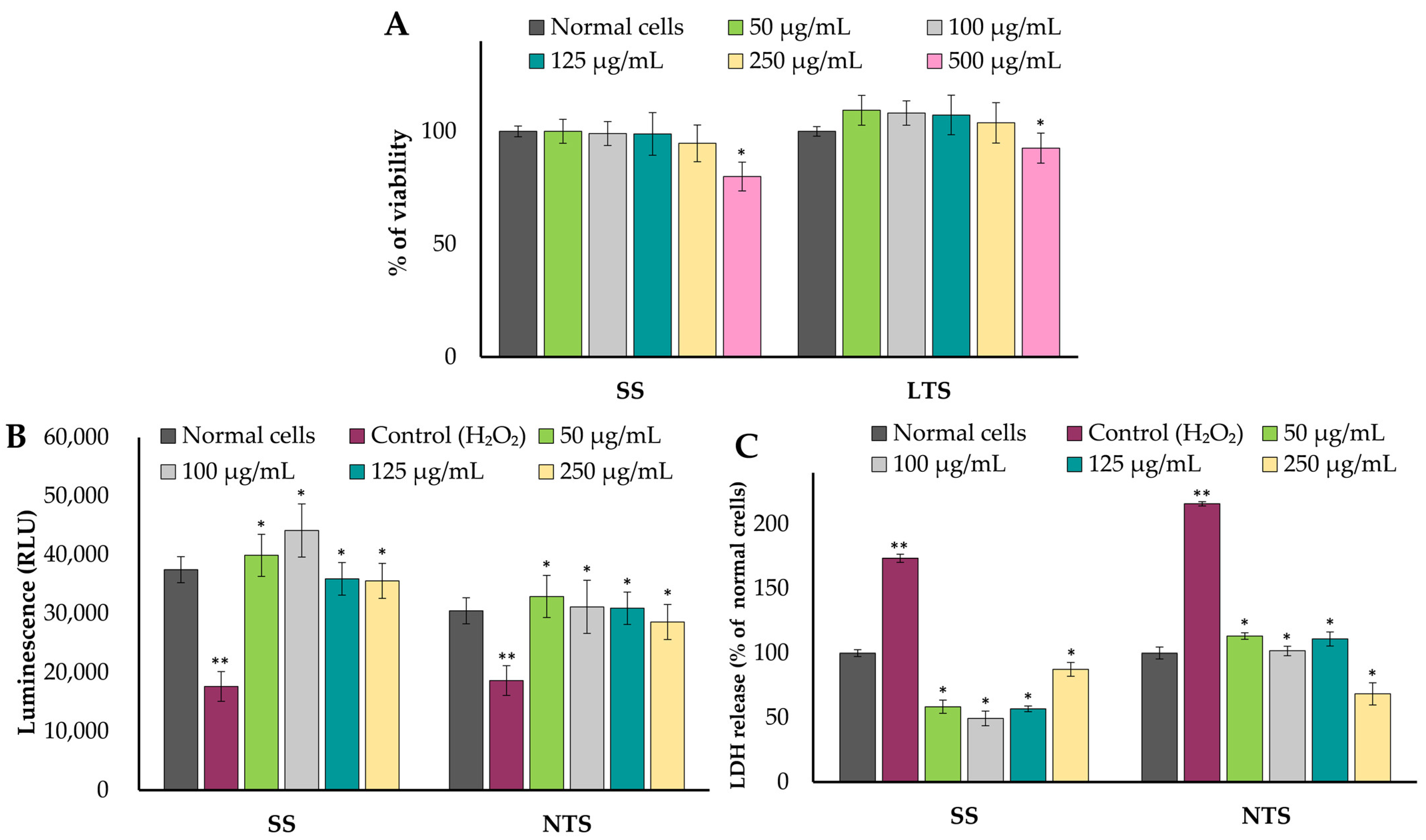
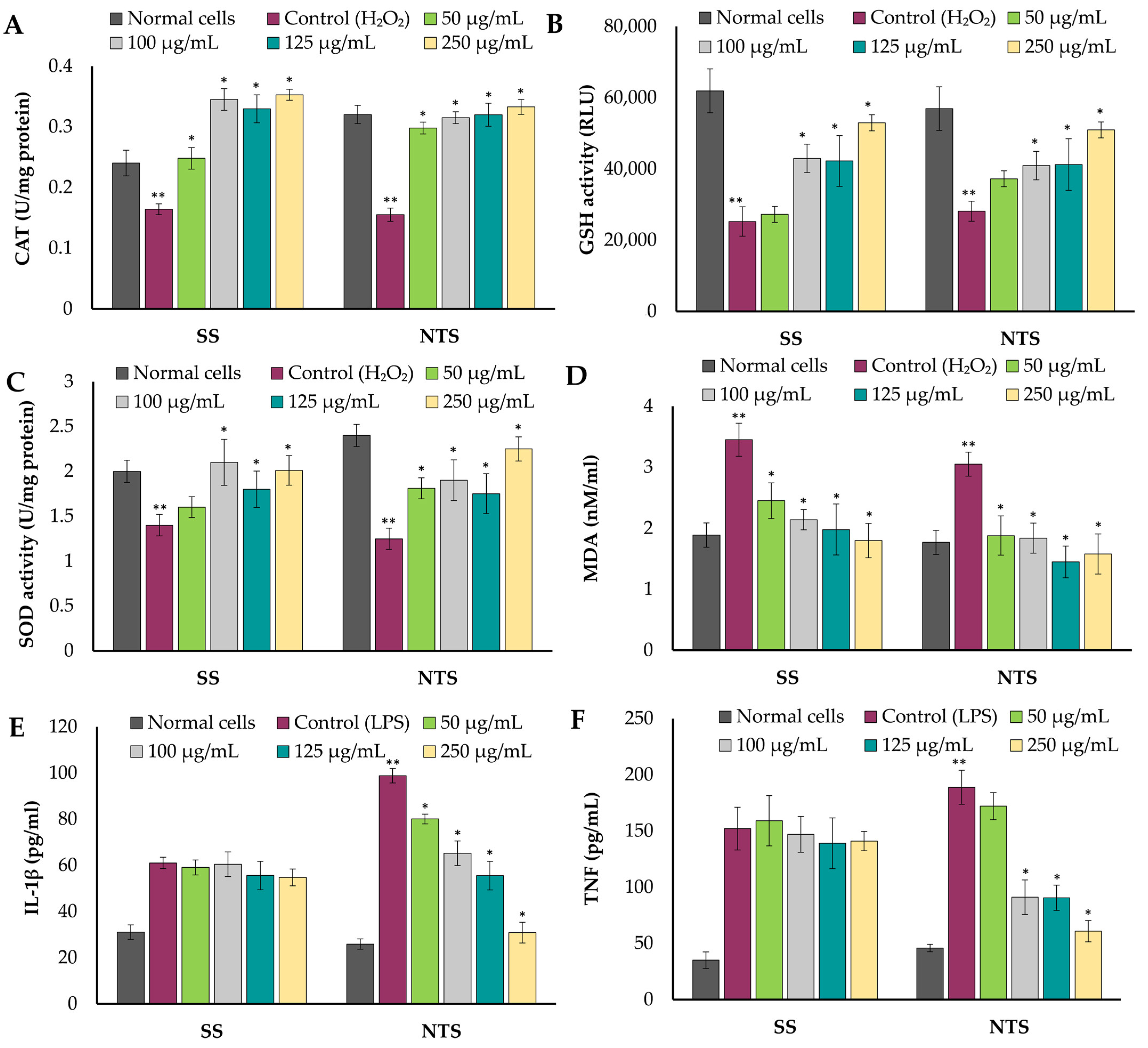
2.4. Cytotoxicity, Cytoprotective, and Immunomodulatory Effects of Extracts on Caco-2 Cell Line
3. Materials and Methods
3.1. Plant Material and Bacterial Strains
3.2. Fermentation Trials
3.3. Extraction Procedure and Extracts Preparation
3.4. Analysis of Polyphenols by HPLC-ESI-QTOF-MS/MS
3.5. Determination of Total Phenolic Content and Antioxidant Activity
3.6. Determination of Cholinesterase Inhibitory Activity
3.7. Cell Cytotoxicity Assay
3.8. Evaluation of the Protective Effect of the Extract
3.8.1. Detection of ATP Content and Cell Membrane Integrity
3.8.2. Determination of Glutathione Levels GSH
3.8.3. Determination of Superoxide Dismutase (SOD) and Catalase (CAT) Activity
3.8.4. Determination of Lipid Membrane Alteration
3.9. Detection of Cytokine Production
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lapuente, M.; Estruch, R.; Shahbaz, M.; Casas, R. Relation of Fruits and Vegetables with Major Cardiometabolic Risk Factors, Markers of Oxidation, and Inflammation. Nutrients 2019, 11, 2381. [Google Scholar] [CrossRef] [PubMed]
- Reda, T.; Thavarajah, P.; Polomski, R.; Bridges, W.; Shipe, E.; Thavarajah, D. Reaching the Highest Shelf: A Review of Organic Production, Nutritional Quality, and Shelf Life of Kale (Brassica oleracea var. acephala). Plants People Planet 2021, 3, 308–318. [Google Scholar] [CrossRef]
- Hahn, C.; Müller, A.; Kuhnert, N.; Albach, D.C. A Cold Case—Glucosinolate Levels in Kale Cultivars Are Differently Influenced by Cold Temperatures. Horticulturae 2023, 9, 953. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a Superfood: Review of the Scientific Evidence behind the Statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Lučić, D.; Pavlović, I.; Brkljačić, L.; Bogdanović, S.; Farkaš, V.; Cedilak, A.; Nanić, L.; Rubelj, I.; Salopek-Sondi, B. Antioxidant and Antiproliferative Activities of Kale (Brassica oleracea L. var. acephala DC.) and Wild Cabbage (Brassica incana Ten.) Polyphenolic Extracts. Molecules 2023, 28, 1840. [Google Scholar] [PubMed]
- Chen, X.; Wang, H.; Huang, X.; Xia, S.; Chen, C.; Nie, Q.; Nie, S. Efficient Enrichment of Total Flavonoids from Kale (Brassica oleracea L. var. acephala L.) Extracts by NKA-9 Resin and Antioxidant Activities of Flavonoids Extract in Vitro. Food Chem. 2022, 374, 131508. [Google Scholar] [CrossRef]
- Hahn, C.; Müller, A.; Kuhnert, N.; Albach, D. Diversity of Kale (Brassica oleracea var. sabellica): Glucosinolate Content and Phylogenetic Relationships. J. Agric. Food Chem. 2016, 64, 3215–3225. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Hernández, E.; Camero-Maldonado, A.V.; Acevedo-Pacheco, L.; Jacobo-Velázquez, D.A.; Antunes-Ricardo, M. Immunomodulatory and Antioxidant Effects of Spray-Dried Encapsulated Kale Sprouts after In Vitro Gastrointestinal Digestion. Foods 2023, 12, 2149. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Mousavi Khaneghah, A.; Barba, F.J.; Nemati, Z.; Sohrabi Shokofti, S.; Alizadeh, F. Fermented Sweet Lemon Juice (Citrus limetta) Using Lactobacillus plantarum LS5: Chemical Composition, Antioxidant and Antibacterial Activities. J. Funct. Foods 2017, 38, 409–414. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In Vitro Comparison of Probiotic Properties of Lactic Acid Bacteria Isolated from Harbin Dry Sausages and Selected Probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous Food Fermentations and Potential Risks for Human Health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food Fermentations: Microorganisms with Technological Beneficial Use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef]
- Sieuwerts, S.; De Bok, F.A.M.; Hugenholtz, J.; Van Hylckama Vlieg, J.E.T. Unraveling Microbial Interactions in Food Fermentations: From Classical to Genomics Approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation Transforms the Phenolic Profiles and Bioactivities of Plant-Based Foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Zhang, M.; Liu, L.; Hu, X.; Liu, J.; Zhou, X.; Chai, Z.; Yin, H. Multiomics Provides Insights into the Succession of Microbiota and Metabolite during Plant Leaf Fermentation. Environ. Res. 2023, 221, 115304. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, U.; Tyagi, A.; Ham, H.J.; Elahi, F.; Oh, D.H. Effect of Fermentation on the Bioactive Compounds of the Black Soybean and Their Anti-Alzheimer’s Activity. Front. Nutr. 2022, 9, 880361. [Google Scholar] [CrossRef]
- Shahbazi, R.; Sharifzad, F.; Bagheri, R.; Alsadi, N.; Yasavoli-Sharahi, H.; Matar, C. Anti-Inflammatory and Immunomodulatory Properties of Fermented Plant Foods. Nutrients 2021, 13, 1516. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Yeboah, P.J.; Ayivi, R.D.; Eddin, A.S.; Wijemanna, N.D.; Paidari, S.; Bakhshayesh, R.V. A Review and Comparative Perspective on Health Benefits of Probiotic and Fermented Foods. Int. J. Food Sci. Technol. 2023, 58, 4948–4964. [Google Scholar] [CrossRef]
- Knez, E.; Kadac-Czapska, K.; Grembecka, M. Effect of Fermentation on the Nutritional Quality of the Selected Vegetables and Legumes and Their Health Effects. Life 2023, 13, 655. [Google Scholar] [CrossRef] [PubMed]
- Gaur, G.; Gänzle, M.G. Conversion of (Poly)Phenolic Compounds in Food Fermentations by Lactic Acid Bacteria: Novel Insights into Metabolic Pathways and Functional Metabolites. Curr. Res. Food Sci. 2023, 6, 100448. [Google Scholar] [CrossRef] [PubMed]
- Olsen, H.; Grimmer, S.; Aaby, K.; Saha, S.; Borge, G.I.A. Antiproliferative Effects of Fresh and Thermal Processed Green and Red Cultivars of Curly Kale (Brassica oleracea L. convar. acephala Var. sabellica). J. Agric. Food Chem. 2012, 60, 7375–7383. [Google Scholar] [CrossRef] [PubMed]
- Ferioli, F.; Giambanelli, E.; D’Antuono, L.F.; Costa, H.S.; Albuquerque, T.G.; Silva, A.S.; Hayran, O.; Koçaoglu, B. Comparison of Leafy Kale Populations from Italy, Portugal, and Turkey for Their Bioactive Compound Content: Phenolics, Glucosinolates, Carotenoids, and Chlorophylls. J. Sci. Food Agric. 2013, 93, 3478–3489. [Google Scholar] [CrossRef]
- Liu, Z.; Shi, J.; Wan, J.; Pham, Q.; Zhang, Z.; Sun, J.; Yu, L.; Luo, Y.; Wang, T.T.Y.; Chen, P. Profiling of Polyphenols and Glucosinolates in Kale and Broccoli Microgreens Grown under Chamber and Windowsill Conditions by Ultrahigh-Performance Liquid Chromatography High-Resolution Mass Spectrometry. ACS Food Sci. Technol. 2022, 2, 101–113. [Google Scholar] [CrossRef]
- Bianchi, G.; Picchi, V.; Tava, A.; Doria, F.; Walley, P.G.; Dever, L.; di Bella, M.C.; Arena, D.; Ben Ammar, H.; Lo Scalzo, R.; et al. Insights into the Phytochemical Composition of Selected Genotypes of Organic Kale (Brassica oleracea L. var. acephala). J. Food Compos. Anal. 2024, 125, 105721. [Google Scholar] [CrossRef]
- Ferreres, F.; Llorach, R.; Gil-Izquierdo, A. Characterization of the Interglycosidic Linkage in Di-, Tri-, Tetra- and Pentaglycosylated Flavonoids and Differentiation of Positional Isomers by Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry. J. Mass Spectrom. 2004, 39, 312–321. [Google Scholar] [CrossRef]
- Ferreres, F.; Fernandes, F.; Sousa, C.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Metabolic and Bioactivity Insights into Brassica oleracea var. acephala. J. Agric. Food Chem. 2009, 57, 8884–8892. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Waśko, A.; Polak-Berecka, M. Fermented Curly Kale as a New Source of Gentisic and Salicylic Acids with Antitumor Potential. J. Funct. Foods 2020, 67, 103866. [Google Scholar] [CrossRef]
- Szutowska, J.; Rybicka, I.; Pawlak-Lemańska, K.; Gwiazdowska, D. Spontaneously Fermented Curly Kale Juice: Microbiological Quality, Nutritional Composition, Antioxidant, and Antimicrobial Properties. J. Food Sci. 2020, 85, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-Based Biotransformation of Bioactive Phenolics and Volatile Compounds from Cashew Apple Juice by Select Lactic Acid Bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Lactobacillus plantarum WCFS1 Fermentation Differentially Affects Antioxidant Capacity and Polyphenol Content in Mung Bean (Vigna radiata) and Soya Bean (Glycine max) Milks. J. Food Process. Preserv. 2017, 41, e12944. [Google Scholar] [CrossRef]
- Szutowska, J.; Gwiazdowska, D.; Rybicka, I.; Pawlak-Lemańska, K.; Biegańska-Marecik, R.; Gliszczyńska-Świgło, A. Controlled Fermentation of Curly Kale Juice with the Use of Autochthonous Starter Cultures. Food Res. Int. 2021, 149, 110674. [Google Scholar] [CrossRef]
- Michalak, M.; Kubik-Komar, A.; Waśko, A.; Polak-Berecka, M. Starter Culture for Curly Kale Juice Fermentation Selected Using Principal Component Analysis. Food Biosci. 2020, 35, 100602. [Google Scholar] [CrossRef]
- Albertini, B.; Schoubben, A.; Guarnaccia, D.; Pinelli, F.; Della Vecchia, M.; Ricci, M.; Di Renzo, G.C.; Blasi, P. Effect of Fermentation and Drying on Cocoa Polyphenols. J. Agric. Food Chem. 2015, 63, 9948–9953. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Razola-Díaz, M.d.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of Lactic Acid Bacteria Fermentation on Phenolic Compounds and Antioxidant Activity of Avocado Leaf Extracts. Antioxidants 2023, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Quispe, C.; Butnariu, M.; Sarac, I.; Marmouzi, I.; Kamle, M.; Tripathi, V.; Kumar, P.; Bouyahya, A.; Capanoglu, E.; et al. Phytotherapy and Food Applications from Brassica Genus. Phytother. Res. 2021, 35, 3590–3609. [Google Scholar] [CrossRef]
- Podsedek, A. Natural Antioxidants and Antioxidant Capacity of Brassica Vegetables: A Review. LWT—Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Ramirez, D.; Abellán-Victorio, A.; Beretta, V.; Camargo, A.; Moreno, D.A. Functional Ingredients from Brassicaceae Species: Overview and Perspectives. Int. J. Mol. Sci. 2020, 21, 1998. [Google Scholar] [CrossRef] [PubMed]
- Bhanja Dey, T.; Chakraborty, S.; Jain, K.K.; Sharma, A.; Kuhad, R.C. Antioxidant Phenolics and Their Microbial Production by Submerged and Solid State Fermentation Process: A Review. Trends Food Sci. Technol. 2016, 53, 60–74. [Google Scholar] [CrossRef]
- Kim, G.Y.; Kim, S.A.; Kong, S.Y.; Seong, H.; Bae, J.H.; Han, N.S. Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800. Antioxidants 2023, 12, 1850. [Google Scholar] [CrossRef]
- Johnson, M.H.; Lucius, A.; Meyer, T.; Gonzalez De Mejia, E. Cultivar Evaluation and Effect of Fermentation on Antioxidant Capacity and in vitro Inhibition of α-Amylase and α-Glucosidase by Highbush Blueberry (Vaccinium corymbosum). J. Agric. Food Chem. 2011, 59, 8923–8930. [Google Scholar] [CrossRef]
- Xiong, T.; Li, X.; Guan, Q.; Peng, F.; Xie, M. Starter Culture Fermentation of Chinese Sauerkraut: Growth, Acidification and Metabolic Analyses. Food Control 2014, 41, 122–127. [Google Scholar] [CrossRef]
- Wilson Iii, D.M.; Cookson, M.R.; Van, L.; Bosch, D.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Leading Edge Hallmarks of Neurodegenerative Diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Ahn, J.W.; Kim, H.S.; Choi, Y.; Jeong, J.; Joo, S.S.; Park, D. Improvement of Cognitive Function by Fermented Panax Ginseng C.A. Meyer Berries Extracts in an AF64A-Induced Memory Deficit Model. Nutrients 2023, 15, 3389. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.N.S.; Ryu, J.K.; Kim, Y.; Jeon, B.H. GABA-Enriched Fermented Laminaria Japonica Improves Cognitive Impairment and Neuroplasticity in Scopolamine- and Ethanol-Induced Dementia Model Mice. Nutr. Res. Pract. 2018, 12, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.Y.; Lee, D.S.; Park, S.K.; Ha, J.S.; Kim, J.M.; Ha, G.J.; Seo, W.T.; Heo, H.J. Cognitive Function of Artemisia Argyi H. Fermented by Monascus Purpureus under TMT-Induced Learning and Memory Deficits in ICR Mice. Evid.-Based Complement. Altern. Med. 2017, 2017, 5809370. [Google Scholar] [CrossRef]
- Yoo, D.H.; Kim, D.H. Lactobacillus pentosus var. plantarum C29 Increases the Protective Effect of Soybean against Scopolamine-Induced Memory Impairment in Mice. Int. J. Food Sci. Nutr. 2015, 66, 912–918. [Google Scholar] [CrossRef]
- Park, H.R.; Lee, H.; Park, H.; Cho, W.K.; Ma, J.Y. Fermented Sipjeondaebo-Tang Alleviates Memory Deficits and Loss of Hippocampal Neurogenesis in Scopolamine-Induced Amnesia in Mice. Sci. Rep. 2016, 6, 22405. [Google Scholar] [CrossRef]
- Boǧa, M.; Hacibekiroǧlu, I.; Kolak, U. Antioxidant and Anticholinesterase Activities of Eleven Edible Plants. Pharm. Biol. 2011, 49, 290–295. [Google Scholar] [CrossRef]
- Mollica, A.; Stefanucci, A.; Zengin, G.; Locatelli, M.; Macedonio, G.; Orlando, G.; Ferrante, C.; Menghini, L.; Recinella, L.; Leone, S.; et al. Polyphenolic Composition, Enzyme Inhibitory Effects Ex-Vivo and in-Vivo Studies on Two Brassicaceae of North-Central Italy. Biomed. Pharmacother. 2018, 107, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Stefanucci, A.; Zengin, G.; Llorent-Martinez, E.J.; Dimmito, M.P.; Della Valle, A.; Pieretti, S.; Ak, G.; Sinan, K.I.; Mollica, A. Chemical Characterization, Antioxidant Properties and Enzyme Inhibition of Rutabaga Root’s Pulp and Peel (Brassica napus L.). Arab. J. Chem. 2020, 13, 7078–7086. [Google Scholar] [CrossRef]
- Geraris Kartelias, I.; Karantonis, H.C.; Giaouris, E.; Panagiotakopoulos, I.; Nasopoulou, C. Kombucha Fermentation of Olympus Mountain Tea (Sideritis scardica) Sweetened with Thyme Honey: Physicochemical Analysis and Evaluation of Functional Properties. Foods 2023, 12, 3496. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, R.; Beaufort, S.; Villarreal-Soto, S.A.; Taillandier, P.; Bouajila, J.; Debouba, M. Kombucha Fermentation of African Mustard (Brassica tournefortii) Leaves: Chemical Composition and Bioactivity. Food Biosci. 2019, 30, 100414. [Google Scholar] [CrossRef]
- Ademosun, A.O.; Oboh, G.; Bello, F.; Ayeni, P.O. Antioxidative Properties and Effect of Quercetin and Its Glycosylated Form (Rutin) on Acetylcholinesterase and Butyrylcholinesterase Activities. J. Evid.-Based Complement. Altern. Med. 2016, 21, NP11–NP17. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhang, G.; Liao, Y.; Fei, X.; Gong, D.; Hu, X. Anti-Acetylcholinesterase Mechanism of Kaempferol and Its Synergistic Effect with Galanthamine Hydrobromide. Food Biosci. 2023, 56, 103174. [Google Scholar] [CrossRef]
- Janarny, G.; Gunathilake, K.D.P.P. Changes in Rice Bran Bioactives, Their Bioactivity, Bioaccessibility and Bioavailability with Solid-State Fermentation by Rhizopus oryzae. Biocatal. Agric. Biotechnol. 2020, 23, 101510. [Google Scholar] [CrossRef]
- Morais, S.G.G.; da Silva Campelo Borges, G.; dos Santos Lima, M.; Martín-Belloso, O.; Magnani, M. Effects of Probiotics on the Content and Bioaccessibility of Phenolic Compounds in Red Pitaya Pulp. Food Res. Int. 2019, 126, 108681. [Google Scholar] [CrossRef] [PubMed]
- Larocca, M.; Perna, A.M.; Simonetti, A.; Gambacorta, E.; Iannuzzi, A.; Perucatti, A.; Rossano, R. Antioxidant and Anti-Inflammatory Effects of Cauliflower Leaf Powder-Enriched Diet against LPS Induced Toxicity in Rabbits. Food Funct. 2017, 8, 3288–3296. [Google Scholar] [CrossRef] [PubMed]
- Iga-Buitrón, D.; Torres-Maravilla, E.; Bermúdez-Humaran, L.G.; Ascacio-Valdes, J.A.; Rodríguez-Herrera, R.; Aguilar, C.N.; Flores-Gallegos, A.C. Lactic Fermentation of Broccoli (Brassica oleracea var. italica) to Enhance the Antioxidant and Antiproliferative Activities. Fermentation 2023, 9, 122. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, S.; Cai, Q.; Miao, J.; Li, J. Antioxidant Capacity and Protective Effects on H2O2-Induced Oxidative Damage in PC12 Cells of the Active Fraction of Brassica rapa L. Foods 2023, 12, 2075. [Google Scholar] [CrossRef]
- Hong, J.Y.; Kim, H.; Lee, J.; Jeon, W.J.; Baek, S.H.; Ha, I.H. Neurotherapeutic Effect of Inula britannica var. chinensis against H2O2-Induced Oxidative Stress and Mitochondrial Dysfunction in Cortical Neurons. Antioxidants 2021, 10, 375. [Google Scholar] [CrossRef]
- An, J.; Liu, J.; Liang, Y.; Ma, Y.; Chen, C.; Cheng, Y.; Peng, P.; Zhou, N.; Zhang, R.; Addy, M.; et al. Characterization, Bioavailability and Protective Effects of Phenolic-Rich Extracts from Almond Hulls against pro-Oxidant Induced Toxicity in Caco-2 Cells. Food Chem. 2020, 322, 126742. [Google Scholar] [CrossRef]
- Li, R.L.; Zhang, Q.; Liu, J.; Sun, J.Y.; He, L.Y.; Duan, H.X.; Peng, W.; Wu, C.J. Hydroxy-α-Sanshool Possesses Protective Potentials on H2O2-Stimulated PC12 Cells by Suppression of Oxidative Stress-Induced Apoptosis through Regulation of PI3K/Akt Signal Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 3481758. [Google Scholar] [CrossRef]
- Radermacher, K.A.; Wingler, K.; Langhauser, F.; Altenhöfer, S.; Kleikers, P.; Hermans, J.J.R.; Hrabě De Angelis, M.; Kleinschnitz, C.; Schmidt, H.H.H.W. Neuroprotection After Stroke by Targeting NOX4 As a Source of Oxidative Stress. Antioxid. Redox Signal. 2013, 18, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Sim, H.W.; Lee, W.Y.; Lee, R.; Yang, S.Y.; Ham, Y.K.; Lim, S.D.; Park, H.J. The Anti-Inflammatory Effects of Broccoli (Brassica oleracea L. var. italica) Sprout Extract in RAW 264.7 Macrophages and a Lipopolysaccharide-Induced Liver Injury Model. Curr. Issues Mol. Biol. 2023, 45, 9117–9131. [Google Scholar] [CrossRef]
- Kubina, R.; Iriti, M.; Kabała-Dzik, A. Anticancer Potential of Selected Flavonols: Fisetin, Kaempferol, and Quercetin on Head and Neck Cancers. Nutrients 2021, 13, 845. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Parrish, W.R.; Rosas-Ballina, M.; Ochani, M.; Puerta, M.; Ochani, K.; Chavan, S.; Al-Abed, Y.; Tracey, K.J. Brain Acetylcholinesterase Activity Controls Systemic Cytokine Levels through the Cholinergic Anti-Inflammatory Pathway. Brain Behav. Immun. 2009, 23, 41–45. [Google Scholar] [CrossRef]
- Michalak, M.; Gustaw, K.; Waśko, A.; Polak-Berecka, M. Composition of Lactic Acid Bacteria during Spontaneous Curly Kale (Brassica oleracea var. sabellica) Fermentation. Microbiol. Res. 2018, 206, 121–130. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A Novel Method for Measuring Antioxidant Capacity and Its Application to Monitoring the Antioxidant Status in Premature Neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Baranowska-Wójcik, E.; Szwajgier, D.; Winiarska-Mieczan, A. Regardless of the Brewing Conditions, Various Types of Tea Are a Source of Acetylcholinesterase Inhibitors. Nutrients 2020, 12, 709. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Ion. (+/−) | Rt. (min) | Molecular Formula | m/z Calculated | m/z Experimental | Delta (mmu) | RDB | Proposed Compound | Sample |
|---|---|---|---|---|---|---|---|---|---|
| 1 | − | 2.19 | C7H12O6 | 191.0561 | 191.0512 | −0.46 | 3.5 | Quinic acid | All |
| 2 | − | 3.52 | C6H8O7 | 191.0197 | 191.0207 | −5.07 | 3 | Citric acid | All |
| 3 | − | 17.26/ 17.76/ 20.09 | C16H18O8 | 337.0929 | 337.0927 | 0.57 | 8 | Coumaroyl-quinic acid isomers | All |
| 4 | − | 15.26/ 18.59 | C9H10O4 | 181.0500 | 181.0505/ 181.0524/ | 0.73/−9.71 | 5 | 4-hydroxy-3-methoxy-phenylactetic acid (homovanillic acid) | All except SB |
| 5 | − | 15.94 | C16H18O9 | 353.0872 | 353.0877 | 0.3 | 8 | Chlorogenic acid | All |
| 6 | − | 16.61 | C16H18O9 | 353.0872 | 353.0880 | −0.55 | 8 | Neochlorogenic acid | All |
| 7 | − | 17.1 | C33H40O22 | 787.1938 | 787.1944 | −0.7 | 14 | Quercetin-hexoside-dihexoside | All except NTC |
| 8 | − | 17.59 | C7H6O4 | 153.0202 | 153.0195 | −1.09 | 5 | Protocatechuic acid | All except HKB, SB and NTB |
| 9 | − | 17.69 | C20H16O12 | 447.0569 | 447.0539 | 6.69 | 13 | Ellagic acid rhamnoside | All |
| 10 | − | 18.86 | C16H18O9 | 353.0872 | 353.0879 | −0.27 | 8 | (z)-Chlorogenic acid | All |
| 11 | − | 19.2 | C16H20N2O6S | 367.0969 | 367.1005 | −9.7 | 8 | Desulfo-glucobrassicin | All except SC |
| 12 | − | 21.8 | C27H30O17 | 625.141 | 625.1425 | −2.36 | 13 | Quercetin dihexoside | All |
| 13 | − | 23.11 | C27H30O16 | 609.144 | 609.1482 | −3.43 | 13 | Rutin | All |
| 14 | − | 24.28 | C21H20O12 | 463.0882 | 463.0898 | −3.45 | 12 | Isoquercetin | All |
| 15 | − | 24.44 | C17H22N2O7S | 397.1075 | 397.1035 | 10.04 | 8 | Desulfo-neoglucobrassicin | All |
| 16 | − | 24.85 | C11H12O5 | 223.0626 | 223.0635 | −10.28 | 5 | Sinapinic acid | HKS, HKC, NTS and NTC |
| 17 | − | 25.4 | C21H20O11 | 447.0933 | 447.0946 | −2.93 | 12 | Quercitrin | All |
| 18 | − | 28.113 | C17H22N2O7S | 397.1075 | 397.1039 | 9.03 | 8 | Desulfo-4-methoxyglucobrassicin | All |
| 19 | − | 29.69 | C15H10O7 | 301.0348 | 301.0358 | −1.4 | 11 | Quercetin | All except SB |
| 20 | − | 31.68 | C15H12O5 | 271.0612 | 271.0639 | −9.93 | 10 | Naringin | SC |
| 21 | − | 31.94 | C15H10O6 | 285.0399 | 285.0404 | 0.22 | 11 | Kaempferol | All except SB |
| Compound | Halbhoner Grüner Krauser | Scarlet | Nero di Toscana | ||||||
|---|---|---|---|---|---|---|---|---|---|
| HKB | HKS | HKC | SB | SS | SC | NTB | NTS | NTC | |
| Quinic acid | 8.05 | 20.86 | 19.90 | 100.00 | 7.75 | 8.78 | 10.06 | 12.65 | 25.13 |
| Citric acid | 51.60 | 20.21 | 46.55 | 100.00 | 26.27 | 10.89 | 37.78 | 14.38 | 8.46 |
| Chlorogenic acid | 100.00 | 96.63 | 9.96 | 25.12 | 48.55 | 53.00 | 59.89 | 36.51 | 42.55 |
| Neochlorogenic acid | 100.00 | 87.72 | 82.22 | 37.21 | 2.06 | 64.43 | 82.82 | 43.16 | 46.08 |
| (z)-Chlorogenic acid | 14.67 | 51.10 | 34.07 | 100.00 | 46.36 | 59.53 | 48.59 | 11.96 | 14.59 |
| Coumaroyl-quinic acid isomer | 27.77 | 45.21 | 29.89 | 15.07 | 13.16 | 13.48 | 43.75 | 72.97 | 100.00 |
| Rutin | 2.97 | 100.00 | 70.93 | 1.37 | 23.15 | 19.38 | 1.42 | 65.36 | 61.76 |
| Isoquercetin | 24.97 | 66.82 | 100.00 | 6.61 | 71.65 | 66.34 | 18.85 | 39.25 | 30.19 |
| 4-hydroxy-3-methoxy-phenylactetic acid | 22.12 | 66.81 | 85.57 | 0.00 | 100.00 | 98.78 | 13.08 | 95.68 | 97.77 |
| Protocatechuic acid | 0.00 | 27.39 | 26.84 | 0.00 | 52.53 | 46.47 | 0.00 | 40.91 | 100.00 |
| Quercetin | 16.81 | 6.23 | 99.13 | 0.00 | 97.62 | 100.00 | 11.24 | 3.97 | 45.62 |
| Kaempferol | 5.41 | 70.21 | 53.84 | 0.00 | 29.08 | 31.86 | 8.25 | 100.00 | 76.40 |
| Naringin | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 100.00 | 0.00 | 0.00 | 0.00 |
| Sinapinic acid | 0.00 | 100.00 | 96.41 | 0.00 | 0.00 | 0.00 | 0.00 | 63.91 | 71.51 |
| Quercetin dihexoside | 37.24 | 93.23 | 62.37 | 0.62 | 100.00 | 90.34 | 29.80 | 24.19 | 32.31 |
| Quercetine-hexoside-dihexoside | 64.09 | 11.67 | 10.83 | 100.00 | 26.71 | 17.16 | 33.95 | 5.36 | 0.11 |
| Desulfo-neoglucobrassicin | 29.85 | 31.93 | 54.19 | 21.34 | 100.00 | 81.79 | 22.19 | 42.32 | 56.05 |
| Desulfo-4-methoxyglucobrassicin | 16.11 | 100.00 | 96.41 | 11.85 | 65.42 | 61.32 | 12.52 | 82.66 | 98.87 |
| Quercitrin | 98.53 | 15.73 | 2.18 | 19.71 | 4.14 | 0.40 | 100.00 | 19.81 | 3.47 |
| Desulfo-glucobrassicin | 100.00 | 88.06 | 82.86 | 54.81 | 64.07 | 0.00 | 68.68 | 0.01 | 58.81 |
| ABTS | DPPH | TPC | AChE | BChE | |
|---|---|---|---|---|---|
| ABTS | 1.00 | 0.93 ** | 0.91 ** | −0.39 * | −0.61 ** |
| DPPH | 0.93 ** | 1.00 | 0.89 ** | −0.44 * | −0.60 ** |
| TPC | 0.91 ** | 0.89 ** | 1.00 | −0.20 | −0.59 ** |
| AChE | −0.39 * | −0.44 * | −0.20 | 1.00 | 0.49 ** |
| BChE | −0.61 ** | −0.60 ** | −0.59 ** | 0.49 ** | 1.00 |
| Kale Cultivar | Type of Sample | Sample Code |
|---|---|---|
| ‘Halbhoner Grüner Krauser’ | Fresh leaves, before fermentation | HKB |
| Leaves subjected to spontaneous fermentation | HKS | |
| Leaves fermented using a starter culture | HKC | |
| ‘Scarlet’ | Fresh leaves, before fermentation | SB |
| Leaves subjected to spontaneous fermentation | SS | |
| Leaves fermented using a starter culture | SC | |
| ‘Nero di Toscana’ | Fresh leaves, before fermentation | NTB |
| Leaves subjected to spontaneous fermentation | NTS | |
| Leaves fermented using a starter culture | NTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalak-Tomczyk, M.; Rymuszka, A.; Kukula-Koch, W.; Szwajgier, D.; Baranowska-Wójcik, E.; Jachuła, J.; Welman-Styk, A.; Kędzierska, K. Studies on the Effects of Fermentation on the Phenolic Profile and Biological Activity of Three Cultivars of Kale. Molecules 2024, 29, 1727. https://doi.org/10.3390/molecules29081727
Michalak-Tomczyk M, Rymuszka A, Kukula-Koch W, Szwajgier D, Baranowska-Wójcik E, Jachuła J, Welman-Styk A, Kędzierska K. Studies on the Effects of Fermentation on the Phenolic Profile and Biological Activity of Three Cultivars of Kale. Molecules. 2024; 29(8):1727. https://doi.org/10.3390/molecules29081727
Chicago/Turabian StyleMichalak-Tomczyk, Magdalena, Anna Rymuszka, Wirginia Kukula-Koch, Dominik Szwajgier, Ewa Baranowska-Wójcik, Jacek Jachuła, Agnieszka Welman-Styk, and Kinga Kędzierska. 2024. "Studies on the Effects of Fermentation on the Phenolic Profile and Biological Activity of Three Cultivars of Kale" Molecules 29, no. 8: 1727. https://doi.org/10.3390/molecules29081727
APA StyleMichalak-Tomczyk, M., Rymuszka, A., Kukula-Koch, W., Szwajgier, D., Baranowska-Wójcik, E., Jachuła, J., Welman-Styk, A., & Kędzierska, K. (2024). Studies on the Effects of Fermentation on the Phenolic Profile and Biological Activity of Three Cultivars of Kale. Molecules, 29(8), 1727. https://doi.org/10.3390/molecules29081727