
Saponins of Paris polyphylla for the Improvement of Acne: Anti-Inflammatory, Antibacterial, Antioxidant and Immunomodulatory Effects
 ,
,
Abstract
:
1. Introduction
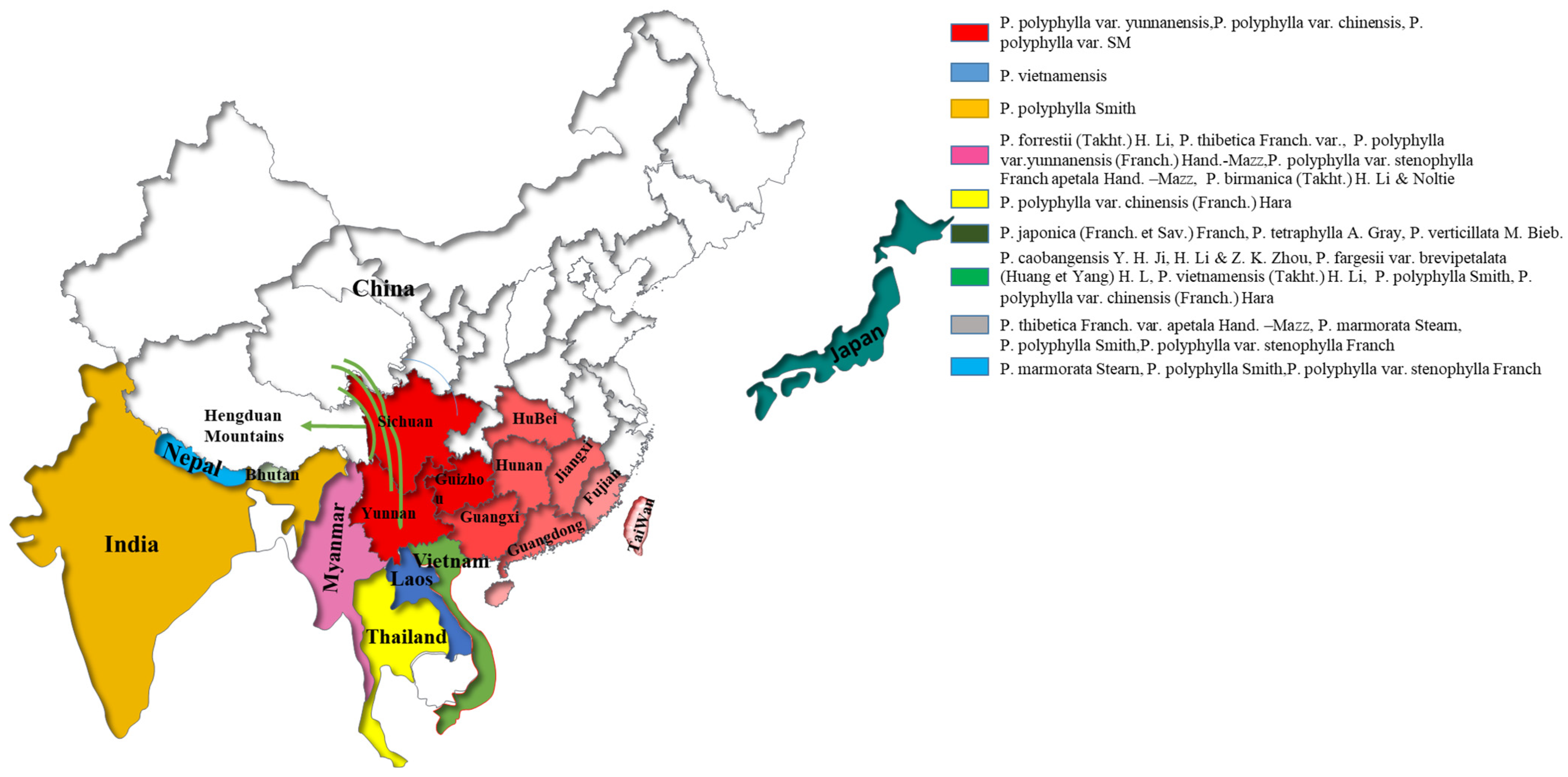
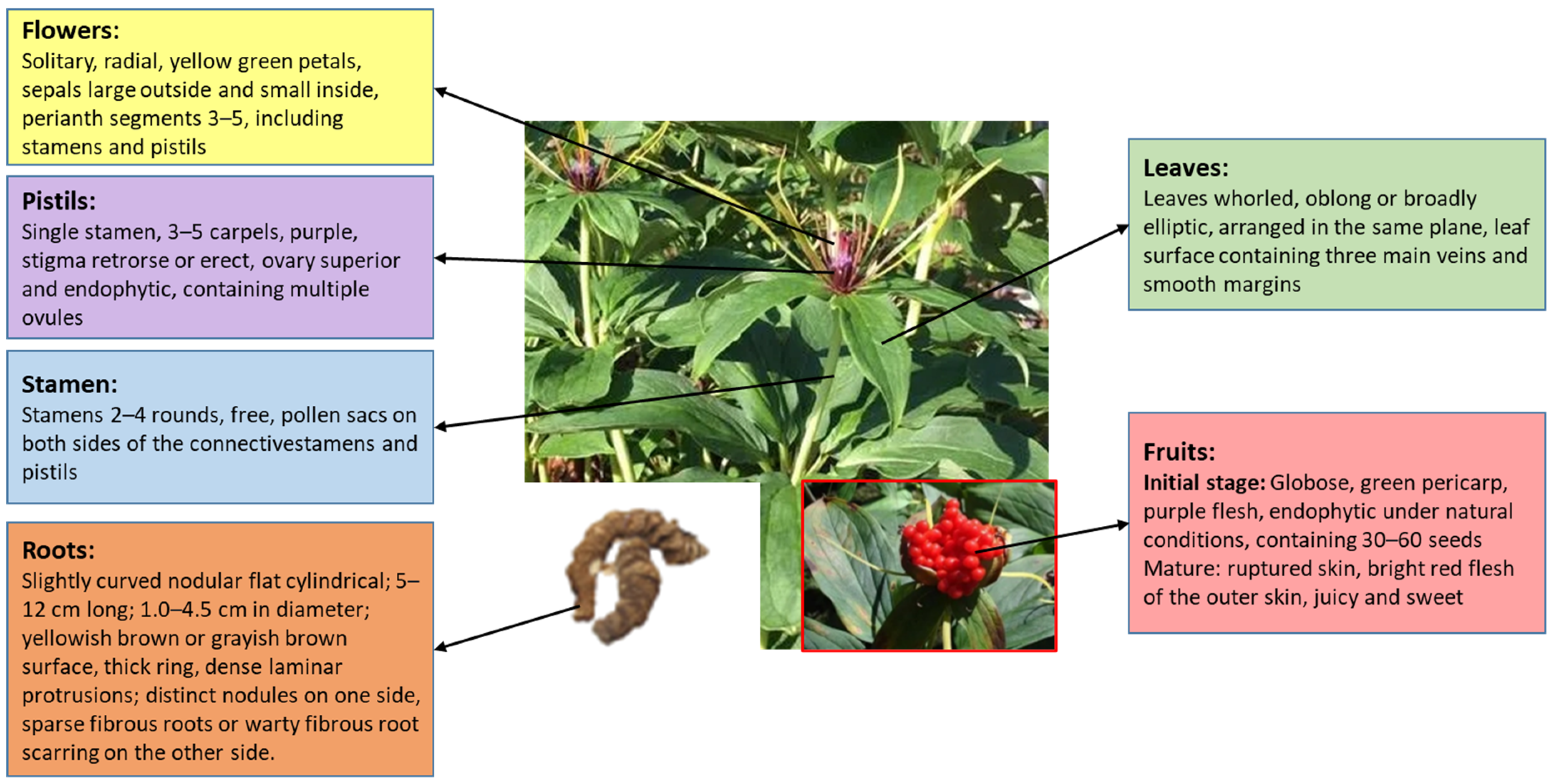
2. Distribution and Characteristics of P. polyphylla
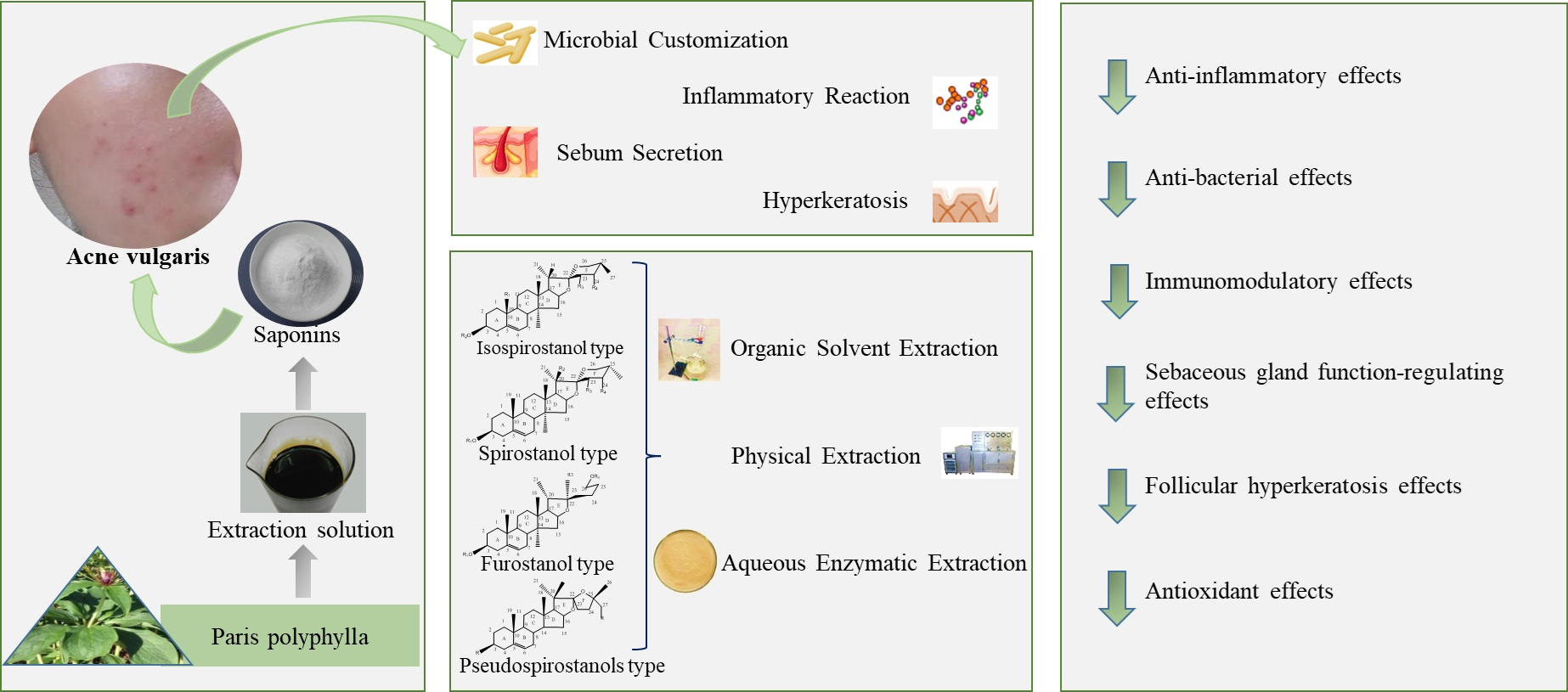
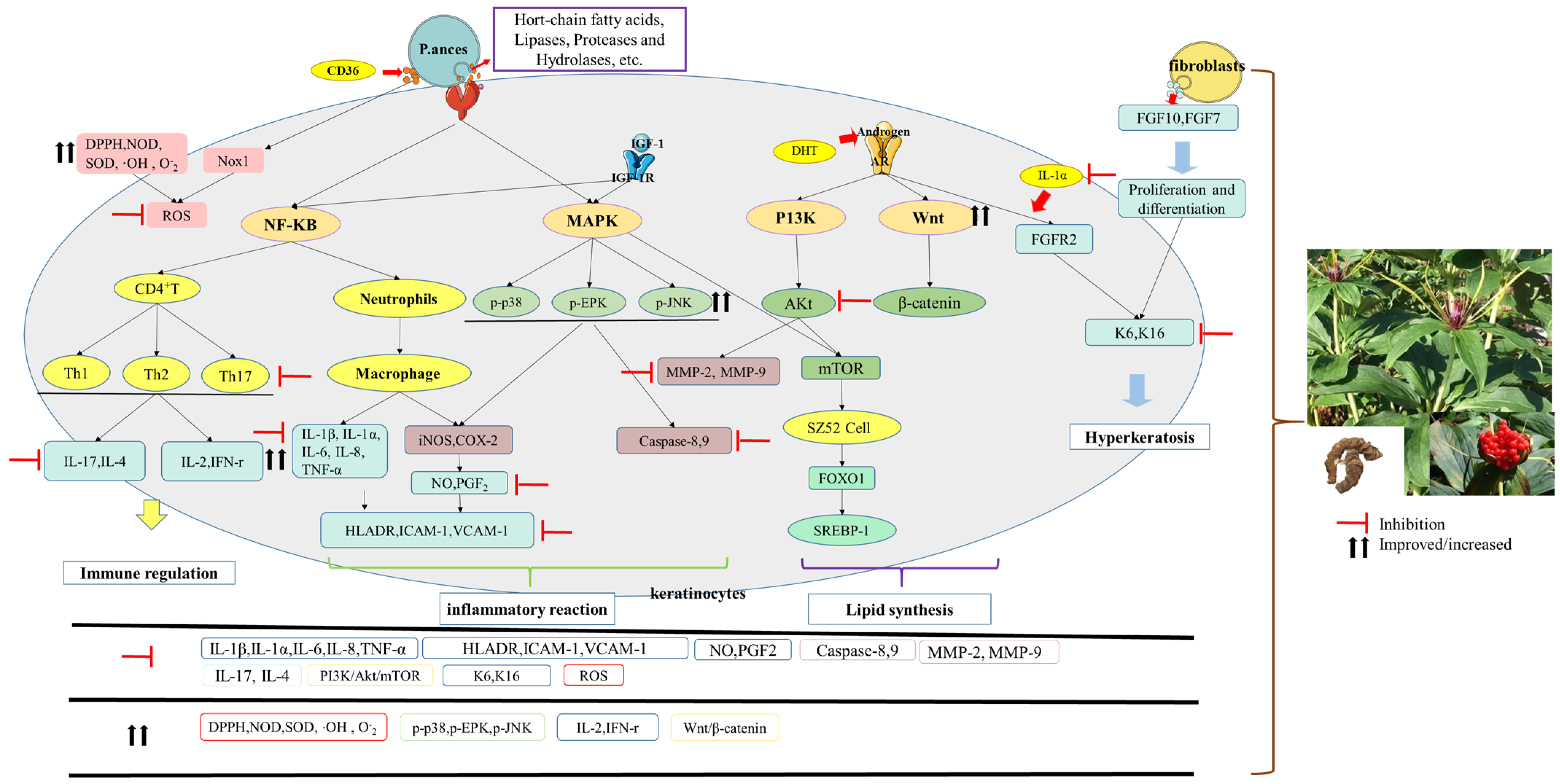
3. Pathogenesis of Acne
3.1. Microbial Customization
3.2. Inflammatory Reaction
3.3. Sebum Secretion
3.4. Hyperkeratosis
4. Bioactives of P. polyphylla Associated with Acne Treatment
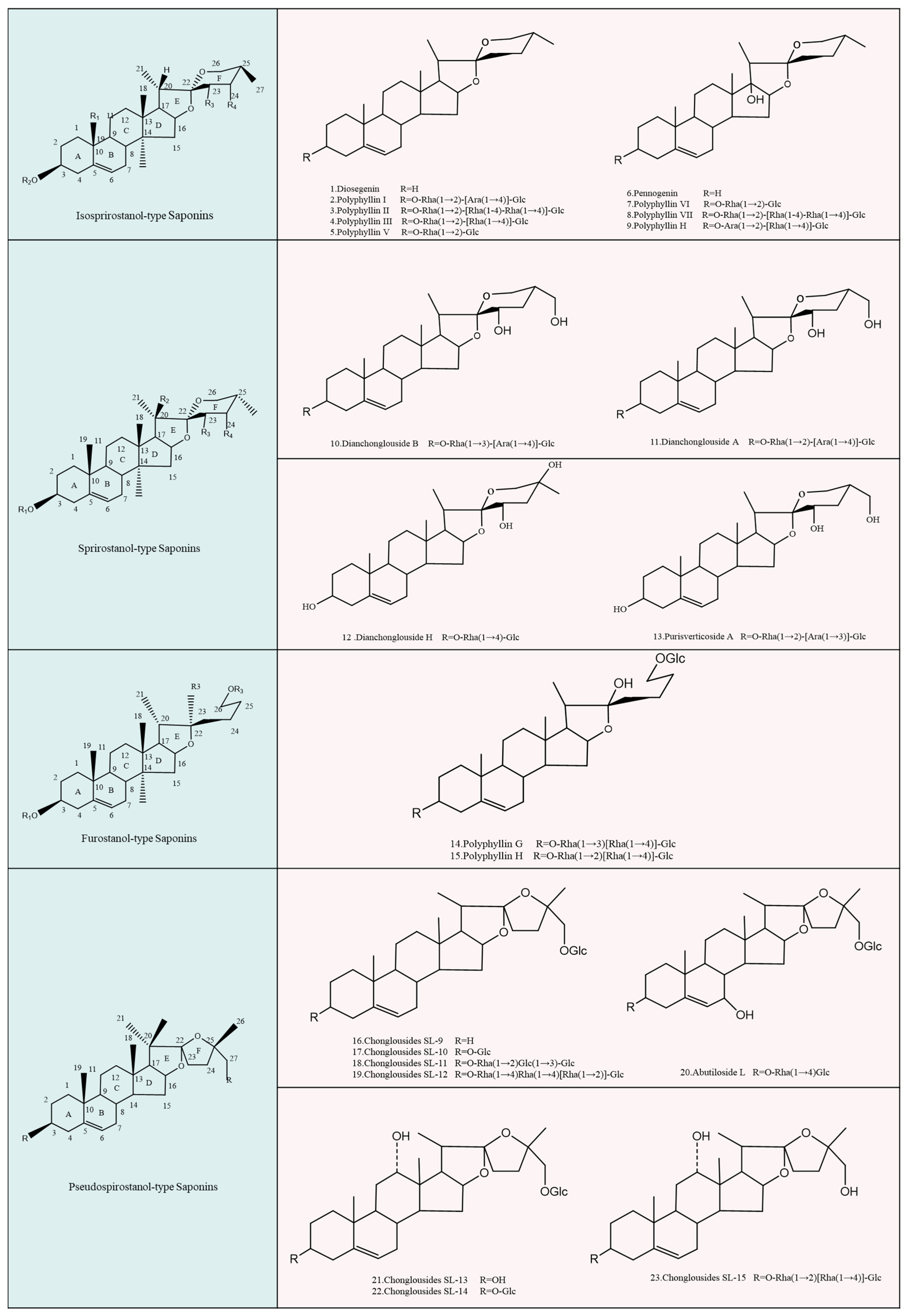
4.1. Isospirostanol Type
4.2. Spirostanol Type
4.3. Furostanol Type
4.4. Pseudospirostanol Type
5. P. polyphylla Saponins Play Roles in Acne Treatment
5.1. Anti-Inflammatory Effects
5.2. Anti-Bacterial Effects
5.3. Immunomodulatory Effects
5.4. Sebaceous Gland Function-Regulating Effects
5.5. Follicular Hyperkeratosis Effects
5.6. Antioxidant Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Substance | Research Subject | Evaluation | Cytokines | References | |
|---|---|---|---|---|---|---|
| 1 | Anti-inflammatory effects | PP I | HACAT cells induced by P. acnes | ELISA PCR/Western blot | ↓ CD36/NOX1/ROS/NLRP3/IL-1β Pathway, IL-8. | [28] |
| 2 | HACAT cells induced by P. acnes | ELISA PCR/Western blot | ↓ IL-6, IL-8, TNF-α, ↓ NF-κB activation, p38 phosphorylation, TLR2 expression. | [37] | ||
| 3 | LPS and IFN-γ induced primary bone marrow-derived macrophages (BMMs) and peritoneal elucidated macrophages (PEMs) cell model in mice | PCR/Western blot | ↓ NF-κB-mediated production of pro-inflammatory effectors in activated macrophages. | [146] | ||
| 4 | IL-17-stimulated HaCaT cell model | CCK-8/PCR | ↓ IL-17 stimulated VEGF, IL-23 content and VEGF mRNA, IL-23 mRNA expression. | [109] | ||
| 5 | PP VII | LPS-induced RAW264.7 cell model of male/zebrafish | ELISA/PCR/Western blot | ↓ NO and PGE2 production as well as pro-inflammatory cytokines (TNF-α, IL-1β and IL-6), enzymes (iNOS, COX-2), MMP-9 protein and mRNA expression. | [26] | |
| 6 | HeLa cells, A549 cells, HepG2 cells | MTT/Western blot | ↓ NF-κB/MMP-9/VEGF pathway. | [112] | ||
| 7 | Induction of apoptosis in an A549 human lung cancer cells model | phase-contrast microscopy/fluorescence microscopy/flow cytometry/Western blot analysis. | ↓ PI3K/Akt and NF-κB pathways. | [27] | ||
| 8 | PP VI | LOVO cell model of intestinal cancer induction | Western blot | ↓ MMP-2, MMP-9 expression. | [111] | |
| 9 | Rhizoma Paridis total saponins | LPS-induced murine macrophage (RAW 264.7) model | CCK8/Griess | ↓ NO release amount. | [110] | |
| 10 | Heat-inactivated Escherichia coli-induced macrophage model in rat peritoneal cavity | ELISA | ↓ TNF-α, IL-1β. | [107] | ||
| 11 | Lewis lung adenoma mouse model of induced lung adenocarcinoma cells | PCR/Western blot | ↓ VEGFD, VEGFR3, RAGE, IL6R, IL17BR and CXCL16, ↑ SOD and CAT, phosphorylation of NF-κB, PI3K/Akt, MAPK (p38, Erk1/2, JNK) signaling pathway. | [8] | ||
| 12 | Dextran induced hind paw edema in rats | MTT/fluorescence microscopy/PCR | Demonstrated potent anti-inflammatory activity by dose-dependently inhibiting dextran-induced paw edema in rats (p < 0.01) over a period of 2 h to 4 h. | [38] | ||
| 13 | Diosgenin | Osteoarthritis (OA)-induced human OA chondrocytes | ELISA | ↓ IL-1β, NO, PGE2 generation, ↓ MMP-3, MMP-13, iNOS, COX-2 expression. | [108] | |
| 14 | A molecular model of TNF-α-induced adhesion in the mouse VSMC cell line MOVAS-1 | ELISA/PCR/Western blot | ↓ VCAM-1 and ICAM-1 mRNA and content expression, ↓ ROS, ↓ p38, ERK, JNK and Akt phosphorylation, ↓ NK-κB activation. | [113] | ||
| 15 | PP II | Human ovarian cancer cell-induced angiogenesis model | MTT/EMSA/Western blot | ↓ NF-κB activity and VEGF-mediated angiogenesis. | [147] | |
| 16 | Primary liver cancer-induced HepG2 and BEL7402 cell model | ELISA/PCR/Western blot | ↓ NF-κB activity andMMP2/MMP9 mRNA and content expression. | [148] | ||
| 17 | PP H | Lipopolysaccharide (LPS)-induced RAW 264.7 and HaCaT cells | ELISA/PCR/Western blot/molecular docking/surface plasmon resonance analysis | ↑ NRF2/HO-1 antioxidant pathway, ↓ activation of the MAPK pathway, ↓ the nuclear translocation of NF-κB and downstream inflammatory genes expression. | [116] | |
| 18 | Antibacterial effects | PP G | Human oral cancer induced Gram-positive and Gram-negative bacteria | The broth microdilution method | Gram-positive and Gram-negative bacteria (MICs = 13.1–78 µg/mL). | [118] |
| 19 | Chonglouoside SL-7 | Positive control: erythromycin | The broth microdilution method | Antibacterial (MIC = 31.3, 3.9 μg/mL). | [149] | |
| 20 | Rhizoma Paridis total saponins (PP I, II, III, V, VI, VII, H) | P. acnes NCTC737, ATCC6919, S. epidermidis ATCC12228, S. aureus ATCC6538/positive control: erythromycin, clindamycin | Liquid microdilution method | Rhizoma Paridis total saponins inhibits P. acnes NCTC737 and ATCC6919, S. epidermidis ATCC12228, S. aureus ATCC6538 (MIC = 2.5, 5.0, 5.0, 1.25 mg/mL), Polyphyllin I, II, III, V, VI, VII, H inhibits P. acnes NCTC737 (MIC = 0.6, 1.2, 2.5, 5.0, 2.5, 2.5, 10.0 mg/mL), S. epidermidis ATCC12228 (MIC = 1.2, 1.2, 5.0, 5.0, 2.5, 2.5, 10.0 mg/mL), S. aureus ATCC6538 (MIC = 0.6, 0.6, 2.5, 5.0, 2.5, 2.5, 10.0 mg/mL). | [120] | |
| 21 | Rhizoma Paridis total saponins | P. acnes NCTC737, ATCC6919, S. epidermidis CMCC26069, S. aureus ATCC29213/positive control: erythromycin, clindamycin | Agar dilution method/broth microdilution method | P. acnes NCTC737 (MIC = 97.5 μg/mL, MBC = 198.0 μg/mL), P. acnes ATCC6919 (MIC = 48.7 μg/mL, MBC = 97.5 μg/mL), S. epidermidis CMCC26069 (MIC = 48.7 μg/mL, MBC = 97.5 μg/mL), S. aureus ATCC29213 (MIC = 12.2 μg/mL, MBC = 24.4 μg/mL), erythromycin inhibits P. acnes NCTC737, ATCC6919 (MIC = 0.0625 μg/mL), S. epidermidis CMCC26069 (MIC = 7.8 μg/mL), S. aureus ATCC29213 (MIC = 0.0625 μg/mL), clindamycin inhibits P. acnes NCTC737 (MIC = 0.125 μg/mL), P. acnes ATCC6919 (MIC = 0.0625 μg/mL). | [23] | |
| 22 | PP I, PP II, PP VI, PP VII | Liquid microdilution method | Polyphyllin I, II, VI, VII inhibition P. acnes (MIC = 125 μg/mL), S. epidermidis (MIC = 15.6, 15.6, 500, 31.2 μg/mL), S. aureus (MIC = 15.6, 15.6, 500, 15.6 μg/mL). | [119] | ||
| 23 | Immunoregulation Sebum secretion | Rhizoma Paridis total saponins | Mouse asthma model | ELISA/PCR | ↓ Th1/Th2, ↓ IL-4 and IFN-γ. | [150] |
| 24 | Polytrauma rat model | ELISA | ↑ TNF-ɑ, IL-1 and IL-6. | [151] | ||
| 25 | Lupus Nephritis-induced lymphocytes model | MTT/ELISA | Regulates Th1/Th2 imbalance and enhances immunosuppressive function of CD4+Treg and CD25+Treg. | [122] | ||
| 26 | Lewis lung adenoma mouse model of induced lung adenocarcinoma cells | PCR/Western blot | ↓ VEGFD, VEGFR3, RAGE, IL6R, IL17BR and CXCL16, ↑ SOD, catalase enzyme content, | [8] | ||
| 27 | Diosgenin | Multiple sclerosis-induced microglia and macrophages modellerosis | Fluorescence microscopy | ↓ microglia and macrophages activation, ↓ CD4+T cell proliferation, ↓ Th1/Th17 cell differentiation. | [114] | |
| 28 | PP I | ELISA/PCR/Western blot | ↓ HIF-1α, VEGF. | [109] | ||
| 29 | Follicular hyperkeratosis effects | PP I | Acne caused by P. acnes | PCR/Western blot | ↓ IL 1α and K16 expression levels in HaCaT keratin cells. | [37] |
| 30 | Anti-oxidation | Rhizoma Paridis total saponins | Ascorbic acid | MTT/AO/PI staining/fluorescence microscopy/PCR | ↑DPPH, NOD, SOD, RP. | [38] |
| 31 | PP I | HacaT photoaging model caused by UV light | ELISA/WB/DCFH-DA | ↑ SIRT3, SOD2, ↓ p53acetylation levels, Bax, cleaved caspase3, ROS. | [143] | |
| 32 | PP II | Glomerular mesangial cells under high glucose intervention (GMC) | MTT/DCFH-DA/xanthine oxidase assay/malondialdehyde kit | ↓ MDA, ROS, ↑ SOD. | [114] | |
| 33 | Rhizoma Paridis total saponins | Lipid peroxidation model induced by light riboflavin and the Fenton reaction | Spectrophotometric methods | Scavenging ·OH, O-2 radicals, ↓ lipid peroxidation and ·OH induced oxidative DNA damage. | [140] |
6. Extraction Methods of P. polyphylla Saponins
| No. | Extraction Method | Extraction Components | Extraction Conditions | Detection Method | Optimal Process Conditions | Rate | References |
|---|---|---|---|---|---|---|---|
| 1 | RE | Diosgenin | Extraction solvent: 75~95% ethanol, solvent dosage: 4~8 mL, extraction time: (0.5, 0.5)–(1.5, 1.5) h, extraction times: 2. | HPLC/orthogonal experimental method | The crude powder of Paris herb was refluxed twice with 8 times the amount of 85% ethanol, 1.5 h/time each. The order of effect was as follows: reflux time > ethanol concentration > ethanol dosage | The average content of diosgenin element was 6.0821 mg/g. | [152] |
| 2 | RE | Rhizoma Paridis total saponins/ PP I/PP II | Extraction solvent: water, 30–90% ethanol, particle size of herbs: drinking tablets, coarsest powder, coarse powder, solvent dosage (first time, second time): 1:(6, 4)–1:(10, 8) (g:mL), extraction time: (1, 0.5)–(2, 1.5) h, extraction times:1~3. | HPLC method | 70% ethanol was extracted twice, the first time with 10 times the amount for 2 h, the second time with 8 times the amount for 1.5 h. The order of effect size was as follows: reflux time > ethanol dosage > ethanol concentration. The effect of the crushing degree of herbs on the total saponin yield was small. | The Rhizoma Paridis total saponins yield was 4.24%, RSD: 4.5%, and the total extraction rate of PP I and PP II was 93.28%, RSD: 1.20%. | [153] |
| 3 | RE | PP VII/ PP H/ PP VI/ PP I/PP II | 50 kg of heavy drug, plus 95% ethanol extraction 3 times, control the temperature between 70–80 °C, extraction time: 3–5 h, parallel extraction 3 times. | HPLC/orthogonal experimental method | The saponin content was PP VII: 90.86 mg/g, PP H: 198.02 mg/g, PP VI: 302.57 mg/g, PP I: 27.22 mg/g, PP II: 137.18 mg/g. | [154] | |
| 4 | RE | PP I | Extraction medium: distilled water, 40~80% ethanol and anhydrous ethanol, solvent dosage: 8–20 times, extraction times: 1–3, extraction time: 1–3 h/time. | HPLC method | 90% ethanol, 1:12 ratio, 1 h each time, 2 extractions. The order of effect size was as follows: number of extractions > extraction time > ratio > extraction solvent | PP I yield of 10.27%. | [155] |
| 5 | RE | PP I | Extraction medium: 55–95% ethanol solution, extraction temperature: 50–90 °C, extraction time: 1–3 h, herb particle size: powder, medium powder, coarse powder. | Colorimetric/Orthogonal experimental method | The ethanol concentration was 75%, the extraction temperature was 90 °C, the extraction time was 1.5 h, and the order of influence was as follows: extraction temperature > solvent concentration > extraction time | The total saponin yield was 12.74%, RSD: 1.90%. | [156] |
| 6 | RE | Rhizoma Paridis total saponins/ PP I/PP II | Extraction solvents: 30–100% methanol, soaking time before extraction: 30–150 min, liquid to material ratio: 10:1~60:1 (mL:g), extraction time: 30–90 min/time, extraction times: 1–4. | HPLC/orthogonal experimental/Response Surface method | The extraction solvent was 70% ethanol, the ratio of liquid to material was 30:1 (mL:g), and the extraction time was 60 min/times after continuous reflux extraction. The order of influence was as follows: extraction time > extraction times > extraction solvent > material to liquid ratio. | The extraction rate was 2.094% | [157] |
| 7 | RE | Rhizoma Paridis total saponins/ PP I/PP II/PP VI/PP VII | Extraction solvents: double-distilled water, 50~95% ethanol and anhydrous ethanol, Solvent dosage: 1:8–1:14 (g:mL), extraction time: 1–3 h, extraction times: 1–3 times. | HPLC/orthogonal experimental method | The extract was refluxed in a water bath with 75% ethanol for 1.5 h at a ratio of 1:12 (B:V) and extracted twice. The order of effect: Extraction times: > Extraction time > Material to liquid ratio > Extraction solvent | The total saponin yield was 10.33%, and PP I, II, VI, VII RSD were 1.3%, 0.70%, 0.94% and 0.58%. | [158] |
| 8 | USA-SE | Rhizoma Paridis total saponins | Material-liquid ratio: 1:30–1:110 (g:mL) add water-saturated n-butanol, placed in the ultrasonic cleaner, extraction temperature: 35–75 °C ultrasonic extraction for 30 min; ultrasonic power: 50–100%. | Colorimetric/L16(45) orthogonal experimental method | The crude powder (40 mesh) was soaked in 30% ethanol solution for 36 h and then sonicated for 30 min; the volume ratio of crude powder to (soaked) ethanol was 1:15. Influence size order was as follows: ultrasonic measurement > soaking time > extraction solvent > material to liquid ratio | The extraction rate of Rhizoma Paridis total saponins was 9.50%. | [159] |
| 9 | USA-SE | PP I/PP II/PP VI/PP VII | Methanol single material-liquid ratio: 1:5–1:15 (g:mL), single extraction time: 10~30 min, extraction temperature: 30–50 °C, extraction times: 1–~3. | HPLC/L9(34) orthogonal experimental method | The extraction was carried out for 20 min at an initial extraction temperature of 40 °C with a single material to liquid ratio of 15 times of methanol and the number of extractions was 3. The order of effect size was as follows: extraction number > initial extraction temperature > material to liquid ratio > single extraction time | The average recoveries of PP I, II/VI/VII were 101.04%, 101.73%, 98.69% and 101.17% with RSDs of 1.33%, 2.34%, 1.64%, and 2.17%, respectively. | [160] |
| 10 | USA-SE | Rhizoma Paridis total saponins | Solvent selection: 40–95% ethano, methanol, acetone, dosage selection: 1:10–1:30 (g:mL) of 75% ethanol, ultrasonic heating time: 20–60 min, ultrasonic temperature: 40~80 °C. | HPLC/Orthogonal experimental method | The extraction solvent was 75% ethanol, 15 times the amount of solvent, the extraction temperature was 70 °C, and the extraction time was 50 min. The order of effect size was as follows: extraction solvent > extraction time > extraction temperature > solvent times | The amount of extract was 0.2651 g, and the Rhizoma Paridis total saponins content was 0.2259%. | [161] |
| 11 | USA-SE | Rhizoma Paridis total saponins | Ethanol concentration: 60–80%, ethanol dosage: 1:30–1:70 (g:mL), extraction time: 15~60 min. | HPLC/L9 (34) Orthogonal experimental method | The extraction was performed by ultrasonication with 60% ethanol at a ratio of 1:50 material to liquid for 30 min. The order of effect size was as follows: solvent times > extraction solvent > extraction time | The total saponin yield was 4.387%. | [162] |
| 12 | Dip method | PP I/PP II | \ | HPLC/Orthogonal experimental method | Take 50 g of the slices of Paris, macerate with 95% ethanol for 24 h with shaking, filter through, and fix the filtrate to 500 mL with 95% ethanol. | The total amount of PP I/PP IIwas 0.356%. | [163] |
| 13 | USA-SE | \ | HPLC/Orthogonal experimental method | Take 50 g of the slices of Paris, sonicate it with 95% ethanol at 500 mL (power 250 W, frequency 33 kHz) for 30 min, let it cool, filter it, and fix the filtrate to 500 mL with 95% ethanol. | The total amount of PP I/PP II was 1.287%. | ||
| 14 | RE | \ | HPLC/Orthogonal experimental method | Take 50 g of the slices of Paris, add 250 mL of 95% ethanol, and reflux twice, 2 h each time. Combine the extracts, filter, and dilute the filtrate to 500 mL with 95% ethanol. There was a significant difference in the ethanol volume fraction factor | The total amount of PP I/PP II was 1.192%, RSD was 2.24%. | ||
| 15 | MWA-SE | Rhizoma Paridis total saponins | Solvent selection: 45–90% ethanol, extraction temperature: 60–80 °C, solvent dosage: 1:10–1:20 (g:mL). | Colorimetric/L9(34) Orthogonal experimental method | The extraction was carried out at 80 °C for 20 min with 10 times the amount of 60% ethanol solution. The order of effect size was as follows: ethanol concentration > extraction temperature > extraction time > ethanol dosage. | The total saponin yield was 11.6% and the RSD was 0.3% | [164] |
| 16 | MWA-SE | PP I/PP VII | Extraction solvent: 0~100% ethanol concentration, microwave power: 300~500 w, liquid-solid ratio: 1:10–1:30 (g:mL), radiation time: 5–15 min, extraction temperature: 30–50 °C. | HPLC/LC-ESI-MS/H NMR Spectroscopic | 70% v/v ethanol aqueous solution concentration, 300 W microwave power, 20:1 mL/g liquid–solid ratio, 50 °C extraction temperature, and 15 min irradiation time. The extraction solvent was the decisive factor, while microwave power and irradiation time were important factors in MAE. | The extraction yields of PP VII and PP I were 5.66 and 15.4 mg/g. | [165] |
| 17 | Room-temperature UPE | Saponin A, saponin B, saponin C, saponin D | Extraction solvent: water, methanol, 95% ethanol, water-saturated n-butanol, ethanol concentration: 30–95%, UHP pressure: 100–600 Mpa, extraction time: 1–5 min. | HPLC/L9 (34) Orthogonal experimental method | The ethanol concentration was 90%, the extraction pressure was 400 Mpa, the extraction time was 2 min, and the liquid–solid ratio was 40:1. | The rates of saponin A, saponin B, saponin C, and saponin D were 1.164%, 0.591%, 0.043%, and 0.053% respectively. | [166] |
| 18 | UHPA-SE | \ | HPLC method | Treat with 40 mL of 90% ethanol in water (v/v), fix the transducer, and sonicate the system in an ultrasonic bath (frequency, 50 Hz; power, 250 W) for 30 min. | The rates of saponin A, saponin B, saponin C, and Saponin D were 1.113%, 0.513%, 0.038%, and 0.045% respectively. | ||
| 19 | Microwave-assisted extraction (MAE). | \ | HPLC method | Using a 40 mL 90% ethanol aqueous solution (v/v) treatment, the system was exposed to a microwave radiation source operating at 300 W and 2450 MHz frequency and subjected to atmospheric pretreatment for 15 min of extraction. | The rates of saponin A, saponin B, saponin C, and saponin D were 1.139%, 0.539%, 0.041%, and 0.049%, respectively. | ||
| 20 | Method of Sohxlet extraction. | \ | HPLC method | Boil the mixture for 3 h with 80 mL of 90% ethanol in water (v/v), | The rates of saponin A, saponin B, saponin C, and saponin D were 1.119%, 0.511%, 0.039%, and 0.044%, respectively. | ||
| 21 | Usual room-temperature extraction (RTE) | \ | HPLC method | Extraction with 40 mL of 90% ethanol in water (v/v), 25 °C room-temperature for 5 days. | The rates of saponin A, saponin B, saponin C, and saponin D were 0.988%, 0.483%, 0.034%, and 0.040%, respectively. | ||
| 22 | SFE-CO2 | Pennogenin | Extraction method: dynamic, static, dynamic + static, entrainment agent: methanol, acetone, ethanol, ethyl acetate, extraction temperature: 5–65 °C, extraction pressure: 150–400 bar, extractant dosage: 100–600 mL, extraction time: 2–7 h, SC-CO2 flow rate: 15–60 g/min. | HPLC/L9 (34) Orthogonal experimental method | The extraction was carried out by a combination of dynamic and static methods with the optimal ratio of ethanol 90%–ethyl acetate 10%, an extraction temperature of 52 °C, an extraction pressure of 315 bar, a SC-CO2 flow rate of 39 g/min, an extractant dosage of 300 mL and an extraction time of 4 h. | The yield of Pennogenin was 16.32%. | [167] |
| 23 | AEE (ultrasound-assisted extraction) | Rhizoma Paridis total saponins | Enzyme dosage:10–50 U/g substrate, enzyme digestion temperature: 30–70 °C, PH: 3.5–5.5, enzyme digestion time: 30–150 min. | Colorimetric/L9(34) Orthogonal experimental/Response Surface method | The amount of cellulase was 32 U/g of substrate, the enzymatic digestion temperature was 52 °C, the enzymatic pH was 4.6, and the enzymatic digestion time was 92 min. | The theoretical value of the extraction rate of Rhizoma Paridis total saponins was 1.66%. | [168] |
| 24 | AEE (water extraction) | PP I | Extraction time: 1–3 h, extraction times: 1–3, extraction temperature: 60–95 °C, solvent dosage: 1:5–1:8 (g:mL), PH value: 5–9, drug particle size: 10–40. | Colorimetric/L9(34) Orthogonal experimental method | Enzyme A and enzyme B were used alternately, where the dosage of enzyme A was 1 mL/kg of raw drug and the dosage of enzyme B was 2 mL/kg of raw drug, crushed into 20 items of herb with 7 times the amount of solvent pH 7 extracted 3 times at 60 °C for 3 h each. | The extraction rate was increased by 24.28% to 61.27%. | [169] |
7. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Oulès, B.; Philippeos, C.; Segal, J.; Tihy, M.; Vietri Rudan, M.; Cujba, A.; Grange, P.A.; Quist, S.; Natsuga, K.; Deschamps, L. Contribution of GATA6 to homeostasis of the human upper pilosebaceous unit and acne pathogenesis. Nat. Commun. 2020, 11, 5067. [Google Scholar] [CrossRef]
- Min, K.J.; Ha, S.; Son, J.A.; Ji, H.S.; Houh, Y.; Cho, E.; Ji, H.C.; Yoon, S.R.; Yang, Y.; Bang, S.I. Polyphenon-60 displays a therapeutic effect on acne by suppression of TLR2 and IL-8 expression via down-regulating the ERK1/2 pathway. Arch. Dermatol. Res. 2012, 304, 655–663. [Google Scholar]
- Ruan, S.F.; Xiang, S.J.; Wu, W.F.; Cao, S.W.; Liu, Q. Potential role of mTORC1 and the PI3K-Akt pathway in anti-acne properties of licorice flavonoids. J. Funct. Foods 2020, 70, 103968. [Google Scholar] [CrossRef]
- Gabsik, Y.; Hye, E.L.; Sang, H.Y.; Han, C. Licochalcone A attenuates acne symptoms mediated by suppression of NLRP3 inflammasome. Phytother. Res. 2018, 32, 2551–2559. [Google Scholar]
- Weber, N.; Biehler, K.; Schwabe, K.; Haarhaus, B.; Quirin, K.W.; Frank, U.; Schempp, C.; Wölfle, U. Hop Extract Acts as an Antioxidant with Antimicrobial Effects against Propionibacterium acnes and Staphylococcus aureus. Molecules 2019, 24, 223. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.Y.; Kim, K. Anti-Propionibacterium acnes and the anti-inflammatory effect of Aloe ferox miller components. J. Herb. Med. 2017, 9, 53–59. [Google Scholar] [CrossRef]
- Kwon, H.C.; Kim, T.Y.; Lee, C.M.; Lee, K.S.; Lee, K.K. Active compound chrysophanol of Cassia tora seeds suppresses heat-induced lipogenesis via inactivation of JNK/p38 MAPK signaling in human sebocytes. Lipids Health Dis. 2019, 18, 135. [Google Scholar] [CrossRef]
- Man, S.L.; Chai, H.Y.; Cui, J.X.; Yao, J.W.; Ma, L.; Gao, W.Y. Antitumor and anti-metastatic mechanisms of Rhizoma paridis saponins in Lewis mice. Environ. Toxicol. 2018, 33, 149–155. [Google Scholar] [CrossRef]
- Qin, X.J.; Sun, D.J.; Ni, W.; Chen, C.X.; Hua, Y.; He, L.; Liu, H.Y. Steroidal saponins with antimicrobial activity from stems and leaves of Paris polyphylla var. yunnanensis. Steroids 2012, 77, 1242–1248. [Google Scholar] [CrossRef]
- Yang, L.; Shou, Y.; Yang, Y.; Xu, J. Elucidating the immune infiltration in acne and its comparison with rosacea by integrated bioinformatics analysis. PLoS ONE 2021, 16, e0248650. [Google Scholar] [CrossRef]
- Lan, M. Dian Nan Ben Cao; Yunnan People’s Publishing House: Yunnan, China, 2017; p. 977. [Google Scholar]
- Liu, X.; Li, Z. Agriculture God’s Canon of Materal Medica; Shanghai Sanlian Bookstore Co., Ltd.: Shanghai, China, 2017; p. 1252. [Google Scholar]
- Li, S. Compendium of Materal Medica; Huaxia Publishing House: Beijing, China, 2013; p. 2000. [Google Scholar]
- China Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume I, p. 1809. [Google Scholar]
- Puwein, A.; Thomas, S.C. An Overview of Paris polyphylla, a Highly Vulnerable Medicinal Herb of Eastern Himalayan Region for Sustainable Exploitation. J. Nat. Prod. 2020, 10, 3–14. [Google Scholar] [CrossRef]
- He, L.; Shi, W.; Liu, X.; Zhao, X.; Zhang, Z. Anticancer action and mechanism of ergosterol peroxide from Paecilomyces cicadae fermentation broth. Int. J. Mol. Sci. 2018, 19, 3935. [Google Scholar] [CrossRef] [PubMed]
- Malla, B.; Gauchan, D.P.; Chhetri, R.B. An ethnobotanical study of medicinal plants used by ethnic people in Parbat district of western Nepal. J. Ethnopharmacol. 2015, 165, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Gao, W. The clinic study of Chong, Lou, Ji, Cai, Sheng, Hua, Tang in promoting uterine recovery after abortion surgery. Contemp. Med. 2014, 20, 148–150. [Google Scholar]
- Guo, T.; Zhou, Z.; Huang, Z.; Wen, H.; Huang, X. Effect of Lianxiuwugong Dilong Tang on the immune function of children with Mycoplasma pneumonia. New Chin. Med. 2007, 19–21. [Google Scholar]
- Su, A.; Duan, H. Qingwenjiedutonglin soup treated with 196 cases of gonorrhea. Mod. Chin. Med. 2000, 30–31. [Google Scholar]
- Zhou, L.; Song, H.; Zhang, Y.; Ren, Z.; Li, M.; Fu, Q. Polyphyllin VII attenuated RANKL-induced osteoclast differentiation via inhibiting of TRAF6/c-Src/PI3K pathway and ROS production. BMC Musculoskel. Dis. 2020, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yi, T.; Lin, K. In Vitro Activity of Paris polyphylla Smith Against Enterovirus 71 and Coxsackievirus B3 and Its Immune Modulation. Am. J. Chin Med. 2011, 39, 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Tu, Y.; He, L. Antimicrobial activity of ethanol extract from Dianchonglou on acne-causing bacteria. Dermatol. Vener. 2013, 35, 67–69. [Google Scholar]
- Deng, D.; Lauren, D.R.; Cooney, J.M.; Jensen, D.J.; Wurms, K.V.; Upritchard, J.E.; Cannon, R.D.; Wang, M.Z.; Li, M.Z. Antifungal Saponins from Paris polyphylla Smith. Planta Med. 2008, 74, 1397–1402. [Google Scholar] [CrossRef]
- Sun, C.; Ni, W.; Yan, H.; Liu, Z.; Yang, L.; Si, Y.; Hua, Y.; Chen, C.; He, L.; Zhao, J.; et al. Steroidal saponins with induced platelet aggregation activity from the aerial parts of Paris verticillata. Steroids 2014, 92, 90–95. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.Y.; Jia, X.J.; Wang, K.; Tu, Y.B.; Wang, R.C.; Liu, K.C.; Lu, T.; He, C.W. In Vitro and In Vivo Anti-Inflammatory Effects of Polyphyllin VII through Downregulating MAPK and NF-κB Pathways. Molecules 2019, 24, 875. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xu, C.; Zheng, L.; Wang, K.; Jin, M.; Sun, Y.; Yue, Z. Polyphyllin VII induces apoptotic cell death via inhibition of the PI3K/Akt and NF-κB pathways in A549 human lung cancer cells. Mol. Med. Rep. 2020, 21, 597–606. [Google Scholar] [PubMed]
- Yang, S.; Jiang, Y.; Yu, X.; Zhu, L.; Wang, L.; Mao, J.; Wang, M.; Zhou, N.; Yang, Z.; Liu, Y.; et al. Polyphyllin I Inhibits Propionibacterium acnes-Induced IL-8 Secretion in HaCaT Cells by Downregulating the CD36/NOX1/ROS/NLRP3/IL-1β Pathway. Evid.-Based Complement. Altern. Med. 2021, 2021, 1821220. [Google Scholar] [CrossRef]
- Ichiro, K.; Layton, L.M.; Rei, O. Updated Treatment for Acne: Targeted Therapy Based on Pathogenesis. Dermatol. Ther. 2021, 11, 1129–1139. [Google Scholar]
- Unkles, S.E.; Gemmell, C.G. Effect of clindamycin, erythromycin, lincomycin, and tetracycline on growth and extracellular lipase production by propionibacteria in vitro. Antimicrob. Agents Chemother. 1982, 21, 39–43. [Google Scholar] [CrossRef]
- Batra, R.; Sadhasivam, S.; Saini, S.; Gupta, S.; Bisen, R.; Sinha, M.; Ghosh, S.; Jain, S. Efficacy and Safety of VB-1953 Topical Gel in Non-Responder Acne Patients with Clindamycin-Resistant Cutibacterium acnes. Drugs R D 2020, 20, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Ceruelos, A.; Romero-Quezada, L.C.; Ruvalcaba Ledezma, J.C.; Lopez Contreras, L. Therapeutic uses of metronidazole and its side effects: An update. Eur. Rev. Med. Pharmacol. 2019, 23, 397–401. [Google Scholar]
- Lowe, L.; Herbert, A.A. Cystic and comedonal acne: A side effect of etretinate therapy. Int. J. Dermatol. 1989, 28, 482. [Google Scholar] [CrossRef]
- Plewig, G.; Schopf, E. Anti-inflammatory effects of antimicrobial agents: An in vivo study. J. Investig. Dermatol. 1975, 65, 532–536. [Google Scholar] [CrossRef]
- Shen, S.; Chen, D.; Li, X.; Li, T.; Yuan, M.; Zhou, Y.; Ding, C. Optimization of extraction process and antioxidant activity of polysaccharides from leaves of Paris polyphylla. Carbohyd. Polym. 2014, 104, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chu, J.; Zhou, Y.; Di, W.; Li, X.; Peng, X.; Li, C.; Ju, R. Study on Extraction Technology and Antioxidant Effect of Flavonoids from Chonglou (Petiolate paris). Guid. J. Tradit. Chin Med. Phar. 2022, 28, 24–29. [Google Scholar]
- Zhu, T.; Wu, W.; Yang, S.; Li, D.; Sun, D.; He, L. Polyphyllin I Inhibits Propionibacterium acnes-Induced Inflammation In Vitro. Inflammation 2019, 42, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, D.; Mishra, S.; Verma, S.S.; Shekher, A.; Rai, V.; Awasthee, N.; Das, T.J.; Paul, D.; Das, S.K.; Tag, H.; et al. Evaluation of antioxidant, anti-inflammatory and anticancer activities of diosgenin enriched Paris polyphylla rhizome extract of Indian Himalayan landraces. J. Ethnopharmacol. 2021, 270, 113842. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.G.; Zhao, Y.L.; Zhang, J.; Zuo, Z.T.; Zhang, Q.Z.; Wang, Y.Z. The traditional uses, phytochem, and pharmacological properties of Paris L. (Liliaceae): A review. J. Ethnopharmacol. 2021, 278, 114293. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Wang, K.; Gao, X.; Yan, B. Research Progress on Chemical Constituents and Pharmacological Activities of Menispermi rhizoma. Molecules 2023, 28, 2701. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Ding, C.; Yang, R.; Zhang, L.; Zhou, Y.; Li, Y. Karyomorphology of some taxa of Paris (Melanthiaceae) from Sichuan province, China. Caryologia 2011, 64, 288–296. [Google Scholar]
- Shah, A.S.; Mazumder, P.B.; Choudhury, M.D. Medicinal properties of Paris polyphylla smith: A review. J. Herb. Med. Toxicol. 2012, 1, 27–33. [Google Scholar]
- Mohd, T.; Belwal, T.; Bhatt, I.D.; Pande, V.; Nandi, S.K. Polyphenolics in leaves of Paris polyphylla: An important high value Himalayan medicinal herb. Ind. Crop. Prod. 2018, 117, 66–74. [Google Scholar] [CrossRef]
- Tariq, M.; Nandi, S.K.; Bhatt, I.D.; Bhavsar, D.; Roy, A.; Pande, V. Phytosociological and niche distribution study of Paris polyphylla smith, an important medicinal herb of Indian Himalayan region. Trop. Ecol. 2021, 62, 163–173. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Shen, T.; Zhang, J. Biomass allocation and allometry of Paris polyphylla var. yunnanensis with different ages. Chin. Tradit. Herb. Drugs 2022, 53, 3456–3462. [Google Scholar]
- Wu, Z.; Zhang, J.; Xu, F.; Wang, Y.; Zhang, J. Rapid and simple determination of polyphyllin I, II, VI, and VII in different harvest times of cultivated Paris polyphylla Smith var. yunnanensis (Franch.) Hand.-Mazz by UPLC-MS/MS and FT-IR. J. Nat. Med. 2017, 71, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Dreno, B.; Dagnelie, M.A.; Khammari, A.; Corvec, S. The Skin Microbiome: A New Actor in Inflammatory Acne. Am. J. Clin. Dermatol. 2020, 21, 18–24. [Google Scholar] [CrossRef]
- Huang, T.Y.; Jiang, Y.E.; Scott, D.A. Culturable bacteria in the entire acne lesion and short-chain fatty acid metabolites of Cutibacterium acnes and Staphylococcus epidermidis isolates. Biochem. Biophys. Res. Commun. 2022, 622, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Gannesen, A.V.; Zdorovenko, E.L.; Botchkova, E.A.; Hardouin, J.; Massier, S.; Kopitsyn, D.S.; Gorbachevskii, M.V.; Kadykova, A.A.; Shashkov, A.S.; Zhurina, M.V.; et al. Composition of the Biofilm Matrix of Cutibacterium acnes Acneic Strain RT5. Front. Microbiol. 2019, 10, 1284. [Google Scholar] [CrossRef] [PubMed]
- Akaza, N.; Akamatsu, H.; Numata, S.; Matsusue, M.; Mashima, Y.; Miyawaki, M.; Yamada, S.; Yagami, A.; Nakata, S.; Matsunaga, K. Fatty acid compositions of triglycerides and free fatty acids in sebum depend on amount of triglycerides, and do not differ in presence or absence of acne vulgaris. J. Dermatol. 2014, 41, 1069–1076. [Google Scholar] [CrossRef]
- Li, Y.H.; Sun, Y.; Tang, T.L.; Niu, Y.B.; Li, X.Q.; Xie, M.; Jin, H.C.; Mei, Q.B. Paris saponin VII reverses chemoresistance in breast MCF-7/ADR cells. J. Ethnopharmacol. 2019, 232, 47–54. [Google Scholar] [CrossRef]
- Xia, P.P.; Wu, Y.P.; Lian, S.Q.; Yan, L.; Meng, X.; Duan, Q.D.; Zhu, G.Q. Research progress on Toll-like receptor signal transduction and its roles in antimicrobial immune responses. Appl. Microbiol. Biotechnol. 2021, 105, 5341–5355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; The, E.; Nedumaran, B.; Ao, L.; Jarrett, M.J.; Xu, D.; Fullerton, D.A.; Meng, X. Monocytes enhance the inflammatory response to TLR2 stimulation in aortic valve interstitial cells through paracrine up-regulation of TLR2 level. Int. J. Biol. Sci. 2020, 16, 3062–3074. [Google Scholar] [CrossRef]
- Jang, M.; Hwang, I.; Hwang, B.; Kim, G. Anti-inflammatory effect of Antirrhinum majus extract in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Sci. Nutr. 2020, 8, 5063–5070. [Google Scholar] [CrossRef]
- Lee, J.W.; Kang, Y.J.; Choi, H.K.; Yoon, Y.G. Fractionated Coptis chinensis Extract and Its Bioactive Component Suppress Propionibacterium acnes-Stimulated Inflammation in Human Keratinocytes. J. Microbiol. Biotechnol. 2018, 28, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Lee, M.Y. The ameliorative effect of hemp seed hexane extracts on the Propionibacterium acnes-induced inflammation and lipogenesis in sebocytes. PLoS ONE 2018, 13, e0202933. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.H.; Lee, W.R.; Wang, P.H.; Cheng, K.T.; Chen, Y.C.; Shen, S.C. Propionibacterium acnes-induced iNOS and COX-2 protein expression via ROS-dependent NF-κB and AP-1 activation in macrophages. J. Dermatol. Sci. 2013, 69, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Jeremy, A.; Holland, D.B.; Roberts, S.G.; Thomson, K.F.; Cunliffe, W.J. Inflammatory events are involved in acne lesion initiation. J. Investig. Dermatol. 2003, 121, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Coenye, T.; He, L.; Kabashima, K.; Kobayashi, T.; Niemann, C.; Nomura, T.; Olah, A.; Picardo, M.; Quist, S.R.; et al. Sebaceous immunobiology-skin homeostasis, pathophysiology, coordination of innate immunity and inflammatory response and disease associations. Fron. Immunol. 2022, 13, 1029818. [Google Scholar] [CrossRef] [PubMed]
- Kistowska, M.; Meier, B.; Proust, T.; Feldmeyer, L.; Cozzio, A.; Kuendig, T.; Contassot, E.; French, L.E. Propionibacterium acnes Promotes Th17 and Th17/Th1 Responses in Acne Patients. J. Investig. Dermatol. 2015, 135, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Agak, G.W.; Qin, M.; Nobe, J.; Kim, M.; Krutzik, S.R.; Tristan, G.R.; Elashoff, D.; Garbán, H.J.; Kim, J. Propionibacterium acnes Induces an IL-17 Response in Acne Vulgaris that Is Regulated by Vitamin A and Vitamin D. J. Investig. Dermatol. 2014, 134, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Karadag, A.S.; Ertugrul, D.T.; Bilgili, S.G.; Takci, Z.; Akin, K.O.; Calka, O. Immunoregulatory effects of isotretinoin in patients with acne. Br. J. Dermatol. 2012, 167, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Bohm, M.; Luger, T.A. The pilosebaceous unit is part of the skin immune system. Dermatology 1998, 196, 75–79. [Google Scholar] [CrossRef]
- Lawrence, D.; Shaw, M.; Katz, M. Elevated free testosterone concentration in men and women with acne vulgaris. Clin. Exp. Dermatol. 1986, 11, 263–273. [Google Scholar] [CrossRef]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell. Endocrinol. 2018, 465, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Miura, S.; Suganami, T.; Akaike, F.; Kanai, S.; Sugita, S.; Katsumata, A.; Aburatani, H.; Unterman, T.G.; Ezaki, O.; et al. Regulation of SREBP1c gene expression in skeletal muscle: Role of retinoid X receptor/liver X receptor and forkhead-O1 transcription factor. Endocrinology 2008, 149, 2293–2305. [Google Scholar] [CrossRef] [PubMed]
- Juhl, C.R.; Bergholdt, H.; Miller, I.M.; Jemec, G.; Kanters, J.K.; Ellervik, C. Dairy Intake and Acne Vulgaris: A Systematic Review and Meta-Analysis of 78,529 Children, Adolescents, and Young Adults. Nutrients 2018, 10, 1049. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Weng, X.; Tian, Y.; Wang, C.; Yang, Y.; Xu, K.; Zhang, C. Effects of omega-3 polyunsaturated fatty acids on steroidogenesis and cellular development in PCOS rats. Food Funct. 2019, 10, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Mirdamadi, Y.; Thielitz, A.; Wiede, A.; Goihl, A.; Papakonstantinou, E.; Hartig, R.; Zouboulis, C.C.; Reinhold, D.; Simeoni, L.; Bommhardt, U.; et al. Insulin and insulin-like growth factor-1 can modulate the phosphoinositide-3-kinase/Akt/FoxO1 pathway in SZ95 sebocytes in vitro. Mol. Cell. Endocrinol. 2015, 415, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Firlej, E.; Kowalska, W.; Szymaszek, K.; Roliński, J.; Bartosińska, J. The Role of Skin Immune System in Acne. J. Clin. Med. 2022, 11, 1579. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K.; Cottle, D.L.; Schweiger, P.J.; Watt, F.M. The Androgen Receptor Antagonizes Wnt/β-Catenin Signaling in Epidermal Stem Cells. J. Investig. Dermatol. 2015, 135, 2753–2763. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; You, L.; Sun, M.; Qu, C.; Dong, X.; Yin, X.; Ni, J. Inhibitory effects of Paris saponin I, II, VI and VII on HUVEC cells through regulation of VEGFR2, PI3K/AKT/mTOR, Src/eNOS, PLC gamma/ERK/MERK, and JAK2-STAT3 pathways. Biomed. Pharmacother. 2020, 131, 110750. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Li, Y.; Wang, X.; Hu, S.; Wang, H.; Shi, Q.; Wang, Y.; Yang, Y. Polyphyllin I suppresses human osteosarcoma growth by inactivation of Wnt/beta-catenin pathway in vitro and in vivo. Sci. Rep. 2017, 7, 7605. [Google Scholar] [CrossRef]
- Laly, A.C.; Sliogeryte, K.; Pundel, O.J.; Ross, R.; Keeling, M.C.; Avisetti, D.; Waseem, A.; Gavara, N.; Connelly, J.T. The keratin network of intermediate filaments regulates keratinocyte rigidity sensing and nuclear mechanotransduction. Sci. Adv. 2021, 7, eabd6187. [Google Scholar] [CrossRef]
- Maruthappu, T.; Chikh, A.; Fell, B.; Delaney, P.J.; Brooke, M.A.; Levet, C.; Moncada-Pazos, A.; Ishida-Yamamoto, A.; Blaydon, D.; Waseem, A.; et al. Rhomboid family member 2 regulates cytoskeletal stress-associated Keratin 16. Nat. Commun. 2017, 8, 14174. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.R.; Morris, C.; Cunliffe, W.J.; Leigh, I.M. Keratin expression in pilosebaceous epithelia in truncal skin of acne patients. Br. J. Dermatol. 1996, 134, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Caillon, F.; O’Connell, M.; Eady, E.A.; Jenkins, G.R.; Cove, J.H.; Layton, A.M.; Mountford, A.P. Interleukin-10 secretion from CD14+ peripheral blood mononuclear cells is downregulated in patients with acne vulgaris. Br. J. Dermatol. 2010, 162, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Akaza, N.; Akamatsu, H.; Kishi, M.; Mizutani, H.; Ishii, I.; Nakata, S.; Matsunaga, K. Effects of Propionibacterium acnes on various mRNA expression levels in normal human epidermal keratinocytes in vitro. J. Dermatol. 2009, 36, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Komine, M.; Rao, L.S.; Freedberg, I.M.; Simon, M.; Milisavljevic, V.; Blumenberg, M. Interleukin-1 induces transcription of keratin K6 in human epidermal keratinocytes. J. Investig. Dermatol. 2001, 116, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Chonglou. Genus. Plants; Science Press: Beijing, China, 1998; p. 220. [Google Scholar]
- Huang, W.; Zhou, J. Study on the steroidal saponin ligand composition of Shigarou. Med. Phar. Yunnan 1962, 64–65. [Google Scholar]
- Huang, X.; Gao, W.; Man, S.; Zhao, Z. Advances in studies on saponins in plants of Paris Land their biosynthetic approach. Chin. Tradit. Herb. Drugs 2009, 40, 483–489. [Google Scholar]
- Chen, C.; Zhou, J.; Zhang, Y.; Gao, C. Studies on the saponin composition of plants in yunnan vim steroidal saponins in paris luquanensis. Plant Divers. 1983, 219–223. [Google Scholar]
- Wang, Y.; Gao, W.; Yuan, L.; Liu, X.; Wang, S.; Chen, C. Chemical constituents from rhizome of Paris polyphylla var. yunnanensis. Chin. Tradit. Herb. Drugs 2007, 17–20. [Google Scholar]
- Huang, X.; Gao, W.; Man, S.; Yan, J.; Wang, Y. Chemical constituents from herbs of Paris verticillata. China J. Chin. Mater. Med. 2009, 34, 1812–1815. [Google Scholar]
- Sha, A.; Liu, Y.; Qiu, X.; Xiong, B. Polysaccharide from Paris polyphylla improves learning and memory ability in D-galactose-induced aging model mice based on antioxidation, p19/p53/p21, and Wnt/β-catenin signaling pathways. Int. J. Biol. Macromol. 2023, 251, 126311. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, L.; Wang, G.; Wang, H.; Dai, Y.; Yang, X.; Ye, W.; Li, Y. Triterpenoid saponins from rhizomes of Paris polyphylla var. yunnanensis. Carbohydr. Res. 2013, 368, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Thuy, T.T.V.; Lien, T.K.V.; Quan, H.N.; Khang, V.P.; Dung, T.N.; Lan, T.N.N.; Mau, H.C. Cytotoxic effects of steroidal glycosides isolated from the Paris vietnamensis plant on cancer cell lines and against bacterial strains. Biotechnol. Biotechnol. Equip. 2019, 33, 1516–1524. [Google Scholar]
- Huang, X.; Gao, W.; Man, S.; Gao, Y.; Huang, L.; Liu, C. Isolation and Identification of Compounds Present in Rhizomes of Paris axialis H. Li and Study of Their Cytotoxic Effects. Lat. Am. J. Pharm. 2011, 30, 540–545. [Google Scholar]
- Yang, Y.; Zhao, Y.; Zuo, Z.; Wang, Y. Determination of Total Flavonoids for Paris polyphylla var. Yunnanensis in Different Geographical Origins Using UV and FT-IR Spectroscopy. J. AOAC Int. 2019, 102, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qiu, P.; Wang, M.; Lu, Y.; He, H.; Tang, H.; Zhang, B. New Steroidal Saponins Isolated from the Rhizomes of Paris mairei. Molecules 2021, 26, 6366. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.A.; Yang, C.Z.; Li, J.Q.; Wang, S.L.; Wu, J.Y. Heptasaccharide and octasaccharide isolated from Paris polyphylla var. yunnanensis and their plant growth-regulatory activity. Plant Sci. 2003, 165, 571–575. [Google Scholar] [CrossRef]
- Yan, L.; Gao, W.; Zhang, Y.; Wang, Y. A new phenylpropanoid glycosides from Paris polyphylla var. yunnanensis. Fitoterapia 2008, 79, 306–307, Erratum in Fitoterapia 2009, 80, 143. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Ni, W.; Chen, C.; Liu, H. Seeing the light: Shifting from wild rhizomes to extraction of active ingredients from above-ground parts of Paris polyphylla var. yunnanensis. J. Ethnopharmacol. 2018, 224, 134–139. [Google Scholar] [CrossRef]
- Song, W.; Zheng, W.; Zhang, J.; Zhang, T.; Liu, S.; Yu, L.; Ma, B. Metabolism of saponins from traditional Chin medicines: A review. Acta Pharm. Sin. 2018, 53, 1609–1619. [Google Scholar]
- Osbourn, A. Saponins and plant defence—A soap story. Trends Plant Sci. 1996, 1, 142. [Google Scholar] [CrossRef]
- Tian, W.S.; Xu, Q.H.; Chen, L.; Zhao, C.F. Synthesis of pennogenin utilizing the intact skeleton of diosgenin. Sci. China Ser. B Chem. 2004, 47, 142–144. [Google Scholar] [CrossRef]
- Liu, F.; Li, L.; Tian, X.; Zhang, D.; Sun, W.; Jiang, S. Chemical Constituents and Pharmacological Activities of Steroid Saponins Isolated from. J. Chem. 2021, 2021, 1442906. [Google Scholar] [CrossRef]
- Singh, S.B.; Thakur, R.S.; Schulten, H. Spirostanol saponins from Paris polyphylla, structures of polyphyllin C, D, E and F. Phytochem 1980, 21, 2925–2929. [Google Scholar] [CrossRef]
- Viet Cuong, L.C.; Nhi, N.P.K.; Ha, T.P.; Anh, L.T.; Dat, T.T.H.; Oanh, P.T.T.; Phuong, N.T.M.; Thu, V.T.K.; Duc, H.V.; Anh, H.L.T. A new steroidal saponin from the aerial parts of Solanum torvum. Nat. Prod. Res. 2022, 36, 4892–4897. [Google Scholar] [CrossRef]
- Yu, L.; Li, Y.; Gao, W.; Ling, S.; Ni, W.; Ji, Y.; Liu, H. Steroidal saponins with cytotoxic activity from the stems and leaves of Paris fargesii. New J. Chem. 2022, 46, 19136–19146. [Google Scholar] [CrossRef]
- Singh, S.B.; Thakur, R.S.; Schulten, H.R. Furostanol saponins from Paris polyphylla: Structures of polyphyllin G and H. Phytochem 1982, 21, 2079–2082. [Google Scholar] [CrossRef]
- Xu, X.; Li, T.; Fong, C.M.V.; Chen, X.; Chen, X.; Wang, Y.; Huang, M.; Lu, J. Saponins from Chin Medicines as Anticancer Agents. Molecules 2016, 21, 1326. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Wang, Y.; Li, X.; Man, S.; Gao, W. Chemical constituents and antitumor activity from Paris polyphylla Smith var. yunnanensis. Nat. Prod. Res. 2017, 31, 660–666. [Google Scholar] [CrossRef]
- Tuchayi, S.M.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne vulgaris. Nat. Rev. Dis. Prim. 2015, 1, 15029. [Google Scholar] [CrossRef]
- Williams, H.C.; Dellavalle, R.P.; Garner, S. Acne vulgaris. Lancet 2012, 379, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Manhong, Z.; Du, W.; Long, S.; Han, J.; Shen, Y.; Li, T. Effect of Rhizoma Paridis Total Saponins on TNF-α and IL-1β Secretion in Rat Peritoneal Macrophages Induced by Lipopolysaccharide. J. Sichuan Tradit. Chin. Med. 2008, 26, 14–16. [Google Scholar]
- Leisheng, W.; Tian, M.; Yanpin, Z.; Shiqiao, L.; Yu, L.; Shaoxian, L. Diosgenin inhibits IL-1β-induced expression of inflammatory mediators in human osteoarthritis chondrocytes. Int. J. Clin. Exp. Pathol. 2015, 8, 4830–4836. [Google Scholar]
- Long, J.; Pi, X.; Tu, Y. Effect of Polyphylin I on proliferation and expressions of HIF-1α, VEGF in laryngeal carcinoma cell line Hep-2 under hypoxia. Acta Univ. Med. Anhui 2016, 30, 453–455. [Google Scholar]
- Wu, L.; Peng, X.; Ju, R.; Yang, S.; Li, H. Optimization of microwave ultrasound-assisted extraction of paris and its anti-inflammatory activity by response surface methodology. J. Chin Med. Mater. 2022, 1700–1704. [Google Scholar]
- Li, Y.; Sun, Y.; Fan, L.; Chen, X.; Mei, Q.; Yue, Z.; Li, Z.; Zheng, H.; Tang, Y. Effects of Polyphyllin VI on Metastasis of colon Cancer Cells LoVo and the Possible Mechanisms. Milit. Med. J. South China 2015, 29, 571–574. [Google Scholar]
- Zhang, C.; Li, Q.R.; Qin, G.Z.; Zhang, Y.; Li, C.Y.; Han, L.W.; Wang, R.C.; Wang, S.D.; Chen, H.X.; Liu, K.C.; et al. Anti-angiogenesis and anti-metastasis effects of Polyphyllin VII on Hepatocellular carcinoma cells in vitro and in vivo. Chin. Med. 2021, 16, 41. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.W.; Park, H.J.; Jung, D.H.; Kim, T.W.; Park, Y.M.; Kim, B.O.; Sohn, E.H.; Moon, E.Y.; Um, S.H.; Rhee, D.K.; et al. Inhibition of TNF-alpha-induced adhesion molecule expression by diosgenin in mouse vascular smooth muscle cells via downregulation of the MAPK, Akt and NF-κB signaling pathways. Vasc. Pharmacol. 2010, 53, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.L.; Zhu, M.; Yu, Z.W.; Yin, D.; Lu, F.F.; Pu, Y.Y.; Zhao, C.; He, C.; Cao, L. Therapeutic effects of diosgenin in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2017, 313, 152–160. [Google Scholar] [CrossRef]
- Quan, Q.; Weng, D.; Li, X.; An, Q.; Yang, Y.; Yu, B.; Ma, Y.; Wang, J. Analysis of drug efficacy for inflammatory skin on an organ-chip system. Front. Bioeng. Biotechnol. 2022, 10, 939629. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, C.; Wang, J.; Yang, L.; Lv, Z.; An, Q.; Wang, Y.; Shao, X.; Wang, F.; Huo, T.; et al. Rhizoma Paridis saponins attenuate Gram-negative bacteria-induced inflammatory acne by binding to KEAP1 and modulating Nrf2 and MAPK pathways. J. Cell. Mol. Med. 2024, 28, e18146. [Google Scholar] [CrossRef] [PubMed]
- Kuehnast, T.; Cakar, F.; Weinhäupl, T.; Pilz, A.; Selak, S.; Schmidt, M.A.; Rüter, C.; Schild, S. Comparative analyses of biofilm formation among different Cutibacterium acnes isolates. Int. J. Med. Microbiol. 2018, 308, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.Q.; Guo, L.L.; Pei, F.; Chang, X.Y.; Zhang, R. Polyphyllin G exhibits antimicrobial activity and exerts anticancer effects on human oral cancer OECM-1 cells by triggering G2/M cell cycle arrest by inactivating cdc25C-cdc2. Arch. Biochem. Biophys. 2018, 644, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, Q.; Liu, B.; Hu, J.; Zhang, C. Antibacterial Effects of Different Saponins from Paris polyphylla on Acne-associated Pathogens In Vitro. Chin. Dermat. Int. Tradit. West. Med. 2022, 21, 225–227. [Google Scholar]
- Wang, Q.; Sun, D.; He, L.; Xu, D.; Wu, W.; Li, Q.; Liu, H.; Du, Y. Evaluation of the Inhibitory Effect of Total Saponins of Polyphylla and its Fraction on Acne-associated Pathogens. Chin. J. Dermatovenereol. 2016, 30, 899–901. [Google Scholar]
- Carlavan, I.; Bertino, B.; Rivier, M.; Martel, P.; Bourdes, V.; Motte, M.; Déret, S.; Reiniche, P.; Menigot, C.; Khammari, A.; et al. Atrophic scar formation in patients with acne involves long-acting immune responses with plasma cells and alteration of sebaceous glands. Br. J. Dermatol. 2018, 179, 906–917. [Google Scholar] [CrossRef]
- Wang, J.; Liu, R.; Xiao, H.; Bi, H.; Li, Z. The effects of Paridis saponin II on the production of cell cytokines in CD4+CD25+ T regulation cells from lupus nephritis patients’ peripheral blood. Prog. Mod. Biomed. 2010, 10, 50–53. [Google Scholar]
- Li, C. The Study on Anti-Breast Cancer and Immunomodulating Effects of Rhizom Paridis Saponins. Ph.D. Thesis, Northeast Normal University, Changchun, China, 2011. [Google Scholar]
- Fallon, M.E.; Hinds, M.T. Single cell morphological metrics and cytoskeletal alignment regulate VCAM-1 protein expression. Biochem. Biophys. Res. Commun. 2021, 555, 160–167. [Google Scholar] [CrossRef]
- Hart, J. Comparative study of serum soluble VCAM-1 and ICAM-1 levels in the early neonatal period. Acta Paediatr. 1999, 88, 1414. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Yang, X.; Cao, Y.; Yu, T.; Xi, A.; Cheng, L.; Jin, Y.; Chen, Q. Clinical and Immunohistochemical Research of Piyan Xiaojin Decoction-2 Combining with Narrow-Spectrum UVB Exposure Treating Atopic Dermatitis (AD) with Blood Deficiency and Wind Dryness. Chin. Arch. Tradit Chin. Med. 2012, 30, 559–561. [Google Scholar]
- Chai, H. Antitumor and Anti-Pulmonary Metastatic Mechanisms of Rhizoma Paridis Saponin. Master’s Thesis, Tianjin University of Science and Technology, Tianjin, China, 2016. [Google Scholar]
- Gao, L.; Li, F.; Kang, L.; Si, Y.; Wang, H. Effects of the pariphyllin on expression of ICAM-1 and VCAM-1 in injury induced by hydrogen peroxide in the human umbiliar vein endothelial cell. China J. Mod. Med. 2009, 19, 1968–1972. [Google Scholar]
- Zouboulis, C.C. Acne and sebaceous gland function. Clin. Dermatol. 2004, 22, 360–366. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef]
- Mu, D.; Li, D.; Li, J.; Yu, H.; Chen, W.; Liang, J.; Wang, D.; Li, A.; Qing, Z.; Zhang, B. Long non-coding RNA HULC protects against atherosclerosis via inhibition of PI3K/AKT signaling pathway. Iubmb Life 2021, 73, 202–212. [Google Scholar] [CrossRef]
- Liu, J. miR-338-3p Inhibits Skin Inflammation and Lung Cancer Progression by Regulating AKT Signaling Pathway. Ph.D. Thesis, Northern Forest University, Harbin, China, 2018. [Google Scholar]
- Chen, M.; Gao, S.; Liu, H.; Tang, Q.; Luo, H. Epigenetic modification regulation of Wnt pathway inhibitors to study the mechanism of polyphyllin V against colorectal cancer. Chin J. Col. Dis. (Electr. Ed.) 2019, 8, 574–579. [Google Scholar]
- Melnik, B.; Schmitz, G. FGFR2 signaling and the pathogenesis of acne. JDDG J. Dtsch. Dermatol. Ges. 2008, 6, 721–728. [Google Scholar] [CrossRef]
- Trenam, C.W.; Dabbagh, A.J.; Morris, C.J.; Blake, D.R. Skin inflammation induced by reactive oxygen species (ROS): An in-vivo model. Br. J. Dermatol. 1991, 125, 325–329. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Adam, W.; Kurz, A.; Saha-Möller, C.R. Peroxidase-catalyzed oxidative damage of DNA and 2′-deoxyguanosine by model compounds of lipid hydroperoxides: Involvement of peroxyl radicals. Chem. Res. Toxicol. 2000, 13, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Ayres, S., Jr.; Mihan, R. Acne Vulgaris and Lipid Peroxidation: New Concepts in Pathogenesis and Treatment. Int. J. Dermatol. 1978, 17, 305–307. [Google Scholar] [CrossRef]
- Li, Y.; Guan, L.; Chen, L.; Zhao, M.; Ding, L.; Meng, C.; Gao, H.; Wang, Z. Qualitative and quantitative analysis of Paris polyphylla var. chinensis by UPLC-Q-TOF-MS/MS and HPLC. China J. Chin. Mater. Med. 2021, 46, 2900–2911. [Google Scholar]
- Gao, Y.; Yang, L.; Yang, Y.; Wang, X. In vitro scavenging of reactive oxygen species and antioxidant effect of Paris polyphylla Sm extract. Chin. Tradit. Patent. Med. 2007, 195–198. [Google Scholar]
- Jiang, W.; Li, X.; Wan, J.; Mei, S.; Fu, X. HPLC Fingerprint of Total Saponins of Aerial Parts of Paris polyphylla var. yunnanenis. Mod. Chin. Med. 2021, 23, 57–61. [Google Scholar]
- Fu, Y.; Fu, Q.; Yang, L.; Bao, Y.; Tian, Q.; Gao, Y.; Liu, X.; Huang, Q. Utilizing the above-ground extract of Paris polyphylla as a Nat antioxidant and antimicrobial additive in soap formulation. Biomass Convers. Biorefinery 2023, 14, 9821–9838. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, D. A study on the protective effect of Paris polyphylla saponin on photoaging. In Proceedings of the 13th Annual Academic Conference of the Chinese Society of Biotechnology and the 2019 National Biotechnology Conference, Chengdu, China, 2019; Yunnan Agriculture University: Kunming, China, 2019; p. 652. [Google Scholar]
- Xu, L. Experimental and Clinical Studies on the Efficacy of a Complex Containing Paris polyphylla Extract and Hyaluronic Acid in Acne Vulgaris. Master’s Thesis, Kunming Medical University, Kunming, China, 2013. [Google Scholar]
- Fang, T. Evaluation of Antimicrobial Activity of Paris polyphylla Extract against Propionibacterium acnes with Topical Dosage Form. Master’s Thesis, Kunming Medical University, Kunming, China, 2023. [Google Scholar]
- Wang, Q.; Zhou, X.; Zhao, Y.; Xiao, J.; Lu, Y.; Shi, Q.; Wang, Y.; Wang, H.; Liang, Q. Polyphyllin I Ameliorates Collagen-Induced Arthritis by Suppressing the Inflammation Response in Macrophages through the NF-κB Pathway. Front. Immunol. 2018, 9, 2091. [Google Scholar] [CrossRef]
- Yang, M.; Zou, J.; Zhu, H.M.; Liu, S.L.; Wang, H.; Bai, P.; Xiao, X. Paris saponin II inhibits human ovarian cancer cell-induced angiogenesis by modulating NF-κB signaling. Oncol. Rep. 2015, 33, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Pang, D.; Yang, C.; Li, C.; Zou, Y.; Feng, B.; Li, L.; Liu, W.; Luo, Q.; Chen, Z.; Huang, C. Polyphyllin II inhibits liver cancer cell proliferation, migration and invasion through downregulated cofilin activity and the AKT/NF-κB pathway. Biol. Open 2020, 9, o46854. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.J.; Chen, C.X.; Ni, W.; Yan, H.; Liu, H.Y. C-22-steroidal lactone glycosides from stems and leaves of Paris polyphylla var. yunnanensis. Fitoterapia 2013, 84, 248–251. [Google Scholar] [CrossRef]
- Tan, L.; Xiang, M.; Mi, C.; Li, Z.; Tian, Z.; Xiang, B.; Zhou, W. Effect of Rhizoma Paridis Total Saponins on airway inflammation in a mouse model of asthma and the mechanism of the effect on airway inflammation in a mouse model of asthma. Chin. J. Geront. 2017, 37, 4703–4704. [Google Scholar]
- Li, L.; Liang, C.; Shan, L.; Xie, X.; Zhao, D.; Zhou, M. Effects of Rhizoma Paridis Total Saponins on Levels of Cytokines in Blood Serum of Rats Subjected to Multiple Trauma. J. Liaoning Univ. Trad. Chin. Med. 2009, 11, 241–244. [Google Scholar]
- Lu, R.; Su, X.; Cai, S. Optimization of Extraction Process of Saponins of Paris polyphylla with Orthogonal Design Method. Lishizhen Med. Mater. Med. Res. 2007, 1044–1045. [Google Scholar]
- Sun, Z.; Zhang, L.; Li, L.; Tian, J. Studies on the Extraction Process of Total Saponins from Paris polyphylla Smith. J. Chin. Med. Mater. 2007, 726–729. [Google Scholar]
- Tan, W.; Dai, J.; Xiong, Y.; Yang, M.; Wang, H. Extraction and Determination of Steroidal Saponins from Paris polyphylla Smith. J. Yunnan Univ. Nation (Nat. Sci. Ed.) 2010, 19, 127–129. [Google Scholar]
- Lu, W.; Yang, G.; Ye, F.; Hu, Y. Optimization of Extraction Technology for Total Saponins in Paris polyphylla Smit by Orthogonal Design. China Pharm. 2015, 18, 1484–1487. [Google Scholar]
- Qian, L.; Chen, Y. Study on Extraction and Stability of Active Components from Paris polyphylla. J. Huaihua Univ. 2019, 38, 11–16. [Google Scholar]
- Wang, S.; Su, Y.; Li, H.; Yang, P.; Wu, P. Optimization of the reflux extraction process of steroidal saponins from paris herbs by response surface methodology. Shaanxi J. Agric. Sci. 2019, 65, 46–52. [Google Scholar]
- Liu, B.; Shao, M.; Yang, Y.; Yang, G. Optimal design of an orthogonal test for the extraction of four saponins from Paris. Lis. Med. Mater. Med. Res. 2020, 31, 1591–1594. [Google Scholar]
- Luo, Y.; Liu, S.; Huang, S. Ultrasonic extraction of total saponins from Paris polyphylla smith by orthogonal design. Pract. Pharm. Clin. Remedies 2010, 13, 185–187. [Google Scholar]
- Liu, J.; Duan, Z.; Duan, B.; Xia, C. Ultrasonic Extraction Technology of Four Kinds of Polyphyllin in Paris vietnamensis. J. Anhui Agric. Sci. 2016, 44, 158–160. [Google Scholar]
- Li, W.; Peng, Y.; Li, Y.; Chen, X. Ultrasonic Extraction of Paris polyphylla saponins from paris. Chin. Tradit. Patent. Med. 2010, 32, 218–220. [Google Scholar]
- Wang, F.; Ma, X.; Li, Z.; Yuan, Y.; Qu, L. Ultrasonic Extraction of Total Saponins and Content Difference of Active Components from Colloidal and Amyloid Paris polyphylla. Pharm. J. Chin People’s Lib. Army 2017, 33, 315–318. [Google Scholar]
- Lan, J.; Yang, M.; Ma, J. Study of the extraction process in Paris. Chin. Tradit. Herb. Drugs 2006, 871–872. [Google Scholar]
- Yu, Z.; Liu, Y.; Li, H.; Li, F. Study on the extraction of total saponins from Paris polyphylla smith by Microwave. Lis. Med. Mater. Med. Res. 2013, 24, 833–834. [Google Scholar]
- Xiao, X.H.; Yuan, Z.Q.; Li, G.K. Separation and purification of steroidal saponins from Paris polyphylla by microwave- assisted extraction coupled with countercurrent chromatography using evaporative light scattering detection. J. Sep. Sci. 2014, 37, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Q.; Zhang, J.S.; Wang, C.Z. Extraction of steroid saponins from Paris polyphylla Sm. var. yunnanensis using novel ultrahigh pressure extraction technology. Pharm. Chem. J. 2007, 41, 424–429. [Google Scholar] [CrossRef]
- Tu, Y.; Jiang, L.; Yang, Y. Study on the Process for Supercritical Carbon Dioxide Extraction of Pennogenin Ingredients of Parispolyphylla Sm. var. yunnanensis. J. Kun Univ. Sci. Tech. (Nat. Sci. Ed.) 2018, 43, 98–104. [Google Scholar]
- Tong, L.; Cai, H. Enzymatic Extraction Technology of Total Saponins in Paris. Aca. Peri. Farm. Pro. Pro. 2012, 62–67. [Google Scholar]
- Nian, S.; Zhang, H.; Zheng, Y. Studies on Water-extracting Technology from Rhizoma Paris. J. Yunnan Univ. Tradit. Chin. Med. 2006, 13–16. [Google Scholar]
- Negi, J.S.; Bisht, V.K.; Bhandari, A.K.; Bhatt, V.P.; Singh, P.; Singh, N. Paris polyphylla: Chemical and biological prospectives. Anti-Cancer Agents Med. Chem. 2014, 14, 833. [Google Scholar] [CrossRef]
- Liu, N.; Chen, L.; Zhang, Y.; Zhang, L.; Fan, J. Response surface optimization on ultrasound-assisted extraction of hemp cannabinoids and study on anti-oxidation property. Mod. Chem. Ind. 2019, 39, 144–149. [Google Scholar]
- Sadowska-Rociek, A.; Surma, M.; Cieslik, E. Comparison of different modifications on QuEChERS sample preparation method for PAHs determination in black, green, red and white tea. Environ. Sci. Pollut. R. 2014, 21, 1326–1338. [Google Scholar] [CrossRef] [PubMed]
- Cheok, C.Y.; Salman, H.A.K.; Sulaiman, R. Extraction and quantification of saponins: A review. Food Res. Int. 2014, 59, 16–40. [Google Scholar] [CrossRef]
- Jun, X. High-Pressure Processing as Emergent Technology for the Extraction of Bioactive Ingredients from Plant Materials. Crit. Rev. Food Sci. 2013, 53, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.C.; Choi, W.Y.; Kim, J.S.; Yoon, C.S.; Lim, H.W.; Cho, J.S.; Ahn, J.H.; Lee, H.Y. Effect of ultra high pressure processing on immuno-modulatory activities of the fruits of Rubus coreanus Miquel. Innov. Food Sci. Emerg. 2011, 12, 207–215. [Google Scholar] [CrossRef]
- Ning, N.; Zhou, J. Research progress of ultra-high pressure extraction technology in Chinese medicine extraction. Tianjin Pharm. 2008, 62–64. [Google Scholar]
- Pan, S.Y.; Chiang, P.C.; Pan, W.B.; Kim, H. Advances in state-of-art valorization technologies for captured CO2 toward sustainable carbon cycle. Crit. Rev. Environ. Sci. Technol. 2018, 48, 471–534. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Chatterjee, D.; Singhal, R.S. Supercritical Carbon Dioxide Extraction of Squalene from Amaranthus paniculatus: Experiments and Process Characterization. Food Bioprocess Technol. 2012, 5, 2506–2521. [Google Scholar] [CrossRef]
- Ye, F.; Yang, G.; Li, Z.; Wang, G. Overview of the extraction process and quality control studies of Paris polyphylla saponins. China Pharm. 2011, 14, 1207–1209. [Google Scholar]
- Gao, Y.H.; Liu, C.; Yao, F.; Chen, F.S. Aqueous enzymatic extraction of peanut oil body and protein and evaluation of its physicochemical and functional properties. Int. J. Food Eng. 2021, 17, 897–908. [Google Scholar] [CrossRef]
- Jian, Q.; Gao, J.; Chen, Z.; Jing, M.; Xue, J.; Liu, X. Extraction of active ingredient from Chinese materia medica by enzyme and enzyme coupling technique. J. Gansu Univ. Chin. Med. 2019, 36, 79–82. [Google Scholar]
- Wang, Z.; Yang, L.; Zeng, X.; Li, P.; Zhang, X. Application Progress on Enzymatic Extraction Technology in Extraction of Chemical Compositions of Chinese Medicine. World Chin. Med. 2013, 8, 104–106. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhang, J.; Cheng, W.; Di, F.; Wang, C.; An, Q. Saponins of Paris polyphylla for the Improvement of Acne: Anti-Inflammatory, Antibacterial, Antioxidant and Immunomodulatory Effects. Molecules 2024, 29, 1793. https://doi.org/10.3390/molecules29081793
Li L, Zhang J, Cheng W, Di F, Wang C, An Q. Saponins of Paris polyphylla for the Improvement of Acne: Anti-Inflammatory, Antibacterial, Antioxidant and Immunomodulatory Effects. Molecules. 2024; 29(8):1793. https://doi.org/10.3390/molecules29081793
Chicago/Turabian StyleLi, Luyao, Jiachan Zhang, Wenjing Cheng, Feiqian Di, Changtao Wang, and Quan An. 2024. "Saponins of Paris polyphylla for the Improvement of Acne: Anti-Inflammatory, Antibacterial, Antioxidant and Immunomodulatory Effects" Molecules 29, no. 8: 1793. https://doi.org/10.3390/molecules29081793