Transcriptome Analysis Did Not Show Endogenous Stem Cell Characteristics in Murine Lgr5+ Retinal Cells


Abstract
:1. Introduction
2. Results
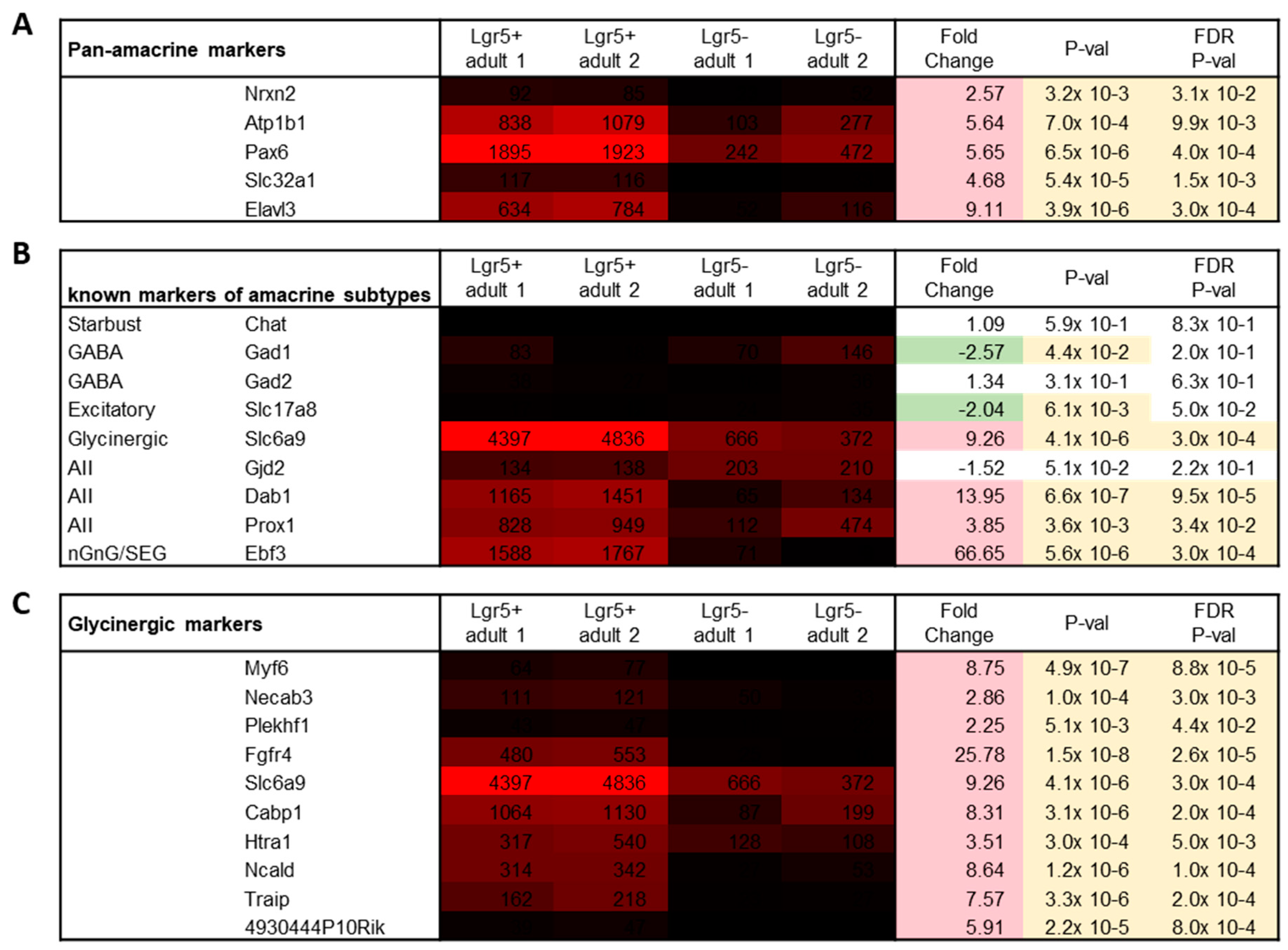
2.1. Lgr5+ Transcriptomics
RNA Quantity and Quality
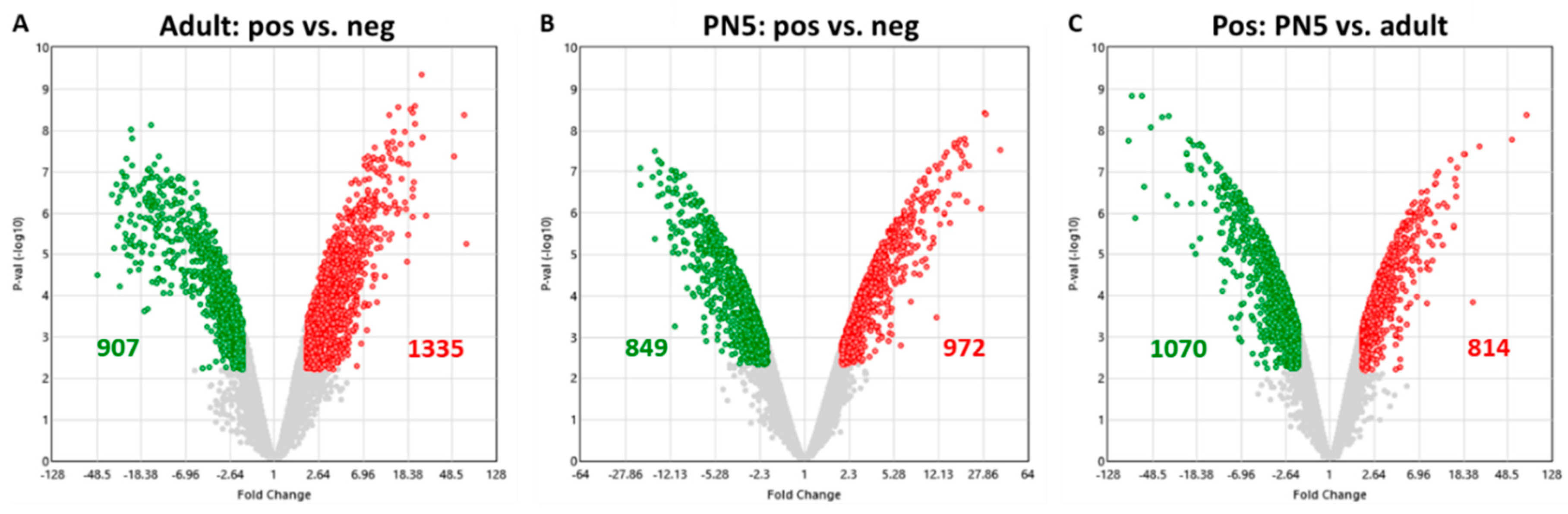
2.2. Quality Control of Microarray Processing
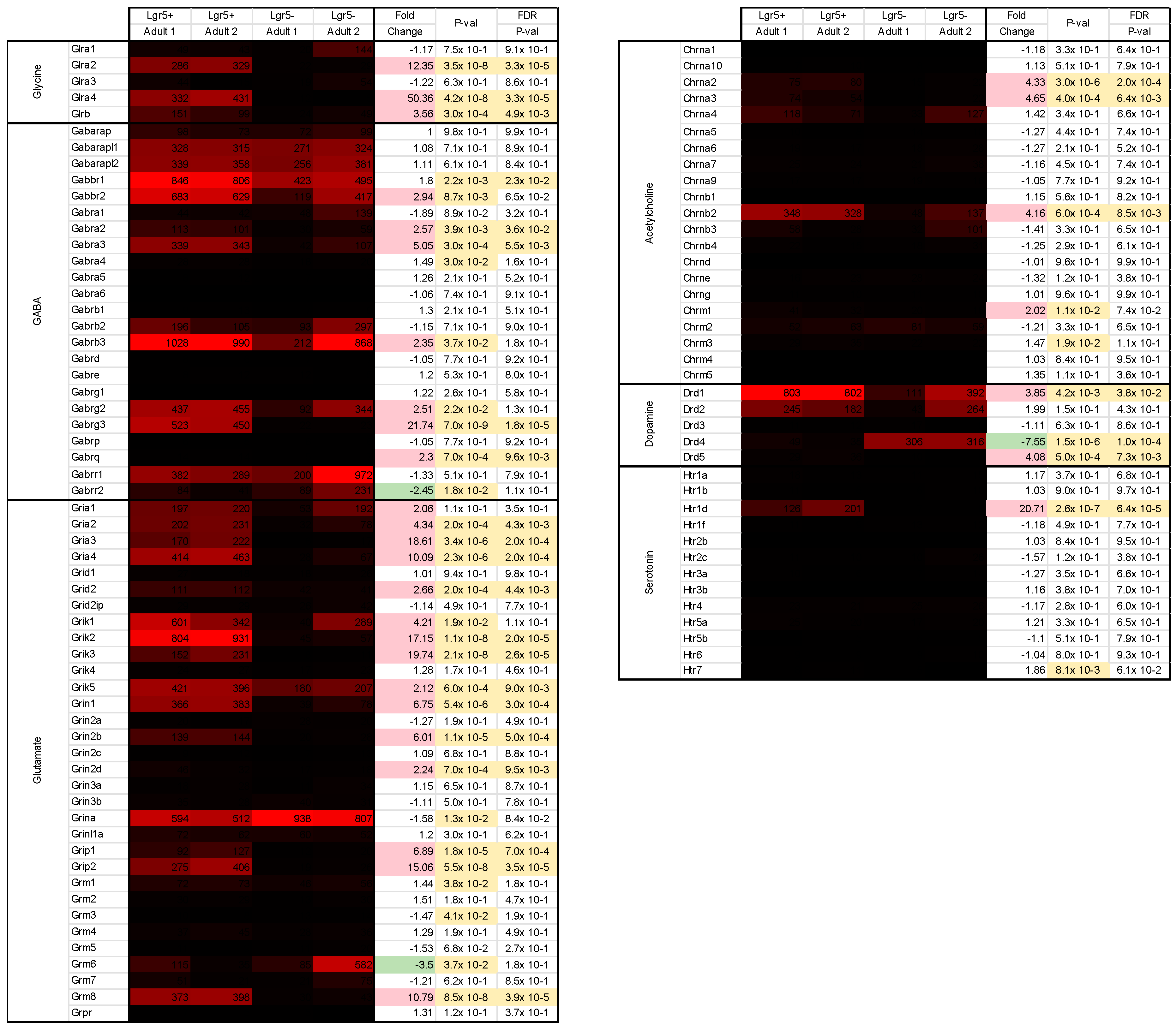
2.3. Differential Gene Expression (DGE) and PANTHER Analysis
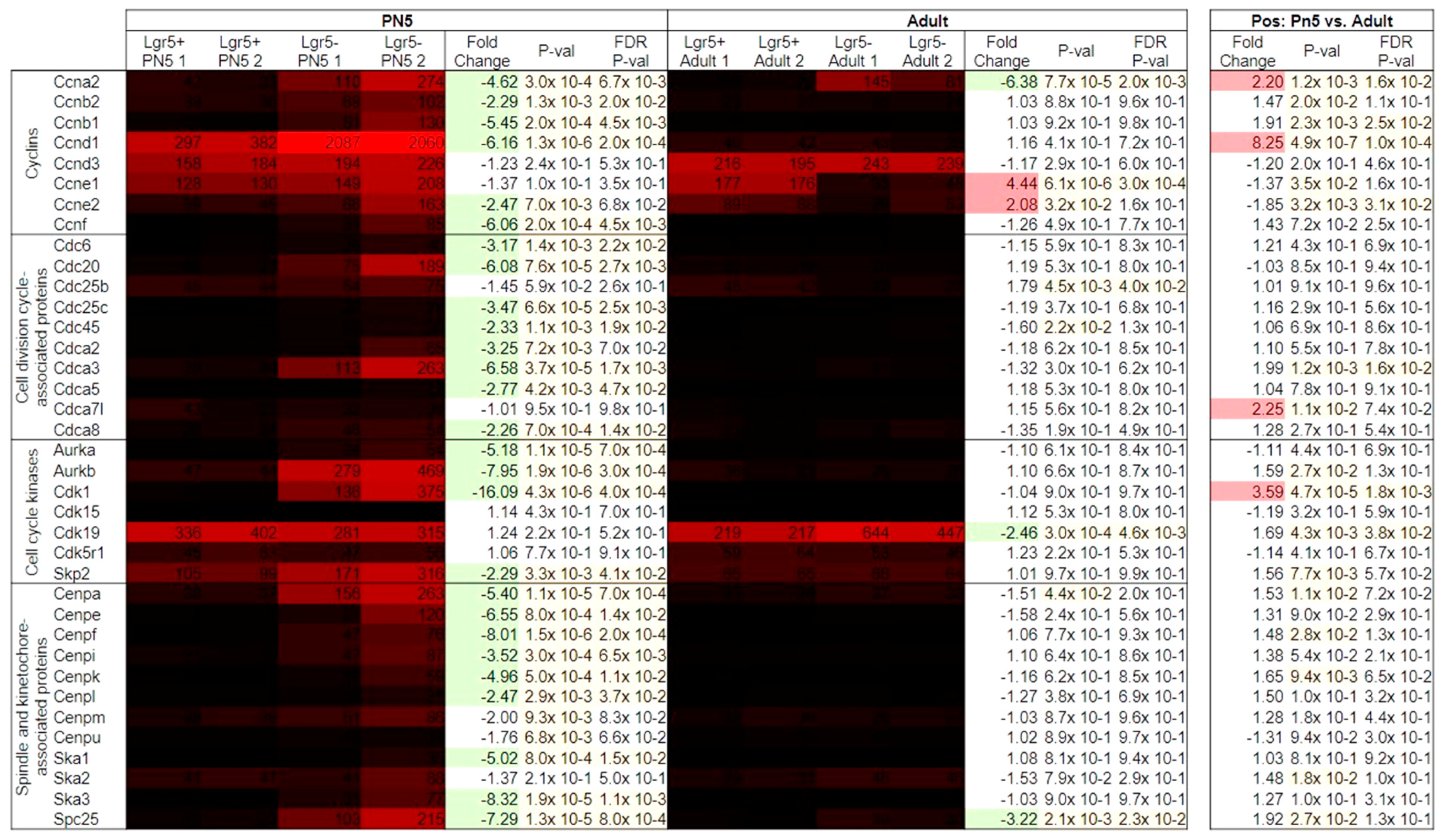
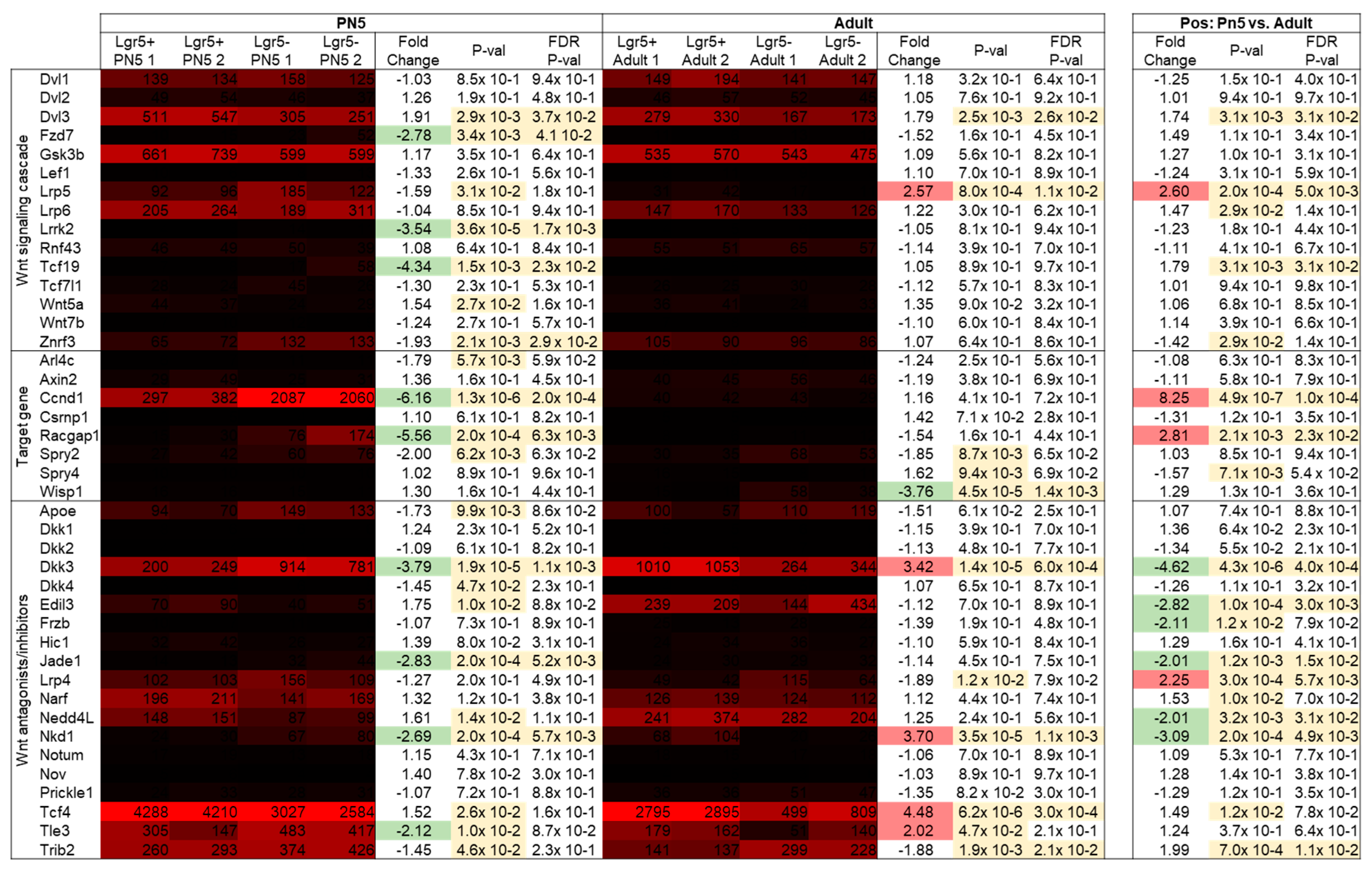
2.4. Gene List Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Lgr5 Transcriptomics
4.3. Analyses of DGE
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DGE | Differential gene expression |
| GO | Gene ontology |
| NSC | Neural stem cells |
| PCA | Principle component analysis |
| PANTHER | Protein ANalysis THrough Evolutionary Relationships |
References
- Chen, M.; Tian, S.; Glasgow, N.G.; Gibson, G.; Yang, X.; Shiber, C.E.; Liu, H. Lgr5(+) amacrine cells possess regenerative potential in the retina of adult mice. Aging Cell 2015, 14, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Qiu, S.; Tian, L.; Snider, W.D.; Flannery, J.G.; Schaffer, D.V.; Chen, B. Wnt Regulates Proliferation and Neurogenic Potential of Müller Glial Cells via a Lin28/let-7 miRNA-Dependent Pathway in Adult Mammalian Retinas. Cell Rep. 2016, 17, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.A.; Montecinos-Oliva, C.; Inestrosa, N.C. Wnt Signaling in the Central Nervous System: New Insights in Health and Disease. Prog. Mol. Biol. Transl. Sci. 2018, 153, 81–130. [Google Scholar] [PubMed]
- De Lau, W.; Peng, W.C.; Gros, P.; Clevers, H. The R-spondin/Lgr5/Rnf43 module: Regulator of Wnt signal strength. Genes Dev. 2014, 28, 305–316. [Google Scholar] [CrossRef]
- Barker, N.; Van Es, J.H.; Kuipers, J.; Kujala, P.; Van Den Born, M.; Cozijnsen, M.; Clevers, H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; Huch, M.; Kujala, P.; van de Wetering, M.; Snippert, H.J.; van Es, J.H.; Danenberg, E. Lgr5(+ve) stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell 2010, 6, 25–36. [Google Scholar] [CrossRef]
- Osawa, H.; Takahashi, H.; Nishimura, J.; Ohta, K.; Haraguchi, N.; Hata, T.; Mori, M. Full-length LGR5-positive cells have chemoresistant characteristics in colorectal cancer. Br. J. Cancer 2016, 114, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Rot, S.; Kaune, T.; Taubert, H.; Greither, T.; Kotrba, J.; Güttler, A.; Bache, M. Prognostic impact of mRNA levels of LGR5 transcript variants in OSCC patients. BMC Cancer 2019, 19, 155. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Clevers, H. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Huch, M.; Bonfanti, P.; Boj, S.F.; Sato, T.; Loomans, C.J.; Van De Wetering, M.; Ringnalda, F. Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 2013, 32, 2708–2721. [Google Scholar] [CrossRef] [Green Version]
- Sukhdeo, K.; Koch, C.E.; Miller, T.E.; Zhou, H.; Rivera, M.; Yan, K.; Rich, J.N. The Lgr5 transgene is expressed specifically in glycinergic amacrine cells in the mouse retina. Exp. Eye Res. 2014, 119, 106–110. [Google Scholar] [CrossRef]
- Kay, J.N.; Voinescu, P.E.; Chu, M.W.; Sanes, J.R. Neurod6 expression defines new retinal amacrine cell subtypes and regulates their fate. Nat Neurosci. 2011, 14, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Trombetta, J.J. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.R.; Jones, B.W.; Watt, C.B.; Shaw, M.V.; Yang, J.H.; DeMill, D.; Koshevoy, P. Exploring the retinal connectome. Mol. Vis. 2011, 17, 355–379. [Google Scholar]
- Grudzinska, J.; Schemm, R.; Haeger, S.; Nicke, A.; Schmalzing, G.; Betz, H.; Laube, B. The beta subunit determines the ligand binding properties of synaptic glycine receptors. Neuron 2005, 45, 727–739. [Google Scholar] [CrossRef]
- Lynch, J.W. Molecular structure and function of the glycine receptor chloride channel. Physiol. Rev. 2004, 84, 1051–1095. [Google Scholar] [CrossRef]
- Livesey, F.J.; Cepko, C.L. Vertebrate neural cell-fate determination: Lessons from the retina. Nat. Rev. Neurosci. 2001, 2, 109–118. [Google Scholar] [CrossRef]
- Cepko, C. Intrinsically different retinal progenitor cells produce specific types of progeny. Nat. Rev. Neurosci. 2014, 15, 615–627. [Google Scholar] [CrossRef]
- Bassett, E.A.; Wallace, V.A. Cell fate determination in the vertebrate retina. Trends Neurosci. 2012, 35, 565–573. [Google Scholar] [CrossRef]
- Lustig, B.; Jerchow, B.; Sachs, M.; Weiler, S.; Pietsch, T.; Karsten, U.; Behrens, J. Negative feedback loop of Wnt signaling through upregulation of conductin/axin2 in colorectal and liver tumors. Mol. Cell. Biol. 2002, 22, 1184–1193. [Google Scholar] [CrossRef]
- Kopp, J.L.; Grompe, M.; Sander, M. Stem cells versus plasticity in liver and pancreas regeneration. Nat. Cell Biol. 2016, 18, 238–245. [Google Scholar] [CrossRef]
- MacNeil, M.A.; Heussy, J.K.; Dacheux, R.F.; Raviola, E.; Masland, R.H. The shapes and numbers of amacrine cells: Matching of photofilled with Golgi-stained cells in the rabbit retina and comparison with other mammalian species. J. Comp. Neurol. 1999, 413, 305–326. [Google Scholar] [CrossRef]
- Mo, Z.; Li, S.; Yang, X.; Xiang, M. Role of the Barhl2 homeobox gene in the specification of glycinergic amacrine cells. Development 2004, 131, 1607–1618. [Google Scholar] [CrossRef]
- Siegert, S.; Scherf, B.G.; Del Punta, K.; Didkovsky, N.; Heintz, N.; Roska, B. Genetic address book for retinal cell types. Nat. Neurosci. 2009, 12, 1197–1204. [Google Scholar] [CrossRef]
- Apaja, P.M.; Tuusa, J.T.; Pietilä, E.M.; Rajaniemi, H.J.; Petäjä-Repo, U.E. Luteinizing hormone receptor ectodomain splice variant misroutes the full-length receptor into a subcompartment of the endoplasmic reticulum. Mol. Biol. Cell. 2006, 17, 2243–2255. [Google Scholar] [CrossRef]
- Scott, D.J.; Layfield, S.; Yan, Y.; Sudo, S.; Hsueh, A.J.; Tregear, G.W.; Bathgate, R.A. Characterization of novel splice variants of LGR7 and LGR8 reveals that receptor signaling is mediated by their unique low density lipoprotein class A modules. J. Biol. Chem. 2006, 281, 34942–34954. [Google Scholar] [CrossRef]
- Hufnagel, R.B.; Brown, N.L. Chapter 27—Specification of Retinal Cell Types. In Patterning and Cell Type Specification in the Developing CNS and PNS, 1st ed.; Rubenstein, J., Rakic, P., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 1, pp. 519–536. [Google Scholar]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | DGE | #GO | #Input | Exp. | FE | p Value |
|---|---|---|---|---|---|---|
| Synaptic vesicle clustering | + | 4 | 4 | 0.25 | 16.00 | 6.89 × 10−4 |
| Postsynaptic density protein 95 clustering | + | 6 | 5 | 0.38 | 13.33 | 2.40 × 10−4 |
| Positive regulation of protein localization to synapse | + | 5 | 4 | 0.31 | 12.80 | 1.18 × 10−3 |
| Synaptic transmission, glycinergic | + | 7 | 5 | 0.44 | 11.43 | 3.91 × 10−4 |
| Neurotransmitter receptor transport, postsynaptic endosome to lysosome | + | 7 | 5 | 0.44 | 11.43 | 3.91 × 10−4 |
| Synaptic membrane adhesion | + | 10 | 7 | 0.63 | 11.20 | 2.77 × 10−5 |
| Neuron cell-cell adhesion | + | 9 | 6 | 0.56 | 10.66 | 1.29 × 10−4 |
| Postsynaptic neurotransmitter receptor diffusion trapping | + | 9 | 6 | 0.56 | 10.66 | 1.29 × 10−4 |
| Anterograde axonal protein transport | + | 8 | 5 | 0.50 | 10.00 | 6.05 × 10−4 |
| Negative regulation of protein kinase activity by regulation of protein phosphorylation | + | 8 | 5 | 0.50 | 10.00 | 6.05 × 10−4 |
| Phototransduction, visible light | − | 9 | 5 | 0.40 | 12.52 | 1.99 × 10−4 |
| Photoreceptor cell maintenance | − | 40 | 20 | 1.77 | 11.27 | 2.48 × 10−13 |
| Detection of light stimulus involved in sensory perception | − | 20 | 10 | 0.89 | 11.27 | 2.52 × 10−7 |
| Protein localization to non-motile cilium | − | 12 | 6 | 0.53 | 11.27 | 6.91 × 10−5 |
| Eye photoreceptor cell development | − | 34 | 13 | 1.51 | 8.62 | 4.94 × 10−8 |
| Camera-type eye photoreceptor cell differentiation | − | 20 | 7 | 0.89 | 7.89 | 1.03 × 10−4 |
| Non-motile cilium assembly | − | 50 | 16 | 2.22 | 7.21 | 1.14 × 10−8 |
| Phospholipid transport | − | 60 | 12 | 2.66 | 4.51 | 4.69 × 10−5 |
| Axoneme assembly | − | 60 | 11 | 2.66 | 4.13 | 1.90 × 10−4 |
| Smoothened signaling pathway | − | 74 | 12 | 3.28 | 3.66 | 2.71 × 10−4 |
| GO Term | DGE | #GO | #Input | Exp. | FE | p Value |
|---|---|---|---|---|---|---|
| Negative regulation of dopamine secretion | + | 5 | 5 | 0.21 | 23.51 | 2.37 × 10−5 |
| Spontaneous synaptic transmission | + | 5 | 4 | 0.21 | 18.81 | 2.94 × 10−4 |
| Postsynaptic neurotransmitter receptor diffusion trapping | + | 9 | 6 | 0.38 | 15.67 | 1.65 × 10−5 |
| Positive regulation of inhibitory postsynaptic potential | + | 8 | 5 | 0.34 | 14.69 | 1.09 × 10−4 |
| Postsynaptic density assembly | + | 8 | 5 | 0.34 | 14.69 | 1.09 × 10−4 |
| Trans-synaptic signaling, modulating synaptic transmission | + | 7 | 4 | 0.30 | 13.44 | 7.21 × 10−4 |
| Neurotransmitter receptor transport, postsynaptic endosome to lysosome | + | 7 | 4 | 0.30 | 13.44 | 7.21 × 10−4 |
| Positive regulation of AMPA receptor activity | + | 9 | 5 | 0.38 | 13.06 | 1.64 × 10−4 |
| Regulation of short-term neuronal synaptic plasticity | + | 18 | 10 | 0.77 | 13.06 | 8.15 × 10−8 |
| Regulation of synaptic vesicle priming | + | 9 | 5 | 0.38 | 13.06 | 1.64 × 10−4 |
| Retinal rod cell differentiation | − | 4 | 4 | 0.17 | 24.18 | 1.52 × 10−4 |
| Compartment pattern specification | − | 4 | 3 | 0.17 | 18.14 | 1.94 × 10−3 |
| Mitotic spindle midzone assembly | − | 7 | 5 | 0.29 | 17.27 | 6.12 × 10−5 |
| Negative regulation of photoreceptor cell differentiation | − | 6 | 4 | 0.25 | 16.12 | 4.28 × 10−4 |
| Regulation of attachment of spindle microtubules to kinetochore | − | 12 | 7 | 0.50 | 14.11 | 5.03 × 10−6 |
| Notch signaling involved in heart development | − | 7 | 4 | 0.29 | 13.82 | 6.51 × 10−4 |
| DNA unwinding involved in DNA replication | − | 8 | 4 | 0.33 | 12.09 | 9.46 × 10−4 |
| Blood vessel endothelial cell differentiation | − | 8 | 4 | 0.33 | 12.09 | 9.46 × 10−4 |
| Protein localization to kinetochore | − | 13 | 6 | 0.54 | 11.16 | 6.70 × 10−5 |
| Positive regulation of mammary gland epithelial cell proliferation | − | 9 | 4 | 0.37 | 10.75 | 1.32 × 10−3 |
| GO Term | DGE | #GO | #Input | Exp. | FE | p Value |
|---|---|---|---|---|---|---|
| Negative regulation of ubiquitin protein ligase activity | + | 10 | 5 | 0.38 | 13.32 | 1.36 × 10−4 |
| DNA damage response, signal transduction by p53 class mediator resulting in transcription of p21 class mediator | + | 11 | 5 | 0.41 | 12.11 | 1.92 × 10−4 |
| Ribosomal small subunit assembly | + | 16 | 6 | 0.60 | 9.99 | 1.00 × 10−4 |
| Regulation of development, heterochronic | + | 15 | 5 | 0.56 | 8.88 | 6.04 × 10−4 |
| Cytoplasmic translation | + | 50 | 16 | 1.88 | 8.52 | 1.17 × 10−9 |
| Ribosomal large subunit assembly | + | 34 | 9 | 1.28 | 7.05 | 1.90 × 10−5 |
| Sympathetic nervous system development | + | 23 | 6 | 0.86 | 6.95 | 5.13 × 10−4 |
| ATP-dependent chromatin remodeling | + | 36 | 8 | 1.35 | 5.92 | 1.58 × 10−4 |
| Positive regulation of Notch signaling pathway | + | 40 | 8 | 1.50 | 5.33 | 2.97 × 10−4 |
| Ventricular septum morphogenesis | + | 44 | 8 | 1.65 | 4.84 | 5.21 × 10−4 |
| Calcium ion export | − | 10 | 5 | 0.52 | 9.61 | 5.82 × 10−4 |
| Membrane repolarization | − | 15 | 7 | 0.78 | 8.96 | 6.30 × 10−5 |
| Cerebellar Purkinje cell differentiation | − | 13 | 6 | 0.68 | 8.87 | 2.25 × 10−4 |
| Sodium ion export across plasma membrane | − | 11 | 5 | 0.57 | 8.73 | 8.12 × 10−4 |
| Relaxation of muscle | − | 18 | 7 | 0.94 | 7.47 | 1.56 × 10−4 |
| G-protein coupled glutamate receptor signaling pathway | − | 16 | 6 | 0.83 | 7.20 | 5.45 × 10−4 |
| CGMP metabolic process | − | 17 | 6 | 0.88 | 6.78 | 7.06 × 10−4 |
| Long term synaptic depression | − | 18 | 6 | 0.94 | 6.40 | 9.03 × 10−4 |
| Neuron cellular homeostasis | − | 18 | 6 | 0.94 | 6.40 | 9.03 × 10−4 |
| Phototransduction | − | 28 | 9 | 1.46 | 6.17 | 6.09 × 10−5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trepp, C.; Quintela Pousa, A.M.; Enzmann, V. Transcriptome Analysis Did Not Show Endogenous Stem Cell Characteristics in Murine Lgr5+ Retinal Cells. Int. J. Mol. Sci. 2019, 20, 3547. https://doi.org/10.3390/ijms20143547
Trepp C, Quintela Pousa AM, Enzmann V. Transcriptome Analysis Did Not Show Endogenous Stem Cell Characteristics in Murine Lgr5+ Retinal Cells. International Journal of Molecular Sciences. 2019; 20(14):3547. https://doi.org/10.3390/ijms20143547
Chicago/Turabian StyleTrepp, Carolyn, Ana Maria Quintela Pousa, and Volker Enzmann. 2019. "Transcriptome Analysis Did Not Show Endogenous Stem Cell Characteristics in Murine Lgr5+ Retinal Cells" International Journal of Molecular Sciences 20, no. 14: 3547. https://doi.org/10.3390/ijms20143547
APA StyleTrepp, C., Quintela Pousa, A. M., & Enzmann, V. (2019). Transcriptome Analysis Did Not Show Endogenous Stem Cell Characteristics in Murine Lgr5+ Retinal Cells. International Journal of Molecular Sciences, 20(14), 3547. https://doi.org/10.3390/ijms20143547