Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis



Abstract
:1. Introduction
2. Results
2.1. RNA-Seq Datasets
2.2. Differentially Expressed Genes
2.3. Toxicity Pathway Analysis
2.4. Canonical Pathway Analysis
2.5. Upstream Regulator Analysis
3. Discussion
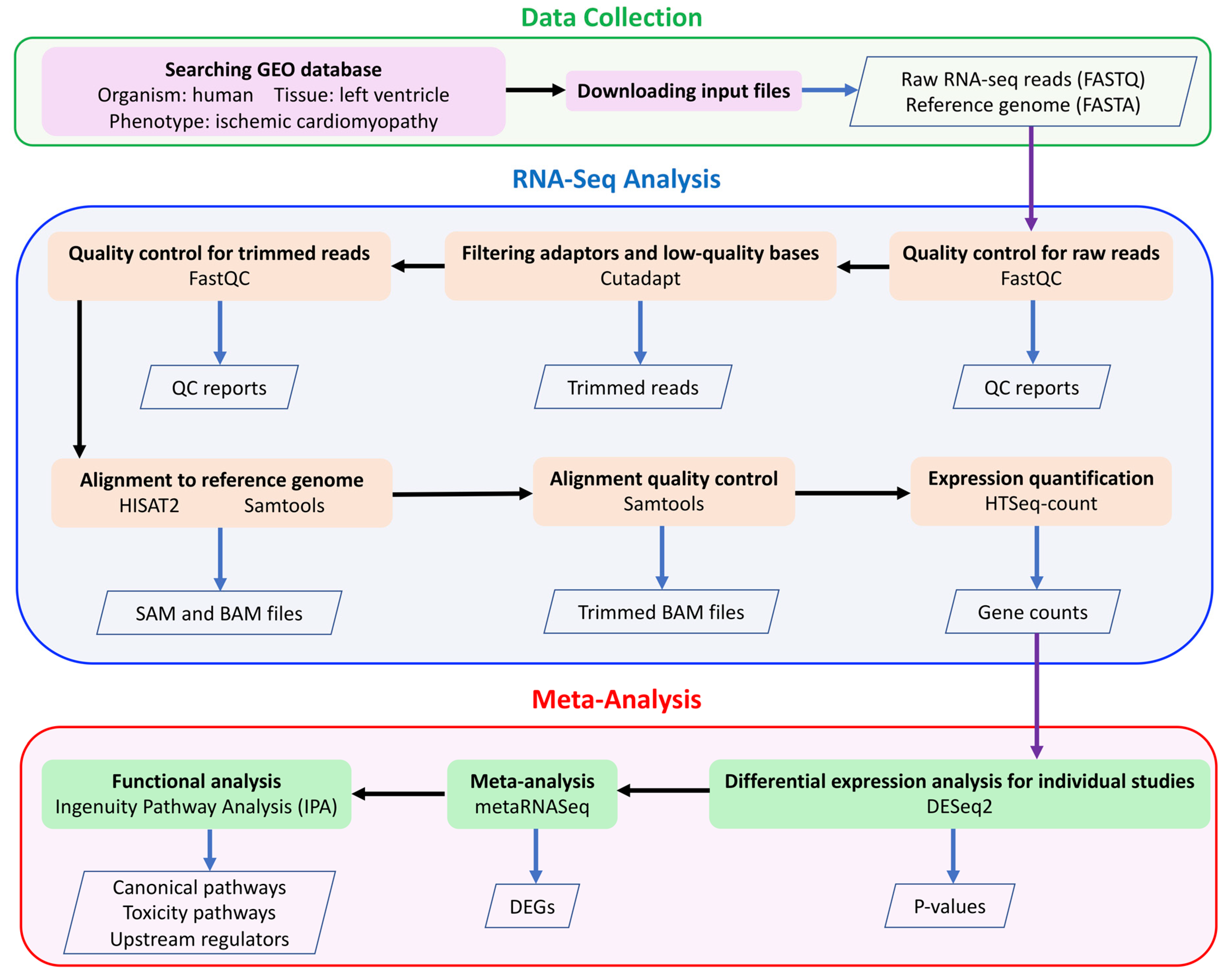
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roger, V.L. Epidemiology of heart failure. Circ. Res. 2013, 113, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.P. (Ed.) Manual of Cardiovascular Medicine, 4th ed.; Lippincott Williams & Wilkins, A Wolters Kluwer Business Company: Philadelphia, PA, USA, 2013; ISBN 978-1451131604. [Google Scholar]
- Kelkar, A.A.; Butler, J.; Schelbert, E.B.; Greene, S.J.; Quyyumi, A.A.; Bonow, R.O.; Cohen, I.; Gheorghiade, M.; Lipinski, M.J.; Sun, W. Mechanisms contributing to the progression of ischemic and nonischemic dilated cardiomyopathy: Possible modulating effects of paracrine activities of stem cells. J. Am. Coll. Cardiol. 2015, 66, 2038–2047. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.I.; Young, R.A. Transcriptional regulation and its misregulation in disease. Cell 2013, 152, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, K.; Nishikimi, T.; Nakao, K. Transcriptional regulation of the fetal cardiac gene program. J. Pharmacol. Sci. 2012, 119, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Dirkx, E.; da Costa Martins, P.A.; De Windt, L.J. Regulation of fetal gene expression in heart failure. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 2414–2424. [Google Scholar] [CrossRef] [Green Version]
- Wirrig, E.E.; Yutzey, K.E. Transcriptional regulation of heart valve development and disease. Cardiovasc. Pathol. 2011, 20, 162–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slagle, C.E.; Conlon, F.L. Emerging field of cardiomics: High-throughput investigations into transcriptional regulation of cardiovascular development and disease. Trends Genet. 2016, 32, 707–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rau, A.; Marot, G.; Jaffrézic, F. Differential meta-analysis of RNA-seq data from multiple studies. BMC Bioinform. 2014, 15, 91. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.; Hernandez, A.V.; Kattan, M.W. Meta-analysis: Its strengths and limitations. Cleve. Clin. J. Med. 2008, 75, 431. [Google Scholar] [CrossRef]
- Sweet, M.; Ambardekar, A.; Bristow, M.; Mestroni, L.; Taylor, M. RNA-seq of Heart Failure in Human Left Ventricles. In GEO Database; 2018. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116250 (accessed on 10 May 2019).
- Rau, C.; Tsai, E. Wipi1 Is a Genetic Hub That Mediates Right Ventricular Failure. In GEO Database; 2018. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE120852 (accessed on 10 May 2019).
- Yang, K.; Nerbonne, J. Deep RNA Sequencing Reveals Dynamic Regulation of Myocardial Noncoding RNA in Failing Human Heart and Remodeling with Mechanical Circulatory Support. In GEO Database; 2014. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE46224 (accessed on 10 May 2019).
- Wang, L.; Hu, Y.; Pu, W. RNA-seq Identifies Novel Transcript Elements and Transcript Processing in the Normal and Failing Hearts. In GEO Database; 2013. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE48166 (accessed on 10 May 2019).
- Van Aelst, L.N.L.; Voss, S.; Carai, P.; Van Leeuwen, R.; Vanhoutte, D.; Sanders-van Wijk, S.; Eurlings, L.; Swinnen, M.; Verheyen, F.K.; Verbeken, E. Osteoglycin prevents cardiac dilatation and dysfunction after myocardial infarction through infarct collagen strengthening. Circ. Res. 2015, 116, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Chen, S.; Ma, A.; Lu, J.; Wang, T. Identification of the difference in the pathogenesis in heart failure arising from different etiologies using a microarray dataset. Clinics 2017, 72, 600–608. [Google Scholar] [CrossRef]
- Pawlak, A.; Gil, R.J.; Grajkowska, W.; Nasierowska-Guttmejer, A.M.; Rzezak, J.; Kulawik, T. Significance of low desmin expression in cardiomyocytes in patients with idiopathic dilated cardiomyopathy. Am. J. Cardiol. 2013, 111, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Asakura, M.; Kitakaze, M. Global gene expression profiling in the failing myocardium. Circ. J. 2009, 73, 1568–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bare, L.A.; Morrison, A.C.; Rowland, C.M.; Shiffman, D.; Luke, M.M.; Iakoubova, O.A.; Kane, J.P.; Malloy, M.J.; Ellis, S.G.; Pankow, J.S. Five common gene variants identify elevated genetic risk for coronary heart disease. Genet. Med. 2007, 9, 682–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2015, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theis, J.L.; Zimmermann, M.T.; Evans, J.M.; Eckloff, B.W.; Wieben, E.D.; Qureshi, M.Y.; O’Leary, P.W.; Olson, T.M. Recessive MYH6 mutations in hypoplastic left heart with reduced ejection fraction. Circ. Cardiovasc. Genet. 2015, 8, 564–571. [Google Scholar] [CrossRef] [Green Version]
- Komamura, K.; Iwai, N.; Kokame, K.; Yasumura, Y.; Kim, J.; Yamagishi, M.; Morisaki, T.; Kimura, A.; Tomoike, H.; Kitakaze, M. The role of a common TNNT2 polymorphism in cardiac hypertrophy. J. Hum. Genet. 2004, 49, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Zhang, W.; Zhang, T. Association of single nucleotide polymorphisms in the 3′ UTR region of TPM1 gene with dilated cardiomyopathy: A case-control study. Medicine 2019, 98, e17710. [Google Scholar] [CrossRef]
- Hitz, M.-P.; Lemieux-Perreault, L.-P.; Marshall, C.; Feroz-Zada, Y.; Davies, R.; Yang, S.W.; Lionel, A.C.; D’Amours, G.; Lemyre, E.; Cullum, R. Rare copy number variants contribute to congenital left-sided heart disease. PLoS Genet. 2012, 8, e1002903. [Google Scholar] [CrossRef]
- BORLAK, J.; THUM, T. Hallmarks of ion channel gene expression in end-stage heart failure. FASEB J. 2003, 17, 1592–1608. [Google Scholar] [CrossRef]
- Hou, L.; Li, F.; Zeng, Q.; Su, L.; Chen, P.; Xu, Z.; Zhu, D.; Liu, C.; Xu, D. Excretion of urinary orosomucoid 1 protein is elevated in patients with chronic heart failure. PLoS ONE 2014, 9, e107550. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.; Azuaje, F. Identification of dilated cardiomyopathy signature genes through gene expression and network data integration. Genomics 2008, 92, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paavola, J.; Schliffke, S.; Rossetti, S.; Kuo, I.Y.-T.; Yuan, S.; Sun, Z.; Harris, P.C.; Torres, V.E.; Ehrlich, B.E. Polycystin-2 mutations lead to impaired calcium cycling in the heart and predispose to dilated cardiomyopathy. J. Mol. Cell. Cardiol. 2013, 58, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crackower, M.A.; Sarao, R.; Oudit, G.Y.; Yagil, C.; Kozieradzki, I.; Scanga, S.E.; Oliveira-dos-Santos, A.J.; da Costa, J.; Zhang, L.; Pei, Y. Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature 2002, 417, 822–828. [Google Scholar] [CrossRef]
- Herrer, I.; Roselló-Lletí, E.; Ortega, A.; Tarazón, E.; Molina-Navarro, M.M.; Triviño, J.C.; Martínez-Dolz, L.; Almenar, L.; Lago, F.; Sánchez-Lázaro, I. Gene expression network analysis reveals new transcriptional regulators as novel factors in human ischemic cardiomyopathy. BMC Med. Genomics 2015, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.U.; Neumann, J.; Schmitz, W. Transcriptional regulation by cAMP in the heart. Mol. Cell. Biochem. 2000, 212, 11–17. [Google Scholar] [CrossRef]
- Alimadadi, A.; Munroe, P.B.; Joe, B.; Cheng, X. Meta-Analysis of Dilated Cardiomyopathy Using Cardiac RNA-Seq Transcriptomic Datasets. Genes 2020, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Tur, M.K.; Etschmann, B.; Benz, A.; Leich, E.; Waller, C.; Schuh, K.; Rosenwald, A.; Ertl, G.; Kienitz, A.; Haaf, A.T. The 140-kD isoform of CD56 (NCAM1) directs the molecular pathogenesis of ischemic cardiomyopathy. Am. J. Pathol. 2013, 182, 1205–1218. [Google Scholar] [CrossRef]
- Schumann, H.; Holtz, J.; Zerkowski, H.-R.; Hatzfeld, M. Expression of secreted frizzled related proteins 3 and 4 in human ventricular myocardium correlates with apoptosis related gene expression. Cardiovasc. Res. 2000, 45, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Steimle, J.D.; Moskowitz, I.P. TBX5: A key regulator of heart development. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 122, pp. 195–221. ISBN 0070-2153. [Google Scholar]
- Ghosh, T.K.; Song, F.F.; Packham, E.A.; Buxton, S.; Robinson, T.E.; Ronksley, J.; Self, T.; Bonser, A.J.; Brook, J.D. Physical interaction between TBX5 and MEF2C is required for early heart development. Mol. Cell. Biol. 2009, 29, 2205–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.-J.; Song, K.; Luo, X.; Daniel, E.; Lambeth, K.; West, K.; Hill, J.A.; DiMaio, J.M.; Baker, L.A.; Bassel-Duby, R. Reprogramming of human fibroblasts toward a cardiac fate. Proc. Natl. Acad. Sci. USA 2013, 110, 5588–5593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Somma, S.; Di Benedetto, M.P.; Salvatore, G.; Agozzino, L.; Ferranti, F.; Esposito, S.; La Dogana, P.; Scarano, M.I.; Caputo, G.; Cotrufo, M. Desmin-free cardiomyocytes and myocardial dysfunction in end stage heart failure. Eur. J. Heart Fail. 2004, 6, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Mercadier, J.-J.; Lompre, A.-M.; Duc, P.; Boheler, K.R.; Fraysse, J.B.; Wisnewsky, C.; Allen, P.D.; Komajda, M.; Schwartz, K. Altered sarcoplasmic reticulum Ca2 (+)-ATPase gene expression in the human ventricle during end-stage heart failure. J. Clin. Invest. 1990, 85, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Gramolini, A.O.; Walsh, M.A.; Zhou, Y.-Q.; Slorach, C.; Friedberg, M.K.; Takeuchi, J.K.; Sun, H.; Henkelman, R.M.; Backx, P.H. Tbx5-dependent pathway regulating diastolic function in congenital heart disease. Proc. Natl. Acad. Sci. USA 2008, 105, 5519–5524. [Google Scholar] [CrossRef] [Green Version]
- Sveinbjornsson, G.; Olafsdottir, E.F.; Thorolfsdottir, R.B.; Davidsson, O.B.; Helgadottir, A.; Jonasdottir, A.; Jonasdottir, A.; Bjornsson, E.; Jensson, B.O.; Arnadottir, G.A. Variants in NKX2-5 and FLNC Cause Dilated Cardiomyopathy and Sudden Cardiac Death. Circ. Genomic Precis. Med. 2018, 11, e002151. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Zhang, J.; Du, Y.; Zhu, E.; Wang, Z.; Que, B.; Miao, H.; Shi, S.; Qin, X.; Zhao, Y. Human epicardial adipose tissue-derived and circulating secreted frizzled-related protein 4 (SFRP4) levels are increased in patients with coronary artery disease. Cardiovasc. Diabetol. 2017, 16, 133. [Google Scholar] [CrossRef] [Green Version]
- Carniel, E.; Taylor, M.R.G.; Sinagra, G.; Di Lenarda, A.; Ku, L.; Fain, P.R.; Boucek, M.M.; Cavanaugh, J.; Miocic, S.; Slavov, D. α-Myosin heavy chain: A sarcomeric gene associated with dilated and hypertrophic phenotypes of cardiomyopathy. Circulation 2005, 112, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Marian, A.J.; Braunwald, E. Hypertrophic cardiomyopathy: Genetics, pathogenesis, clinical manifestations, diagnosis, and therapy. Circ. Res. 2017, 121, 749–770. [Google Scholar] [CrossRef]
- Gigante, B.; Vikström, M.; Meuzelaar, L.S.; Chernogubova, E.; Silveira, A.; van’t Hooft, F.; Hamsten, A.; de Faire, U. Variants in the coagulation factor 2 receptor (F2R) gene influence the risk of myocardial infarction in men through an interaction with interleukin 6 serum levels. Thromb. Haemost. 2009, 101, 943–953. [Google Scholar]
- Gigante, B.; Bellis, A.; Visconti, R.; Marino, M.; Morisco, C.; Trimarco, V.; Galasso, G.; Piscione, F.; De Luca, N.; Prince, J.A. Retrospective analysis of coagulation factor II receptor (F2R) sequence variation and coronary heart disease in hypertensive patients. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1213–1219. [Google Scholar] [CrossRef] [Green Version]
- Tamamori-Adachi, M.; Takagi, H.; Hashimoto, K.; Goto, K.; Hidaka, T.; Koshimizu, U.; Yamada, K.; Goto, I.; Maejima, Y.; Isobe, M. Cardiomyocyte proliferation and protection against post-myocardial infarction heart failure by cyclin D1 and Skp2 ubiquitin ligase. Cardiovasc. Res. 2008, 80, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2012, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Sweet, M.E.; Cocciolo, A.; Slavov, D.; Jones, K.L.; Sweet, J.R.; Graw, S.L.; Reece, T.B.; Ambardekar, A.V.; Bristow, M.R.; Mestroni, L. Transcriptome analysis of human heart failure reveals dysregulated cell adhesion in dilated cardiomyopathy and activated immune pathways in ischemic heart failure. BMC Genom. 2018, 19, 812. [Google Scholar] [CrossRef] [Green Version]
- Tzimas, C.; Rau, C.D.; Buergisser, P.E.; Jean-Louis, G., Jr.; Lee, K.; Chukwuneke, J.; Dun, W.; Wang, Y.; Tsai, E.J. WIPI1 is a conserved mediator of right ventricular failure. JCI Insight 2019, 4, e122929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.-C.; Yamada, K.A.; Patel, A.Y.; Topkara, V.K.; George, I.; Cheema, F.H.; Ewald, G.A.; Mann, D.L.; Nerbonne, J.M. Deep RNA sequencing reveals dynamic regulation of myocardial noncoding RNAs in failing human heart and remodeling with mechanical circulatory support. Circulation 2014, 129, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, C.Y.; Li, Y.S.; Xu, J.; Li, D.G.; Li, X.; Han, D. Deep RNA sequencing elucidates microRNA-regulated molecular pathways in ischemic cardiomyopathy and nonischemic cardiomyopathy. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, L.; Zhang, S.; Zhang, C.; Huang, H.; Li, Y.; Hu, E.; Deng, G.; Guo, S.; Wang, Y. Identification of potential genes for human ischemic cardiomyopathy based on RNA-Seq data. Oncotarget 2016, 7, 82063. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects (accessed on 14 May 2019).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd: Edinburgh, Scotland, 1925; ISBN 0-05-002170-2. [Google Scholar]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. gplots: Various R Programming Tools for Plotting Data. R Package Version. 2015. Available online: https://cran.r-project.org/web/packages/gplots/index.html (accessed on 14 May 2019).
- Ohio Supercomputer Center; Ohio Supercomputer Center: Columbus, OH, USA, 1987; Available online: https://www.osc.edu (accessed on 15 June 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Dataset | Platform | Sample Size | Tissue |
|---|---|---|---|---|
| Study_1 | GSE116250 [11] | Illumina HiSeq 2500 | 13 ICM 14 NF | left ventricle |
| Study_2 | GSE120852 [12] | Illumina HiSeq 2500 | 5 ICM 5 NF | left ventricle |
| Study_3 | GSE46224 [13] | Illumina HiSeq 2000 | 8 ICM 8 NF | left ventricle |
| Study_4 | GSE48166 [14] | Illumina Genome Analyzer II | 15 ICM 15 NF | left ventricle |
| Ensembl_ID | Gene_Symbol | Adj_P 1 | Average_Log2FC 2 | Effect 3 |
|---|---|---|---|---|
| ENSG00000008311 | AASS | 0 | −1.03 | Down |
| ENSG00000075413 | MARK3 | 0 | −0.88 | Down |
| ENSG00000076351 | SLC46A1 | 0 | 0.48 | Up |
| ENSG00000100196 | KDELR3 | 0 | 0.72 | Up |
| ENSG00000103415 | HMOX2 | 0 | −0.81 | Down |
| ENSG00000105894 | PTN | 0 | 1.47 | Up |
| ENSG00000106809 | OGN | 0 | 2.26 | Up |
| ENSG00000106819 | ASPN | 0 | 1.99 | Up |
| ENSG00000106823 | ECM2 | 0 | 1.17 | Up |
| ENSG00000118194 | TNNT2 | 0 | −0.55 | Down |
| ENSG00000122034 | GTF3A | 0 | −0.54 | Down |
| ENSG00000123689 | G0S2 | 0 | −1.57 | Down |
| ENSG00000126106 | TMEM53 | 0 | −0.58 | Down |
| ENSG00000129250 | KIF1C | 0 | −0.56 | Down |
| ENSG00000130528 | HRC | 0 | −0.58 | Down |
| ENSG00000139329 | LUM | 0 | 1.81 | Up |
| ENSG00000140416 | TPM1 | 0 | −0.53 | Down |
| ENSG00000141905 | NFIC | 0 | −0.57 | Down |
| ENSG00000145934 | TENM2 | 0 | −0.67 | Down |
| ENSG00000156219 | ART3 | 0 | −1.31 | Down |
| ENSG00000161970 | RPL26 | 0 | −0.76 | Down |
| ENSG00000175084 | DES | 0 | −0.80 | Down |
| ENSG00000176293 | ZNF135 | 0 | 0.51 | Up |
| ENSG00000179526 | SHARPIN | 0 | −0.34 | Down |
| ENSG00000197256 | KANK2 | 0 | −0.57 | Down |
| ENSG00000197616 | MYH6 | 0 | −2.59 | Down |
| ENSG00000198542 | ITGBL1 | 0 | 1.69 | Up |
| ENSG00000198624 | CCDC69 | 0 | −0.87 | Down |
| ENSG00000210127 | MT-TA | 0 | −1.66 | Down |
| ENSG00000233098 | CCDC144NL-AS1 | 0 | 1.17 | Up |
| ENSG00000140319 | SRP14 | 6.91 ×10−14 | −0.53 | Down |
| ENSG00000197586 | ENTPD6 | 6.91 × 10−14 | −0.77 | Down |
| ENSG00000267280 | TBX2-AS1 | 6.91 × 10−14 | 0.83 | Up |
| ENSG00000152086 | TUBA3E | 1.33 × 10−13 | −1.99 | Down |
| ENSG00000170448 | NFXL1 | 1.33 × 10−13 | −1.84 | Down |
| ENSG00000165124 | SVEP1 | 1.89 × 10−13 | 1.24 | Up |
| ENSG00000152580 | IGSF10 | 2.46 × 10−13 | 1.54 | Up |
| ENSG00000143603 | KCNN3 | 3.04 × 10−13 | 1.43 | Up |
| ENSG00000187837 | HIST1H1C | 3.60 × 10−13 | −0.89 | Down |
| ENSG00000075886 | TUBA3D | 4.15 × 10−13 | −1.60 | Down |
| ENSG00000189060 | H1F0 | 8.87 × 10−13 | −0.80 | Down |
| ENSG00000134247 | PTGFRN | 2.21 × 10−12 | 0.93 | Up |
| ENSG00000116690 | PRG4 | 3.94 × 10−12 | 0.84 | Up |
| ENSG00000160392 | C19orf47 | 4.61 × 10−12 | −0.95 | Down |
| ENSG00000129009 | ISLR | 7.14 × 10−12 | 1.49 | Up |
| ENSG00000129116 | PALLD | 8.77 × 10−12 | −0.72 | Down |
| ENSG00000173991 | TCAP | 1.11 × 10−11 | −0.55 | Down |
| ENSG00000104879 | CKM | 1.27 × 10−11 | −0.79 | Down |
| ENSG00000108298 | RPL19 | 1.66 × 10−11 | −0.54 | Down |
| ENSG00000142748 | FCN3 | 1.75 × 10−11 | −1.59 | Down |
| Ensembl_ID | Gene_Symbol | Adj_P 1 | Average_Log2FC 2 | Effect 3 |
|---|---|---|---|---|
| ENSG00000171517 | LPAR3 | 8.64 × 10−4 | −1.16 | Down |
| ENSG00000178607 | ERN1 | 2.24× 10−3 | 0.88 | Up |
| ENSG00000048707 | VPS13D | 3.17 × 10−3 | −0.60 | Down |
| ENSG00000179604 | CDC42EP4 | 3.18 × 10−3 | −0.66 | Down |
| ENSG00000072832 | CRMP1 | 3.42 × 10−3 | 0.75 | Up |
| ENSG00000162458 | FBLIM1 | 6.49 × 10−3 | −0.79 | Down |
| ENSG00000178307 | TMEM11 | 6.86 × 10−3 | −0.49 | Down |
| ENSG00000166278 | C2 | 7.49 × 10−3 | 1.02 | Up |
| ENSG00000228526 | MIR34AHG | 8.17 × 10−3 | 0.97 | Up |
| ENSG00000184007 | PTP4A2 | 8.29 × 10−3 | −0.40 | Down |
| ENSG00000160818 | GPATCH4 | 8.45 × 10−3 | −0.48 | Down |
| ENSG00000100949 | RABGGTA | 8.63 × 10−3 | −0.38 | Down |
| ENSG00000255248 | MIR100HG | 9.29 × 10−3 | 0.35 | Up |
| ENSG00000165028 | NIPSNAP3B | 9.48 × 10−3 | −0.52 | Down |
| ENSG00000133678 | TMEM254 | 9.98 × 10−3 | 0.63 | Up |
| ENSG00000128272 | ATF4 | 1.02 × 10−2 | −0.55 | Down |
| ENSG00000103342 | GSPT1 | 1.13 × 10−2 | −0.33 | Down |
| ENSG00000163866 | SMIM12 | 1.27 × 10−2 | −0.46 | Down |
| ENSG00000198355 | PIM3 | 1.29 × 10−2 | −0.61 | Down |
| ENSG00000163399 | ATP1A1 | 1.32 × 10−2 | −0.47 | Down |
| ENSG00000160862 | AZGP1 | 1.33 × 10−2 | −0.85 | Down |
| ENSG00000180758 | GPR157 | 1.42 × 10−2 | −0.73 | Down |
| ENSG00000115461 | IGFBP5 | 1.53 × 10−2 | 0.48 | Up |
| ENSG00000037280 | FLT4 | 1.76 × 10−2 | 0.48 | Up |
| ENSG00000135272 | MDFIC | 1.78 × 10−2 | 0.69 | Up |
| ENSG00000131781 | FMO5 | 1.83 × 10−2 | −0.73 | Down |
| ENSG00000184887 | BTBD6 | 1.93 × 10−2 | −0.53 | Down |
| ENSG00000142494 | SLC47A1 | 2.39 × 10−2 | 0.58 | Up |
| ENSG00000113811 | SELENOK | 2.63 × 10−2 | −0.34 | Down |
| ENSG00000186567 | CEACAM19 | 2.63 × 10−2 | −0.60 | Down |
| ENSG00000100767 | PAPLN | 2.66 × 10−2 | 0.97 | Up |
| ENSG00000159674 | SPON2 | 2.85 × 10−2 | 0.62 | Up |
| ENSG00000169155 | ZBTB43 | 2.95 × 10−2 | −0.40 | Down |
| ENSG00000103034 | NDRG4 | 3.09 × 10−2 | −0.34 | Down |
| ENSG00000106034 | CPED1 | 3.11 × 10−2 | 0.37 | Up |
| ENSG00000179262 | RAD23A | 3.28 × 10−2 | −0.32 | Down |
| ENSG00000169718 | DUS1L | 3.31 × 10−2 | −0.37 | Down |
| ENSG00000107736 | CDH23 | 3.35 × 10−2 | 0.71 | Up |
| ENSG00000108883 | EFTUD2 | 3.43 × 10−2 | −0.26 | Down |
| ENSG00000139990 | DCAF5 | 3.45 × 10−2 | −0.34 | Down |
| ENSG00000019995 | ZRANB1 | 3.49 × 10−2 | −0.24 | Down |
| ENSG00000160877 | NACC1 | 3.49 × 10−2 | −0.50 | Down |
| ENSG00000175602 | CCDC85B | 3.51 × 10−2 | 0.56 | Up |
| ENSG00000143869 | GDF7 | 3.65 × 10−2 | 0.76 | Up |
| ENSG00000182287 | AP1S2 | 3.90 × 10−2 | −0.34 | Down |
| ENSG00000114670 | NEK11 | 3.94 × 10−2 | 0.77 | Up |
| ENSG00000197977 | ELOVL2 | 3.96 × 10−2 | −0.71 | Down |
| ENSG00000170004 | CHD3 | 4.20 × 10−2 | 0.48 | Up |
| ENSG00000168615 | ADAM9 | 4.38 × 10−2 | −0.39 | Down |
| ENSG00000118762 | PKD2 | 4.42 × 10−2 | 0.32 | Up |
| ENSG00000078061 | ARAF | 4.54 × 10−2 | −0.27 | Down |
| ENSG00000113140 | SPARC | 4.57 × 10−2 | 0.49 | Up |
| ENSG00000144645 | OSBPL10 | 4.62 × 10−2 | 0.85 | Up |
| ENSG00000003096 | KLHL13 | 4.99 × 10−2 | 0.83 | Up |
| Ensembl_ID | Gene_Symbol | Average_Log2FC 1 | Effect 2 |
|---|---|---|---|
| ENSG00000120907 | ADRA1A | −0.67 | Down |
| ENSG00000163399 | ATP1A1 | −0.47 | Down |
| ENSG00000174437 | ATP2A2 | −0.74 | Down |
| ENSG00000007402 | CACNA2D2 | 0.59 | UP |
| ENSG00000175084 | DES | −0.80 | Down |
| ENSG00000126218 | F10 | 0.54 | Up |
| ENSG00000152642 | GPD1L | −0.75 | Down |
| ENSG00000171385 | KCND3 | −0.91 | Down |
| ENSG00000055118 | KCNH2 | −0.84 | Down |
| ENSG00000187486 | KCNJ11 | −1.03 | Down |
| ENSG00000134571 | MYBPC3 | −0.55 | Down |
| ENSG00000092054 | MYH7 | −0.45 | Down |
| ENSG00000175206 | NPPA | 1.86 | UP |
| ENSG00000104368 | PLAT | 0.64 | Up |
| ENSG00000183873 | SCN5A | −0.54 | Down |
| ENSG00000118194 | TNNT2 | −0.55 | Down |
| ENSG00000155657 | TTN | −0.66 | Down |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alimadadi, A.; Aryal, S.; Manandhar, I.; Joe, B.; Cheng, X. Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 3472. https://doi.org/10.3390/ijms21103472
Alimadadi A, Aryal S, Manandhar I, Joe B, Cheng X. Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis. International Journal of Molecular Sciences. 2020; 21(10):3472. https://doi.org/10.3390/ijms21103472
Chicago/Turabian StyleAlimadadi, Ahmad, Sachin Aryal, Ishan Manandhar, Bina Joe, and Xi Cheng. 2020. "Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis" International Journal of Molecular Sciences 21, no. 10: 3472. https://doi.org/10.3390/ijms21103472
APA StyleAlimadadi, A., Aryal, S., Manandhar, I., Joe, B., & Cheng, X. (2020). Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis. International Journal of Molecular Sciences, 21(10), 3472. https://doi.org/10.3390/ijms21103472