Comparative Transcriptomics of Rice Genotypes with Contrasting Responses to Nitrogen Stress Reveals Genes Influencing Nitrogen Uptake through the Regulation of Root Architecture



Abstract
:1. Introduction
2. Results
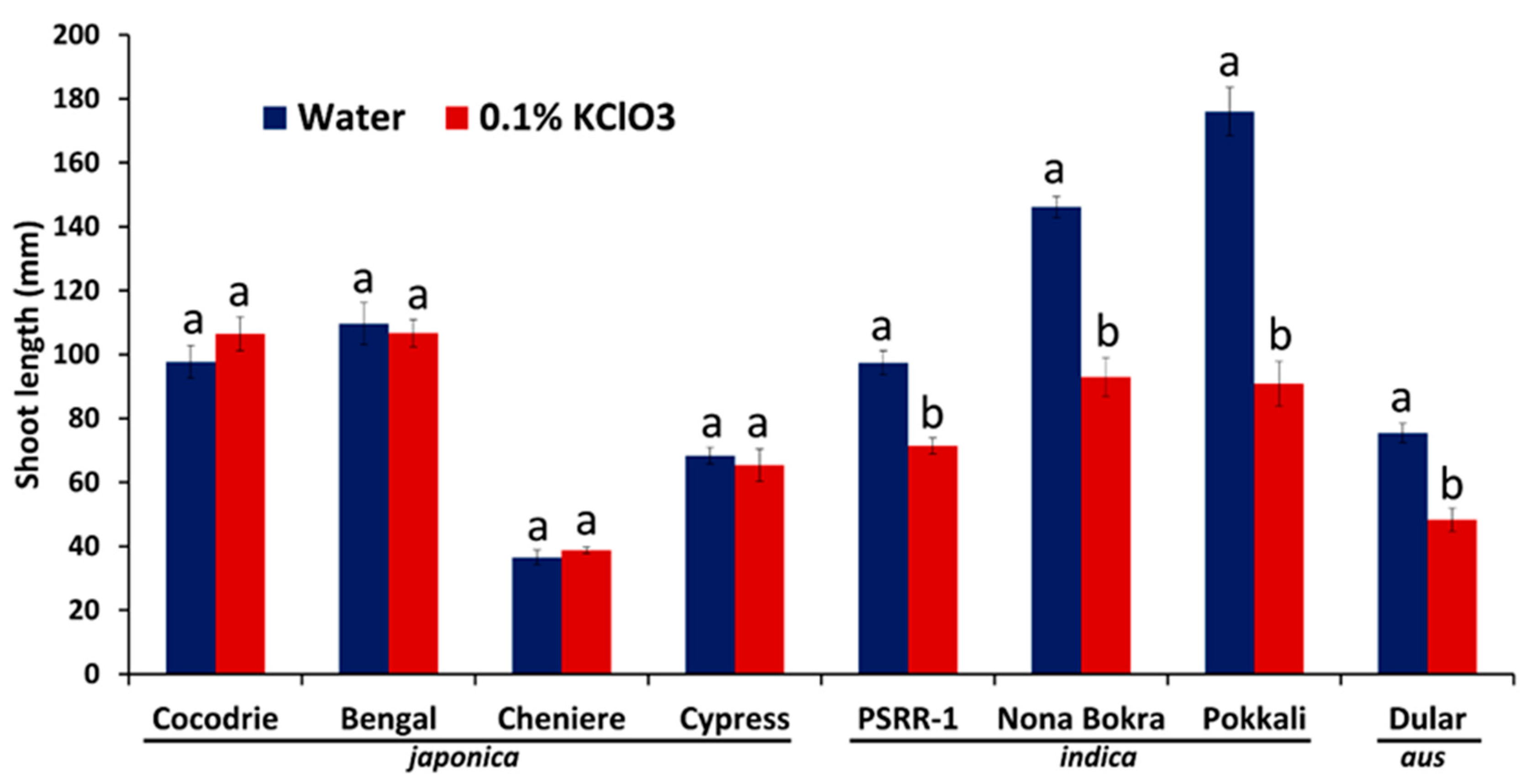
2.1. Chlorate Experiment
2.2. Nitrogen Stress Experiment
2.3. Transcriptome Analysis under Nitrogen Stress
2.4. Gene Ontology (GO) and Pathway Analysis of Differentially Expressed Genes under Nitrogen Stress
2.5. Expression Pattern of Nitrogen Utilization and Long Distance Signaling
2.6. Transcription Factors and Signaling for Nitrogen Utilization and Root Growth and Development Genes
2.7. Phytohormones Related to N Stress and Root Growth and Development
2.8. Differentially Expressed Genes Overlapping the Nitrogen Stress-Related QTL
2.9. Alternate Splicing Events under Nitrogen Stress
2.10. Gene Expression Validation of Selected Genes by Quantitative Reverse Transcription PCR (qRT-PCR)
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Cultivation
4.2. Chlorate Assay
4.3. Nitrogen Stress Response Experiment
4.4. Hydroponic Experiment for RNA-Seq
4.5. RNA Isolation, Library Preparation, and Sequencing
4.6. Reference Genome-Based Mapping
4.7. Transcript Assembly and Analysis of Alternative Splicing Events
4.8. Differential Expression, Genotype and Treatment Comparison, and Ontology
4.9. Transcription Factors among DEGs
4.10. DEGs Co-Localized in Root Development-Related, ChloratE-Resistance, and NUE QTLs
4.11. Validation of Expression of NUE-Related Genes: qRT-PCR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| N | Nitrogen |
| NUE | Nitrogen use efficiency |
| DEG | Differentially expressed genes |
| QTL | Quantitative trait loci |
| TF | Transcription factor |
| PK | Pokkali |
| BG | Bengal |
| PKFN | Pokkali-Full nitrogen |
| PKLN | Pokkali-low nitrogen |
| PK1H | Pokkali-1h after transfer from low N to full N |
| BGFN | Bengal-full nitrogen |
| BGLN | Bengal-low nitrogen |
| BG1H | Bengal-1h after transfer from low N to full N |
| GO | Gene ontology |
| SEA | Singular enrichment analysis |
| CEP | C-terminally encoded peptides |
| CLE | CLV3/ESR-related |
| rtnb | Root number |
| rtlg | Root length |
| rtth | Root thickness |
| rtvol | Root volume |
| rtdp | Root depth |
| kclo3rs | Potassium chlorate resistance |
| AS | Alternate splicing |
| IR | Intron retention |
| A3SS | Alternative 3′ splice site |
| ES | Exon skipping |
| A5SS | Alternative 5′ splice site |
| MXE | Mutually exclusive exon |
| GOGAT | Glutamine oxoglutarate aminotransferase |
| qRT-PCR | Quantitative reverse transcription PCR |
| NIGT1 | Nitrate-Inducible, GARP-type Transcriptional Repressor 1 |
| GID1 | GA insensitive dwarf1 |
| NGR5 | Nitrogen-mediated tiller growth response 5 |
| PRC2 | Polycomb repressive complex 2 |
References
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, E. Nitrogen pollution knows no bounds. Science 2017, 356, 700–701. [Google Scholar] [CrossRef] [PubMed]
- Golden, B.R.; Slaton, N.A.; Norman, R.J.; Wilson, C.E., Jr.; DeLong, R.E. Evaluation of polymer-coated urea for direct-seeded, delayed-flood rice production. Soil Sci. Soc. Am. J. 2009, 73, 375–383. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.; Coruzzi, G.M.; et al. Nitrate 2020: Thirty years from transport to signaling networks. Plant Cell 2020. [Google Scholar] [CrossRef]
- International Rice Genome Sequencing Project (IRGSP). The map-based sequence of the rice genome. Nature 2005, 436, 793–800. [Google Scholar] [CrossRef]
- McCouch, S.; Zhao, K.; Wright, M.; Tung, C.W.; Ebana, K.; Thomson, M.; Reynolds, A.; Wang, D.; DeClerck, G.; Ali, L.; et al. Development of genome-wide SNP assays for rice. Breed. Sci. 2010, 60, 524–535. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y.; et al. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef]
- Ta, T.C.; Ohira, K. Effects of various environmental and medium conditions on the response of indica and japonica rice plants to ammonium and nitrate nitrogen. Soil Sci. Plant Nutr. 1981, 27, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Ta, T.; Tsutsumi, M.; Kurihara, K. Comparative study on the response of indica and japonica rice plants to ammonium and nitrate nitrogen. Soil Sci. Plant Nutr. 1981, 27, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Lin, H.X. Nitrogen-use efficiency: Transport solution in rice variations. Nat. Plants 2015, 1, 15096. [Google Scholar] [CrossRef] [PubMed]
- Good, A.G.; Johnson, S.J.; De Pauw, M.; Carroll, R.T.; Savidov, N. Engineering nitrogen use efficiency with alanine aminotransferase. Can. J. Bot. 2007, 85, 252–262. [Google Scholar] [CrossRef]
- Lea, P.J.; Azevedo, R.A. Nitrogen use efficiency. 2. Amino acid metabolism. Annal. Appl. Biol. 2007, 151, 269–275. [Google Scholar] [CrossRef]
- Pathak, R.R.; Ahmad, A.; Lochab, S.; Raghuram, N. Molecular physiology of plant nitrogen use efficiency and biotechnological options for its enhancement. Curr. Sci. 2008, 94, 1394–1403. [Google Scholar]
- Shrawat, A.K.; Carroll, R.T.; DePauw, M.; Taylor, G.J.; Good, A.G. Genetic engineering of improved nitrogen use efficiency in rice by the tissue-specific expression of alanine aminotransferase. Plant Biotech. J. 2008, 6, 722–732. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Yang, S.; Hao, D.; Song, Z.; Yang, G.; Wang, L.; Su, Y. RNA-Seq analysis of differentially expressed genes in rice under varied nitrogen supplies. Gene 2015, 555, 305–317. [Google Scholar] [CrossRef]
- Yang, W.; Yoon, J.; Choi, H.; Fan, Y.; Chen, R.; An, G. Transcriptome analysis of nitrogen-starvation-responsive genes in rice. BMC Plant Biol. 2015, 15, 31. [Google Scholar] [CrossRef] [Green Version]
- Xin, W.; Zhang, L.; Zhang, W.; Gao, J.; Yi, J.; Zhen, X.; Li, Z.; Zhao, Y.; Peng, C.; Zhao, C. An integrated analysis of the rice transcriptome and metabolome reveals differential regulation of carbon and nitrogen metabolism in response to nitrogen availability. Int. J. Mol. Sci. 2019, 20, 2349. [Google Scholar] [CrossRef] [Green Version]
- Sinha, S.K.; Sevanthi, V.A.M.; Chaudhary, S.; Tyagi, P.; Venkadesan, S.; Rani, M.; Mandal, P.K. Transcriptome analysis of two rice varieties contrasting for nitrogen use efficiency under chronic N starvation reveals differences in chloroplast and starch metabolism-related genes. Genes 2018, 9, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhou, J.; Huang, N.; Mo, L.; Lv, M.; Gao, Y.; Chen, C.; Yin, S.; Ju, J.; Dong, G.; et al. Transcriptomic and co-expression network profiling of shoot apical meristem reveal contrasting response to nitrogen rate between indica and japonica rice subspecies. Int. J. Mol. Sci. 2019, 20, 5922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaya, T.; Obara, M.; Nakajima, H.; Sasaki, S.; Hayakawa, T.; Sato, T. Genetic manipulation and quantitative-trait loci mapping for nitrogen recycling in rice. J. Exp. Bot. 2002, 53, 917–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quraishi, U.M.; Abrouk, M.; Murat, F.; Pont, C.; Foucrier, S.; Desmaizieres, G.; Confolent, C.; Rivière, N.; Charmet, G.; Paux, E.; et al. Cross-genome map based dissection of a nitrogen use efficiency ortho-meta QTL in bread wheat unravels concerted cereal genome evolution. Plant J. 2011, 65, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Tang, H.; Liu, H.; Shunzong, S.; Zhang, S.; Wu, L.; Liu, D.; Gao, S. Mining for low-nitrogen tolerance genes by integrating meta-analysis and large-scale gene expression data from maize. Euphytica 2015, 206, 117–131. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, X.; Wu, K.; Fu, X. Nitrogen signaling and use efficiency in plants: What’s new? Curr. Opin. Plant Biol. 2015, 27, 192–198. [Google Scholar] [CrossRef]
- Forde, B. Nitrogen signaling pathways shaping root system architecture: An update. Curr. Opin. Plant Biol. 2014, 21, 30–36. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Ruan, B.; Liu, C.; Jiang, H.; Dong, G.; et al. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10, 5207. [Google Scholar] [CrossRef]
- Manneh, B.; Stam, P.; Struik, P.C.; Bruce-Oliver, S.; van Eeuwijk, F.A. QTL-based analysis of genotype-by-environment interaction for grain yield of rice in stress and non-stress environments. Euphytica 2007, 156, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S.; Valencia, M.O.; Ishitani, M.; Selvaraj, M.G. Root system architecture variation in response to different NH4+ concentrations and its association with nitrogen-deficient tolerance traits in rice. Acta Physiol. Plant. 2014, 36, 2361–2372. [Google Scholar] [CrossRef]
- Chrispeels, H.E.; Oettinger, H.; Janvier, N.; Tague, B.W. AtZFP1, encoding Arabidopsis thaliana C2H2 zinc-finger protein 1, is expressed downstream of photomorphogenic activation. Plant Mol. Biol. 2000, 42, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Krouk, G. Hormones and nitrate: A two-way connection. Plant Mol. Biol. 2016, 91, 599–606. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Coruzzi, G.; Gutierrez, R.A. Nitrate transport, sensing and responses in plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Fredes, I.; Moreno, S.; Diaz, F.P.; Gutierrez, R.A. Nitrate signaling and the control of Arabidopsis growth and development. Curr. Opin. Plant Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef]
- Sui, Z.; Wang, T.; Li, H.; Zhang, M.; Li, Y.; Xu, R.; Xing, G.; Ni, Z.; Xin, M. Overexpression of peptide-encoding OsCEP6.1 results in pleiotropic effects on growth in rice (O. sativa). Front. Plant Sci. 2016, 7, 228. [Google Scholar] [CrossRef] [Green Version]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Katayama, H.; Mori, M.; Kawamura, Y.; Tanaka, T.; Mori, M.; Hasegawa, H. Production and characterization of transgenic rice plants carrying a high-affinity nitrate transporter gene (OsNRT2.1). Breed. Sci. 2009, 59, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Naz, M.; Luo, B.; Guo, X.; Li, B.; Chen, J.; Fan, X. Overexpression of nitrate transporter OsNRT2.1 enhances nitrate-dependent root elongation. Genes 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, B.; Chen, J.; Zhu, L.; Liu, S.; Li, B.; Lu, H.; Ye, G.; Xu, G.; Fan, X. Overexpression of a high-affinity nitrate transporter OsNRT2.1 increases yield and manganese accumulation in rice under alternating wet and dry condition. Front. Plant Sci. 2018, 9, 1192. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Qi, T.; Hu, Z.; Fan, X.; Zhu, L.; Iqbal, M.F.; Yin, X.; Xu, G.; Fan, X. OsNAR2.1 positively regulates drought tolerance and grain yield under drought stress conditions in rice. Front. Plant Sci. 2019, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.H.; Kohler, B.; Cao, F.Q.; Liu, G.W.; Gong, Y.Y.; Sheng, S.; Song, Q.C.; Cheng, X.Y.; Garnett, T.; Okamoto, M.; et al. Rice DUR3 mediates high-affinity urea transport and plays an effective role in improvement of urea acquisition and utilization when expressed in Arabidopsis. New Phytol. 2012, 193, 432–444. [Google Scholar] [CrossRef]
- Beier, M.P.; Fujita, T.; Sasaki, K.; Kanno, K.; Ohashi, M.; Tamura, W.; Konishi, N.; Saito, M.; Imagawa, F.; Ishiyama, K.; et al. The urea transporter DUR3 contributes to rice production under nitrogen-deficient and field conditions. Physiol. Plant. 2019, 167, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Sawaki, N.; Tsujimoto, R.; Shigyo, M.; Konishi, M.; Toki, S.; Fujiwara, T.; Yanagisawa, S. A nitrate-inducible GARP family gene encodes an auto-repressible transcriptional repressor in rice. Plant Cell Physiol. 2013, 54, 506–517. [Google Scholar] [CrossRef]
- Wu, K.; Wang, S.; Song, W.; Zhang, J.; Wang, Y.; Liu, Q.; Yu, J.; Ye, Y.; Li, S.; Chen, J.; et al. Enhanced sustainable green revolution yield via nitrogen-responsive chromatin modulation in rice. Science 2020, 367, eaaz2046. [Google Scholar] [CrossRef]
- Maeda, Y.; Konishi, M.; Kiba, T.; Sakuraba, Y.; Sawaki, N. Molecular interactions of a soluble gibberellin receptor, GID1, with a rice DELLA protein, SLR1, and Gibberellin. Nat Commun. 2018, 9, 1376. [Google Scholar] [CrossRef] [Green Version]
- Shankar, R.; Bhattacharjee, A.; Jain, M. Transcriptome analysis in different rice cultivars provides novel insights into desiccation and salinity stress responses. Sci. Rep. 2016, 6, 23719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansuri, R.M.; Shobbar, Z.S.; Jelodar, N.B.; Ghaffari, M.R.; Nematzadeh, G.A.; Asari, S. Dissecting molecular mechanisms underlying salt tolerance in rice: A comparative transcriptional profiling of the contrasting genotypes. Rice 2019, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Ashwini, N.; Sajeevan, R.S.; Udayakumar, M.; Nataraja, K.N. Identification of splice variant of OsGBF1 in Oryza sativa ssp. indica genotypes under salinity stress. 3 Biotech 2018, 8, 345. [Google Scholar] [PubMed]
- Sellamuthu, G.; Jegadeeson, V.; Sajeevan, R.; Rajakani, R.; Parthasarathy, P.; Raju, K.; Shabala, L.; Chen, Z.; Zhou, M.; Sowdhamini, R.; et al. Distinct evolutionary origins of intron retention splicing events in NHX1 antiporter transcripts relate to sequence specific distinctions in Oryza Species. Front. Plant Sci. 2020, 11, 267. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Pujar, A.; Youens-Clark, K.; Yap, I.; Jaiswal, P.; Tecle, I.; Tung, C.W.; Ren, L.; Spooner, W.; Wei, X.; et al. Gramene QTL database: Development, content and applications. Database (Oxford) 2009. [Google Scholar] [CrossRef]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 20, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Cai, W. OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance. PLoS ONE 2012, 7, e45117. [Google Scholar] [CrossRef] [Green Version]
- Senthilvel, S.; Vinod, K.K.; Malarvizhi, M.; Maheswaran, M. QTL and QTL × environment effects on agronomic and nitrogen acquisition traits in rice. J. Integr. Plant Biol. 2008, 50, 1108–1117. [Google Scholar] [CrossRef] [Green Version]
- Tobias, C.M.; Chow, E.K. Structure of the cinnamyl-alcohol dehydrogenase gene family in rice and promoter activity of a member associated with lignification. Planta 2005, 220, 678–688. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Xiao, G.; Zhai, M.; Pan, X.; Huang, R.; Zhang, H. CYP71D8L is a key regulator involved in growth and stress responses by mediating gibberellin homeostasis in rice. J. Exp. Bot. 2020, 71, 1160–1170. [Google Scholar]
- Lee, D.S.; Kim, B.K.; Kwon, S.J.; Jin, H.C.; Park, O.K. Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem. Biophys. Res. Commun. 2009, 379, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.Á.; Forment, J.; Roldán, M.; Serrano, R.; Vicente, O. Overexpression of Arabidopsis thaliana LTL1, a salt-induced gene encoding a GDSL-motif lipase, increases salt tolerance in yeast and transgenic plants. Plant Cell Environ. 2006, 29, 1890–1900. [Google Scholar] [CrossRef] [PubMed]
- De Datta, S.K.; Chang, T.T.; Yoshida, S. Drought tolerance in upland rice. In Major Research in Upland Rice; International Rice Research Institute: Los Baños, Philippines, 1975; pp. 101–116. [Google Scholar]
- Senadhira, D.; Neue, H.U.; Akbar, M. Development of improved donors for salinity tolerance in rice through somaclonal variation. SABRAO J. 1994, 26, 19–25. [Google Scholar]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef]
- Subudhi, P.K.; Parco, A.; Singh, P.K.; Deleon, T.; Karan, R.; Biradar, H.; Cohn, M.A.; Brar, D.S.; Sasaki, T. Genetic architecture of seed dormancy in US weedy rice in different genetic backgrounds. Crop Sci. 2012, 52, 2564–2575. [Google Scholar] [CrossRef]
- Chai, C.; Shankar, R.; Jain, M.; Subudhi, P.K. Genome-wide discovery of DNA polymorphisms by whole genome sequencing differentiates weedy and cultivated rice. Sci. Rep. 2018, 8, 14218. [Google Scholar] [CrossRef]
- Teng, S.; Tian, C.; Chen, M.; Zeng, D.; Guo, L.; Zhu, L.; Han, B.; Qian, Q. QTLs and candidate genes for chlorate resistance in rice (Oryza sativa L). Euphytica 2006, 152, 141–148. [Google Scholar] [CrossRef]
- Lobet, G.; Pagès, L.; Draye, X. A novel image-analysis toolbox enabling quantitative analysis of root system architecture. Plant Physiol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast-spliced aligner with 192low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foissac, S.; Sammeth, M. ASTALAVISTA: Dynamic and flexible analysis of alternative splicing events in custom gene datasets. Nucl. Acids Res. 2007, 35, W297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sammeth, M.; Foissac, S.; Guigo, R. A general definition and nomenclature for alternative splicing events. PLoS Comput. Biol. 2008, 4, e1000147. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucl. Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Naithani, S.; Gupta, P.; Preece, J.; D’Eustachio, P.; Elser, J.; Garg, P.; Dikeman, D.A.; Kiff, J.; Cook, J.; Olson, A.; et al. Plant Reactome: A knowledgebase and resource for comparative pathway analysis. Nucl. Acids Res. 2019. [Google Scholar] [CrossRef]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucl. Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Cui, K.; Ye, G.; Pan, J.; Xiang, J.; Huang, J.; Nie, L. QTL mapping for nitrogen-use efficiency and nitrogen-deficiency tolerance traits in rice. Plant Soil 2012, 359, 281–295. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karan, R.; Subudhi, P.K. A stress inducible SUMO conjugating enzyme gene of a grass halophyte Spartina alterniflora (SaSce9) enhances salinity and drought stress tolerance in Arabidopsis. BMC Plant Biol. 2012, 12, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Average Lateral Root Length (cm) | Average Lateral Root Diameter (cm) | Total Lateral Roots (range) | Total Surface Area (cm2) | Total Volume (cm3) |
|---|---|---|---|---|---|---|
| Bengal | Full N | 8.20 ± 0.29 a,b | 0.029 ± 0.003 a | 5–8 | 4.19 ± 0.23 a | 0.036 ± 0.003 a |
| Low N | 9.09 ± 0.56 a | 0.032 ± 0.003 a | 7–9 | 7.40 ± 0.13 b | 0.089 ± 0.003 b | |
| Pokkali | Full N | 7.61 ± 0.40 b | 0.039 ± 0.001 b | 6–10 | 5.30 ± 0.33 c | 0.054 ± 0.006 c |
| Low N | 11.60 ± 0.46 c | 0.036 ± 0.001 a,b | 8–13 | 9.67 ± 0.45 d | 0.107 ± 0.006 d |
| Major Pathway | Pathway Name | Low N (%) a | 1-h after Full N (%) b |
|---|---|---|---|
| Cellular process | DNA replication: activation of the pre-replicative complex | - | 1.4 |
| Protein metabolism: translation | 3.0 | 7.1 | |
| Growth and developmental process | Reproductive structure development | 5.0 | 5.7 |
| Vegetative structure development | 3.0 | - | |
| Metabolism and regulation | Amine and polyamine biosynthesis | 1.0 | - |
| Amino acid metabolism | 13.0 | 18.6 | |
| Carbohydrate metabolism | 8.0 | 5.7 | |
| Cofactor biosynthesis | 9.0 | 18.6 | |
| Cytokinin 7-N-glucoside biosynthesis | 2.0 | 1.4 | |
| Cytokinin 9-N-glucoside biosynthesis | 2.0 | 1.4 | |
| Detoxification | 2.0 | - | |
| Fatty acid and lipid metabolism | 2.0 | - | |
| Generation of precursor metabolites and energy | 2.0 | - | |
| Hormone signaling, transport, and metabolism | 25.0 | 21.4 | |
| Inorganic nutrients metabolism | 7.0 | 7.1 | |
| Photorespiration | 2.0 | - | |
| Secondary metabolism | 9.0 | 7.1 | |
| Response to stimuli: abiotic and stimuli and stresses | Response to cold temperature | 2.0 | 2.9 |
| Response to heavy metals | 1.0 | 1.4 | |
| Response to phosphate deficiency | 1.0 | - | |
| Response to stimuli: biotic and stimuli and stresses | Recognition of fungal and bacterial pathogens and immunity response | 1.0 | - |
| Function | MSU ID | Description | Low N | Early N Recovery | Low N | Early N Recovery |
|---|---|---|---|---|---|---|
| Pokkali | Bengal | |||||
| Nitrate transporters | LOC_Os10g40600 | Nitrate transporter 1.1B | - | U | - | U |
| LOC_Os02g02170 | High-affinity nitrate transporter 2.1 | U | U | - | U | |
| LOC_Os02g02190 | High-affinity nitrate transporter 2.2 | U | U | U | U | |
| LOC_Os01g50820 | High-affinity nitrate transporter 2.3 | U | - | U | - | |
| Ammonium transporter | LOC_Os02g40730 | Ammonium transporter 1 member 2 | D | U | - | U |
| Nitrate reductase | LOC_Os08g36480 | Nitrate reductase 1 | - | - | - | U |
| LOC_Os08g36500 | Nitrate reductase 2 | - | - | - | U | |
| LOC_Os02g53130 | NADH/NADPH-dependent NO3-reductase 2 | D | U | D | U | |
| Glutamine synthetase | LOC_Os03g12290 | Glutamine synthetase 1;2 | D | U | D | U |
| GOGAT | LOC_Os01g48960 | NADH-GOGAT | - | U | - | U |
| TF Family | Description | Low N | Early N Recovery | Low N | Early N Recovery |
|---|---|---|---|---|---|
| Pokkali | Bengal | ||||
| AP2 | AP2 family protein | 1 | 0 | 0 | 0 |
| B3 | B3 family protein | 0 | 0 | 3 | 1 |
| bHLH | basic/helix-loop-helix family proteins | 10 | 5 | 6 | 7 |
| bZIP | bZIP family protein | 3 | 6 | 0 | 6 |
| C2H2 | C2H2 zinc finger domain | 9 | 6 | 5 | 3 |
| C3H | Cys3His -containing zinc finger domain | 0 | 1 | 0 | 0 |
| CO-like | CO (CONSTANS) family protein | 1 | 1 | 0 | 1 |
| Dof | Dof (DNA binding with one finger) family | 2 | 0 | 0 | 0 |
| EIL | Ethylene-insensitive3 (EIN3) and EIN3-like (EIL) family proteins | 0 | 2 | 0 | 1 |
| ERF | ERF family protein | 9 | 18 | 8 | 15 |
| FAR1 | FAR1 family protein | 1 | 0 | 1 | 1 |
| G2-like | G2-like family protein | 5 | 3 | 1 | 0 |
| GATA | GATA family protein | 2 | 0 | 2 | 1 |
| GRAS | GRAS family protein | 1 | 0 | 1 | 1 |
| GRF | GROWTH-REGULATING FACTOR family protein | 0 | 1 | 0 | 1 |
| HD-ZIP | HD-ZIP family protein | 0 | 3 | 3 | 1 |
| HSF | Heat stress transcription factors | 1 | 1 | 0 | 1 |
| LBD | LBD family protein | 2 | 5 | 3 | 4 |
| MIKC-MADS | MIKC-MADS family protein | 1 | 0 | 1 | 0 |
| M-type MADS | M-type MADS family protein | 0 | 0 | 0 | 1 |
| MYB | MYB family protein | 8 | 1 | 4 | 4 |
| MYB-related | MYB-related family protein | 3 | 2 | 2 | 2 |
| NAC | NAM, ATAF, and CUC (NAC) transcription factors | 8 | 3 | 7 | 3 |
| NF-YA | NF-YA family protein | 2 | 0 | 2 | 0 |
| NF-YB | NF-YB family protein | 0 | 1 | 0 | 1 |
| Nin-like | Nin (for nodule inception)-like family protein | 1 | 1 | 1 | 0 |
| RAV | RAV family protein | 0 | 1 | 0 | 1 |
| SBP | SQUAMOSA promoter binding proteins (SBPs) | 1 | 0 | 1 | 0 |
| WOX | WOX family protein | 0 | 1 | 0 | 0 |
| WRKY | WRKY family protein | 17 | 2 | 9 | 9 |
| Total | 88 | 64 | 60 | 65 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subudhi, P.K.; Garcia, R.S.; Coronejo, S.; Tapia, R. Comparative Transcriptomics of Rice Genotypes with Contrasting Responses to Nitrogen Stress Reveals Genes Influencing Nitrogen Uptake through the Regulation of Root Architecture. Int. J. Mol. Sci. 2020, 21, 5759. https://doi.org/10.3390/ijms21165759
Subudhi PK, Garcia RS, Coronejo S, Tapia R. Comparative Transcriptomics of Rice Genotypes with Contrasting Responses to Nitrogen Stress Reveals Genes Influencing Nitrogen Uptake through the Regulation of Root Architecture. International Journal of Molecular Sciences. 2020; 21(16):5759. https://doi.org/10.3390/ijms21165759
Chicago/Turabian StyleSubudhi, Prasanta K., Richard S. Garcia, Sapphire Coronejo, and Ronald Tapia. 2020. "Comparative Transcriptomics of Rice Genotypes with Contrasting Responses to Nitrogen Stress Reveals Genes Influencing Nitrogen Uptake through the Regulation of Root Architecture" International Journal of Molecular Sciences 21, no. 16: 5759. https://doi.org/10.3390/ijms21165759