Deimination, Intermediate Filaments and Associated Proteins


Abstract
:1. Introduction
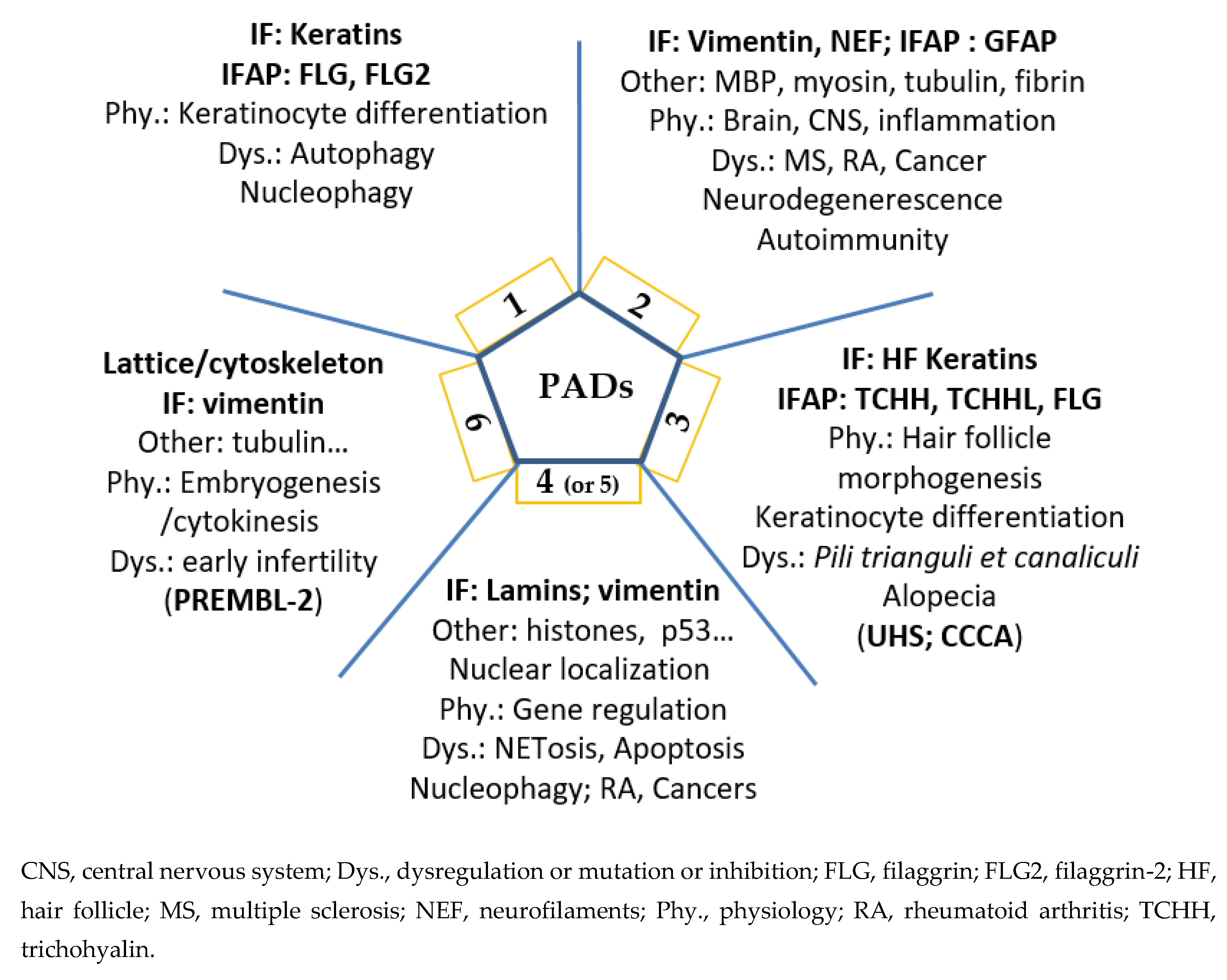
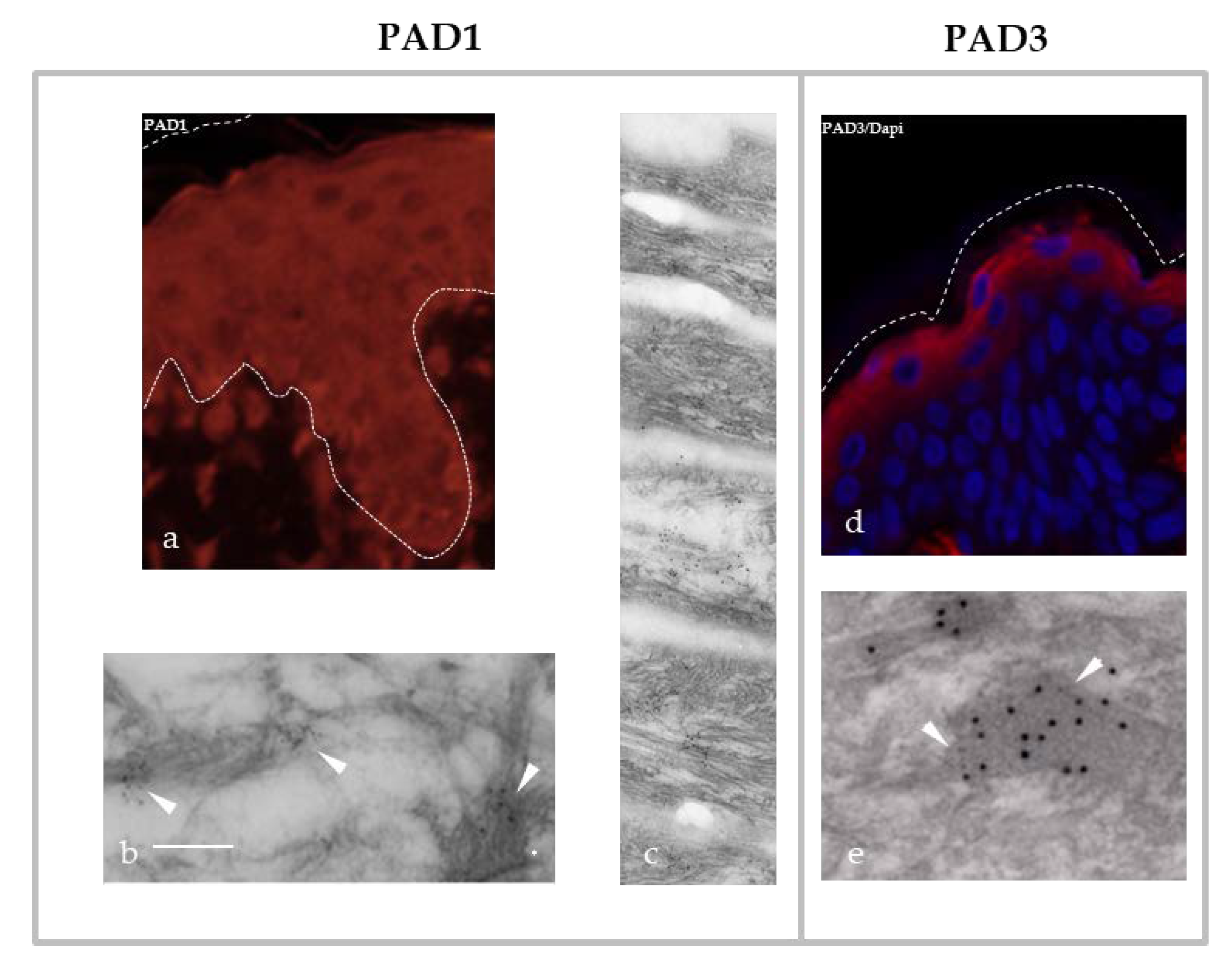
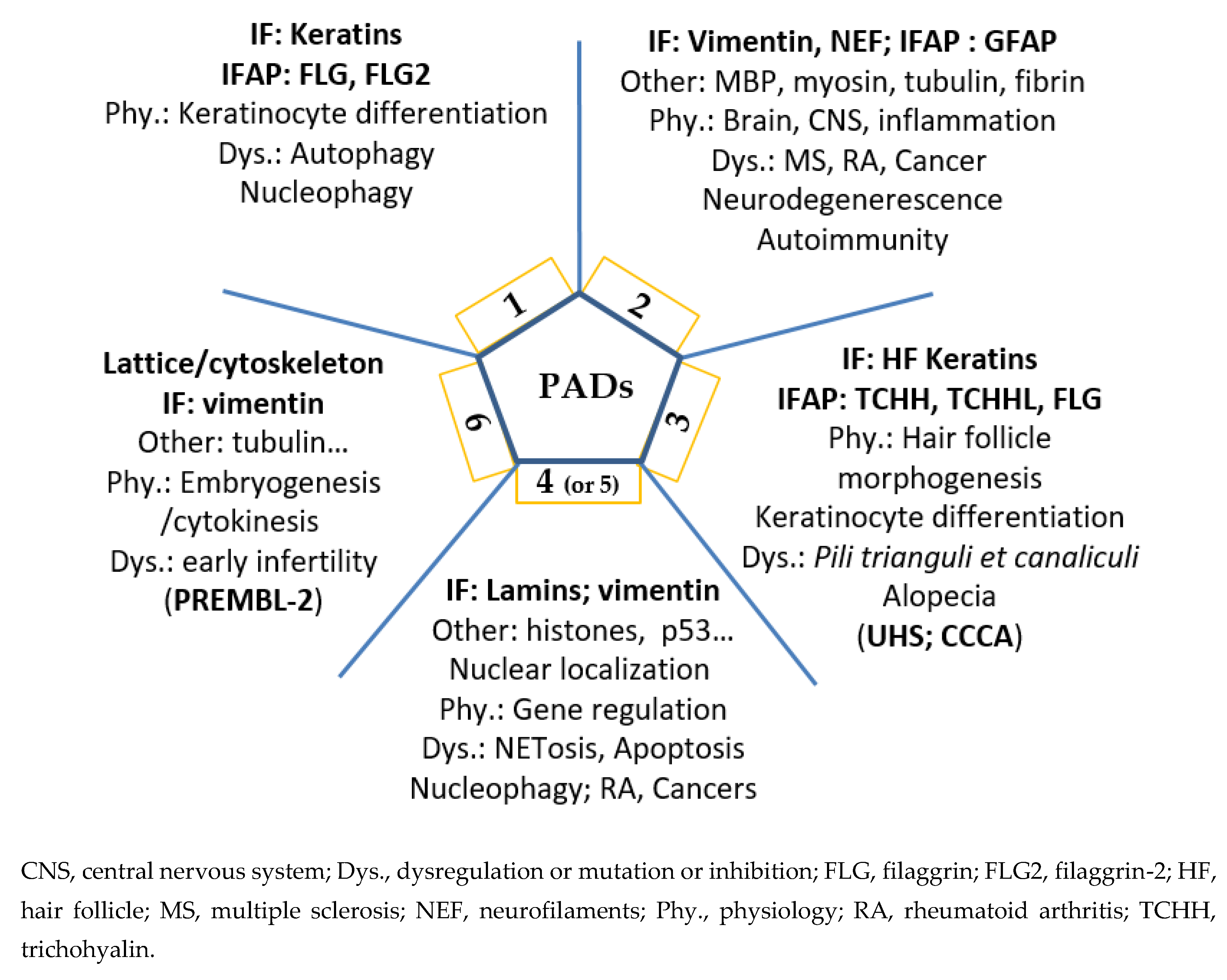
2. Five Human PADs, Their Differential Expression, Localization and Implication
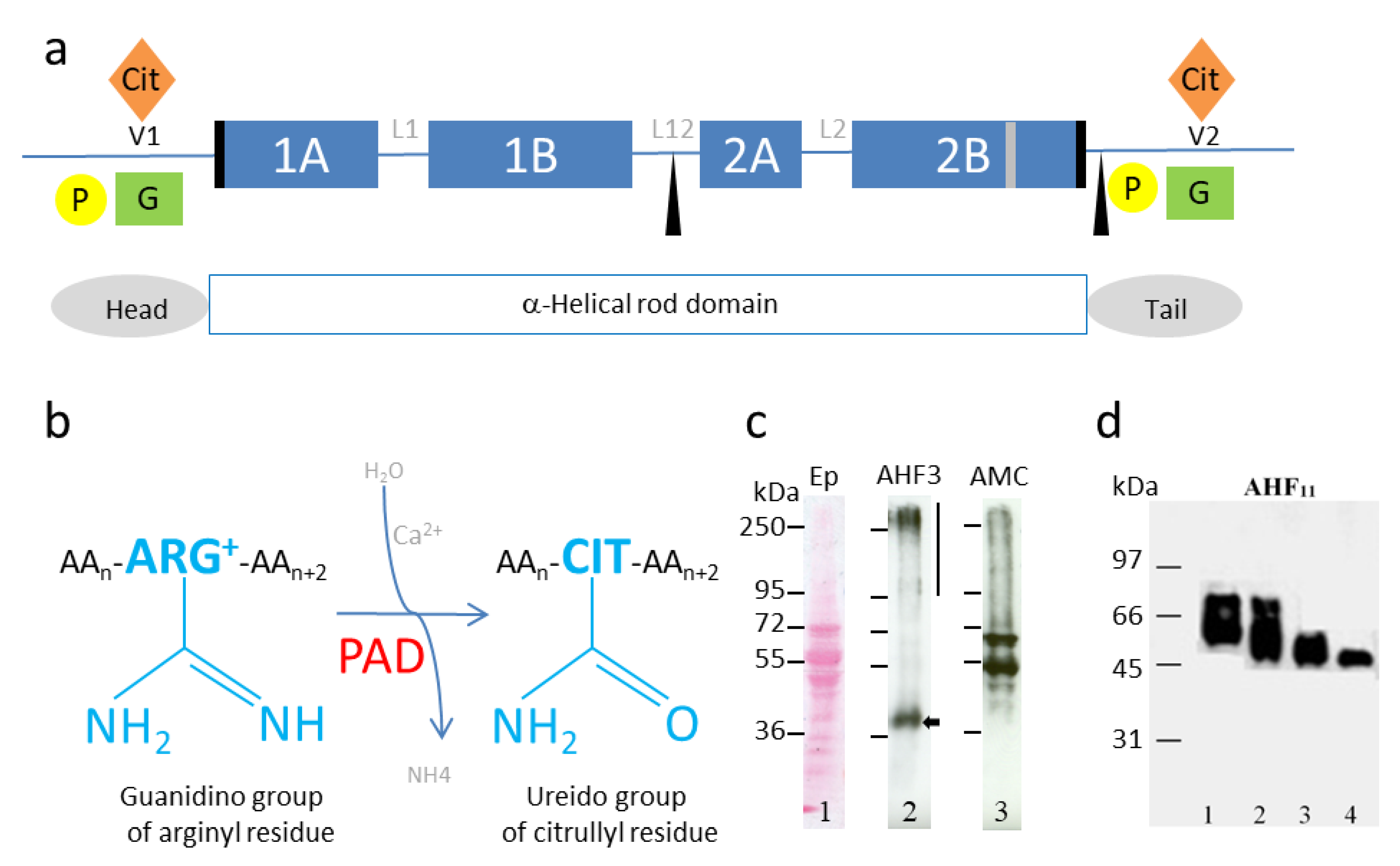
3. Deimination of IF Proteins, Impact on Their Polymerization and Solubility
4. Deimination of IFAP, Impact on Their Proteolysis
5. Hypercitrullination or Hypocitrullination
6. Proteomic Approaches of “Citrullinomes”
7. Major highlights and Summary
- Influences their polymerization and interaction
- Increases their solubility in low ionic strength buffer
- Promotes their proteolysis by several peptidases
- Promotes their cross-linking by transglutaminases
- By various exogenous environmental stimuli (chemical trauma, pressure and atmospheric dryness)
- By various chemical or biological agents (retinoic acid, calcium ionophores and inductors of apoptosis or NETosis)
- In many pathological contexts (rheumatoid arthritis, multiple sclerosis, Alzheimer’s disease, cancers, etc.)
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CCCA | Central Centrifugal Cicatricial Alopecia |
| GFAP | Glial fibrous acidic protein |
| IF | Intermediate filament |
| IFAP | Intermediate filament associated protein |
| MBP | Myelin basic protein |
| NETosis | Neutrophil extracellular trap activation and release |
| PAD | Peptidylarginine deiminase |
| UHS | Uncombable Hair Syndrome |
References
- Fisher, D.Z.; Chaudhary, N.; Blobel, G. cDNA sequencing of nuclear lamins A and C reveals primary and secondary structural homology to intermediate filament proteins. Proc. Natl. Acad. Sci. USA 1986, 83, 6450–6454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeon, F.D.; Kirschner, M.W.; Caput, D. Homologies in both primary and secondary structure between nuclear envelope and intermediate filament proteins. Nature 1987, 319, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Chernyatina, A.A.; Guzenko, D.; Strelkov, S.V. Intermediate filament structure: The bottom-up approach. Curr. Opin. Cell Biol. 2015, 32, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Windoffer, R.; Beil, M.; Magin, T.M.; Leube, R.E. Cytoskeleton in motion: The dynamics of keratin intermediate filaments in epithelia. J. Cell Biol. 2011, 194, 669–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mc Lean, W.H.I. and Moore, C.B.T. Keratin disorders: From gene to therapy. Hum. Mol. Genet. 2011, 20, R189–R197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, D.J.; Walsh, T.R.; Noro, M.G.; Notman, R. Complete structure of an epithelial keratin dimer: Implications for intermediate filament assembly. PLoS ONE 2015, 10, e0132706. [Google Scholar] [CrossRef] [Green Version]
- Strelkov, V.S.; Herrmann, H.; Aebi, U. Molecular architecture of intermediate filaments. BioEssays 2003, 25, 243–251. [Google Scholar] [CrossRef]
- Cau, L.; Méchin, M.-C.; Simon, M. Peptidylarginine deiminases and deiminated proteins at the epidermal barrier. Exp. Dermatol. 2018, 27, 852–858. [Google Scholar] [CrossRef] [Green Version]
- Henry, J.; Toulza, E.; Hsu, C.Y.; Pellerin, L.; Balica, S.; Mazereeuw-Hautier, J.; Paul, C.; Serre, G.; Jonca, N.; Simon, M. Update on the epidermal differentiation complex. Front. Biosci. (Landmark Ed) 2012, 17, 1517–1532. [Google Scholar] [CrossRef] [Green Version]
- Pellerin, L.; Henry, J.; Chiung-Yueh, H.; Balica, S.; Jean-Devoster, C.; Méchin, M.-C.; Hansmann, B.; Rodriguez, E.; Weindinger, S.; Schmitt, A.M.; et al. Defects of filaggrin-like proteins in both lesional and nonlesional atopic skin. J. Allergy Clin. Immunol. 2013, 131, 1094–1102. [Google Scholar] [CrossRef]
- Pendaries, V.; Malaisse, J.; Pellerin, L.; Le Lamer, M.; Nachat, R.; Kezic, S.; Schmitt, A.M.; Paul, C.; Poumay, Y.; Serre, G.; et al. Knockdown of filaggrin in a three-dimensional reconstructed human epidermis impairs keratinocyte differentiation. J. Investig. Dermatol. 2014, 134, 2938–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Lamer, M.; Pellerin, L.; Reynier, M.; Cau, L.; Pendaries, V.; Leprince, C.; Méchin, M.-C.; Serre, G.; Paul, C.; Simon, M. Defects of corneocyte structural proteins and epidermal barrier in atopic dermatitis. Biol. Chem. 2015, 396, 1163–1179. [Google Scholar] [CrossRef] [PubMed]
- Tarcsa, E.; Marekov, L.N.; Mei, G.; Melino, G.; Lee, S.-C.; Steinert, P.M. Protein unfolding by peptidylarginine deiminase substrate specificity and structural relationships of the natural substrates trichohyalin and filaggrin. J. Biol. Chem. 1996, 271, 30709–30716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manabe, M.; Sanchez, M.; Sun, T.T.; Dale, B.A. Interaction of filaggrin with keratin filaments during advanced stages of normal human epidermal differentiation and in ichthyosis vulgaris. Differentiation 1991, 48, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Dale, B.A.; Presland, R.B.; Lewis, S.P.; Underwood, R.A.; Fleckman, P. Transient expression of epidermal filaggrin in cultured cells causes collapse of intermediate filament networks with alteration of cell shape and nuclear integrity. J. Investig. Dermatol. 1997, 108, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Snider, N.; Omary, M.B. Assays for posttranslational modifications of intermediate filament proteins. Meth. Enz. 2016, 568, 113–138. [Google Scholar]
- Senshu, T.; Akiyama, K.; Kan, S.; Asaga, H.; Ishigami, A.; Manabe, M. Detection of deiminated proteins in rat skin: Probing with a monospecific antibody after modification of citrulline residues. J. Investig. Dermatol. 1995, 105, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Senshu, T.; Kan, S.; Ogawa, H.; Manabe, M.; Asaga, H. Preferential deimination of keratin K1 and filaggrin during the terminal differentiation of human epidermis. Biochem. Biophys. Res. Commun. 1996, 225, 712–719. [Google Scholar] [CrossRef]
- Senshu, T.; Akiyama, K.; Nomura, K. Identification of citrulline residues in the V subdomains of keratin K1 derived from the cornified layer of newborn mouse epidermis. Exp. Dermatol. 1999, 8, 392–401. [Google Scholar] [CrossRef]
- Inagaki, M.; Takahara, H.; Nishi, Y.; Sugawara, K.; Sato, C. Ca2+-dependent deimination-induced disassembly of intermediate filaments involves specific modification of the amino-terminal head domain. J. Biol. Chem. 1989, 264, 18119–18127. [Google Scholar]
- Hojo-Nakashima, I.; Sato, R.; Nakashima, K.; Hagiwara, T.; Yamada, M. Dynamic expression of peptidylarginine deiminase 2 in human monocytic leukaemia THP-1 cells during macrophage differentiation. J. Biochem. 2009, 146, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Jang, B.; Jin, J.-K.; Jeon, Y.-C.; Cho, H.J.; Ishigami, A.; Choi, K.-C.; Carp, R.I.; Maruyama, N.; Kim, Y.-S.; Choi, E.-K. Involvement of peptidylarginine deiminase-mediated post-translational citrullination in pathogenesis of sporadic Creutzfeldt-Jakob disease. Acta Neuropathol. 2009, 119, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, C.; Espinosa, M.; Suzuki, A.; Masuda, K.; Yamamoto, K.; Tsuchiya, E.; Ueda, K.; Daigo, Y.; Nakamura, Y.; Matsuda, K. Regulation of histone modification and chromatin structure by the p53-PADI4 pathway. Nat. Comm. 2012, 3, 676. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.E.; Harding, H.W.; Llewelyn-Smith, I.J. The origin of citrulline-containing proteins in the hair follicle and the chemical nature of trichohyalin, an intracellular precursor. Biochim. Biophys. Acta. 1977, 495, 159–175. [Google Scholar] [CrossRef]
- Tarcsa, E.; Marekov, L.N.; Andreoli, J.; Idler, W.W.; Candi, E.; Chung, S.; Steinert, P.M. The fate of trichohyalin. Sequential post-translational modifications by peptidylarginine deiminase and transglutaminases. Biol. Chem. 1997, 272, 27893–27901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, P.M.; Parry, D.A.D.; Marekov, L.N. Trichohyalin mechanically strengthens the hair follicle. Multiple cross-bridging roles in the inner root sheath. J. Biol. Chem. 2003, 278, 41409–41419. [Google Scholar] [CrossRef]
- Moscarello, M.A.; Mastronardi, F.G.; Wood, D.D. The role of citrullinated proteins suggests a novel mechanism in the pathogenesis of multiple sclerosis. Neurochem. Res. 2007, 32, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Mastronardi, F.G.; Moscarello, M. Molecules affecting myelin stability: A novel hypothesis regarding the pathogenesis of multiple sclerosis. J. Neurosci. Res. 2005, 80, 301. [Google Scholar] [CrossRef]
- Chen, L.Y.; Lin, K.R.; Chen, Y.J.; Chiang, Y.J.; Ho, K.C.; Shen, L.F.; Song, I.W.; Liu, K.M.; Yang-Yen, H.F.; Chen, Y.J.; et al. Palmitoyl acyltransferase activity of ZDHHC13 regulates skin barrier development partly by controlling PADI3 and TGM1 protein stability. J. Investig. Dermatol. 2020, 140, 959–970.e3. [Google Scholar] [CrossRef]
- Méchin, M.-C.; Takahara, H.; Simon, M. Deimination and peptidylarginine deiminases in skin physiology and diseases. Int. J. Mol. Sci. 2020, 21, 566. [Google Scholar] [CrossRef] [Green Version]
- Méchin, M.-C.; Sebbag, M.; Arnaud, J.; Nachat, R.; Foulquier, C.; Adoue, V.; Coudane, F.; Duplan, H.; Schmitt, A.-M.; Chavanas, S.; et al. Update on peptidylarginine deiminases and deimination in skin physiology and severe human diseases. Int. J. Cosmet. Sci. 2007, 29, 147–168. [Google Scholar] [CrossRef] [PubMed]
- Baka, Z.; György, B.; Géher, P.; Buzás, E.I.; Falus, A.; György, N.B. Citrullination under physiological and pathological conditions. Jt. Bone Spine. 2012, 79, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Cau, L.; Pendaries, V.; Lhuillier, E.; Thompson, P.R.; Serre, G.; Takahara, H.; Méchin, M.-C.; Simon, M. Lowering relative humidity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 2017, 86, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.-Y.; Henry, J.; Raymond, A.-A.; Méchin, M.-C.; Pendaries, V.; Nassar, D.; Hansmann, B.; Balica, S.; Burlet-Schiltz, O.; Schmitt, A.M.; et al. Deimination of human filaggrin-2 promotes its proteolysis by calpain 1. J. Biol. Chem. 2011, 286, 23222–23233. [Google Scholar] [CrossRef] [Green Version]
- Kizawa, K.; Unno, M.; Heizmann, C.W.; Takahara, H. Importance of Citrullination on Hair Protein Molecular Assembly during Trichocytic Differentiation; Springer: New York, NY, USA, 2002; Volume 8, pp. 129–148. [Google Scholar]
- Chermnykh, E.S.; Alpeeva, E.V.; Vorotelyak, E.A. Transglutaminase 3: The involvement in epithelial differentiation and cancer. Cells 2020, 9, 1996. [Google Scholar] [CrossRef]
- Arslan, M.; Qin, Z.; Buehler, M.J. Coiled-coil intermediate filament stutter instability and molecular unfolding. Compt. Methods Biomech. Biomed. Engin. 2011, 14, 483–489. [Google Scholar] [CrossRef]
- Simon, M.; Sebbag, M.; Haftek, M.; Vincent, C.; Girbal-Neuhauser, E.; Rakotoarivony, J.; Schmitt, D.; Serre, G.; Somme, G. Monoclonal antibodies to human epidermal filaggrin, some not recognizing profilaggrin. J. Investig. Dermatol. 1995, 105, 132–437. [Google Scholar] [CrossRef] [Green Version]
- Méchin, M.-C.; Enji, M.; Nachat, R.; Chavanas, S.; Charveron, M.; Ishida-Yamamoto, A.; Serre, G.; Takahara, H.; Simon, M. The peptidylarginine deiminases expressed in human epidermis differ in their substrate specificities and subcellular locations. Cell. Mol. Life Sci. 2005, 62, 1984–1995. [Google Scholar] [CrossRef]
- Chavanas, S.; Méchin, M.-C.; Takahara, H.; Kawada, A.; Nachat, R.; Serre, G.; Simon, M. Comparative analysis of the mouse and human peptidylarginine deiminase gene clusters reveals highly conserved non-coding segments and a new human gene. PADI Gene 2004, 14, 19–27. [Google Scholar] [CrossRef]
- Guerrin, M.; Ishigami, A.; Méchin, M.-C.; Nachat, R.; Valmary, S.; Sebbag, M.; Simon, M.; Senshu, T.; Serre, G. cDNA cloning, gene organization and expression analysis of human peptidylarginine deiminase type I. Biochem. J. 2003, 370, 167–174. [Google Scholar] [CrossRef]
- Nachat, R.; Méchin, M.-C.; Takahara, H.; Chavanas, S.; Charveron, M.; Serre, G.; Simon, M. Peptidylarginine deiminase isoforms 1–3 are expressed in the epidermis and involved in the deimination of K1 and filaggrin. J. Investig. Dermatol. 2005, 124, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuckley, B.; Causey, C.P.; Jones, J.E.; Bhatia, M.; Dreyton, C.J.; Osborme, T.C.; Takahara, H.; Thompson, P.R. Substrate specificity and kinetic studies of PADs 1, 3, and 4 identify potent and selective inhibitors of protein arginine deiminase 3. Biochemistry 2010, 49, 4852–4863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama-Hamada, M.; Suzuki, A.; Kubota, K.; Takazawa, T.; Ohsaka, M.; Kawaida, R.; Ono, M.; Kasuya, A.; Furukawa, H.; Yamada, R.; et al. Comparison of enzymatic properties between hPADI2 and hPADI4. Biochem. Biophys. Res. Commun. 2005, 327, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Dong, S.; Kawada, A.; Kojima, T.; Chavanas, S.; Méchin, M.-C.; Adoue, V.; Serre, G.; Simon, M.; Takahara, H. Transcriptional regulation of peptidylarginine deiminase expression in human keratinocytes. J. Dermatol. Sci. 2009, 53, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Adoue, V.; Chavanas, S.; Coudane, F.; Méchin, M.C.; Caubet, C.; Ying, S.; Dong, S.; Duplan, H.; Charveron, M.; Takahara, H.; et al. Long-range enhancer differentially regulated by c-Jun and JunD controls peptidylarginine deiminase-3 gene in keratinocytes. J. Mol. Biol. 2008, 384, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Méchin, M.-C.; Coudane, F.; Adoue, V.; Arnaud, J.; Duplan, H.; Charveron, M.; Schmitt, A.M.; Takahara, H.; Serre, G.; Simon, M. Deimination is regulated at multiple levels including auto-deimination of peptidylarginine deiminases. Cell. Mol. Life Sci. 2010, 67, 1491–1503. [Google Scholar] [CrossRef]
- Nakashima, K.; Hagiwara, T.; Ishigami, A.; Nagata, S.; Asaga, H.; Kuramoto, M.; Senshu, T.; Yamada, M. Molecular characterization of peptidylarginine deiminase in HL-60 cells induced by retinoic acid and 1alpha,25-dihydroxyvitamin D(3). J. Biol. Chem. 1999, 274, 27786–27792. [Google Scholar] [CrossRef] [Green Version]
- Andrade, F.; Darrah, E.; Gucek, M.; Cole, R.N.; Rosen, A.; Zhu, X. Autocitrullination of human peptidyl arginine deiminase type 4 regulates protein citrullination during cell activation. Arthritis Rheum. 2010, 62, 1630–1640. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.L.; Jones, L.E., Jr.; Bhatia, M.M.; Thompson, P.R. Autodeimination of protein arginine deiminase 4 alters protein-protein interactions but not activity. Biochemistry 2011, 50, 3997–4010. [Google Scholar] [CrossRef] [Green Version]
- Méchin, M.-C.; Nachat, R.; Coudane, F.; Adoue, V.; Arnaud, J.; Serre, G.; Simon, M. Deimination or citrullination: A post-translational modification with many facettes. Med. Sci. 2011, 27, 49–54. [Google Scholar]
- Rud’s, A.A.; Ikejiri, Y.; Ono, H.; Yonekawa, T.; Shiraiwa, M.; Kawada, A.; Takahara, H. Molecular cloning of cDNA of mouse peptidylarginine deiminase type I, type III and type IV, and the expression pattern of type I in mouse. J. Biochem. 1999, 259, 660–669. [Google Scholar]
- Garcia Quiroz, F.; Fiore, V.F.; Levorse, J.; Polak, L.; Wong, E.; Pasolli, H.A.; Fuchs, E. Liquid-liquid phase separation drives skin barrier formation. Science 2020, 367, eaax9554. [Google Scholar]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.-M.; Hsiao, Y.-C.; Chien, K.-Y.; Chen, S.-F.; Chuang, Y.-N.; Lin, S.-Y.; Wang, W.S.; Chang, I.Y.-F.; Yang, C.; Chu, L.J.; et al. Assessment of candidate biomarkers in paired saliva and plasma samples from oral cancer patients by targeted mass spectrometry. J. Proteom. 2020, 211, 103571. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Liu, X.; Li, F.; Miao, L.; Li, T.; Xu, B.; An, X.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; et al. PAD1 promotes epithelial-mesenchymal transition and metastasis in triple-negative breast cancer cells by regulating MEK1-ERK1/2-MMP2 signaling. Cancer Lett. 2017, 409, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahara, H.; Okamoto, H.; Sugawara, K. Specific modification of the functional arginine residue in soybean trypsin inhibitor (Kunitz) by peptidylarginine deiminase. J. Biol. Chem. 1985, 260, 8378–8383. [Google Scholar] [PubMed]
- Takahara, H.; Okamoto, H.; Sugawara, K. Affinity chromatography of peptidylarginine deiminase from rabbit skeletal muscle on a column of soybean trypsin inhibitor (Kunitz)-sepharose. J. Biochem. 1986, 99, 1417–1424. [Google Scholar] [CrossRef]
- Ishiyama, N.; Bates, I.R.; Hill, C.M.; Wood, D.D.; Matharu, P.; Viner, N.J.; Moscarello, M.A.; Harauz, G. The effects of deimination of myelin basic protein on structures formed by its interaction with phosphoinositide-containing lipid monolayers. J. Struct. Biol. 2001, 136, 30–45. [Google Scholar] [CrossRef]
- Ishigami, A.; Ohsawa, T.; Hiratsuka, M.; Taguchi, H.; Kobayashi, S.K.; Saito, Y.; Murayama, S.; Asaga, H.; Toda, T.; Kimura, N.; et al. Abnormal accumulation of citrullinated proteins catalyzed by peptidylarginine deiminase in hippocampal extracts from patients with Alzheimer’s disease. J. Neurosci. Res. 2005, 80, 120–128. [Google Scholar] [CrossRef]
- Bhattacharya, S.H.; Crabb, J.S.; Bonilha, V.L.; Gu, X.; Takahara, H.; Crabb, J.W. Proteomics implicates peptidylarginine deiminase 2 and optic nerve citrullination in glaucome pathogenesis. Investig. Ophtalmol. Vis. Sci. 2006, 47, 2508–2514. [Google Scholar] [CrossRef]
- Raijmakers, R.; Vogelzangs, J.; Raats, J.; Panzenbeck, M.; Corby, M.; Jiang, H.; Thibodeau, M.; Haynes, N.; van Venrooij, W.J.; Pruijn, G.J.M.; et al. Experimental autoimmune encephalomyelitis induction in peptidylarginine deiminase 2 knockout mice. J. Comp. Neurol. 2006, 498, 217–226. [Google Scholar] [CrossRef]
- Coudane, F.; Méchin, M.-C.; Huchenq, A.; Henry, J.; Nachat, R.; Ishigami, A.; Adoue, V.; Sebbag, M.; Serre, G.; Simon, M. Deimination and expression of peptidylarginine deiminases during cutaneous wound healing in mice. Eur. J. Dermatol. 2011, 21, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Mendanha Falcão, A.; Meijer, M.; Scaglione, A.; Nielsen, M.L.; Casaccia, P.; Costelo-Branco, G. PAD2-mediated citrullination contributes to efficient oligodendrocyte differentiation and myelination. Cell Rep. 2019, 27, 1090–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachat, R.; Méchin, M.-C.; Charveron, M.; Serre, G.; Constans, J.; Simon, M. Peptidylarginine deiminase isoforms are differentially expressed in the anagen hair follicles and other human skin appendages. J. Investig. Dermatol. 2005, 125, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basmanav, Ü.F.B.; Cau, L.; Tafazzoli, A.; Méchin, M.-C.; Wolf, S.; Romano, M.T.; Valentin, F.; Wiegmann, H.; Huchenq, A.; Kandil, R.; et al. Mutations in Three Genes Encoding Proteins Involved in Hair Shaft Formation Cause Uncombable Hair Syndrome. Am. J. Hum. Genet. 2016, 99, 1292–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malki, L.; Sarig, O.; Romano, M.T.; Méchin, M.-C.; Peled, A.; Pavlovsky, M.; Warshauer, E.; Samuelov, L.; Uwakwe, L.; Briskin, V.; et al. Variant PADI3 in Central Centrifugal Cicatricial Alopecia. N. Engl. J. Med. 2019, 380, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Pong, K.; Subramanian, V.; Nicholas, A.P.; Thompson, P.R.; Ferretti, P. Modulation of calcium-induced cell death in human neural stem cells by the novel peptidylarginine deiminase-AIF pathway. Biochim. Biophys. Acta. 2014, 1843, 1162–1171. [Google Scholar]
- Ishigami, A.; Ohsawa, T.; Asaga, H.; Akiyama, K.; Kuramoto, M.; Maruyama, N. Human peptidylarginine deiminase type II: Molecular cloning, gene organization, and expression in human skin. Arch. Biochem. Biophys. Eng. 2011, 14, 483–489. [Google Scholar] [CrossRef]
- Kanno, T.; Kawada, A.; Yamanouchi, J.; Yosida-Noro, C.; Yoshiki, A.; Shiraiwa, M.; Kusakabe, M.; Manabe, M.; Tezuka, T.; Takahara, H. Human peptidylarginine deiminase type III: Molecular cloning and nucleotide sequence of the cDNA, properties of the recombinant enzyme, and immunohistochemical localization in human skin. J. Investig. Dermatol. 2000, 115, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wysocka, J.; Sayegh, J.; Lee, Y.H.; Perlin, J.R.; Leonelli, L.; Sonbuchner, L.S.; Mc Donald, C.H.; Cook, R.G.; Dou, Y.; et al. Human PAD4 regulates histone arginine methylation levels via demethylimination. Science 2004, 306, 279–283. [Google Scholar] [CrossRef]
- Cuthbert, G.L.; Daujat, S.; Snowden, A.W.; Erdjument-Bromage, H.; Hagiwara, T.; Yamada, M.; Schneider, R.; Gregory, P.D.; Tempst, P.; Bannister, A.J.; et al. Histone deimination antagonizes arginine methylation. Cell 2004, 118, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, A.; Yamada, R.; Chang, X.; Tokuhiro, S.; Sawada, T.; Suzuki, M.; Nagasaki, M.; Nakayama-Hamada, M.; Kawaida, R.; Ono, M.; et al. Functional haplotypes of PADI4, encoding citrullinating enzyme peptidylarginine deiminase 4, are associated with rheumatoid arthritis. Nat. Genet. 2003, 34, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.W.; Bolling, L.C.; Calvert, M.E.; Sarmento, O.F.; Berkeley, E.V.; Shea, M.C.; Hao, Z.; Jayes, F.C.; Bush, L.A.; Shetty, J.; et al. ePAD, an oocyte and early embryo-abundant peptidylarginine deiminase-like protein that localizes to egg cytoplasmic sheets. Dev. Biol. 2003, 256, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, Y.; Fu, J.; Yu, M.; Feng, R.; Sang, Q.; Liang, B.; Chen, B.; Qu, R.; Li, B.; et al. Mutations in PADI6 cause female infertility characterized by early embryonic arrest. Am. J. Hum. Genet. 2016, 99, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Maddirevula, S.; Coskun, S.; Awartani, K.; Alsaif, H.; Abdulwahab, F.M.; Alkuraya, F.S. The human knockout phenotype of PADI6 is female sterility caused by cleavage failure of their fertilized eggs. Clin. Genet. 2017, 91, 344–345. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, A.; Kuramoto, M.; Yamada, M.; Watanabe, K.; Senshu, T. Molecular cloning of two novel types of peptidylarginine deiminase cDNAs from retinoic acid-treated culture of a newborn rat keratinocyte cell line. FEBS Lett. 1998, 433, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Hagiwara, T.; Yamada, M. Nuclear localization of peptidylarginine deiminase V and histone deimination in granulocytes. J. Biol. Chem. 2002, 277, 49562–49568. [Google Scholar] [CrossRef] [Green Version]
- Foulquier, C.; Sebbag, M.; Clavel, C.; Chapuy-Regaud, S.; Al Badine, R.; Méchin, M.-C.; Vincent, C.; Nachat, R.; Yamada, M.; Takahara, H.; et al. Peptidylarginine deiminase type 2 (PAD-2) and PAD-4 but not PAD-1, PAD-3, and PAD-6 are expressed in rheumatoid arthritis synovium in close association with tissue inflammation. Arthritis. Rheum. 2007, 11, 3541–3553. [Google Scholar] [CrossRef]
- Yang, X.-K.; Liu, J.; Liu, J.; Liang, Y.; Xu, W.-D.; Leng, R.-X.; Pan, H.-F.; Ye, D.-Q. Associations between PADI4 gene polymorphisms and rheumatoid arthritis: An update meta-analysis. Arch. Med. Res. 2015, 12, 33–51. [Google Scholar] [CrossRef]
- Barton, A.; Bowes, J.; Eyre, S.; Symmons, D.; Worthington, J.; Silman, A. Investigation of polymorphisms in the PADI4 gene in determining severity of inflammatory polyarthritis. Ann. Rheum. Dis. 2005, 64, 1311–1315. [Google Scholar] [CrossRef] [Green Version]
- Massarenti, L.; Enevold, C.; Damgaard, D.; Ødum, N.; Nielsen, C.H.; Jacobsen, S. Peptidylarginine deiminase-4 gene polymorphisms are associated with systemic lupus erythematosus and lupus nephritis. Scand. J. Rheumatol. 2019, 48, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An overview of the intrinsic role of citrullination in autoimmune disorders. J. Immunol. Res. 2019, 25, 7592851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell. Biol. 2009, 184, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konig, M.F.; Andrade, F. A critical reappraisal of neutrophil extracellular traps and NETosis mimics based on differential requirements for protein citrullination. Front. Immunol. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Mol. 2010, 207, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Guiducci, E.; Lenberg, C.; Küng, N.; Schraner, E.; Theocharides, A.P.A.; LeibundGut-Landmann, S. Candida albicans-Induced NETosis is independent of peptidylarginine deiminase 4. Front. Immunol. 2018, 9, 1573. [Google Scholar] [CrossRef]
- Gößwein, S.; Lindemann, A.; Mahajan, A.; Maueröder, C.; Martini, E.; Patankar, J.; Schett, G.; Becker, C.; Wirtz, S.; Naumann-Bartsch, N.; et al. Citrullination licenses calpain to decondense nuclei in neutrophil extracellular trap formation. Front. Immunol. 2019, 22, 2481. [Google Scholar] [CrossRef]
- Masson-Bessière, C.; Sebbag, M.; Girbal-Neuhauser, E.; Nogueira, L.; Vincent, C.; Senshu, T.; Serre, G. The major synovial targets of the rheumatoid arthritis-specific antifilaggrin autoantibodies are deiminated forms of the alpha- and beta-chains of fibrin. J. Immunol. 2001, 166, 4177–4184. [Google Scholar] [CrossRef] [Green Version]
- Cornillet, M.; Sebbag, M.; Verrouil, E.; Magyar, A.; Babos, F.; Ruyssen-Witrand, A.; Hudecz, F.; Cantagrel, A.; Serre, G.; Nogueira, L. The fibrin-derived citrullinated peptide β60–74Cit60,72,74 bears the major ACPA epitope recognised by the rheumatoid arthritis-specific anticitrullinated fibrinogen autoantibodies and anti-CCP2 antibodies. Ann. Rheum. Dis. 2014, 73, 1246–1252. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, B.; Mittereder, N.; Chaerkady, R.; Strain, M.; An, L.-L.; Rahman, S.; Ma, W.; Low, C.P.; Chan, D.; et al. Spontaneous secretion of the citrullination enzyme PAD2 and cell surface exposure of PAD4 by neutrophils. Front. Immunol. 2017, 8, 1200. [Google Scholar] [CrossRef] [Green Version]
- Criscitiello, M.F.; Kraev, I.; Petersen, L.H.; Lange, S. Deimination protein profiles in Alligator mississippiensis reveal plasma and extracellular vesicle-specific signatures relating to immunity, metabolic function, and gene regulation. Front. Immunol. 2020, 11, 651. [Google Scholar] [CrossRef] [PubMed]
- Horibata, S.; Coonrod, S.A.; Cherrington, B.D. Role for peptidylarginine deiminase enzymes in disease and female reproduction. J. Reprod. Dev. 2012, 58, 274–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vossenaar, E.R.; Després, N.; Lapointe, E.; van der Heijden, A.; Lora, M.; Senshu, T.; van Venrooij, W.J.; Ménard, H.A. Rheumatoid arthritis specific anti-Sa antibodies target citrullinated vimentin. Arthitis Res. Ther. 2004, 6, R142–R150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, A.P.; Sambandam, T.; Echols, J.D.; Tourtelotte, W.W. Increased citrullinated glial fibrillary acid protein in secondary progressive multiple sclerosis. J. Comp. Neurol. 2004, 473, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Union, A.; Meheus, L.; Humbel, R.L.; Conrad, K.; Steiner, G.; Moereels, H.; Pottel, H.; Serre, G.; De Keyser, F. Identification of citrullinated rheumatoid arthritis-specific epitopes in natural filaggrin relevant for antifilaggrin autoantibody detection by line immunoassay. Arthitis. Rheum. 2002, 46, 1185–1195. [Google Scholar] [CrossRef]
- Wizeman, J.W.; Nicholas, A.P.; Ishigami, A.; Mohan, R. Citrullination of glial intermediate filaments is early response in retinal injury. Mol. Vis. 2016, 22, 1137–1155. [Google Scholar]
- Ishida-Yamamoto, A.; Senshu, T.; Takahashi, H.; Akiyama, K.; Nomura, K.; Iizuka, H. Decreased deiminated keratin K1 in psoriatic hyperproliferative epidermis. J. Investig. Dermatol. 2000, 114, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Yamamoto, A.; Senshu, T.; Eady, R.A.; Takahashi, H.; Shimizu, H.; Akiyama, M.; Lizuka, H. Sequential reorganization of cornified cell keratin filaments involving filaggrin-mediated compaction and keratin 1 deimination. J. Investig. Dermatol. 2002, 118, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Méchin, M.-C.; Cau, L.; Galliano, M.F.; Daunes-Marion, S.; Poigny, S.; Vidaluc, J.L.; Bessou-Touya, S.; Takahara, H.; Serre, G.; Duplan, H.; et al. Acefylline activates filaggrin deimination by peptidylarginine deiminases in the upper epidermis. J. Dermatol. Sci. 2016, 81, 101–106. [Google Scholar] [CrossRef]
- Daunes-Marion, S.; Duplan, H.; Méchin, M.-C.; Poigny, S.; Serre, G.; Simon, M. Peptidylarginine Deiminase 1 and/or 3 Activator Compounds in the Epidermis and Uses. Thereof. Patent n°WO2012140095A1, 18 October 2012. [Google Scholar]
- Arita, S.; Hatta, M.; Uchida, K.; Kita, T.; Okamura, K.; Ryu, T.; Murakami, H.; Sakagami, R.; Yamazaki, J. Peptidylarginine deiminase is involved in maintaining the cornified oral mucosa of rats. J. Periodontal Res. 2018, 53, 750–761. [Google Scholar] [CrossRef]
- Steinert, P.M.; Cantieri, J.S.; Teller, D.C.; Lonsdale-Eccles, J.D.; Dale, B.A. Characterization of a class of cationic proteins that specifically interact with intermediate filaments. Proc. Natl. Acad. Sci. USA 1981, 78, 4097–4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamata, Y.; Taniguchi, A.; Yamamoto, M.; Nomura, J.; Ishiara, K.; Takahara, H.; Hibino, T.; Takeda, A. Neutral cysteine protease bleomycin hydrolase is essential for the breakdown of deiminated filaggrin into amino acids. J. Biol. Chem. 2009, 284, 12829–12836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, W.M.; Steven, A.C.; Steinert, P.M. The mechanism of interaction of filaggrin with intermediate filaments. The ionic zipper hypothesis. J. Mol. Biol. 1993, 232, 50–66. [Google Scholar]
- Pritzker, L.B.; Joshi, S.; Gowan, J.J.; Harauz, G.; Moscarello, M.A. Deimination of myelin basic protein. 1. Effect of deimination of arginyl residues of myelin basic protein on its structure and susceptibility to digestion by cathepsin D. Biochemistry 2000, 39, 5374–5381. [Google Scholar] [CrossRef] [PubMed]
- Nakayama-Hamada, M.; Suzuki, A.; Furukawa, H.; Yamada, R.; Yamamoto, K. Citrullinated fibrinogen inhibits thrombin-catalysed fibrin polymerization. J. Biochem. 2008, 144, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Okumura, N.; Haneishi, A.; Terasawa, F. Citrullinated fibrinogen shows defects in FPA and FPB release and fibrin polymerization catalyzed by thrombin. Clin. Chim. Acta. 2009, 401, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Bezuidenhout, J.A.; Venter, C.; Roberts, T.; Tarr, G.; Kell, D.B.; Pretorius, E. The atypical fibrin fibre network in rheumatoid arthritis and its relation to autoimmunity, inflammation and thrombosis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Proost, P.; Loos, T.; Mortier, A.; Schutyser, E.; Gouwy, M.; Noppen, S.; Dillen, C.; Ronsse, I.; Conings, R.; Struyf, S.; et al. Citrullination of CXCL8 by peptidylarginine deiminase alters receptor usage, prevents proteolysis, and dampens tissue inflammation. J. Exp. Mol. 2008, 205, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Singh, P.; Azhar, A.; Naseem, A.; Rashid, Q.; Kabir, M.A.; Jairajpuri, M.A. Serpin inhibition mechanism: A delicate balance between native metastable state and polymerization. J. Amino Acids 2011, 2001, 606797. [Google Scholar] [CrossRef] [Green Version]
- Tilvawala, R.; Nguyen, S.H.; Maurais, A.J.; Nemmara, V.V.; Nagar, M.; Salinger, A.J.; Nagpal, S.; Weerapana, E.; Thompson, P.R. The rheumatoid arthritis-associated citrullinome. Cell. Chem. Biol. 2018, 25, 691–704.e6. [Google Scholar] [CrossRef] [Green Version]
- Teshima, H.; Kato, M.; Tatsukawa, H.; Hitomi, K. Analysis of the expression of transglutaminases in the reconstructed human epidermis using a three-dimensional cell culture. Anal. Biochem. 2020, 603, 113606. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Gasc, G.; Raymond, A.A.; Burlet-Schiltz, O.; Takahara, H.; Serre, G.; Mechin, M.C.; Simon, M. Deimination of human hornerin enhances its processing by calpain-1 and its cross-linking by transglutaminases. J. Invest. Dermatol. 2017, 137, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albérola, G.; Schröder, J.M.; Froment, C.; Simon, M. The amino-terminal part of human FLG2 is a component of cornified envelopes. J. Investig. Dermatol. 2019, 139, 1395–1397. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Di Pucchio, T.; Sims, G.P.; Mittereder, N.; Mustelin, T. Characterization of the hypercitrullination reaction in human neutrophils and other leukocytes. Mediators Inflamm. 2015, 2015, 236451. [Google Scholar] [CrossRef] [Green Version]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter actinomycetemcomitans-induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Pingxin, L.; Wang, S.; Hu, J.; Chen, X.A.; Wu, J.; Fisher, M.; Oshaben, K.; Zhao, N.; Gu, Y.; et al. Anticancer peptidylarginine deiminase (PAD) inhibitors regulate the autophagy flux and the mammalian target of rapamycin complex 1 activity. J. Biol. Chem. 2012, 287, 25941–25953. [Google Scholar] [CrossRef] [Green Version]
- Cau, L.; Takahara, H.; Thompson, P.R.; Serre, G.; Méchin, M.-C.; Simon, M. Peptidylarginine deiminase inhibitor cl-amidine attenuates cornification and interferes with the regulation of autophagy in reconstructed human epidermis. J. Investig. Dermatol. 2019, 139, 1889–1897.e4. [Google Scholar] [CrossRef]
- Luo, Y.; Arita, K.; Bhatia, M.; Knuckley, B.K.; Lee, Y.-H.; Stallcup, M.R.; Sato, M.; Thompson, P.R. Inhibitors and inactivators of protein arginine deiminase 4: Functional and structural characterization. Biochemistry 2006, 45, 11727–11736. [Google Scholar] [CrossRef] [Green Version]
- Akinduro, O.; Sullu, K.; Patel, A.; Robinson, D.; Chikh, A.; McPhail, G.; Braun, K.M.; Philpott, M.P.; Harwood, C.A.; Byrne, C.; et al. Constitutive autophagy and nucleophagy during epidermal differentiation. J. Investig. Dermatol. 2016, 136, 1460–1470. [Google Scholar] [CrossRef] [Green Version]
- Bai, W.S.; Herrera-Abreu, M.T.; Rohn, J.L.; Racine, V.; Tajadura, V.; Suryavanshi, N.; Bechtel, S.; Wiemann, S.; Baum, B.; Ridley, A.J. Identification and characterization of a set of conserved and new regulators of cytoskeletal organization, cell morphology and migration. BMC Biol. 2011, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- De Ceuleneer, M.; De Wit, V.; Van Steendam, K.; Van Nieuwerburgh, F.; Tilleman, K.; Deforce, D. Modification of citrulline residues with 2,3-butanedione facilitates their detection by liquid chromatography/mass spectrometry. Rapid Commun. Mass. Spec. 2011, 25, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- De Ceuleneer, M.; Van Steendam, K.; Dhaenens, M.; Deforce, D. In vivo relevance of citrullinated proteins and the challenges in their detection. Proteomics 2012, 12, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Bicker, K.L.; Subramanian, V.; Chumanevich, A.A.; Hofseth, L.J.; Thompson, P.R. Seeing citrulline: Development of a phenylglyoxal-based probe to visualize protein citrullination. J. Am. Chem. Soc. 2012, 134, 17015–17018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ceuleneer, M.; Van steendam, K.V.; Dhaenens, M.; Elewaut, D.; Deforce, D. Quantification of citrullination by means of skewed isotope distribution pattern. J. Proteom. Res. 2012, 11, 5245–5251. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Fu, Z.; Yang, J.; Troncosco, J.; Everett, A.D.; Van Eyk, J.E. Identification and characterization of citrulline-modified brain proteins by combining HCD and CID fragmentation. Proteomics 2013, 17, 2682–2691. [Google Scholar] [CrossRef] [Green Version]
- Winget, J.M.; Finlay, D.; Mills, K.J.; Huggins, T.; Bascom, C.; Isfort, R.J.; Moritz, R. Quantitative proteomic analysis of stratum corneum dysfunction in adult chronic atopic dermatitis. J. Investig. Dermatol. 2016, 136, 1732–1735. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-Y.; Wang, D.; Wilhelm, M.; Zolg, D.P.; Schmidt, T.; Schnatbaum, K.; Reimer, U.; Pontén, F.; Uhlén, M.; Hahne, H.; et al. Mining the human tissue proteome for protein citrullination. Mol. Cell. Proteom. 2018, 17, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Fert-Bober, J.; Sokolove, J. Proteomics of citrullination in cardiovascular disease. Proteomics Clin. Appl. 2014, 8, 522–533. [Google Scholar] [CrossRef]
- Faigle, W.; Cruciani, C.; Wolski, W.; Roschitzki, B.; Puthenparampil, M.; Tomas-Ojer, P.; Sellés-Moreno, C.; Zeis, T.; Jelcic, I.; Schaeren-Wiemers, N.; et al. Brain citrullination patterns and T-cell reactivity of cerebrospinal fluid-derived CD4+ T cells in multiple sclerosis. Front. Immunol. 2019, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Mittereder, N.; Sims, G.P. Perspective on protein arginine deiminase activity-Bicarbonate is a pH-independent regulator of citrullination. Front. Immunol. 2018, 18, 34. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Abbreviation | Uniprot Identifier | Type | Mass (kDa) | Cell Type/ Compartment | References 1 |
|---|---|---|---|---|---|---|
| (Cyto)-keratin | CK or K | >50 genes | I (acidic) | 40–60 | Keratinocyte/cytosol (epidermis, hair follicle, nail) | [17,18,19] |
| II (basic) | 50–70 | |||||
| Vimentin | VIM | P08670 | III | 54 | Mesenchymal cell/cytosol | [20,21] |
| Desmin | DES | P17661 | III | 54 | Muscular cell/sarcomere | / |
| Glial fibrillary acidic protein | GFAP | P14136 | III | 50 | Glial cell, astrocyte/cytosol | [22] |
| Peripherin | PRPH | P14136 | III | 54 | Peripheral neuron/cytosol | / |
| Neurofilaments | NF-L | P07196 | IV | 62 | Neuronal axon/cytosol | / |
| NF-M | P07197 | IV | 102 | Neuronal axon/cytosol | ||
| NF-H | P12036 | IV | 112 | Neuronal axon/cytosol | ||
| Nestin | NES | P48681 | IV | 177 | Neuroepithelial cell | / |
| Lamin A/C | LMNA | P02545 | V | 74 | Nuclear lamina and nucleoplasm | [23] |
| Lamin B | LMNB1 | P20700 | V | 66 | Nuclear lamina and nucleoplasm | / |
| LMNB2 | Q03252 | V | 68 | Nuclear lamina and nucleoplasm | ||
| Filensin | BFSP1 | Q12934 | VI | 75 | Lens epithelial cell | / |
| Phakinin | BFSP2 | Q13515 | VI | 46 | Lens epithelial cell | / |
| (Pro)filaggrin | FLG | P20930 | IFAP | >250/37 | Granular keratinocyte | [17,18,19,22] |
| Trichohyalin | TCHH | Q07283 | IFAP | >250 | Hair follicle keratinocyte (IRS, medulla)/cytosol | [13,24,25,26] |
| Observed PTMs (√) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Proteins | Phosphorylation | O-Linked Glycosylation | Ubiquitination | Sumoylation | Acetylation | Transamidation | Farnesylation | ADP-Ribosylation | Proteolysis | Cross-linked | N-Acylation | GTP-Ribosylation | Deamidation | Arg. Methylation | S-Palmitoylation | Deimination 1 |
| Keratins | √ | √ | √ | √ | √ | √ | √ | |||||||||
| Lamins | √ | √ | √ | √ | √ | |||||||||||
| Vimentin | √ | √ | √ | √ | √ | √ | ||||||||||
| GFAP | √ | √ | ||||||||||||||
| Neurofilaments | √ | √ | √ | √ | ||||||||||||
| Desmin | √ | √ | √ | √ | ||||||||||||
| (Pro)Filaggrin | √ | √ | √ | √ | ||||||||||||
| TCHH | √ | √ | √ | √ | ||||||||||||
| MBP 2 | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||
| PADs | √ | √ 3 | √ 4 | |||||||||||||
| Genes | OMIM | Diseases or Human Gene Identification | Reference |
|---|---|---|---|
| PADI1 | 607934 | PADI1 cDNA cloning from a normal human keratinocyte library | [41] |
| PADI2 | 607935 | PADI2 cDNA cloning from a human cutaneous squamous cell carcinoma | [69] |
| PADI3 | 606755 | PADI3 cDNA cloning from cultured human keratinocytes | [70] |
| 191480 | Uncombable hair syndrome (UHS) | [66] | |
| 618352 | Central centrifugal cicatricial alopecia (CCCA) | [67] | |
| PADI4 | 605347 | PADI4 cDNA cloning from cultured human keratinocytes | [48] |
| 180300 | Haplotypes associated with susceptibility to rheumatoid arthritis (RA) | [71,72,73] | |
| PADI6 | 610363 | PADI6 cDNA cloning from a normal human ovary library | [40] |
| Proteomic identification in mouse eggs (ePAD) | [74] | ||
| 617234 | Preimplantation embryonic lethality-2 (PREMBL-2) | [75,76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briot, J.; Simon, M.; Méchin, M.-C. Deimination, Intermediate Filaments and Associated Proteins. Int. J. Mol. Sci. 2020, 21, 8746. https://doi.org/10.3390/ijms21228746
Briot J, Simon M, Méchin M-C. Deimination, Intermediate Filaments and Associated Proteins. International Journal of Molecular Sciences. 2020; 21(22):8746. https://doi.org/10.3390/ijms21228746
Chicago/Turabian StyleBriot, Julie, Michel Simon, and Marie-Claire Méchin. 2020. "Deimination, Intermediate Filaments and Associated Proteins" International Journal of Molecular Sciences 21, no. 22: 8746. https://doi.org/10.3390/ijms21228746